Can a Single Amphibian Species Be a Good Biodiversity Indicator?

Abstract
:
1. Introduction
2. Methods
2.1. Analysis of Indicator Criteria
2.2. Study Area
2.3. Data Collection
2.4. Data Analysis
3. Results and Discussion
3.1. Indicator Criteria

{kind=link}
{kind=link}
| Criteria | Applicable to type (i) or type (ii) indicator, or both | Number of occurrences |
|---|---|---|
| 1. The species should be easy and cost effective to observe, identify and monitor | Both | 6 |
| 2. Patterns of species richness should be closely correlated with those of other, non-related groups | Type (i) | 3 |
| 3. The species should be sufficiently sensitive to provide early warning of change in the environment | Type (ii) | 4 |
| 4. Species should be capable of providing a continuous assessment over a wide range of stresses, including anthropogenic stresses | Type (ii) | 4 |
| 5. It should be possible to differentiate between the species responses to natural and anthropogenic stresses | Type (ii) | 4 |
| 6. The species should be distributed over a broad geographical area, if the indicator is at family or higher level, ideally worldwide | Both | 4 |
| 7. It may be better to use a number of species rather than have just one | Both | 7 |
| 8. The indicator should be relevant to ecologically significant phenomena | Both | 2 |
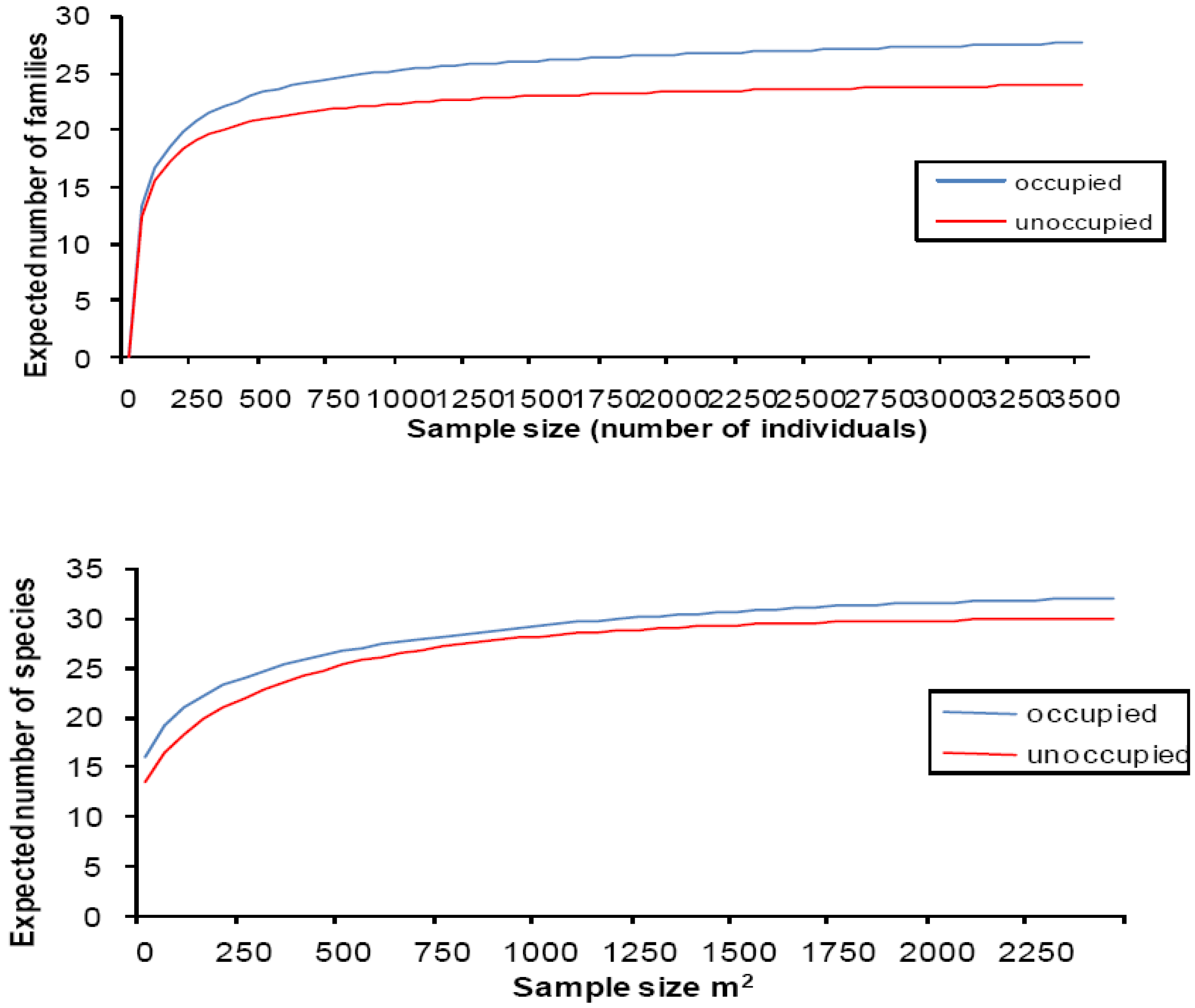
3.2. Relationship between Newts and Plant and Invertebrate Diversity
| Occupied ponds mean ± SD | Unoccupied ponds mean ± SD | ANCOVA with pond area as a covariate | |
|---|---|---|---|
| Aquatic plants | |||
| Number of species per pond | 6.9 ± 3.85 | 3.8 ± 3.11 | F 1, 62 = 19.22 *** |
| Percentage cover per pond | 45.2 ± 24.00 | 25.6 ± 28.97 | F 1, 62 = 0.05 NS |
| Shannon–Wiener function | 1.1 ± 0.67 | 0.67 ± 0.65 | F 1, 62 = 19.30 *** |
| Simpson's index | 0.54 ± 0.35 | 0.36 ± 0.36 | F 1, 62 = 12.82 *** |
| Macroinvertebrates | |||
| Number of families per pond | 12.6 ± 3.36 | 11.6 ± 3.86 | F 1,24 = 0.04 NS |
| Individuals per pond | 276.8 ± 125.30 | 287.7 ± 265.44 | F 1,24 = 0.002 NS |
| Shannon–Wiener function | 3.36 ± 0.88 | 3.05 ± 0.93 | F 1,24 = 0.04 NS |
| Simpson's index | 0.71 ± 0.12 | 0.67 ± 0.15 | F 1,24 = 0.026 NS |

4. Conclusions
Acknowledgements
References
- Caro, T.M.; O’Doherty, G. On the use of surrogate species in conservation biology. Conserv. Biol. 1999, 13, 805–814. [Google Scholar] [CrossRef]
- Simberloff, D. Flagships, umbrellas and keystones: is single species management passé in the landscape era? Biol. Conserv. 1998, 83, 247–257. [Google Scholar] [CrossRef]
- Leader-Williams, N.; Dublin, H.T. Charismatic megafauna as ‘flagship species’. In Priorities for the Conservation of Mammalian Diversity: Has the Panda had its Day? Entwistle, A., Dunstone, N., Eds.; Cambridge University Press: Cambridge, UK, 2000; pp. 53–81. [Google Scholar]
- Akutsu, K.; Khen, C.V.; Toda, M.J. Assessment of higher insect taxa as bioindicators for different logging-disturbance regimes in lowland tropical rain forest in Sabah, Malaysia. Ecol. Res. 2007, 22, 542–550. [Google Scholar] [CrossRef]
- Espinosa, F.; Guerra-Garcia, J.M.; Garcia-Gomez, J.C. Sewage pollution and extinction risk: an endangered limpet as a bioindicator? Biodivers. Conserv. 2007, 16, 377–397. [Google Scholar] [CrossRef]
- Limaverde, A.M.; Wagener, A.D.L.R.; Fernandez, M.A.; Schofield, A.D.L.; Coutinho, R. Stramonita haemastoma as a bioindicator for organotin contamination in coastal environments. Mar. Environ. Res. 2007, 64, 384–398. [Google Scholar] [CrossRef]
- Moganti, S.; Richardson, B.J.; McClellan, K.; Martin, M.; Lam, P.K.S.; Zheng, G.J. Use of the clam Asaphis deflorata as a potential indicator of organochlorine bioaccumulation in Hong Kong coastal sediments. Mar. Pollut. Bull. 2008, 57, 672–680. [Google Scholar] [CrossRef]
- Ugolini, A.; Ungherese, G.; Somigli, S.; Galanti, G.; Baroni, D.; Borghini, F.; Cipriani, N.; Nebbiai, M.; Passaponti, M.; Focardi, S. The amphipod Talitrus saltator as a bioindicator of human trampling on sandy beaches. Mar. Environ. Res. 2008, 65, 349–357. [Google Scholar] [CrossRef]
- Noss, R.F. Indicators for monitoring biodiversity: a hierarchical approach. Conserv. Biol. 1990, 4, 355–364. [Google Scholar] [CrossRef]
- Pearson, D.L.; Cassola, F. World-wide species richness patterns of tiger beetles (Coleoptera: Cicindelidae): indicator taxon for biodiversity and conservation studies. Conserv. Biol. 1992, 6, 376–391. [Google Scholar]
- Kremen, C.; Colwell, R.K.; Erwin, T.L.; Murphy, D.D.; Noss, R.F.; Sanjayan, M.A. Terrestrial arthropod assemblages: their use in conservation planning. Conserv. Biol. 1993, 7, 794–808. [Google Scholar]
- Brooks, R.P.; O’Connell, T.J.; Wardrop, D.H.; Jackson, L.E. Towards a regional index of biological integrity: the example of forested riparian ecosystems. Environ. Monit. Assess. 1998, 51, 131–143. [Google Scholar] [CrossRef]
- Howard, P.C.; Viscanic, P.; Davenport, T.R.B.; Kigenyi, F.W.; Baltzer, M.; Dickinson, C.J.; Lwanga, J.S.; Matthews, R.A.; Balmford, A. Complementarity and the use of indicator groups for reserve selection in Uganda. Nature 1998, 394, 472–475. [Google Scholar] [CrossRef]
- Dale, V.H.; Beyeler, S.C. Changes in the development and use of ecological indicators. Ecol. Indic. 2001, 1, 3–10. [Google Scholar] [CrossRef]
- Fleishman, E.; Murphy, D.D. A realistic assessment of the Indicator potential of butterflies and other charismatic taxonomic groups. Conserv. Biol. 2009, 23, 1109–1116. [Google Scholar] [CrossRef]
- Gregory, R.D.; van Strien, A.; Vorisek, P.; Gmelig Meyling, A.W.; Noble, D.G.; Foppen, R.P.B.; Gibbons, D.W. Developing indicators for European birds. Phil. Trans. R. Soc. B. 2005, 360, 269–288. [Google Scholar] [CrossRef]
- Gregory, R.D.; Vořišek, P.; Noble, D.G.; van Strien, A.; Klvaňová, A.; Eaton, M.; Gmelig Meyling, A.W.; Joys, A.; Foppen, R.P.B.; Burfield, I.J. The generation and use of bird population indicators in Europe. Bird Conserv. Int. 2008, 18, S223–S244. [Google Scholar]
- Grenyer, R.; Orme, C.D.L.; Jackson, S.F.; Thomas, G.H.; Davies, R.G.; Davies, T.J.; Jones, K.E.; Olson, V.A.; Ridgely, R.S.; Rasmussen, P.C.; Ding, T.S.; Bennett, P.M.; Blackburn, T.M.; Gaston, K.J.; Gittleman, J.L.; Owens, I.P.F. Global distribution and conservation of rare and threatened vertebrates. Nature 2006, 444, 93–96. [Google Scholar] [CrossRef]
- Sebastiăo, H.; Grelle, C.E.V. Taxon surrogates among Amazonian mammals: Can total species richness be predicted by single orders? Ecol. Indic. 2009, 9, 160–166. [Google Scholar] [CrossRef]
- Loyola, R.D.; Kubota, U.; Lewinsohn, T.M. Endemic vertebrates are the most effective surrogates for identifying conservation priorities among Brazilian ecoregions. Diversity Distrib. 2007, 13, 389–396. [Google Scholar] [CrossRef]
- Franco, A.M.A.; Anderson, B.J.; Roy, D.B.; Gillings, S.; Fox, R.; Moilanen, A.; Thomas, C.D. Surrogacy and persistence in reserve selection: landscape prioritization for multiple taxa in Britain. J. Appl. Ecol. 2009, 46, 82–91. [Google Scholar] [CrossRef]
- Smith, G.F.; Gittings, T.; Wilson, M.; French, L.; Oxbrough, A.; O’Donoghue, S.; O’Halloran, J.; Kelly, D.L.; Mitchell, F.J.G.; Kelly, T.; Iremonger, S.; McKee, A.-M.; Giller, P. Identifying practical indicators of biodiversity for stand-level management of plantation forests. Biodivers. Conserv. 2008, 17, 991–1015. [Google Scholar] [CrossRef]
- Blaustein, A.R.; Wake, D.B. Declining amphibian populations: a global phenomenon. Trends Ecol. Evol. 1990, 5, 203–204. [Google Scholar] [CrossRef]
- Wyman, R.L. What’s happening to the amphibians? Conserv. Biol. 1990, 4, 350–352. [Google Scholar] [CrossRef]
- Lips, K.R. Decline of a tropical montane amphibian fauna. Conserv. Biol. 1998, 12, 106–112. [Google Scholar] [CrossRef]
- Vitt, L.J.; Caldwell, J.P.; Wilbur, H.M.; Smith, D.C. Amphibians as harbingers of decay. Bioscience 1990, 40, 417. [Google Scholar]
- Wake, D.B. Declining amphibian populations. Science 1991, 253, 860. [Google Scholar] [CrossRef]
- Collins, J.P.; Storfer, A. Global amphibian declines: sorting the hypotheses. Divers. Distrib. 2003, 9, 89–98. [Google Scholar] [CrossRef]
- Storfer, A. Amphibian declines: future directions. Divers. Distrib. 2003, 9, 151–163. [Google Scholar] [CrossRef]
- Blaustein, A.R.; Romansic, J.M.; Kiesecker, J.M.; Hatch, A.C. Ultraviolet radiation, toxic chemicals and amphibian population declines. Divers. Distrib. 2003, 9, 123–140. [Google Scholar] [CrossRef]
- Gascon, C.; Collins, J.P.; Moore, R.D.; Church, D.R.; McKay, J.E.; Mendelson, J.R., III (Eds.) Amphibian Conservation Action Plan; IUCN/SSC Amphibian Specialist Group: Gland, Switzerland, 2008; pp. 17–19.
- Pechmann, J.H.K.; Wilbur, H.M. Putting declining amphibian populations in perspective: natural fluctuations and human impacts. Herpetologica 1994, 50, 65–84. [Google Scholar]
- Halliday, T. Do frogs make good canaries? Biologist 2000, 47, 143–146. [Google Scholar]
- Relyea, R.A.; Schoeppner, N.M.; Hoverman, J.T. Pesticides and amphibians: The importance of community context. Ecol. Appl. 2005, 15, 1125–1134. [Google Scholar] [CrossRef]
- Relyea, R.A. A cocktail of contaminants: how mixtures of pesticides at low concentrations affect aquatic communities. Oecologia 2009, 159, 363–376. [Google Scholar] [CrossRef]
- Kerby, J.L.; Richards-Hrdlicka, K.L.; Storfer, A.; Skelly, D.K. An examination of amphibian sensitivity to environmental contaminants: are amphibians poor canaries? Ecol. Lett. 2009, 12, 1–8. [Google Scholar]
- Beebee, T.J.C.; Griffiths, R.A. The amphibian decline crisis: A watershed for conservation biology? Biol. Conserv. 2005, 125, 271–285. [Google Scholar] [CrossRef]
- DeGarady, C.J.; Halbrook, R.S. Using anurans as bioindicators of PCB contaminated streams. J. Herpetol. 2006, 40, 127–130. [Google Scholar] [CrossRef]
- Lebboroni, M.; Ricchiardino, G.; Bellavita, M.; Chelazzi, G. Potential use of anurans as indicators of biological quality in upstreams of central Italy. Amphibia-Reptilia 2006, 27, 73–79. [Google Scholar] [CrossRef]
- Capaldo, A.; Gay, F.; DeFalco, M.; Virgilio, F.; Valiante, S.; Laforgia, V.; Varano, L. The newt Triturus carnifex as a model for monitoring the ecotoxic impact of the fungicide thiophanate methyl: adverse effects on the adrenal gland. Comp. Biochem. Phys. C 2006, 143, 86–93. [Google Scholar]
- Maletzky, A.; Kyek, M.; Goldschmid, A. Monitoring status, habitat features and amphibian species richness of Crested newt (Triturus cristatus superspecies) ponds at the edge of the species range (Salzburg, Austria). Ann. Limnol. Int. J. Lim. 2007, 43, 107–115. [Google Scholar] [CrossRef]
- Gustafson, D.H.; Pettersson, C.J.; Malmgren, J.C. Great crested newts (Triturus cristatus) as indicators of aquatic plant diversity. Herpetol. J. 2007, 16, 347–352. [Google Scholar]
- Corbett, K. European perspective and status. In Conservation and Management of Great Crested Newts, Proceedings of a Symposium, Kew Gardens, Richmond, Surrey, 11 January 1994; Gent, A., Bray, R., Eds.; English Nature: Peterborough, UK, 1994; pp. 7–17. [Google Scholar]
- Wildcare. Farm Wildlife Habitat Scheme. Available online: http://www.wildcare.co.uk/species.asp?navid=3 (accessed 22 June 2009).
- Jehle, R. The terrestrial summer habitat of radio-tracked great crested newts (Triturus cristatus) and marbled newts (T. mamoratus). Herpetol. J. 2000, 10, 137–142. [Google Scholar]
- Waddle, J.H. Use of Amphibians as Ecosystem Indicator Species. PhD thesis, University of Florida, Gainesville, FL, USA, 2006. [Google Scholar]
- Williams, C. Metapopulation Dynamics of the Crested Newt (Triturus cristatus). PhD thesis, University of Kent, Canterbury, UK, 1999. [Google Scholar]
- Griffiths, R.A. A simple funnel trap for studying newt populations and an evaluation of trap behaviour in smooth and palmate newts, Triturus vulgaris and T. helveticus. Herpetol. J. 1985, 1, 5–10. [Google Scholar]
- Griffiths, R.A.; Raper, S.J.; Brady, L.D. Evaluation of a Standard Method for Surveying Common Frogs Rana temporaria and Newts Triturus cristatus, T. helveticus and T. vulgaris; JNCC Report no. 259; Joint Nature Conservation Committee: Peterborough, UK, 1996. [Google Scholar]
- Biggs, J.; Fox, G.; Nicolet, P.; Walker, D.; Whitfield, M.; Williams, P. A Guide to the Methods of the National Pond Survey; Pond Action: Oxford, UK, 1998. [Google Scholar]
- Bursche, E.M. A Handbook of Water Plants; Frederick Warne & Co.: London, UK, 1971; pp. 1–126. [Google Scholar]
- Fitter, R.; Fitter, A.; Blamey, M. The Wild Flowers of Britain and Northern Europe, 3rd ed.; Collins: London, UK, 1978. [Google Scholar]
- Preston, C.D.; Croft, J.M. Aquatic Plants in Britain and Ireland; Harley Books: Colchester, UK, 1997. [Google Scholar]
- Philp, E.G. Atlas of the Kent Flora; Kent Field Club: West Malling, UK, 1982. [Google Scholar]
- Croft, P.S. A Key to the Major Groups of British Freshwater Invertebrates; Field Studies Council: Preston Montford, UK, 1986. [Google Scholar]
- Olsen, L.-H.; Sunesen, J.; Pedersen, B.V. Small Freshwater Creatures; Oxford University Press: Oxford, UK, 2001. [Google Scholar]
- Henderson, P.A. Practical Methods in Ecology; Blackwell Science: Malden, MA, USA, 2003; pp. 126–128. [Google Scholar]
- Beebee, T.J.C.; Griffiths, R.A. Amphibians and Reptiles; Harper Collins: London, UK, 2000; pp. 63–72. [Google Scholar]
- Oldham, R.S.; Keeble, J.; Swan, M.J.S.; Jeffcote, M. Evaluating the suitability of habitat for the great crested newt (Triturus cristatus). Herpetol. J. 2000, 10, 143–155. [Google Scholar]
- Oertli, B.; Joye, D.A.; Castella, E.; Juge, R.; Cambin, D.; Lachavanne, J.-B. Does size matter? The relationship between pond area and biodiversity. Biol. Conserv. 2002, 104, 59–70. [Google Scholar] [CrossRef]
- MacKenzie, D.I.; Nichols, J.D.; Lachman, G.B.; Droege, S.; Royle, J.A.; Langtim, C.A. Estimating site occupancy rates when detection probabilities are less than one. Ecology 2002, 83, 2248–2255. [Google Scholar] [CrossRef]
- Sanders, H.L. Marine benthic diversity: a comparative study. Am. Nat. 1968, 102, 243–282. [Google Scholar]
- Hurlbert, S.H. The non-concept of species diversity: a critique and alternative parameters. Ecology 1971, 52, 577–586. [Google Scholar] [CrossRef]
- Simberloff, D.S. Properties of the rarefaction diversity measurement. Am. Nat. 1972, 106, 414–418. [Google Scholar]
- Sewell, D.L. Great Crested Newts (Triturus cristatus) as Indicators of Pond Biodiversity. PhD thesis, University of Kent, Canterbury, UK, 2006. [Google Scholar]
- MacKenzie, D.I.; Nichols, J.D.; Hines, J.E.; Knutson, M.G.; Franklin, A.B. Estimating site occupancy, colonization, and local extinction when a species is detected imperfectly. Ecology 2003, 84, 2200–2207. [Google Scholar] [CrossRef]
- Schmidt, B.R. Count data, detection probabilities, and the demography, dynamics, distribution and declines of amphibians. C. R. Biol. 2003, 326, S119–S124. [Google Scholar] [CrossRef]
- Schmidt, B.R. Declining amphibian populations: the pitfalls of count data in the study of diversity, distributions, dynamics and demography. Herpetol. J. 2004, 14, 167–174. [Google Scholar]
- Schmidt, B.R. Monitoring the distribution of pond-breeding amphibians when species are detected imperfectly. Aquat. Conserv. 2005, 15, 681–692. [Google Scholar] [CrossRef]
- Kati, V.; DeVilliers, P.; Dufrêne, M.; Legakis, A.; Vokou, D.; Lebrun, P. Testing the value of six taxonomic groups as biodiversity indicators at a local scale. Conserv. Biol. 2004, 18, 667–675. [Google Scholar] [CrossRef]
- Francillon-Vieillot, H.; Arntzen, J.W.; Geraudie, J. Age, growth and longevity of sympatric Triturus cristatus, T. mamoratus and their hybrids (Amphibia: Urodela): a skeletochronological comparison. J. Herpetol. 1990, 24, 13–22. [Google Scholar] [CrossRef]
- Griffiths, R.A. Newts and Salamanders of Europe; Academic Press: San Diego, CA, USA, 1996; pp. 143–144. [Google Scholar]
- Marsh, D.M. Fluctuations in amphibian populations: a meta-analysis. Biol. Conserv. 2001, 101, 327–335. [Google Scholar] [CrossRef]
- Green, D.M. The ecology of extinction: population fluctuation and decline in amphibians. Biol. Conserv. 2003, 111, 331–343. [Google Scholar] [CrossRef]
- Pechmann, J.H.K.; Scott, D.E.; Semlitsch, R.D.; Caldwell, J.P.; Vitt, L.J.; Gibbons, J.W. Declining amphibian populations—the problem of separating human impacts from natural fluctuations. Science 1991, 253, 892–895. [Google Scholar] [CrossRef]
- Lambeck, R.J. Focal species: a multi-species umbrella for nature conservation. Conserv. Biol. 1997, 11, 849–856. [Google Scholar]
- Lindenmayer, D.B.; Margules, C.R.; Botkin, D.B. Indicators of biodiversity for ecologically sustainable forest management. Conserv. Biol. 2000, 14, 941–950. [Google Scholar] [CrossRef]
- Biggs, J.; Williams, P.J.; Corfield, A.; Whitfield, M.A.; Barr, C.J.; Cummins, C.P. Pond Survey 1996 Stage 1 Scoping Survey; Pond Action: Oxford, UK, 1996. [Google Scholar]
- Environment Agency. The National Pond Monitoring Network, a Strategy for Pond Monitoring, Surveillance and Inventory Development; Ponds Conservation Trust: Oxford, UK, 2003. [Google Scholar]
- Collinson, N.H.; Biggs, J.; Corfield, A.; Hodson, M.J.; Walker, D.; Whitfield, M.; Williams, P.J. Temporary and permanent ponds: An assessment of the effects of drying out on the conservation value of aquatic macroinvertebrate communities. Biol. Conserv. 1995, 74, 125–133. [Google Scholar] [CrossRef]
- Bazzanti, M.; Della Bella, V.; Seminara, M. Factors affecting macroinvertebrate communities in astatic ponds in central Italy. J. Freshwater Ecol. 2003, 18, 537–548. [Google Scholar] [CrossRef]
- Larsen, F.W.; Bladt, J.; Rahbek, C. Indicator taxa revisited: useful for conservation planning? Diversity Distrib. 2009, 15, 70–79. [Google Scholar] [CrossRef]
- Everard, M. Selection of taxa as indicators of river and freshwater wetland quality in the U.K. Aquat. Conserv. 2008, 18, 1052–1061. [Google Scholar] [CrossRef]
- DETR. Indicators of Sustainable Development: Quality of Life Counts; Department of Environment, Transport and the Regions: London, UK, 1999.
- Department of the Environment, Food and Rural Affairs (Defra). Working with the Grain of Nature: a Biodiversity Strategy for England; Defra Publications: London, UK, 2002.
- Department of the Environment, Food and Rural Affairs (Defra). The UK Biodiversity Action Plan: Highlights from the 2005 Reporting Round; Defra Publications: London, UK, 2006. [Google Scholar]
- UK Biodiversity Partnership. A Biodiversity Strategy for England–Part Two: Indicators. Draft report. 2003. Available online: http://www.ukbap.org.uk/ebg/library/ebg/ebg-03-01.pdf (accessed 29 July, 2009).
- Gibbons, D.W. Societal values and attitudes: Their history and sociological influences on amphibian conservation problems. In Amphibian Conservation; Semlitsch, R.D., Ed.; Smithsonian Institution Press: Washington, DC, USA, 2003; pp. 214–222. [Google Scholar]
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sewell, D.; Griffiths, R.A. Can a Single Amphibian Species Be a Good Biodiversity Indicator? Diversity 2009, 1, 102-117. https://doi.org/10.3390/d1020102
Sewell D, Griffiths RA. Can a Single Amphibian Species Be a Good Biodiversity Indicator? Diversity. 2009; 1(2):102-117. https://doi.org/10.3390/d1020102
Chicago/Turabian StyleSewell, David, and Richard A. Griffiths. 2009. "Can a Single Amphibian Species Be a Good Biodiversity Indicator?" Diversity 1, no. 2: 102-117. https://doi.org/10.3390/d1020102
APA StyleSewell, D., & Griffiths, R. A. (2009). Can a Single Amphibian Species Be a Good Biodiversity Indicator? Diversity, 1(2), 102-117. https://doi.org/10.3390/d1020102