

Physiological, Transcriptome, and Metabolome Analyses Reveal the Tolerance to Cu Toxicity in Red Macroalgae Gracilariopsis lemaneiformis
 and
and
Abstract
:1. Introduction
2. Results and Discussion
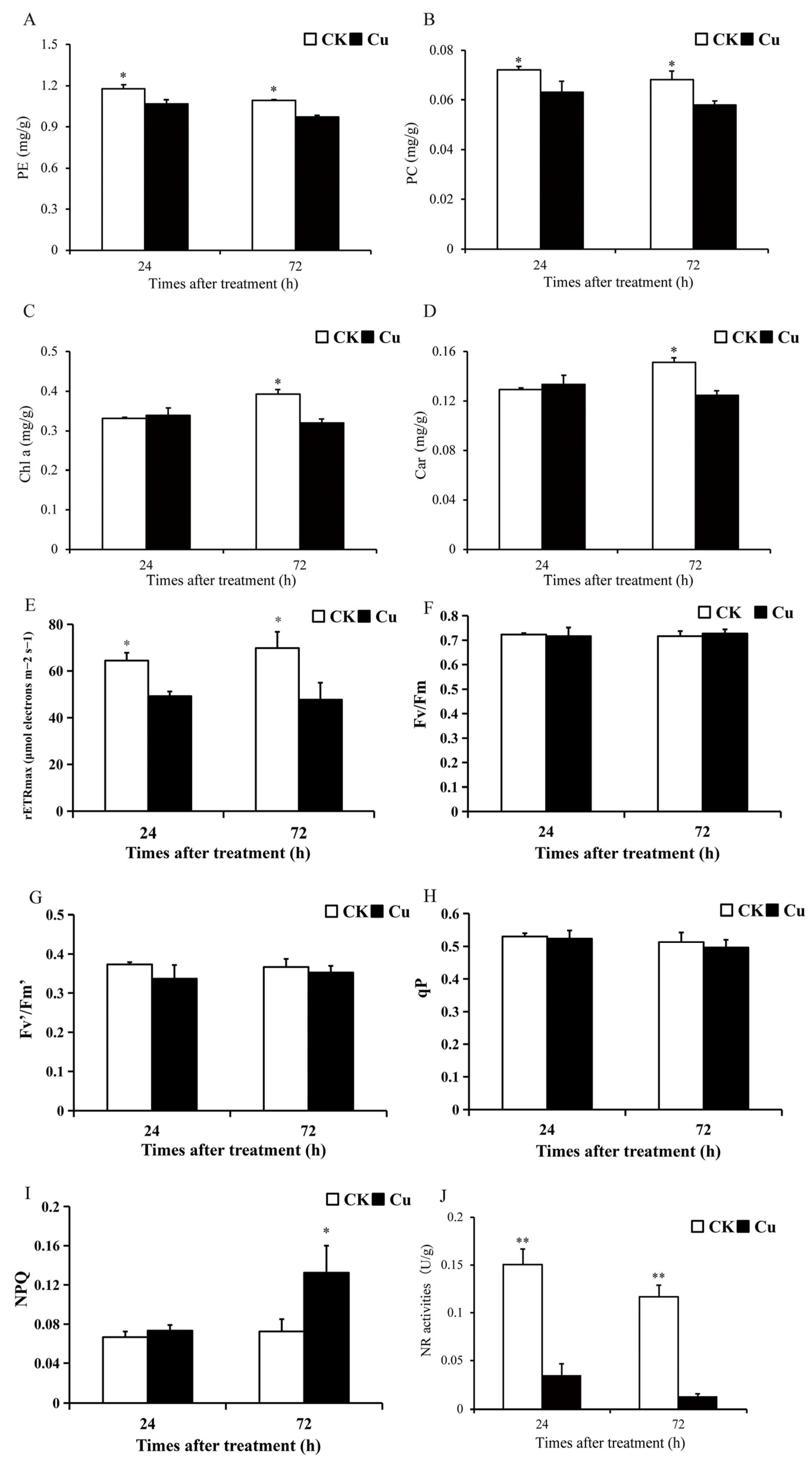
2.1. Photosynthesis and Nitrogen Metabolism
2.2. Content of H2O2 and Flavonoids
2.3. Identification and Validation of Differentially Expressed Genes
2.4. GO and KEGG Enrichment Analyses
2.5. Specific DEGs in Response to Cu
2.5.1. DEGs Involved in Cu and Other Element Transporters
2.5.2. DEGs Involved in Antioxidant System
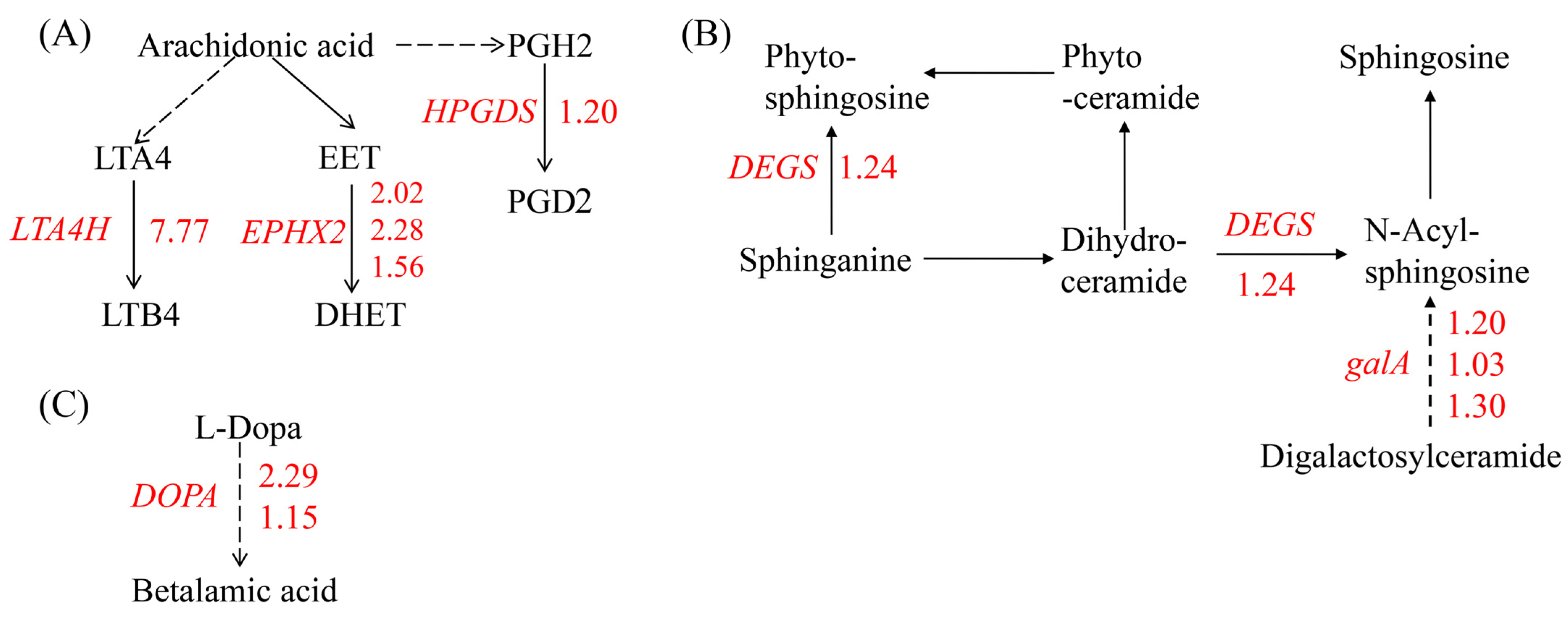
2.5.3. DEGs Involved in Lipid Metabolism and Betalain Biosynthesis
2.5.4. Cu Is Negatively Regulated in Photosynthesis, Nitrogen Metabolism, Protein Synthesis, and Amino Acid Metabolism
2.6. Metabolomics Analysis of G. lemaneiformis in Response to Cu Exposure
3. Materials and Methods
3.1. Materials, Culture Conditions, and Treatment Methods
3.2. Photosynthetic Pigments
3.3. Chlorophyll Fluorescence Parameters
3.4. Nitrate Reductase Activity
3.5. Content of H2O2 and Flavonoids
3.6. Total RNA Extraction
3.7. Transcriptome Analysis
3.8. Metabolome Analysis
3.9. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nagajyoti, P.C.; Lee, K.D.; Sreekanth, T.V.M. Heavy metals, occurrence and toxicity for plants: A review. Environ. Chem. Lett. 2010, 8, 199–216. [Google Scholar] [CrossRef]
- Zeraatkar, A.K.; Ahmadzadeh, H.; Talebi, A.F.; Moheimani, N.R.; Mchenry, M.P. Potential use of algae for heavy metal bioremediation, a critical review. J. Environ. Manag. 2016, 181, 817–831. [Google Scholar] [CrossRef] [PubMed]
- Burda, K.; Kruk, J.; Schmid, G.H.; Strzalka, K. Inhibition of oxygen evolution in Photosystem II by Cu (II) ions is associated with oxidation of cytochrome b559. Biochem. J. 2003, 371, 597–601. [Google Scholar] [CrossRef] [PubMed]
- Nazir, F.; Hussain, A.; Fariduddin, Q. Hydrogen peroxide modulate photosynthesis and antioxidant systems in tomato (Solanum lycopersicum L.) plants under copper stress. Chemosphere 2019, 230, 544–558. [Google Scholar] [CrossRef] [PubMed]
- Nowicka, B.; Kruk, J. Powered by light: Phototrophy and photosynthesis in prokaryotes and its evolution. Microbiol. Res. 2016, 186–187, 99–118. [Google Scholar] [CrossRef]
- Pinto, E.; Teresa, C.S.; Sigaud-Kutner, M.A.S.; Oswaldo, O.K.; Morse, D.; Colepicolo, P. Heavy metal-induced oxidative stress in algae. J. Phycol. 2003, 39, 1008–1018. [Google Scholar] [CrossRef]
- Gonzalez, A.; Cabrera Mde, L.; Henriquez, M.J.; Contreras, R.A.; Morales, B.; Moenne, A. Cross talk among calcium, hydrogen peroxide, and nitric oxide and activation of gene expression involving calmodulins and calcium-dependent protein kinases in Ulva compressa exposed to copper excess. Plant Physiol. 2012, 158, 1451–1462. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Li, J.; Han, H.; Du, R.; Wang, X. Physiological and Molecular Mechanisms of Plant Responses to Copper Stress. Int. J. Mol. Sci. 2022, 23, 12950. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Chen, X.; Zhang, H.; Sun, X.; Xu, N. 1-Aminocyclopropane-1-carboxylic acid mitigates copper stress by decreasing copper transport and inducing jasmonic acid synthesis in Gracilariopsis lemaneiformis. Environ. Exp. Bot. 2023, 208, 105237–105249. [Google Scholar] [CrossRef]
- Shabbir, Z.; Sardar, A.; Shabbir, A.; Abbas, G.; Shamshad, S.; Khalid, S.; Natasha; Murtaza, G.; Dumat, C.; Shahid, M. Copper uptake, essentiality, toxicity, detoxification and risk assessment in soil-plant environment. Chemosphere 2020, 259, 127436. [Google Scholar] [CrossRef]
- Nakajima, H.; Fujimoto, N.; Yamamoto, Y.; Amemiya, T.; Itoh, K. Response of secondary metabolites to Cu in the Cu-hyperaccumulator lichen Stereocaulon japonicum. Environ. Sci. Pollut. Res. Int. 2019, 26, 905–912. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; He, T.; Xu, F.; Li, X.; Yuan, L.; Wang, Q.; Liu, H. Analysis of physiological and metabolite response of Celosia argentea to copper stress. Plant Biol. 2021, 23, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Wang, Q.; Liu, Z.; Wang, S.; Long, A.; Yang, Y. Potential bioremediation effects of seaweed Gracilaria lemaneiformis on heavy metals in coastal sediment from a typical mariculture zone. Chemosphere 2020, 245, 125636. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Wang, Q.; Zhang, C.; Zhang, L.; Yang, Y. Bioaccumulation and release of heavy metals during growth and decomposition of cultivated Gracilaria lemaneiformis. Mar. Pollut. Bull. 2021, 173, 113130. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, X.; Ke, C. Bioaccumulation of trace metals by the live macroalga Gracilaria lemaneiformis. J. Appl. Phycol. 2014, 26, 1889–1897. [Google Scholar] [CrossRef]
- Ata, A.; Nalcaci, O.O.; Ovez, B. Macro algae Gracilaria verrucosa as a biosorbent: A study of sorption mechanisms. Algal Res. 2012, 1, 194–204. [Google Scholar] [CrossRef]
- Yu, H.; Yang, A.; Wang, K.; Li, Q.; Ye, D.; Huang, H.; Zhang, X.; Wang, Y.; Zheng, Z.; Li, T. The role of polysaccharides functional groups in cadmium binding in root cell wall of a cadmium-safe rice line. Ecotoxicol. Environ. Saf. 2021, 226, 112818–112825. [Google Scholar] [CrossRef] [PubMed]
- Qi, Z.; Liu, H.; Li, B.; Mao, Y.; Jiang, Z.; Zhang, J.; Fang, J. Suitability of two seaweeds, Gracilaria lemaneiformis and Sargassum pallidum, as feed for the abalone Haliotis discus hannai Ino. Aquaculture 2010, 300, 189–193. [Google Scholar] [CrossRef]
- Diwa, R.R.; Elvira, M.V.; Deocaris, C.C.; Fukuyama, M.; Belo, L.P. Transport of toxic metals in the bottom sediments and health risk assessment of Corbicula fluminea (Asiatic clam) collected from Laguna de Bay, Philippines. Sci. Total Environ. 2022, 838, 156522–156533. [Google Scholar] [CrossRef]
- Dalcorso, G. Heavy Metal Toxicity in Plants; Springer: Dordrecht, The Netherlands, 2012. [Google Scholar]
- Brown, M.T.; Newman, J.E.; Han, T. Inter-population comparisons of copper resistance and accumulation in the red seaweed, Gracilariopsis longissima. Ecotoxicology 2012, 21, 591–600. [Google Scholar] [CrossRef]
- Jiang, J.; Li, Y.; Hu, C.; Zhu, W.; Xu, N. Transcriptomic analysis reveals the regulation mechanism of sporogenesis in Ulva prolifera. Front. Mar. Sci. 2022, 9, 802006. [Google Scholar] [CrossRef]
- Zhou, R.; Yu, X.; Kjær, K.H.; Rosenqvist, E.; Ottosen, C.O.; Wu, Z. Screening and validation of tomato genotypes under heat stress using Fv/Fm to reveal the physiological mechanism of heat tolerance. Environ. Exp. Bot. 2015, 118, 1–11. [Google Scholar] [CrossRef]
- Zhao, P.; Lin, Z.; Wang, Y.; Chai, H.; Li, Y.; He, L.; Zhou, J. Facilitating effects of plant hormones on biomass production and nutrients removal by Tetraselmis cordiformis for advanced sewage treatment and its mechanism. Sci. Total Environ. 2019, 693, 133650–133658. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.Y.; Wang, S.C.; Zhang, Y.X.; Liu, F.F. Single and combined toxicity of polystyrene nanoplastics and copper on Platymonas helgolandica var. tsingtaoensis: Perspectives from growth inhibition, chlorophyll content and oxidative stress. Sci. Total Environ. 2022, 829, 154571–154580. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Kabir, A.H.; Mandal, A.; Roy, S.K.; Song, Y.; Ji, H.C.; Lee, K.W. Glutathione restores Hg-induced morpho-physiological retardations by inducing phytochelatin and oxidative defense in Alfalfa. Biology 2020, 9, 364–380. [Google Scholar] [CrossRef] [PubMed]
- Keramat, B.; Kalantari, K.M.; Arvin, M. Effects of methyl jasmonate in regulating cadmium induced oxidative stress in soybean plant (Glycine max L.). Afr. J. Microbiol. Res. 2009, 3, 240–244. [Google Scholar]
- Migocka, M.; Malas, K. Plant Responses to Copper: Molecular and Regulatory Mechanisms of Copper Uptake, Distribution and Accumulation in Plants. In Plant Micronutrient Use Efficiency; Academic Press: Cambridge, MA, USA, 2018; pp. 71–86. [Google Scholar]
- Bernal, M.; Casero, D.; Singh, V.; Wilson, G.T.; Grande, A.; Yang, H.; Dodani, S.C.; Pellegrini, M.; Huijser, P.; Connolly, E.L.; et al. Transcriptome sequencing identifies SPL7-regulated copper acquisition genes FRO4/FRO5 and the copper dependence of iron homeostasis in Arabidopsis. Plant Cell 2012, 24, 738–761. [Google Scholar] [CrossRef] [PubMed]
- Sancenón, V.; Puig, S.; Mira, H.; Thiele, D.J.; Peñarrubia, L. Identification of a copper transporter family in Arabidopsis thaliana. Plant Mol. Biol. 2003, 51, 577–587. [Google Scholar] [CrossRef]
- Rodriguez, F.I.; Esch, J.J.; Hall, A.E.; Binder, B.M.; Schaller, G.E.; Bleecker, A.B. A copper cofactor for the ethylene receptor ETR1 from Arabidopsis. Science 1999, 283, 996–998. [Google Scholar] [CrossRef]
- Cobbett, C.; Goldsbrough, P. Phytochelatins and metallothioneins: Roles in heavy metal detoxification and homeostasis. Annu. Rev. Plant Biol. 2002, 53, 159–182. [Google Scholar] [CrossRef]
- Song, W.Y.; Mendoza-Cózatl, D.G.; Lee, Y.; Schroeder, J.I.; Ahn, S.N.; Lee, H.S.; Wicker, T.; Martinoia, E. Phytochelatin-metal (loid) transport into vacuoles shows different substrate preferences in barley and Arabidopsis. Plant Cell Environ. 2014, 37, 1192–1201. [Google Scholar] [CrossRef] [PubMed]
- den Toom, W.T.F.; van Soest, D.M.K.; Polderman, P.E.; van Triest, M.H.; Bruurs, L.J.M.; De Henau, S.; Burgering, B.M.T.; Dansen, T.B. Oxygen-consumption based quantification of chemogenetic H2O2 production in live human cells. Free Radic. Biol. Med. 2023, 206, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Antonenkov, V.D.; Isomursu, A.; Mennerich, D.; Vapola, M.H.; Weiher, H.; Kietzmann, T.; Hiltunen, J.K. The human mitochondrial DNA depletion syndrome gene MPV17 encodes a non-selective channel that modulates membrane potential. J. Biol. Chem. 2015, 290, 13840–13861. [Google Scholar] [CrossRef] [PubMed]
- Pawlik-Skowronska, B.; Pirszel, J.; Brown, M.T. Concentrations of phytochelatins and glutathione found in natural assemblages of seaweeds depend on species and metal concentrations of the habitat. Aquat. Toxicol. 2007, 83, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Chen, B.; Lin, H.; Liu, Y.; Wei, Y.; Chen, F.; Li, W. Identification and characterization of the glutathione S-transferase (GST) family in radish reveals a likely role in anthocyanin biosynthesis and heavy metal stress tolerance. Gene 2020, 743, 144484–144494. [Google Scholar] [CrossRef] [PubMed]
- Gerwick, W.H.; Roberts, M.A.; Vulpanovici, A.; Ballantine, D.L. Biogenesis and biological function of marine algal oxylipins. Adv. Exp. Med. Biol. 1999, 447, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Bouarab, K.; Adas, F.; Gaquerel, E.; Kloareg, B.; Salaün, J.P.; Potin, P. The innate immunity of a marine red alga involves oxylipins from both the eicosanoid and octadecanoid pathways. Plant Physiol. 2004, 135, 1838–1848. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Zhang, Y.; Zhang, X.; Shi, Y. Long-chain base kinase1 affects freezing tolerance in Arabidopsis thaliana. Plant Sci. 2017, 259, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Markham, J.E.; Cahoon, E.B. Sphingolipid Δ8 unsaturation is important for glucosylceramide biosynthesis and low-temperature performance in Arabidopsis. Plant J. 2012, 69, 769–781. [Google Scholar] [CrossRef]
- Zhou, Y.; Zeng, L.; Fu, X.; Mei, X.; Cheng, S.; Liao, Y.; Deng, R.; Xu, X.; Jiang, Y.; Duan, X.; et al. The sphingolipid biosynthetic enzyme Sphingolipid delta8 desaturase is important for chilling resistance of tomato. Sci. Rep. 2016, 6, 38742. [Google Scholar] [CrossRef]
- Gargouri, O.D.; Gargouri, B.; Trabelsi, S.K.; Bouaziz, M.; Abdelhédi, R. Synthesis of 3-O-methylgallic acid a powerful antioxidant by electrochemical conversion of syringic acid. Biochim. Biophys. Acta 2013, 1830, 3643–3649. [Google Scholar] [CrossRef] [PubMed]
- Maoka, T.; Etoh, T.; Osawa, A.; Shindo, K. Characterization and singlet oxygen quenching activity of (3R)-3-hydroxy-4-ketotorulene and (3R)-3-hydroxy-4-keto-γ-carotene isolated from the yeast Xanthophyllomyces dendrorhrous. J. Oleo Sci. 2012, 61, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Mason, R.P.; Libby, P.; Bhatt, D.L. Emerging mechanisms of cardiovascular protection for the omega-3 fatty acid eicosapentaenoic acid. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 1135–1147. [Google Scholar] [CrossRef]
- He, M.; Ding, N.Z. Plant Unsaturated Fatty Acids: Multiple Roles in Stress Response. Front. Plant Sci. 2020, 11, 562785. [Google Scholar] [CrossRef]
- Othman, A.; Lau, B.Y.C.; Nurazah, Z.; Shahwan, S.; Rusli, M.H.; Singh, R.; Abdullah, M.H.; Marjuni, M.; Yaakub, Z.; Sundram, S.; et al. Comparative proteomic and metabolomic studies between partial resistant and susceptible oil palm reveal the molecular mechanism associated with Ganoderma boninense infection. Physiol. Mol. Plant Pathol. 2024, 129, 102198. [Google Scholar] [CrossRef]
- Kumar, M.; Kumari, P.; Gupta, V.; Reddy, C.R.K.; Jha, B. Biochemical responses of red alga Gracilaria corticata (Gracilariales, Rhodophyta) to salinity induced oxidative stress. J. Exp. Mar. Biol. Ecol. 2010, 391, 27–34. [Google Scholar] [CrossRef]
- Kumar, M.; Gupta, V.; Trivedi, N.; Kumari, P.; Bijo, A.J.; Reddy, C.R.K.; Jha, B. Desiccation induced oxidative stress and its biochemical responses in intertidal red alga Gracilaria corticata (Gracilariales, Rhodophyta). Environ. Exp. Bot. 2011, 72, 194–201. [Google Scholar] [CrossRef]
- Provasoli, L. Media and prospects for the cultivation of marine algae. In Cultures and Collections of Algae, Proceedings of the US-Japan Conference, Hakone, Japan, 12–15 September 1966; Japanese Society of Plant Physiologists: Kyoto, Japan, 1968; pp. 63–75. [Google Scholar]
- Fan, Y.; Chen, X.; Chen, Z.; Zhou, X.; Lu, X.; Liu, J. Pollution characteristics and source analysis of heavy metals in surface sediments of Luoyuan Bay, Fujian. Environ. Res. 2022, 203, 111911. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Yu, S. Anthropogenic metal loads in nearshore sediment along the coast of China mainland interacting with provincial socioeconomics in the period 1980–2020. Sci. Total Environ. 2022, 839, 156286. [Google Scholar] [CrossRef]
- Beer, S.; Eshel, A. Determining phycoerythrin and phycocyanin concentrations in aqueous crude extracts of red algae. Aust. J. Mar. Freshw. Res. 1985, 13, 133–141. [Google Scholar] [CrossRef]
- Ji, Z.; Zou, D.; Gong, J.; Liu, C.; Ye, C.; Chen, Y. The different responses of growth and photosynthesis to NH4+ enrichments between Gracilariopsis lemaneiformis and its epiphytic alga Ulva lactuca grown at elevated atmospheric CO2. Mar. Pollut. Bull. 2019, 144, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Misra, A.N.; Misra, M.; Singh, R. Chlorophyll fluorescence in plant biology. Biophysics 2012, 7, 171–192. [Google Scholar]
- Eilers, P.H.C.; Peeters, J.C.H. A model for the relationship between light intensity and the rate of photosynthesis in phytoplankton. Ecol. Model. 1988, 42, 199–215. [Google Scholar] [CrossRef]
- Van Kooten, O.; Snel, J.F.H. The use of chlorophyll fluorescence nomenclature in plant stress physiology. Photosynth. Res. 1990, 25, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Kitajima, M.; Butler, W.L. Excitation spectra for Photosystem I and Photosystem II in chloroplasts and the spectral characteristics of the distribution of quanta between the two photosystems. Biochim. Biophys. Acta Bioenerg. 1975, 408, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Ralph, P.J.; Short, F.T. Impact of the wasting disease pathogen, Labyrinthula zosterae, on the photobiology of eelgrass Zostera marina. Mar. Ecol. Prog. Ser. 2002, 226, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthesis electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Roháček, K. Chlorophyll Fluorescence Parameters: The Definitions, Photosynthetic Meaning, and Mutual Relationships. Photosynthetica 2002, 40, 13–29. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Jaworski, E.G. Nitrate reductase activity in intact plant tissues. Biochem. Biophys. Res. Commun. 1971, 43, 1274–1279. [Google Scholar] [CrossRef]
- GB 5009.226-2016. Determination of Residual Hydrogen Peroxide in Food. National Standard for Food Safety; Standards Press of China: Beijing, China, 2016.
- Woisky, R.G.; Salatino, A. Analysis of propolis: Some parameters and procedures for chemical quality control. J. Apicult. Res. 1998, 37, 99–105. [Google Scholar] [CrossRef]
- Sun, X.; Wu, J.; Wang, G.; Kang, Y.; Ooi, H.S.; Shen, T.; Wang, F.; Yang, R.; Xu, N.; Zhao, X. Genomic analyses of unique carbohydrate and phytohormone metabolism in the macroalga Gracilariopsis lemaneiformis (Rhodophyta). BMC Plant Biol. 2018, 18, 94. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Tang, Y.Y.; Yin, H.; Sun, X.; Zhang, X.; Xu, N. A survey of the full-length transcriptome of Gracilariopsis lemaneiformis using single-molecule long-read sequencing. BMC Plant Biol. 2022, 22, 597. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.L.; Zhu, Q.H.; Xie, Z.Q.; Xu, B.; Wang, X.Q.; Chen, J.Y. Genome-wide gene expression analysis of amphioxus (Branchiostoma belcheri) following lipopolysaccharide challenge using strand-specific RNA-seq. RNA Biol. 2017, 14, 1799–1809. [Google Scholar] [CrossRef]
- Chen, X.; Tang, Y.; Sun, X.; Zhang, X.; Xu, N. Comparative transcriptome analysis reveals the promoting effects of IAA on biomass production and branching of Gracilariopsis lemaneiformis. Aquaculture 2022, 548, 737678. [Google Scholar] [CrossRef]
- Meng, J.J.; Shen, J.W.; Li, G.; Ouyang, C.J.; Hu, J.X.; Li, Z.S.; Zhao, H.; Shi, Y.M.; Zhang, M.; Liu, R.; et al. Light modulates glucose metabolism by a retina-hypothalamus-brown adipose tissue axis. Cell 2023, 186, 398–412.e17. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Level 1 | Level 2 | Pathway ID | Pathway Name | p Value |
|---|---|---|---|---|
| Up-regulated DEGs | ||||
| Metabolism | Lipid metabolism | ko00590 | Arachidonic acid metabolism | 0.0041 |
| ko00600 | Sphingolipid metabolism | 0.019 | ||
| Metabolism of cofactors and vitamins | ko00730 | Thiamine metabolism | 0.013 | |
| Biosynthesis of other secondary metabolites | ko00965 | Betalain biosynthesis | 0.015 | |
| Carbohydrate metabolism | ko00052 | Galactose metabolism | 0.036 | |
| Environmental Information Processing | Membrane transport | ko02010 | ABC transporters | 0.036 |
| Cellular Processes | Transport and catabolism | ko04146 | Peroxisome | 0.0057 |
| Genetic Information Processing | Replication and repair | ko03030 | DNA replication | 0.024 |
| Transcription | ko03020 | RNA polymerase | 0.048 | |
| ko03022 | Basal transcription factors | 0.010 | ||
| Down-regulated DEGs | ||||
| Genetic Information Processing | Translation | ko03010 | Ribosome | 6.2 × 10−18 |
| Metabolism | Energy metabolism | ko00196 | Photosynthesis—antenna proteins | 8.2 × 10−10 |
| ko00910 | Nitrogen metabolism | 7.0 × 10−5 | ||
| Amino acid metabolism | ko00360 | Phenylalanine metabolism | 0.013 | |
| ko00350 | Tyrosine metabolism | 0.039 | ||
| ko00340 | Histidine metabolism | 0.0047 | ||
| Carbohydrate metabolism | ko00010 | Glycolysis/gluconeogenesis | 0.025 | |
| Metabolism of terpenoids and polyketides | ko00903 | Limonene and pinene degradation | 0.032 | |
| Biosynthesis of other secondary metabolites | ko00960 | Tropane, piperidine, and pyridine alkaloid biosynthesis | 0.035 | |
| GENE | Gene ID | log2 (Fold Change) |
|---|---|---|
| Zinc transporters 6 (ZIP6) | PB.6159.1 | 1.00 |
| Zinc transporters 3 (ZIP3) | PB.1088.4 | 1.04 |
| Zinc transporter ZupT | PB.4500.2 | 1.27 |
| Zinc transporters 1 (ZIP1) | PB.4116.1 | 1.60 |
| Iron transporter (IRT) | PB.552.1 | 2.87 |
| Copper-transporting ATPase HMA5 | PB.4965.1 | 1.54 |
| ATP-binding cassette, subfamily C (CFTR/MRP), member 1 (ABCC1) | PB.4380.6 | 1.08 |
| ATP-binding cassette, subfamily D (ALD), member 3 (ABCD3) | Gene.10182 | 1.45 |
| PB.425.20 | 1.40 | |
| ATP-binding cassette, subfamily N (ALN), member 7 (ABCN7) | Gene.670 | 1.15 |
| Gene.674 | 1.83 | |
| Gene.678 | 1.09 | |
| ABCD (PXA1) | Gene.10182 | 1.45 |
| PB.425.20 | 1.40 | |
| Phosphate transporter | PB.117.55 | −1.03 |
| Inorganic phosphate cotransporter | PB.2081.2 | −1.28 |
| High-affinity nitrate transporter 2.5 (NR 2.5) | PB.207.1 | −3.92 |
| Magnesium transporter | PB.4084.1 | −1.52 |
| GENE | Gene ID | log2 (Fold Change) |
|---|---|---|
| Catalase (CAT) | PB.5635.1 | −1.06 |
| Mitochondrial inner membrane protein (MPV17) | PB.5254.4 | −1.07 |
| PB.5254.6 | −1.10 | |
| Ascorbate peroxidase (APX) | PB.6120.1 | 1.36 |
| Glutathione-S-transferase (GST) | PB.6753.1 | 1.20 |
| Glutathione reductase (GR) | PB.5.219 | 0.98 |
| Superoxide dismutase (SOD) | PB.4633.17 | 1.67 |
| PB.4633.18 | 1.11 | |
| D-aspartate oxidase (DDO) | PB.1107.1 | 1.20 |
| Epoxide hydrolase 2 (EPHX2) | Gene.42815 | 2.02 |
| PB.2662.1 | 2.28 | |
| PB.7028.1 | 1.56 | |
| Peroxisomal protein 13 (PEX13) | PB.4437.3 | 1.87 |
| Peroxisome membrane protein 70 (PMP70) | Gene.10182 | 1.45 |
| PB.425.20 | 1.40 | |
| Peroxiredoxins (PRXS) | PB.4818.4 | 1.16 |
| Pathway_Hierarchy1 | Pathway_Hierarchy2 | Map_Name | Test |
|---|---|---|---|
| Metabolism | Nucleotide metabolism | Purine metabolism | 4 |
| Metabolism | Carbohydrate metabolism | Galactose metabolism | 3 |
| Metabolism | Carbohydrate metabolism | Ascorbate and aldarate metabolism | 3 |
| Organismal Systems | Digestive system | Protein digestion and absorption | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.; Tang, Y.; Zhang, H.; Zhang, X.; Sun, X.; Zang, X.; Xu, N. Physiological, Transcriptome, and Metabolome Analyses Reveal the Tolerance to Cu Toxicity in Red Macroalgae Gracilariopsis lemaneiformis. Int. J. Mol. Sci. 2024, 25, 4770. https://doi.org/10.3390/ijms25094770
Chen X, Tang Y, Zhang H, Zhang X, Sun X, Zang X, Xu N. Physiological, Transcriptome, and Metabolome Analyses Reveal the Tolerance to Cu Toxicity in Red Macroalgae Gracilariopsis lemaneiformis. International Journal of Molecular Sciences. 2024; 25(9):4770. https://doi.org/10.3390/ijms25094770
Chicago/Turabian StyleChen, Xiaojiao, Yueyao Tang, Hao Zhang, Xiaoqian Zhang, Xue Sun, Xiaonan Zang, and Nianjun Xu. 2024. "Physiological, Transcriptome, and Metabolome Analyses Reveal the Tolerance to Cu Toxicity in Red Macroalgae Gracilariopsis lemaneiformis" International Journal of Molecular Sciences 25, no. 9: 4770. https://doi.org/10.3390/ijms25094770