Kelps’ Long-Distance Dispersal: Role of Ecological/Oceanographic Processes and Implications to Marine Forest Conservation

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
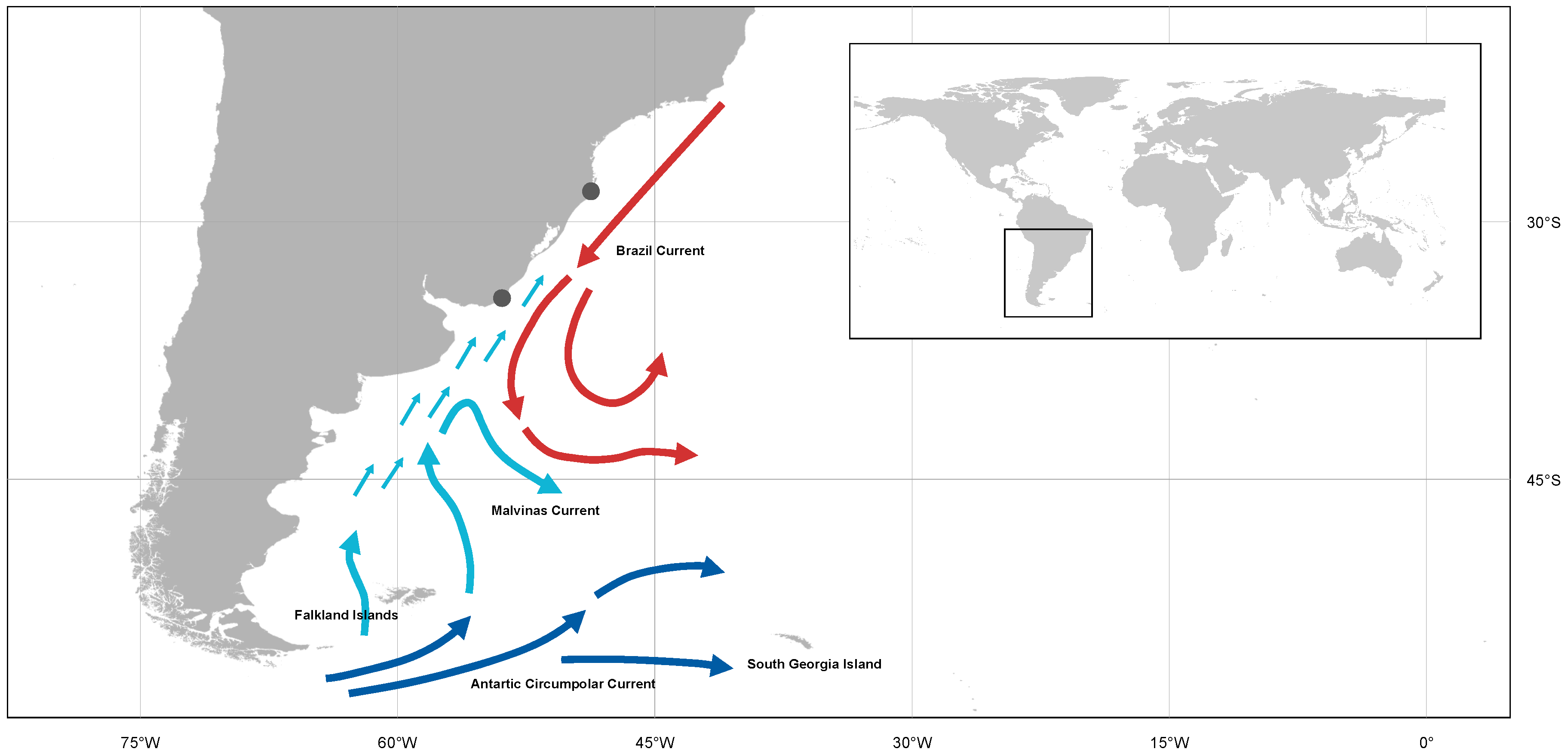
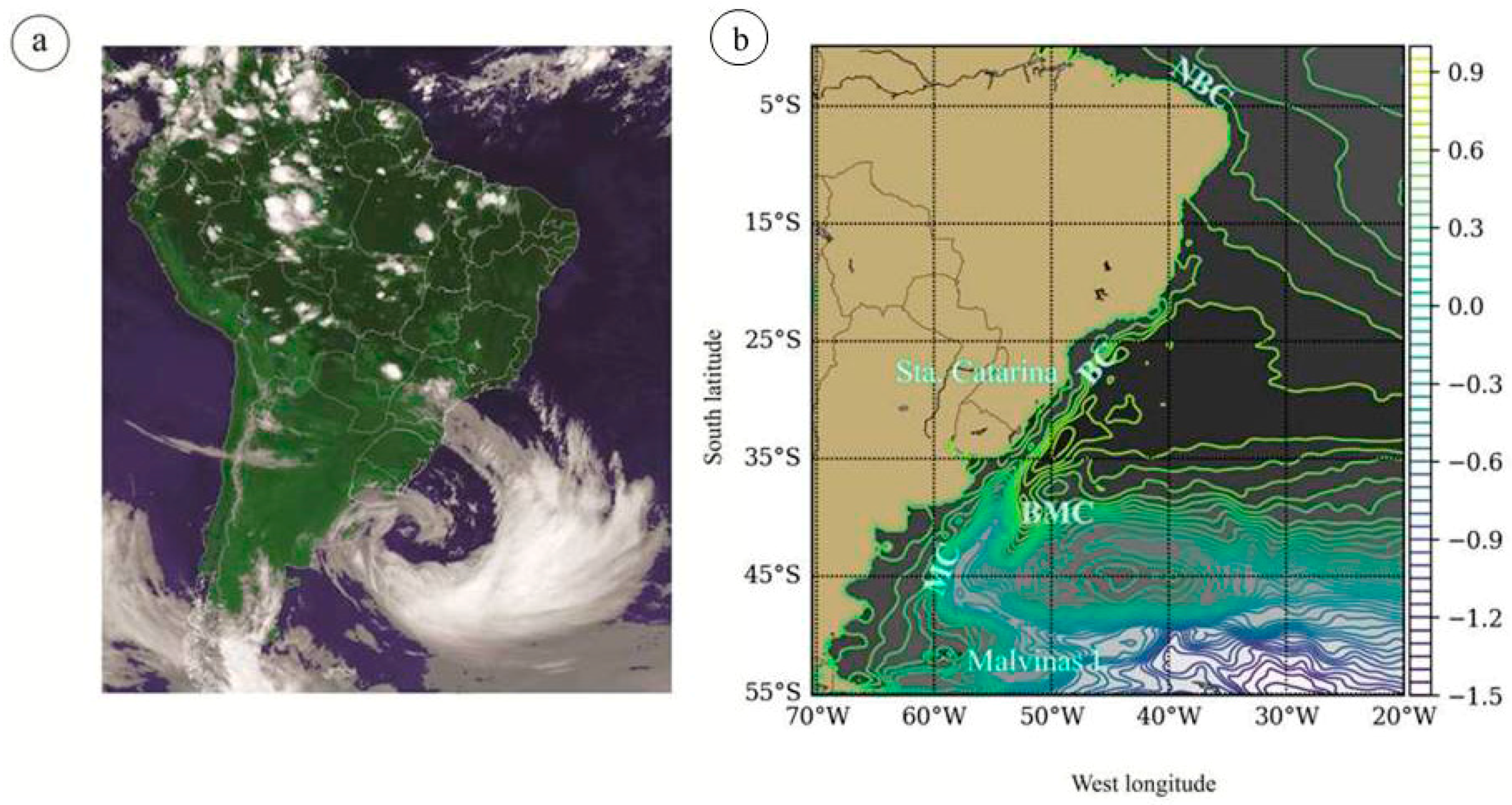
2.1. Study Area
2.2. Collection and Characterization
2.3. Niche Modeling Procedures
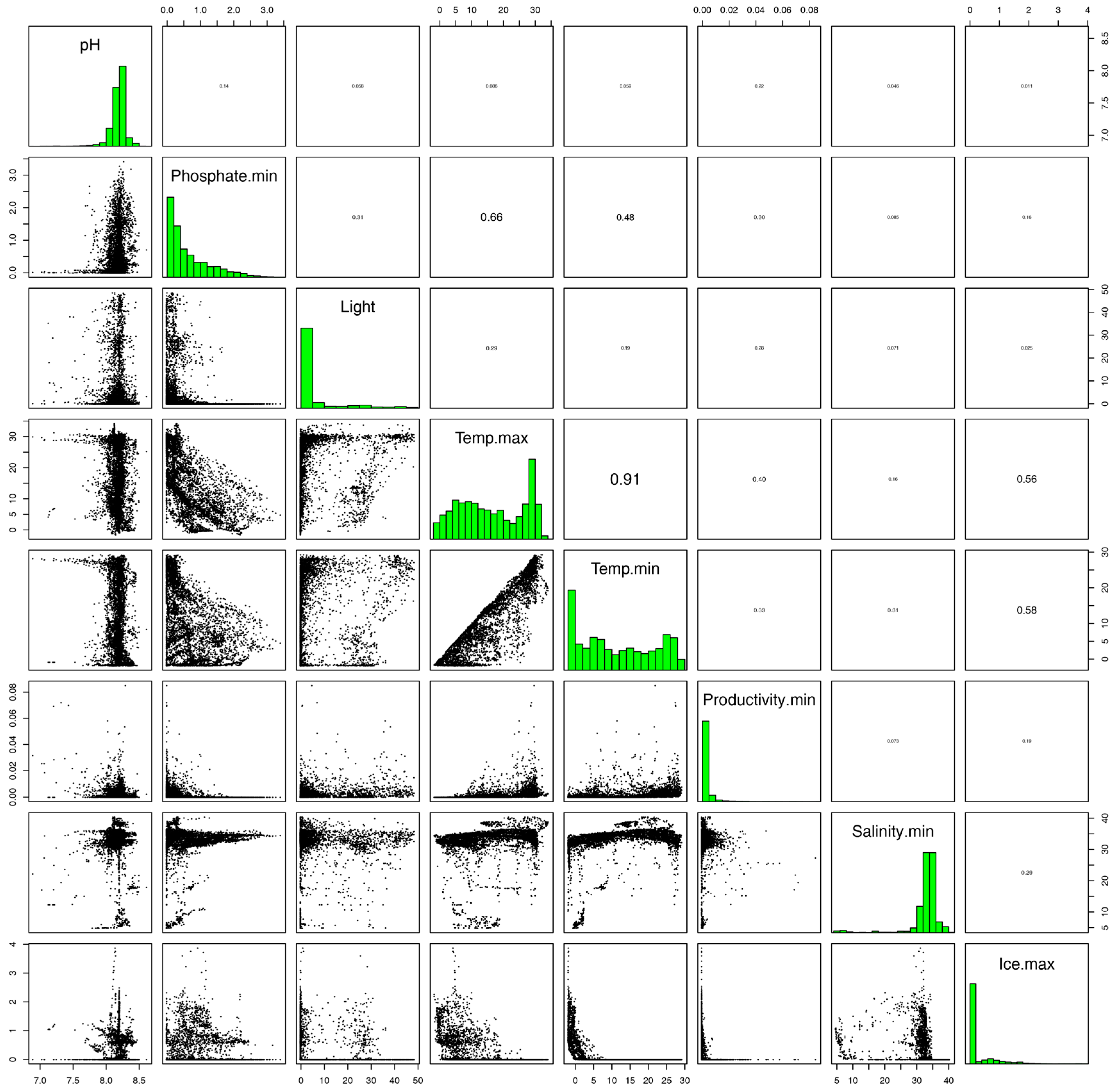
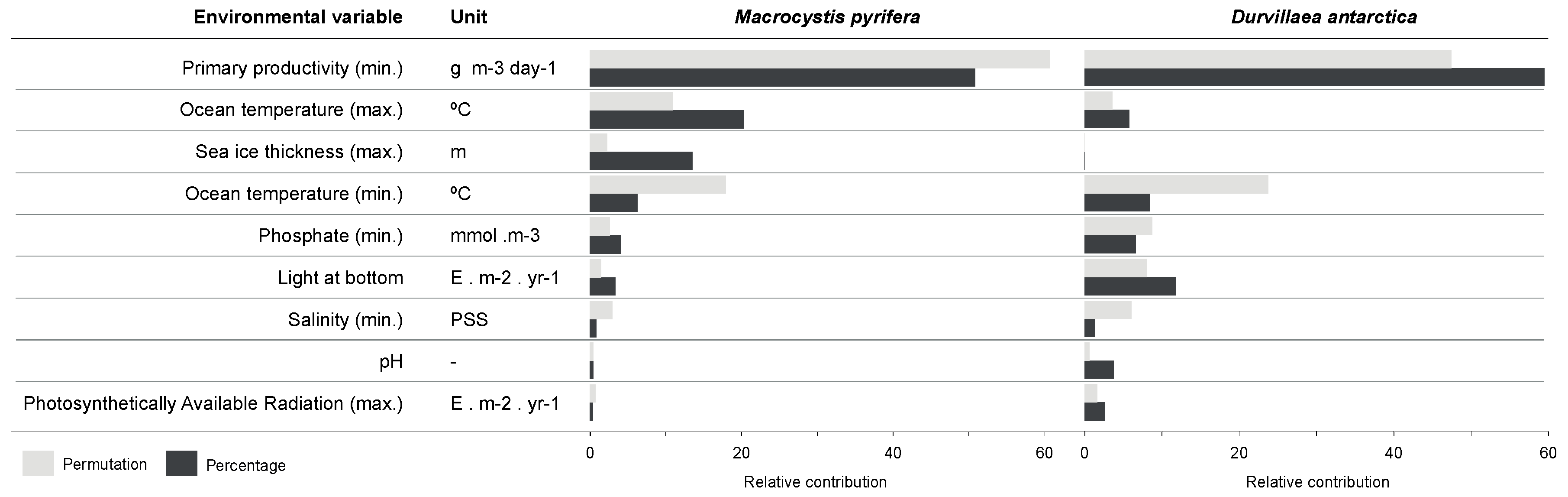
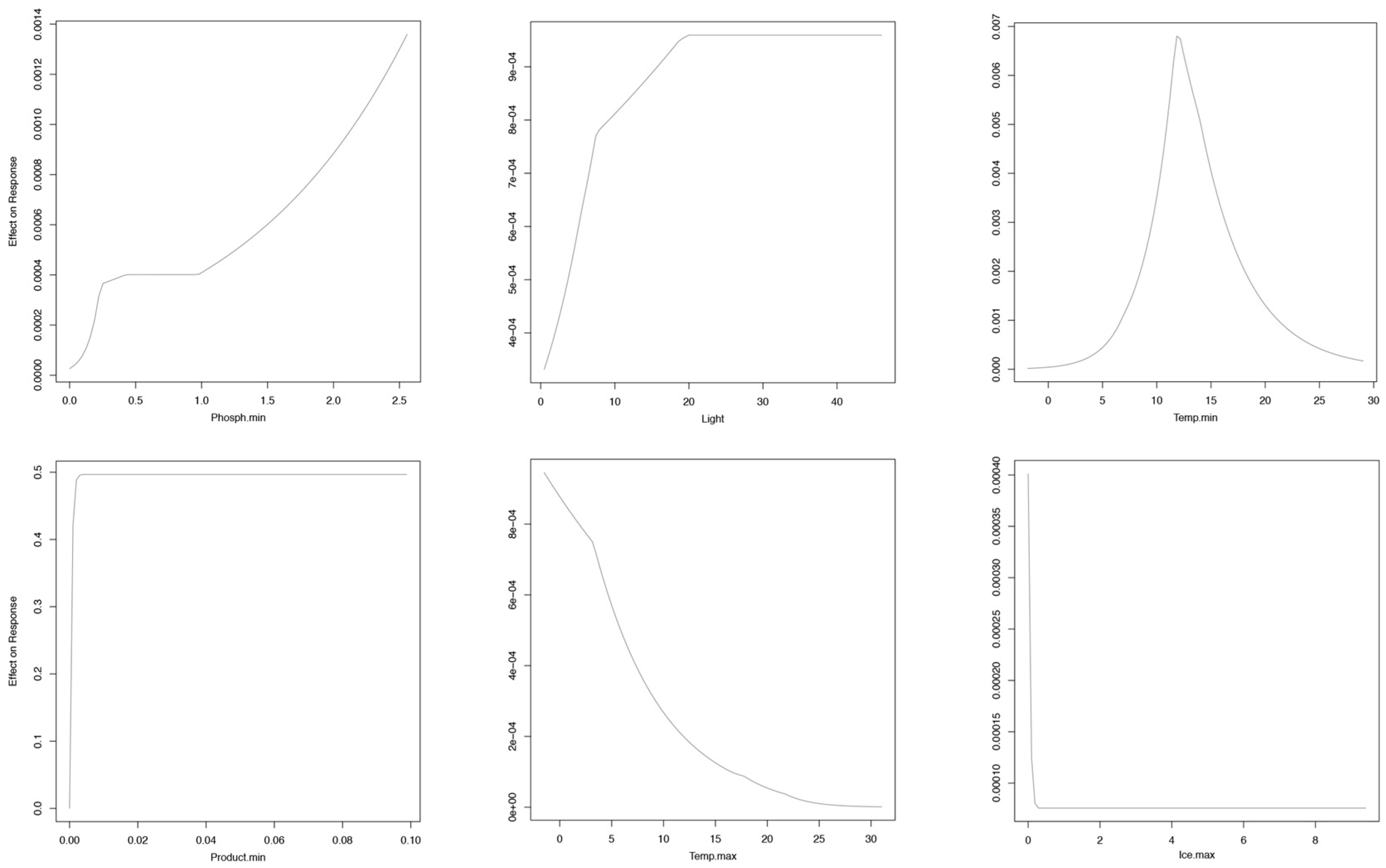
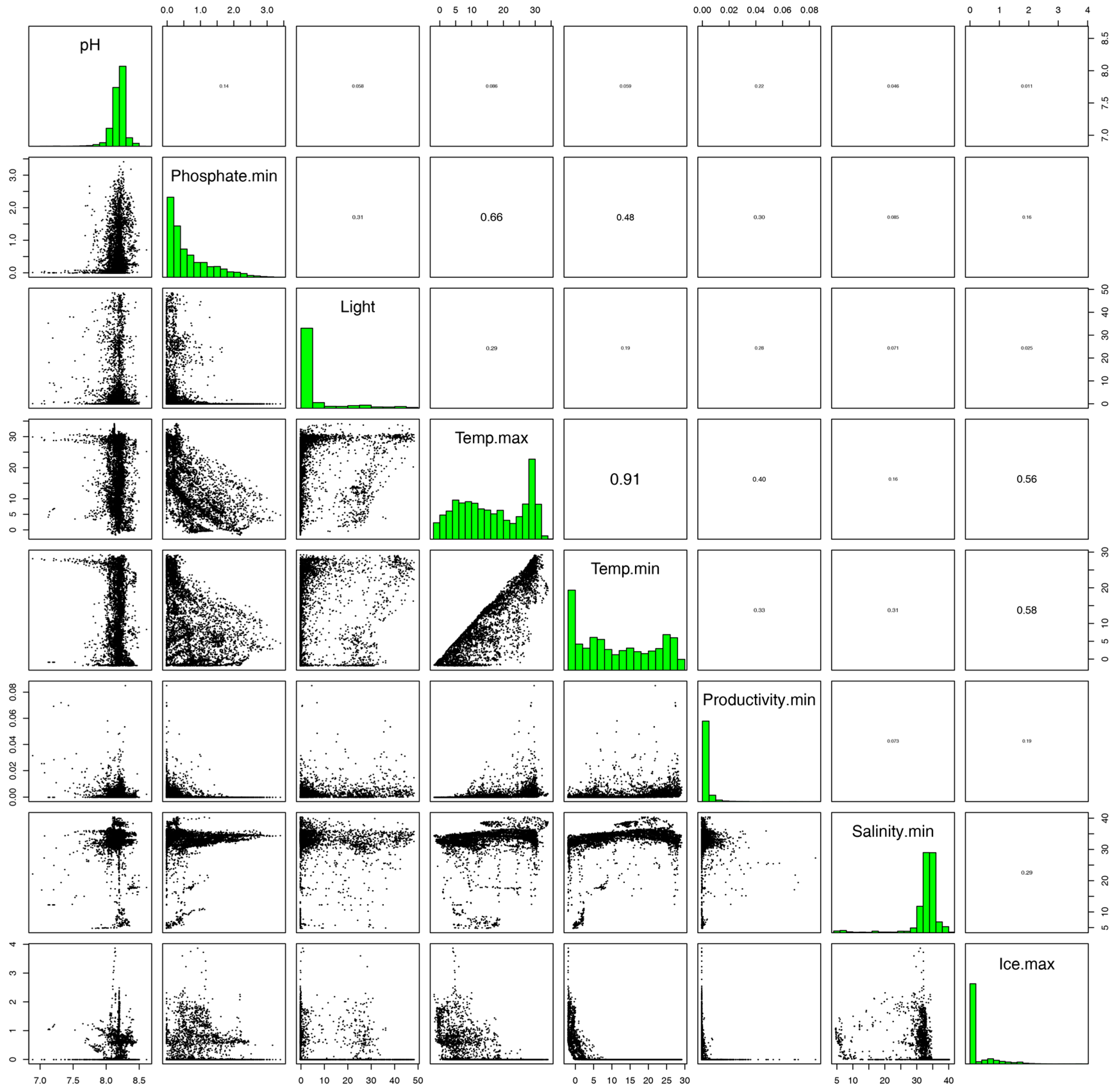
2.4. Environmental Variables and Occurrence Data
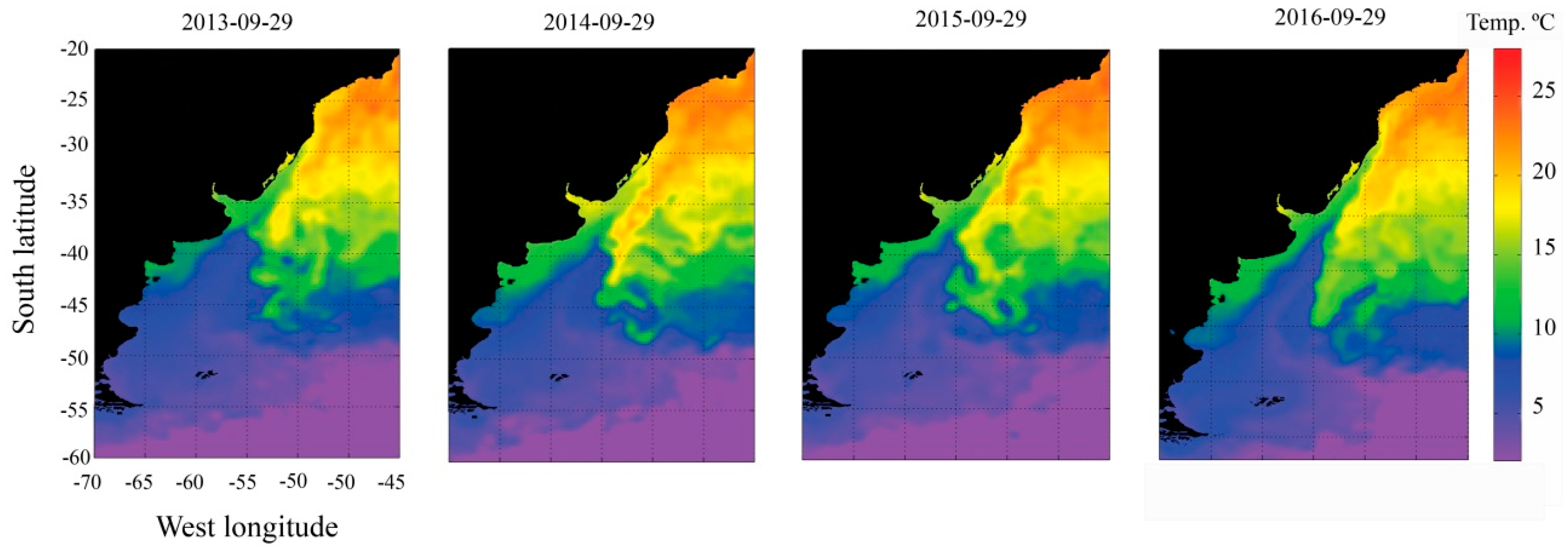
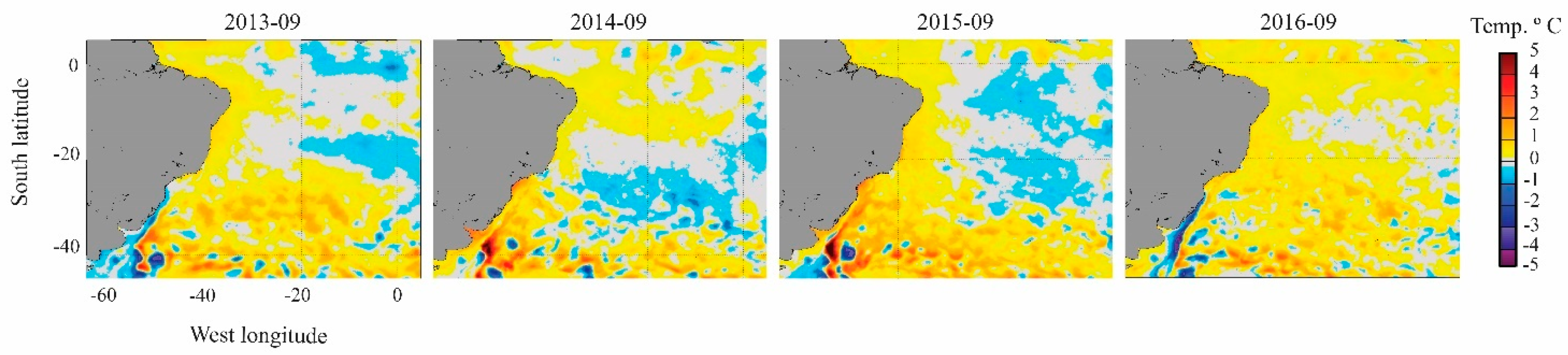
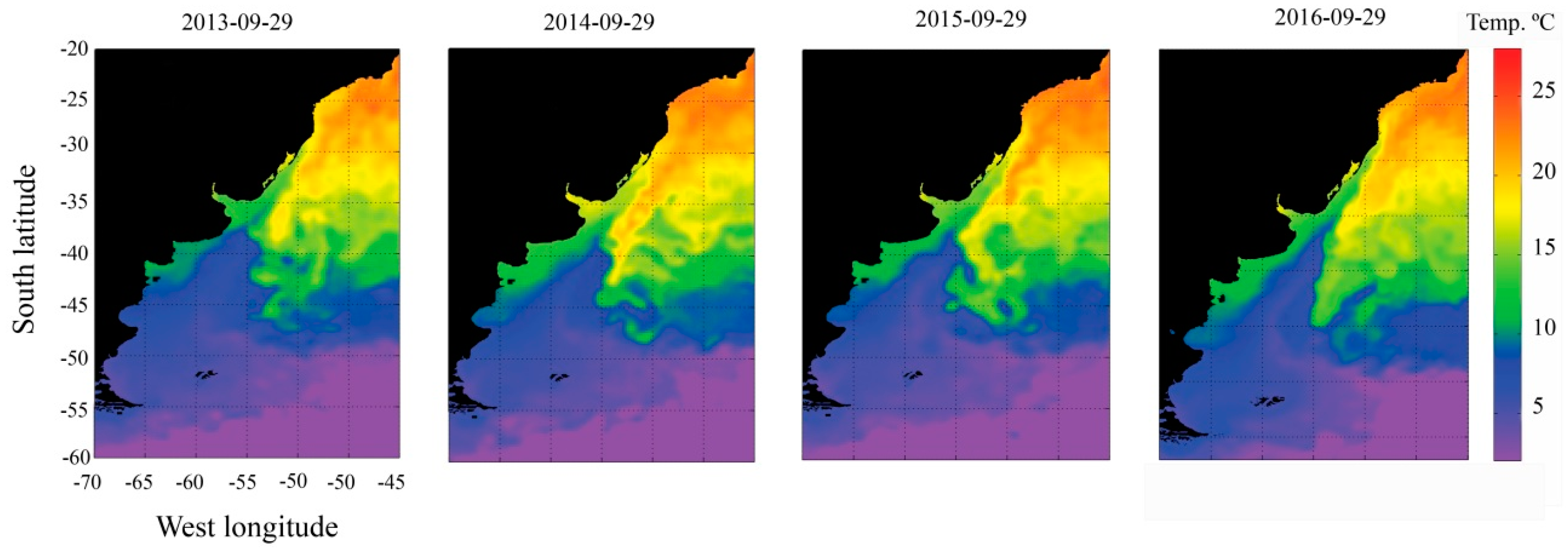
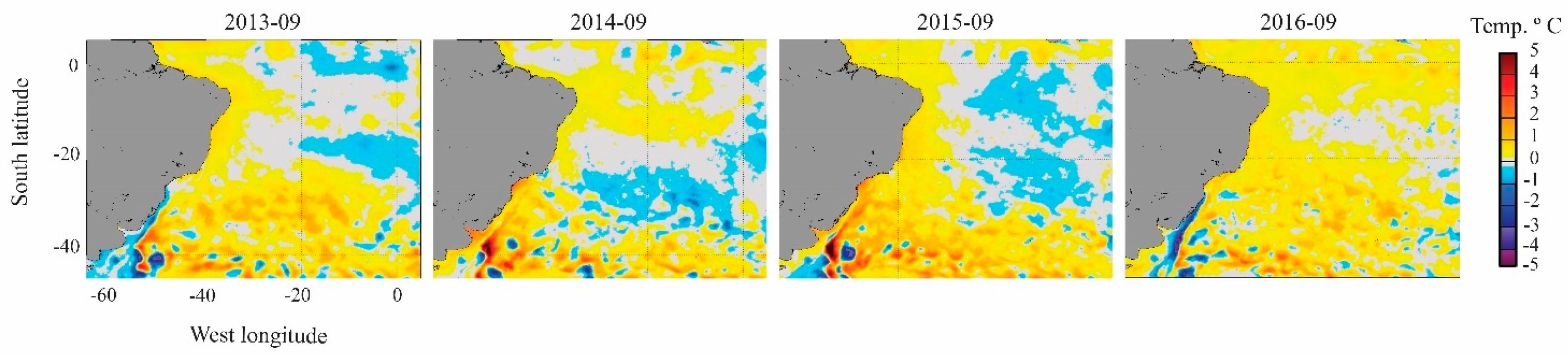
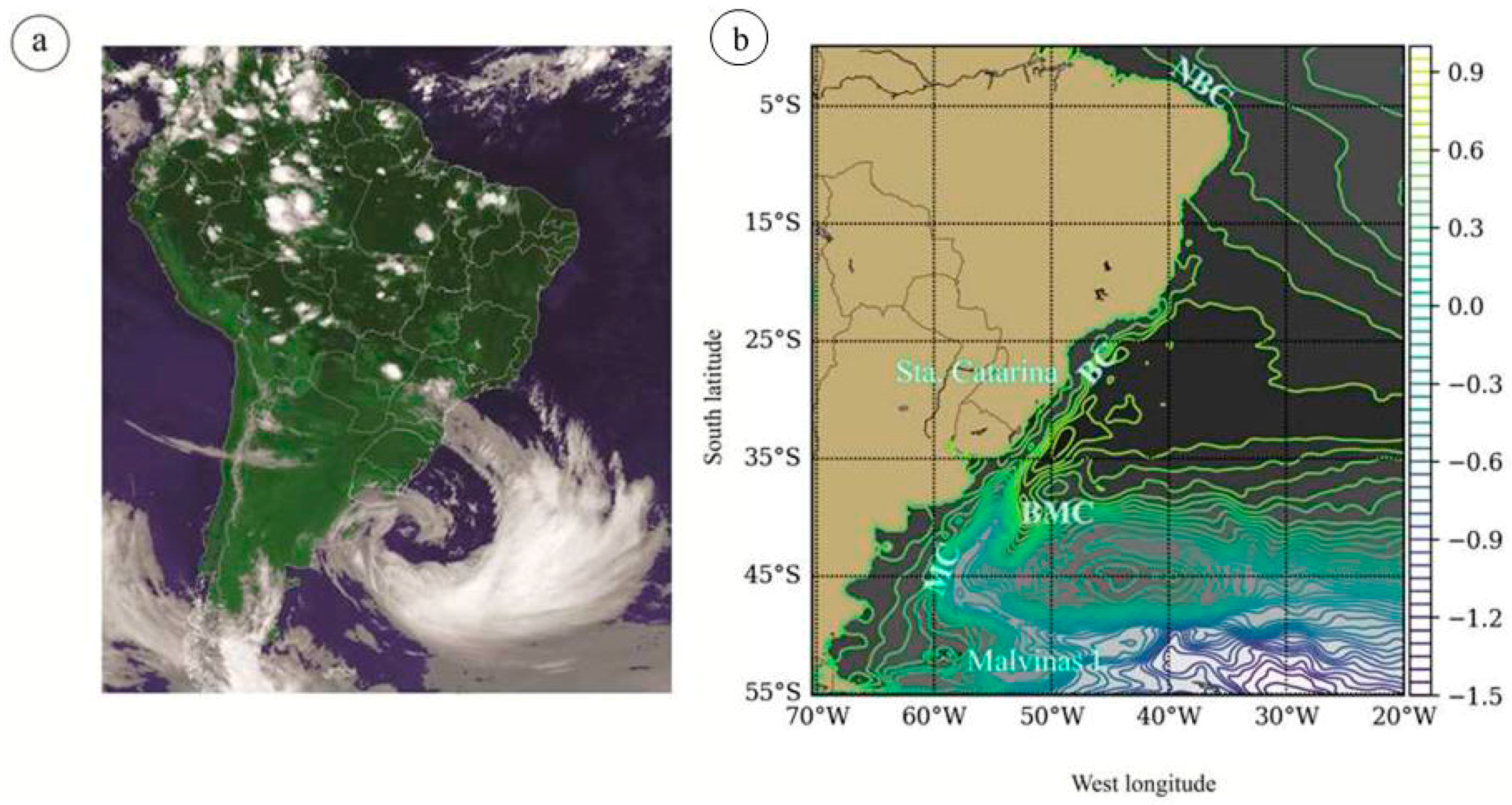
2.5. Oceanographic Characterization before and during Dispersion Event
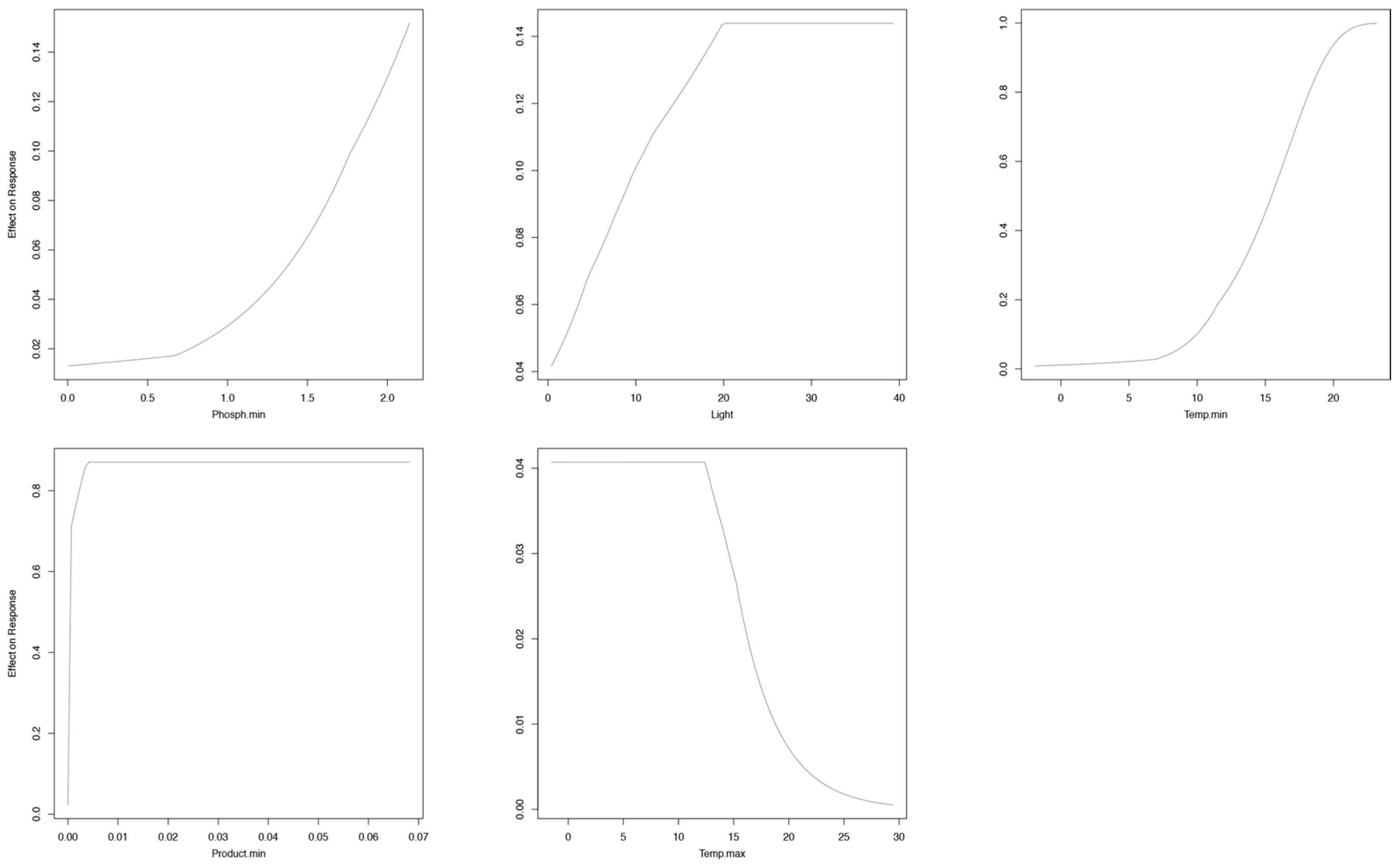
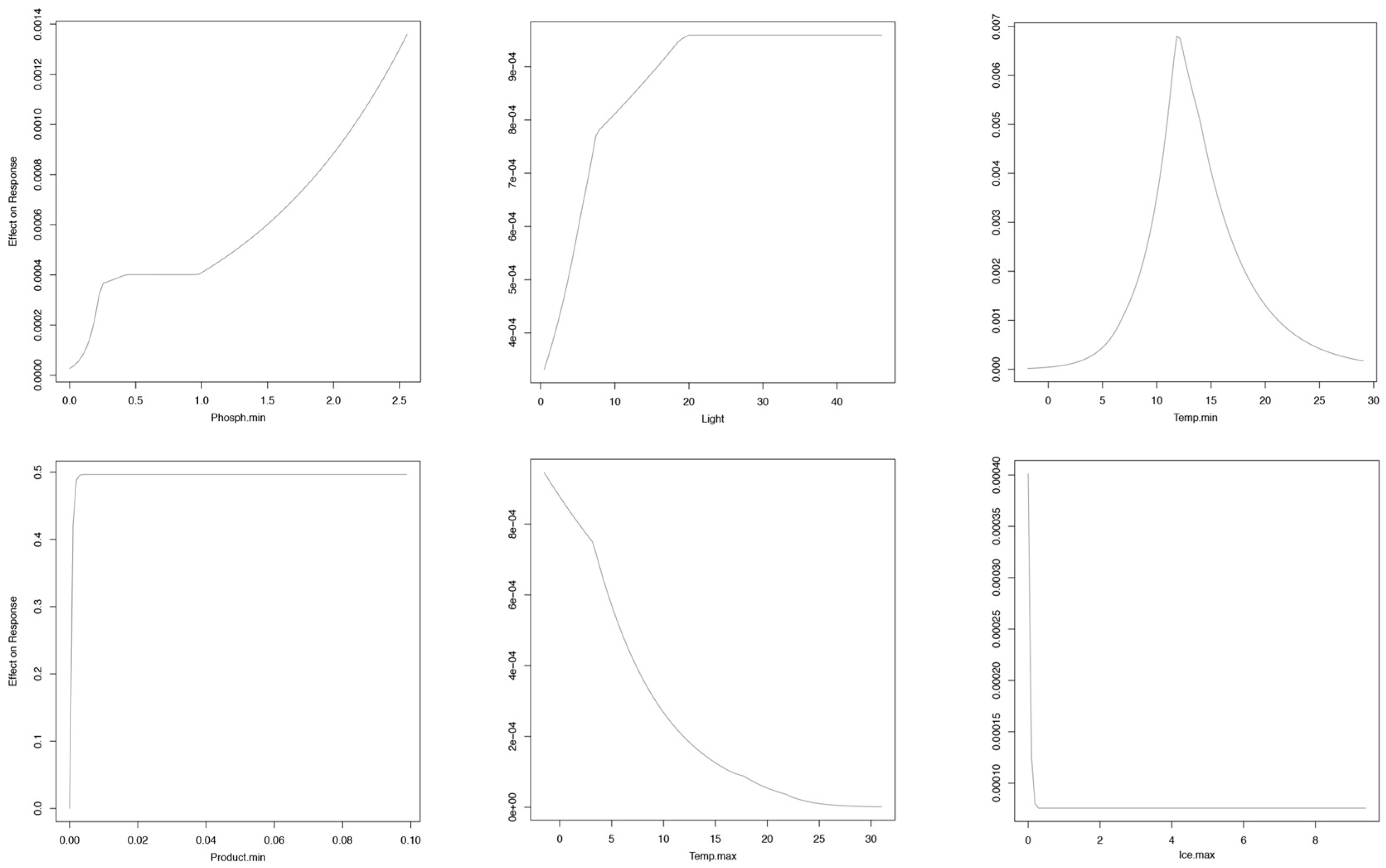
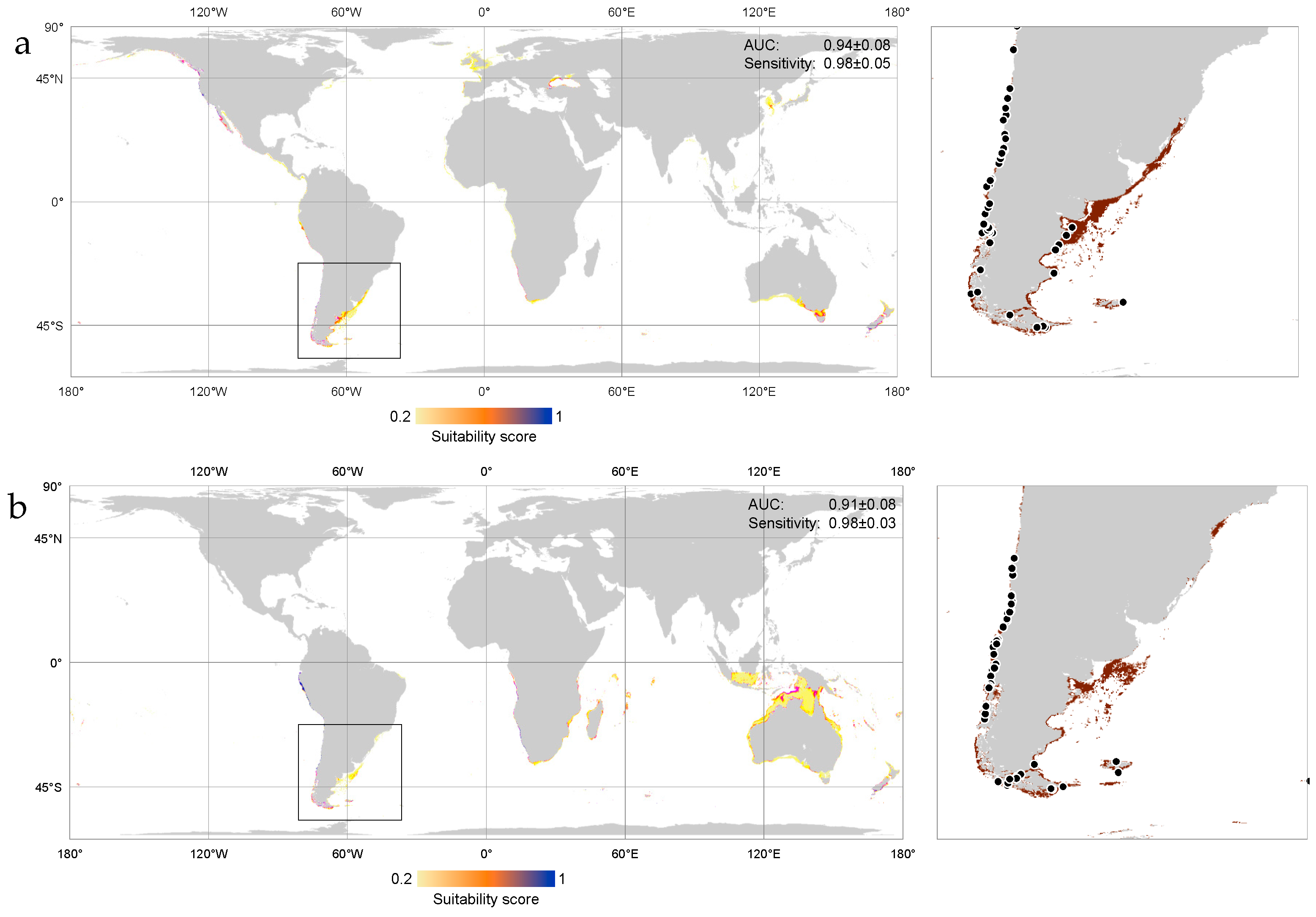
3. Results
Niche Models at the Global Scale
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Citation |
|---|---|
| A | Barrales, H.L.; Lobban, C.S. The comparative ecology of Macrocystis pyrifera, with emphasis on the forests of Chubut, Argentina. The Journal of Ecology 1975, 657–677. |
| A | Dayton, P. Kelp communities of southern south America. Antarct J US 1974. |
| A | Zimmerman, R.C.; Kremer, J.N. In situ growth and chemical composition of the giant kelp, Macrocystis pyrifera: Response to temporal changes in ambient nutrient availability. Marine Ecology Progress Series 1986, 277–285. |
| A | Macaya, E.C.; Zuccarello, G.C. Genetic structure of the giant kelp em Macrocystis pyrifera along the southeastern pacific. Marine Ecology Progress Series 2010, 420, 103–112. |
| A | Beas-Luna, R.; Ladah Lydia, B. Latitudinal, seasonal, and small-scale spatial differences of the giant kelp, Macrocystis pyrifera, and an herbivore at their southern range limit in the northern hemisphere. In Botanica Marina, 2014; Vol. 57, p. 73. |
| A | Johansson, M.L.; Alberto, F.; Reed, D.C.; Raimondi, P.T.; Coelho, N.C.; Young, M.A.; Drake, P.T.; Edwards, C.A.; Cavanaugh, K.; Assis, J. Seascape drivers of Macrocystis pyrifera population genetic structure in the northeast pacific. Molecular Ecology 2015, 24, 4866–4885. |
| A | Riosmena-Rodríguez, R.; Boo, G.H.; López-Vivas, J.M.; Hernández-Velasco, A.; Sáenz-Arroyo, A.; Boo, S.M. The invasive seaweed Sargassum filicinum (Fucales, Phaeophyceae) is on the move along the Mexican pacific coastline. Botanica Marina 2012, 55, 547–551. |
| A | Starko, S.; Martone, P.T. Evidence of an evolutionary-developmental trade-off between drag avoidance and tolerance strategies in wave-swept intertidal kelps (Laminariales, Phaeophyceae). Journal of Phycology 2016, 52, 54–63. |
| A | Ladah Lydia, B.; Zertuche-González José, A. Giant kelp (Macrocystis pyrifera) survival in deep water (25–40 m) during el Niño of 1997–1998 in Baja California, Mexico. In Botanica Marina, 2004; Vol. 47, p. 367. |
| A | Asensi, A.O.; Küpper, F.C. Seasonal periodicity and reproduction of brown algae (Phaeophyceae) at Puerto Deseado (Patagonia). Botanica Marina 2012, 55, 217–228. |
| A | Cazón, J.P.; Viera, M.; Sala, S.; Donati, E. Biochemical characterization of Macrocystis pyrifera and Undaria pinnatifida (Phaeophyceae) in relation to their potentiality as biosorbents. Phycologia 2014, 53, 100–108. |
| A | Macaya, E.C.; Boltaña, S.; Hinojosa, I.A.; Macchiavello, J.E.; Valdivia, N.A.; Vásquez, N.R.; Buschmann, A.H.; Vásquez, J.A.; Alonso Vega, J.M.; Thiel, M. Presence of sporophylls in floating kelp rafts of Macrocystis spp. (Phaeophyceae) along the Chilean pacific coast. Journal of Phycology 2005, 41, 913–922. |
| A | Engel, C.R.; Billard, M.V., Emmanuelle; Viard, F. Conservation and polymorphism of mitochondrial intergenic sequences in brown algae (Phaeophyceae). European Journal of Phycology 2008, 43, 195–205. |
| A | Rothäusler, E.; Gómez, I.; Karsten, U.; Tala, F.; Thiel, M. Physiological acclimation of floating Macrocystis pyrifera to temperature and irradiance ensures long-term persistence at the sea surface at mid-latitudes. Journal of Experimental Marine Biology and Ecology 2011, 405, 33–41. |
| A | Van Tussenbroek, B. Plant and frond dynamics of the giant kelp, Macrocysti spyrifera, forming a fringing zone in the Falkland Islands. European Journal of Phycology 1993, 28, 161–165. |
| A | Hurd, C.L.; Pilditch, C.A. Flow-induced morphological variations affect diffusion boundary-layer thickness of Macrocystis pyrifera (Heterokontophyta, Laminariales). Journal of Phycology 2011, 47, 341–351. |
| A | Ricker, R.W. Taxonomy and biogeography of Macquarie Island seaweeds. British Museum (Natural History): 1987. |
| A | John, D.M.; Pugh, P.J.; Tittley, I. Observations on the benthic marine algal flora of South Georgia: A floristic and ecological analysis. Bulletin of the Natural History Museum. Botany Series 1994. |
| A | Hay, C.H. A new species of Macrocystisc. Ag. (Phaeophyta) from Marion Island, Southern Indian ocean. Phycologia 1986, 25, 241–252. |
| B | Boraso, A.; Zaixso, J. Atlas de sensibilidad ambiental de la costa y el mar argentino. Algas marinas bentónicas 2011, 1–28. |
| B | Boraso, A.L. Elementos para el estudio de las macroalgas de argentina. Comodoro Rivadavia: Editorial Universitaria de la Patagonia EDUPA 2013. |
| B | Graiff, A.; Karsten, U.; Meyer, S.; Pfender, D.; Tala, F.; Thiel, M. Seasonal variation in floating persistence of detached Durvillaea antarctica (chamisso) hariot thalli. Botanica Marina 2013, 56, 3–14. |
| B | López, B.A.; Macaya, E.C.; Tala, F.; Tellier, F.; Thiel, M. The variable routes of rafting: Stranding dynamics of floating bull kelp Durvillaea antarctica (Fucales, Phaeophyceae) on beaches in the se pacific. Journal of Phycology 2017, 53, 70–84. |
| B | Cho, G.Y.; Rousseau, F.; De Reviers, B.; Boo, S.M. Phylogenetic relationships within the Fucales (Phaeophyceae) assessed by the photosystem i coding psa a sequences. Phycologia 2006, 45, 512–519. |
| B | Fraser, C.I.; Hay, C.H.; Spencer, H.G.; Waters, J.M. Genetic and morphological analyses of the southern bull kelp Durvillaea antarctica (Phaeophyceae: Durvillaeales) in new Zealand reveal cryptic species. Journal of Phycology 2009, 45, 436–443. |
| B | Wells, E.; Brewin, P.; Brickle, P. Intertidal and subtidal benthic seaweed diversity of South Georgia. Norfolk, UK 2011. |
| B | Ortiz, J.; Romero, N.; Robert, P.; Araya, J.; Lopez-Hernandez, J.; Bozzo, C.; Navarrete, E.; Osorio, A.; Rios, A. Dietary fiber, amino acid, fatty acid and tocopherol contents of the edible seaweeds Ulva lactuca and Durvillaea antarctica. Food Chemistry 2006, 99, 98–104. |
| B | Bustamante, R.; Castilla, J. Impact of human exploitation on populations of the intertidal southern bull-kelp Durvillaea antarctica (Phaeophyta, Durvilleales) in central Chile. Biological Conservation 1990, 52, 205–220. |
| B | Smith, S.; Simpson, R. Effects of the ‘nella dan’ oil spill on the fauna of Durvillaea antarctica holdfasts. Marine Ecology Progress Series 1995, 73–89. |
| B | Collins, C.J.; Fraser, C.I.; Ashcroft, A.; Waters, J.M. Asymmetric dispersal of southern bull-kelp (Durvillaea antarctica) adults in coastal New Zealand: Testing an oceanographic hypothesis. Molecular Ecology 2010, 19, 4572–4580. |
| B | Smith, J.; Bayliss-Smith, T. Kelp-plucking: Coastal erosion facilitated by bull-kelp Durvillaea antarctica at subantarctic Macquarie Island. Antarctic Science 1998, 10, 431–438. |
| B | Dillehay, T.D.; Ramirez, C.; Pino, M.; Collins, M.B.; Rossen, J.; Pino-Navarro, J. Monte verde: Seaweed, food, medicine, and the peopling of South America. Science 2008, 320, 784–786. |
| B | Taylor, D.I.; Schiel, D.R. Self-replacement and community modification by the southern bull kelp Durvillaea antarctica. Marine Ecology Progress Series 2005, 288, 87–102. |
| B | Chancho, M.J.R.; Sánchez, J.F.L.; Rubio, R. Occurrence of arsenic species in the seagrass Posidonia oceanica and in the marine algae Lessonia nigrescens and Durvillaea antarctica. Journal of Applied Phycology 2010, 22, 465–472. |
| B | Castilla, J.; Campo, M.; Bustamante, R. Recovery of Durvillaea antarctica (durvilleales) inside and outside las cruces marine reserve, Chile. Ecological Applications 2007, 17, 1511–1522. |
| B | Smith, S.D. Kelp rafts in the Southern Ocean. Global Ecology and Biogeography 2002, 11, 67–69. |
| B | Matsuhiro, B.; Zúñiga, E.; Jashes, M.; Guacucano, M. Sulfated polysaccharides from Durvillaea antarctica. Hydrobiologia 1996, 321, 77–81. |
| B | Kelly, B.J.; Brown, M.T. Variations in the alginate content and composition of Durvillaea antarctica and D. willana from southern New Zealand. Journal of Applied Phycology 2000, 12, 317–324. |
| B | Collantes, G.; Merino, A.; Lagos, V. Fenología de la gametogénesis, madurez de conceptáculos, fertilidad y embriogénesis en Durvillaea antarctica (chamisso) Hariot (Phaeophyta, Durvillaeales). Revista de Biología Marina y Oceanografía 2002, 37, 83–112. |
| B | Tala, F.; Gómez, I.; Luna-Jorquera, G.; Thiel, M. Morphological, physiological and reproductive conditions of rafting bull kelp (Durvillaea antarctica) in northern-central Chile (30 s). Marine Biology 2013, 160, 1339–1351. |
| B | Castilla, J.C.; Defeo, O. Latin American benthic shellfisheries: Emphasis on co-management and experimental practices. Reviews in Fish Biology and Fisheries 2001, 11, 1–30. |
| B | Nikula, R.; Fraser, C.I.; Spencer, H.G.; Waters, J.M. Circumpolar dispersal by rafting in two subantarctic kelp-dwelling crustaceans. Marine Ecology Progress Series 2010, 405, 221–230. |
| B | Stevens, C.L.; Hurd, C.L.; Smith, M.J. An idealized model of interaction between fronds of the large seaweed Durvillaea antarctica. Journal of Marine Systems 2004, 49, 145–156. |
| B | Goecke, F.; Wiese, J.; Núñez, A.; Labes, A.; Imhoff, J.F.; Neuhauser, S. A novel phytomyxean parasite associated with galls on the bull-kelp Durvillaea antarctica (Chamisso) Hariot. PLoS ONE 2012, 7, e45358. |
| B | Castilla, J.C.; Manríquez, P.H.; Camaño, A. Effects of rocky shore coseismic uplift and the 2010 Chilean mega-earthquake on intertidal biomarker species. Marine Ecology Progress Series 2010, 418, 17–23. |
| B | Miller, I.J. Alginate composition of some New Zealand brown seaweeds. Phytochemistry 1996, 41, 1315–1317. |
| B | Fraser, C.I.; Kay, G.M.; Plessis, M.d.; Ryan, P.G. Breaking down the barrier: Dispersal across the Antarctic Polar Front. Ecography 2017, 40, 235–237. |
| B | Duarte, C.; Acuña, K.; Navarro, J.M.; Gómez, I. Intra-plant differences in seaweed nutritional quality and chemical defenses: Importance for the feeding behavior of the intertidal amphipod Orchestoidea tuberculata. Journal of Sea Research 2011, 66, 215–221. |
| B | Astorga-España, M.S.; Mansilla, A. Sub-antarctic macroalgae: Opportunities for gastronomic tourism and local fisheries in the region of Magallanes and Chilean Antarctic territory. Journal of Applied Phycology 2014, 26, 973–978. |
| B | Beckley, L.E.; Branch, G.M. A quantitative scuba-diving survey of the sublittoral macrobenthos at subantarctic Marion Island. Polar Biology 1992, 11, 553–563. |
| B | Bussolini, L.T.; Waters, J.M. Genetic analyses of rafted macroalgae reveal regional oceanographic connectivity patterns. Journal of Biogeography 2015, 42, 1319–1326. |
| B | Cruces, E.; Huovinen, P.; Gomez, I. Phlorotannin and antioxidant responses upon short-term exposure to uv radiation and elevated temperature in three south pacific kelps. Photochemistry and Photobiology 2012, 88, 58–66. |
| B | Díaz, O.; Tapia, Y.; Muñoz, O.; Montoro, R.; Velez, D.; Almela, C. Total and inorganic arsenic concentrations in different species of economically important algae harvested from coastal zones of Chile. Food and Chemical Toxicology 2012, 50, 744–749. |
| B | Dimartino, S.; Savory, D.M.; Fraser-Miller, S.J.; Gordon, K.C.; McQuillan, A.J. Microscopic and infrared spectroscopic comparison of the underwater adhesives produced by germlings of the brown seaweed species Durvillaea antarctica and Hormosira banksii. Journal of The Royal Society Interface 2016, 13, 2015–1083. |
| B | Dufour, C.; Probert, P.; Savage, C. Macrofaunal colonisation of stranded Durvillaea antarctica on a southern New Zealand exposed sandy beach. New Zealand Journal of Marine and Freshwater Research 2012, 46, 369–383. |
| B | Fraser, C.I.; Nikula, R.; Waters, J.M. Oceanic rafting by a coastal community. Proceedings of the Royal Society of London B: Biological Sciences 2011, 278, 649–655. |
| B | González, A.V.; Beltrán, J.; Flores, V.; Santelices, B. Morphological convergence in the inter-holdfast coalescence process among kelp and kelp-like seaweeds (Lessonia, Macrocystis, Durvillaea). Phycologia 2015, 54, 283–291. |
| B | Garden, C.J.; Smith, A.M. The role of kelp in sediment transport: Observations from southeast New Zealand. Marine Geology 2011, 281, 35–42. |
| B | Haye, P.A.; Varela, A.I.; Thiel, M. Genetic signatures of rafting dispersal in algal-dwelling brooders Limnoria spp. (isopoda) along the se Pacific (Chile). Marine Ecology Progress Series 2012, 455, 111–122. |
| B | Hinojosa, I.; González, E.; Ugalde, P.; Valdivia, N.; Macaya, E.; Thiel, M. Distribución y abundancia de macroalgas flotando a la deriva y su fauna peracarida asociada en los canales de la xl región, Chile. Ciencia y Tecnología del Mar 2007, 30. |
| B | Huovinen, P.; Leal, P.; Gómez, I. Interacting effects of copper, nitrogen and ultraviolet radiation on the physiology of three south pacific kelps. Marine and Freshwater Research 2010, 61, 330–341. |
| B | Fraser, C.I.; Thiel, M.; Spencer, H.G.; Waters, J.M. Contemporary habitat discontinuity and historic glacial ice drive genetic divergence in Chilean kelp. BMC Evolutionary Biology 2010, 10, 203. |
| B | Jaramillo, E.; De la Huz, R.; Duarte, C.; Contreras, H. Algal wrack deposits and macroinfaunal arthropods on sandy beaches of the Chilean coast. Revista Chilena de Historia Natural 2006, 79. |
| B | Lizée-Prynne, D.; López, B.; Tala, F.; Thiel, M. No sex-related dispersal limitation in a dioecious, oceanic long-distance traveller: The bull kelp durvillaea antarctica. In Botanica Marina, 2016; Vol. 59, p. 39. |
| B | Miranda, L.; Thiel, M. Active and passive migration in boring isopods Limnoria spp. (Crustacea, Peracarida) from kelp holdfasts. Journal of Sea Research 2008, 60, 176–183. |
| B | Nikula, R.; Spencer, H.; Waters, J. Evolutionary consequences of microhabitat: Population-genetic structuring in kelp-vs. Rock-associated chitons. Molecular Ecology 2011, 20, 4915–4924. |
| B | Smith, S.D.A. The effects of a small sewage outfall on an algal epifaunal community at Macquarie Island (sub-antarctic): A drop in the southern ocean? Marine Pollution Bulletin 2000, 40, 873–878. |
| B | Smith, J.; Summers, G.; Wong, R. Nutrient and heavy metal content of edible seaweeds in New Zealand. New Zealand Journal of Crop and Horticultural Science 2010, 38, 19–28. |
| B | Taylor, D.I.; Schiel, D.R. Wave-related mortality in zygotes of habitat-forming algae from different exposures in southern New Zealand: The importance of ‘stickability’. Journal of Experimental Marine Biology and Ecology 2003, 290, 229–245. |
| B | Alestra, T.; Schiel, D. Impacts of local and global stressors in intertidal habitats: Influence of altered nutrient, sediment and temperature levels on the early life history of three habitat-forming macroalgae. Journal of Experimental Marine Biology and Ecology 2015, 468, 29–36. |
| B | Anderson, M.J.; Diebel, C.E.; Blom, W.M.; Landers, T.J. Consistency and variation in kelp holdfast assemblages: Spatial patterns of biodiversity for the major phyla at different taxonomic resolutions. Journal of Experimental Marine Biology and Ecology 2005, 320, 35–56. |
| B | Collantes, G.; Riveros, R.; Acevedo, M. (fenologia reproductiva de Durvillaea antarctica (Phaeophyta, Durvillaeales) del intermareal de caleta Montemar, Chile Central). Oceanographic Literature Review 1998, 8, 1412. |
| B | De Souza, B.W.S.; Teixeira, D.I.A.; Andrade, F.K.; Melo, M.R.S.; Muñoz, A.M.; Freitas, A.L.P. A survey of Antarctic algae for agglutinins. Oecologia Australis 2009, 11, 122–130. |
| B | Eriksson, C.; Burton, H.; Fitch, S.; Schulz, M.; van den Hoff, J. Daily accumulation rates of marine debris on sub-antarctic island beaches. Marine Pollution Bulletin 2013, 66, 199–208. |
| B | Tait, L.W.; South, P.M.; Lilley, S.A.; Thomsen, M.S.; Schiel, D.R. Assemblage and understory carbon production of native and invasive canopy-forming macroalgae. Journal of Experimental Marine Biology and Ecology 2015, 469, 10–17. |
| B | Tala, F.; Velásquez, M.; Mansilla, A.; Macaya, E.C.; Thiel, M. Latitudinal and seasonal effects on short-term acclimation of floating kelp species from the south-east pacific. Journal of Experimental Marine Biology and Ecology 2016, 483, 31–41. |
| B | Aguilera, M.A.; Valdivia, N.; Broitman, B.R. Herbivore-alga interaction strength influences spatial heterogeneity in a kelp-dominated intertidal community. PloS one 2015, 10, e0137287. |
| B | Astorga-España, M.S.; Mansilla, A.; Ojeda, J.; Marambio, J.; Rosenfeld, S.; Mendez, F.; Rodriguez, J.; Ocaranza, P. Nutritional properties of dishes prepared with sub-antarctic macroalgae an opportunity for healthy eating. Journal of Applied Phycology 2017, 1–8. |
| B | Mansilla, A.O.; Avila, M.; Cáceres, J. Reproductive biology of Durvillaea antarctica (Chamisso) Hariot in the sub-antartic ecoregion of Magallanes (51–56° s). Journal of Applied Phycology 2017, 1–8. |
References
- MacArthur, R.H.; Wilson, E.O. Theory of Island Biogeography (MPB-1); Princeton University Press: Princeton, NJ, USA, 2015; Volume 1, p. 216. [Google Scholar]
- Jackson, S.T.; Sax, D.F. Balancing biodiversity in a changing environment: Extinction debt, immigration credit and species turnover. Trends Ecol. Evol. 2010, 25, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, H.T.; Bernardi, G.; Simon, T.; Joyeux, J.-C.; Macieira, R.M.; Gasparini, J.L.; Rocha, C.; Rocha, L.A. Island biogeography of marine organisms. Nature 2017, 549, 82–85. [Google Scholar] [CrossRef] [PubMed]
- Mackas, D.L.; Denman, K.L.; Abbott, M.R. Plankton patchiness: Biology in the physical vernacular. Bull. Mar. Sci. 1985, 37, 652–674. [Google Scholar]
- Gilg, M.R.; Hilbish, T.J. The geography of marine larval dispersal: Coupling genetics with fine-scale physical oceanography. Ecology 2003, 84, 2989–2998. [Google Scholar] [CrossRef]
- Zakas, C.; Binford, J.; Navarrete, S.A.; Wares, J.P. Restricted gene flow in Chilean barnacles reflects an oceanographic and biogeographic transition zone. Mar. Ecol. Prog. Ser. 2009, 394, 165–177. [Google Scholar] [CrossRef]
- Gaylord, B.; Gaines, S.D. Temperature or transport? Range limits in marine species mediated solely by flow. Am. Nat. 2000, 155, 769–789. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.H. Macroecology; University of Chicago Press: Chicago, IL, USA, 1995; p. 284. [Google Scholar]
- Brown, J.H.; Maurer, B.A. Macroecology: The division of food and space among species on continents. Science 1989, 243, 1145–1150. [Google Scholar] [CrossRef] [PubMed]
- Beal, L.M.; De Ruijter, W.P.M.; Biastoch, A.; Zahn, R. On the role of the agulhas system in ocean circulation and climate. Nature 2011, 472, 429–436. [Google Scholar] [CrossRef] [PubMed]
- O’Gorman, E.J. It's only a matter of time: The altered role of subsidies in a warming world. J. Anim. Ecol. 2016, 85, 1133–1135. [Google Scholar] [CrossRef] [PubMed]
- Sissini, M.N.; Oliveira, M.C.; Gabrielson, P.W.; Robinson, N.M.; Okolodkov, Y.B.; Riosmena-Rodríguez, R.; Horta, P.A. Mesophyllum erubescens (corallinales, rhodophyta)-so many species in one epithet. Phytotaxa 2014, 190, 299–319. [Google Scholar] [CrossRef]
- Knutson, T.R.; McBride, J.L.; Chan, J.; Emanuel, K.; Holland, G.; Landsea, C.; Held, I.; Kossin, J.P.; Srivastava, A.K.; Sugi, M. Tropical cyclones and climate change. Nat. Geosci. 2010, 3, 157–163. [Google Scholar] [CrossRef] [Green Version]
- Coumou, D.; Rahmstorf, S. A decade of weather extremes. Nat. Clim. Chang. 2012, 2, 491–496. [Google Scholar] [CrossRef]
- Easterling, D.R.; Meehl, G.A.; Parmesan, C.; Changnon, S.A.; Karl, T.R.; Mearns, L.O. Climate extremes: Observations, modeling, and impacts. Science 2000, 289, 2068–2074. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.-C.; Hill, J.K.; Ohlemüller, R.; Roy, D.B.; Thomas, C.D. Rapid range shifts of species associated with high levels of climate warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef] [PubMed]
- Pearson, R.G.; Dawson, T.P. Predicting the impacts of climate change on the distribution of species: Are bioclimate envelope models useful? Glob. Ecol. Biogeogr. 2003, 12, 361–371. [Google Scholar] [CrossRef]
- Anderson, A.B.; Salas, E.M.; Rocha, L.A.; Floeter, S.R. The recent colonization of south brazil by the azores chromis chromis limbata. J. Fish Biol. 2017, 91, 558–573. [Google Scholar] [CrossRef] [PubMed]
- Tegner, M.J.; Dayton, P.K.; Edwards, P.B.; Riser, K.L. Large-scale, low-frequency oceanographic effects on kelp forest succession: A tale of two cohorts. Mar. Ecol. Prog. Ser. 1997, 146, 117–134. [Google Scholar] [CrossRef]
- Garbary, D.J. Biogeography of marine algae. In eLS; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2001; pp. 1–9. [Google Scholar]
- Soberón, J.; Nakamura, M. Niches and distributional areas: Concepts, methods, and assumptions. Proc. Natl. Acad. Sci. USA 2009, 106, 19644–19650. [Google Scholar] [CrossRef] [PubMed]
- Bruno, J.F.; Stachowicz, J.J.; Bertness, M.D. Inclusion of facilitation into ecological theory. Trends Ecol. Evol. 2003, 18, 119–125. [Google Scholar] [CrossRef]
- Thomsen, M.; McGlathery, K. Facilitation of macroalgae by the sedimentary tube forming polychaete Diopatra cuprea. Estuar. Coast. Shelf Sci. 2005, 62, 63–73. [Google Scholar] [CrossRef]
- Hutchinson, G.E. Cold spring harbor symposium on quantitative biology. Concluding Remarks 1957, 22, 415–427. [Google Scholar]
- Dayton, P.K.; Tegner, M.J. Catastrophic storms, el niño, and patch stability in a southern california kelp community. Science 1984, 224, 283–285. [Google Scholar] [CrossRef] [PubMed]
- Dayton, P.K.; Tegner, M.J.; Edwards, P.B.; Riser, K.L. Temporal and spatial scales of kelp demography: The role of oceanographic climate. Ecol. Monogr. 1999, 69, 219–250. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol., Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Peterson, A.T.; Vieglais, D.A. Predicting species invasions using ecological niche modeling: New approaches from bioinformatics attack a pressing problem: A new approach to ecological niche modeling, based on new tools drawn from biodiversity informatics, is applied to the challenge of predicting potential species’ invasions. BioScience 2001, 51, 363–371. [Google Scholar]
- Tyberghein, L.; Verbruggen, H.; Pauly, K.; Troupin, C.; Mineur, F.; De Clerck, O. Bio-oracle: A global environmental dataset for marine species distribution modelling. Glob. Ecol. Biogeogr. 2012, 21, 272–281. [Google Scholar] [CrossRef]
- Gotelli, N.J.; Stanton-Geddes, J. Climate change, genetic markers and species distribution modelling. J. Biogeogr. 2015, 42, 1577–1585. [Google Scholar] [CrossRef]
- Graham, M.H.; Kinlan, B.P.; Druehl, L.D.; Garske, L.E.; Banks, S. Deep-water kelp refugia as potential hotspots of tropical marine diversity and productivity. Proc. Natl. Acad. Sci. USA 2007, 104, 16576–16580. [Google Scholar] [CrossRef] [PubMed]
- Verbruggen, H.; Leliaert, F.; Maggs, C.A.; Shimada, S.; Schils, T.; Provan, J.; Booth, D.; Murphy, S.; De Clerck, O.; Littler, D.S.; et al. Species boundaries and phylogenetic relationships within the green algal genus codium (bryopsidales) based on plastid DNA sequences. Mol. Phylogenet. Evol. 2007, 44, 240–254. [Google Scholar] [CrossRef] [PubMed]
- Assis, V.D.L.; Carvalho, T.S.G.; Pereira, V.M.; Freitas, R.T.F.; Saad, C.E.P.; Costa, A.C.; Silva, A.A.A. Environmental enrichment on the behavior and welfare of cockatiels (nymphicus hollandicus). Arq. Bras. Med. Vet. Zootec. 2016, 68, 562–570. [Google Scholar] [CrossRef]
- Highsmith, R.C. Floating and algal rafting as potential dispersal mechanisms in brooding invertebrates. Mar. Ecol. Prog. Ser. 1985, 25, 169–179. [Google Scholar] [CrossRef]
- Steneck, R.S.; Graham, M.H.; Bourque, B.J.; Corbett, D.; Erlandson, J.M.; Estes, J.A.; Tegner, M.J. Kelp forest ecosystems: Biodiversity, stability, resilience and future. Environ. Conserv. 2003, 29, 436–459. [Google Scholar] [CrossRef]
- Dayton, P.K. Ecology of kelp communities. Annu. Rev. Ecol. Syst. 1985, 16, 215–245. [Google Scholar] [CrossRef]
- Darwin, C. The descent ofman. In The Great Books of the Western World; Encyclopædia Britannica, Inc.: Chatswood, NSW, Australia, 1871; Volume 49, p. 320. [Google Scholar]
- Macaya, E.C.; Zuccarello, G.C. Genetic structure of the giant kelp Macrocystis pyrifera along the southeastern pacific. Mar. Ecol. Prog. Ser. 2010, 420, 103–112. [Google Scholar] [CrossRef]
- Griffiths, H.J.; Waller, C.L. The first comprehensive description of the biodiversity and biogeography of antarctic and sub-antarctic intertidal communities. J. Biogeogr. 2016, 43, 1143–1155. [Google Scholar] [CrossRef] [Green Version]
- Kuhnemann, O. Algunas consideraciones sobre los bosques de macrocystis pyrifera. Physis 1970, 25, 273–296. [Google Scholar]
- Raffo, M.P.; Eyras, M.C.; Iribarne, O.O. The invasion of undaria pinnatifida to a Macrocystis pyrifera kelp in Patagonia (argentina, south-west atlantic). J. Mar. Biol. Assoc. UK 2009, 89, 1571–1580. [Google Scholar] [CrossRef]
- Rothman, M.D.; Mattio, L.; Anderson, R.J.; Bolton, J.J. A phylogeographic investigation of the kelp genus laminaria (laminariales, phaeophyceae), with emphasis on the South Atlantic Ocean. J. Phycol. 2017, 4, 778–789. [Google Scholar] [CrossRef] [PubMed]
- Westermeier, R.; Patiño, D.J.; Murúa, P.; Müller, D.G. Macrocystis mariculture in chile: Growth performance of heterosis genotype constructs under field conditions. J. Appl. Phycol. 2011, 23, 819–825. [Google Scholar] [CrossRef]
- Ladah, L.B.; Zertuche-González, J.A. Survival of microscopic stages of a perennial kelp (Macrocystis pyrifera) from the center and the southern extreme of its range in the northern hemisphere after exposure to simulated el niño stress. Mar. Biol. 2007, 152, 677–686. [Google Scholar] [CrossRef]
- Macaya, E.C.; Boltaña, S.; Hinojosa, I.A.; Macchiavello, J.E.; Valdivia, N.A.; Vásquez, N.R.; Buschmann, A.H.; Vásquez, J.A.; Alonso Vega, J.M.; Thiel, M. Presence of sporophylls in floating kelp rafts of Macrocystis spp. (phaeophyceae) along the chilean pacific coast. J. Phycol. 2005, 41, 913–922. [Google Scholar] [CrossRef]
- Hernández-Carmona, G.; Hughes, B.; Graham, M.H. Reproductive longevity of drifting kelp Macrocystis pyrifera (phaeophyceae) in Monterey bay, USA. J. Phycol. 2006, 42, 1199–1207. [Google Scholar] [CrossRef]
- Hay, C.H. Durvillaea (bory). In Biology of Economic Seaweeds; SPB Academic Publishing: The Hague, The Netherlands, 1994. [Google Scholar]
- Castañeda, V.; Reyes-Bonilla, H. A tropical assemblage of benthic macroalgae on rocky reefs in a temperate zone on the western Baja California peninsula, mexico. Bot. Mar. 2010, 53, 195–203. [Google Scholar]
- Marins, B.; Amado-Filho, G.; Barbarino, E.; Pereira-Filho, G.; Longo, L. Seasonal changes in population structure of the tropical deep-water kelp laminaria abyssalis. Phycol. Res. 2014, 62, 55–62. [Google Scholar] [CrossRef]
- Emiliani, C. Pleistocene temperatures. J. Geol. 1955, 63, 538–578. [Google Scholar] [CrossRef]
- Lüning, K. Seaweeds: Their Environment, Biogeography, and Ecophysiology; John Wiley & Sons: Hoboken, BJ, USA, 1990; p. 544. [Google Scholar]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Schiel, D.R.; Foster, M.S. The population biology of large brown seaweeds: Ecological consequences of multiphase life histories in dynamic coastal environments. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 343–372. [Google Scholar] [CrossRef]
- Fernández, C. The retreat of large brown seaweeds on the north coast of Spain: The case of Saccorhiza polyschides. Eur. J. Phycol. 2011, 46, 352–360. [Google Scholar] [CrossRef]
- Falvey, M.; Garreaud, R.D. Regional cooling in a warming world: Recent temperature trends in the southeast pacific and along the west coast of subtropical South America (1979–2006). J. Geophys. Res. Atmos. 2009, 114, D4. [Google Scholar] [CrossRef]
- Bolton, J.; Anderson, R.; Smit, A.; Rothman, M. South African kelp moving eastwards: The discovery of Ecklonia maxima (osbeck) papenfuss at de hoop nature reserve on the south coast of South Africa. Afr. J. Mar. Sci. 2012, 34, 147–151. [Google Scholar] [CrossRef]
- Verbruggen, H.; Tyberghein, L.; Pauly, K.; Vlaeminck, C.; Nieuwenhuyze, K.V.; Kooistra, W.H.C.F.; Leliaert, F.; Clerck, O.D. Macroecology meets macroevolution: Evolutionary niche dynamics in the seaweed halimeda. Glob. Ecol. Biogeogr. 2009, 18, 393–405. [Google Scholar] [CrossRef]
- Palumbi, S.R.; Gaines, S.D.; Leslie, H.; Warner, R.R. New wave: High-tech tools to help marine reserve research. Front. Ecol. Environ. 2003, 1, 73–79. [Google Scholar] [CrossRef]
- Anderson, A.; Bonaldo, R.; Barneche, D.; Hackradt, C.; Félix-Hackradt, F.; García-Chartón, J.; Floeter, S. Recovery of grouper assemblages indicates effectiveness of a marine protected area in southern brazil. Mar. Ecol. Prog. Ser. 2014, 514, 207–215. [Google Scholar] [CrossRef]
- Shefer, S.; Abelson, A.; Mokady, O.; Geffen, E. Red to mediterranean sea bioinvasion: Natural drift through the suez canal, or anthropogenic transport? Mol. Ecol. 2004, 13, 2333–2343. [Google Scholar] [CrossRef] [PubMed]
- Facon, B.; Genton, B.J.; Shykoff, J.; Jarne, P.; Estoup, A.; David, P. A general eco-evolutionary framework for understanding bioinvasions. Trends Ecol. Evol. 2006, 21, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A.B.; Carvalho-Filho, A.; Morais, R.A.; Nunes, L.T.; Quimbayo, J.P.; Floeter, S.R. Brazilian tropical fishes in their southern limit of distribution: Checklist of santa catarina’s rocky reef ichthyofauna, remarks and new records. J. Biodivers. Data 2015, 11, 1688. [Google Scholar] [CrossRef]
- Olson, B.; Richards, J. Spatial arrangement of tiller replacement in agropyron desertorum following grazing. Oecologia 1988, 76, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Peterson, R.G.; Stramma, L. Upper-level circulation in the South Atlantic Ocean. Prog. Oceanogr. 1991, 26, 1–73. [Google Scholar] [CrossRef]
- Pereira, M.D.; Schettini, C.A.F.; Omachi, C.Y. Caracterização de feições oceanográficas na plataforma de santa catarina através de imagens orbitais. Rev. Bras. Geofís. 2009, 27, 81–93. [Google Scholar] [CrossRef]
- Acha, E.M.; Mianzan, H.W.; Guerrero, R.A.; Favero, M.; Bava, J. Marine fronts at the continental shelves of austral south america. J. Mar. Syst. 2004, 44, 83–105. [Google Scholar] [CrossRef]
- Piola, A.R.; Campos, E.J.D.; Möller, O.O.; Charo, M.; Martinez, C. Subtropical shelf front off eastern South America. J. Geophys. Res. Oceans 2000, 105, 6565–6578. [Google Scholar] [CrossRef]
- Piola, A.R.; Matano, R.P.; Palma, E.D.; Möller, O.O.; Campos, E.J.D. The influence of the plata river discharge on the western south Atlantic shelf. Geophys. Res. Lett. 2005, 32. [Google Scholar] [CrossRef]
- Anderson, A.B. Peixes Tropicais no seu Limite de Distribuição: Dinâmica Temporal da Ictiofauna Recifal no sul do Brasil. Ph.D. Thesis, Universidade Federal de Santa Catarina, Florianópolis, Santa Catarina, Brazil, 2017; p. 202. [Google Scholar]
- Horta, P.; Amancio, E.; Coimbra, C.; Oliveira, E. Considerações sobre a distribuição e origem da flora de macroalgas marinhas brasileiras. Hoehnea 2001, 28, 243–265. [Google Scholar]
- Spalding, M.D.; Fox, H.E.; Allen, G.R.; Davidson, N.; FerdaÑA, Z.A.; Finlayson, M.A.X.; Halpern, B.S.; Jorge, M.A.; Lombana, A.L.; Lourie, S.A.; et al. Marine ecoregions of the world: A bioregionalization of coastal and shelf areas. BioScience 2007, 57, 573–583. [Google Scholar] [CrossRef]
- Pires Gouvêa, L.; Schubert, N.; Dalcuche Leal Martins, C.; Sissini, M.; Ramlov, F.; Regina de Oliveira Rodrigues, E.; Oliveira Bastos, E.; Carvalho Freire, V.; Maraschin, M.; Simonassi, J.; et al. Interactive effects of marine heatwaves and eutrophication on the ecophysiology of a widespread and ecologically important macroalga: Temperature and nutrient effect on macroalgae. Limnol. Oceanogr. 2017, 62, 2056–2075. [Google Scholar] [CrossRef]
- Freire, A.S.; Varela, A.R.D.; Fonseca, A.L.D.; Menezes, B.S.; Fest, C.B.; Obata, C.S.; Gorril, C.; Franco, D.; Machado, E.C.; Barros, G.; et al. O ambiente oceanográfico. In Maare—Monitoramento Ambiental da Reserva Biológica Marinha do Arvoredo e Entorno; UFSC-EdUFSC: Florianópolis, Brazil, 2017; Volume 1, pp. 159–197. [Google Scholar]
- Fraser, C.I.; Hay, C.H.; Spencer, H.G.; Waters, J.M. Genetic and morphological analyses of the southern bull kelp Durvillaea antarctica (phaeophyceae: Durvillaeales) in New Zealand reveal cryptic species. J. Phycol. 2009, 45, 436–443. [Google Scholar] [CrossRef] [PubMed]
- Leal, P.P.; Hurd, C.L.; Roleda, M.Y. Meiospores produced in sori of nonsporophyllous laminae of macrocystis pyrifera (laminariales, phaeophyceae) may enhance reproductive output. J. Phycol. 2014, 50, 400–405. [Google Scholar] [CrossRef] [PubMed]
- Elith, J.; Graham, C.; Anderson, R.; Dudık, M.; Ferrier, S. Novel methods improve prediction of species’ distribution models. Ecography 2006, 32, 66–77. [Google Scholar] [CrossRef]
- Philips, S.J.; Dudík, M. Modeling of species distributions with maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander, J.A. A practical guide to maxent for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Hernandez, P.A.; Graham, C.H.; Master, L.L.; Albert, D.L. The effect of sample size and species characteristics on performance of different species distribution modeling methods. Ecography 2006, 29, 773–785. [Google Scholar] [CrossRef]
- Assis, J.; Coelho, N.C.; Lamy, T.; Valero, M.; Alberto, F.; Serrão, E.Á. Deep reefs are climatic refugia for genetic diversity of marine forests. J. Biogeogr. 2016, 43, 833–844. [Google Scholar] [CrossRef]
- Assis, J.; Araújo, M.B.; Serrão, E.A. Projected climate changes threaten ancient refugia of kelp forests in the north Atlantic. Glob. Chang. Biol. 2017. [Google Scholar] [CrossRef] [PubMed]
- O’brien, R.M. A caution regarding rules of thumb for variance inflation factors. Qual. Quant. 2007, 41, 673–690. [Google Scholar] [CrossRef]
- Segurado, P.; Araújo, M.B.; Kunin, W. Consequences of spatial autocorrelation for niche-based models. J. Appl. Ecol. 2006, 43, 433–444. [Google Scholar] [CrossRef]
- Patrão, C.; Assis, J.; Rufino, M.; Silva, G.; Jordaens, K.; Backeljau, T.; Castilho, R. Habitat suitability modelling of four terrestrial slug species in the iberian peninsula (arionidae: Geomalacus species). J. Molluscan Stud. 2015, 81, 427–434. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M.; Elith, J.; Graham, C.H.; Lehmann, A.; Leathwick, J.; Ferrier, S. Sample selection bias and presence-only distribution models: Implications for background and pseudo-absence data. Ecol. Appl. 2009, 19, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Peterson, A.T. Ecological niche conservatism: A time-structured review of evidence. J. Biogeogr. 2011, 38, 817–827. [Google Scholar] [CrossRef]
- Muscarella, R.; Galante, P.J.; Soley-Guardia, M.; Boria, R.A.; Kass, J.M.; Uriarte, M.; Anderson, R.P. Enmeval: An r package for conducting spatially independent evaluations and estimating optimal model complexity for maxent ecological niche models. Methods Ecol. Evol. 2014, 5, 1198–1205. [Google Scholar] [CrossRef]
- Madon, B.; Warton, D.I.; Araújo, M.B. Community-level vs. species-specific approaches to model selection. Ecography 2013, 36, 1291–1298. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Fraser, C.I.; Winter, D.J.; Spencer, H.G.; Waters, J.M. Multigene phylogeny of the southern bull-kelp genus durvillaea (phaeophyceae: Fucales). Mol. Phylogenet. Evol. 2010, 57, 1301–1311. [Google Scholar] [CrossRef] [PubMed]
- Tala, F.; Velásquez, M.; Mansilla, A.; Macaya, E.C.; Thiel, M. Latitudinal and seasonal effects on short-term acclimation of floating kelp species from the south-east pacific. J. Exp. Mar. Biol. Ecol. 2016, 483, 31–41. [Google Scholar] [CrossRef]
- Coyer, J.A.; Smith, G.J.; Andersen, R.A. Evolution of Macrocystis spp. (phaeophyceae) as determined by its1 and its2 sequences1. J. Phycol. 2001, 37, 574–585. [Google Scholar] [CrossRef]
- Van den Hoek, C.; Donze, M. Algal phytogeography of the European Atlantic coasts. Blumea 1967, 15, 63–85. [Google Scholar]
- Smith, S.D. Kelp rafts in the southern ocean. Glob. Ecol. Biogeogr. 2002, 11, 67–69. [Google Scholar] [CrossRef]
- Garden, C.J.; Currie, K.; Fraser, C.I.; Waters, J.M. Rafting dispersal constrained by an oceanographic boundary. Mar. Ecol. Prog. Ser. 2014, 501, 297–302. [Google Scholar] [CrossRef]
- Edwards, M.S.; Kim, K.Y. Diurnal variation in relative photosynthetic performance in giant kelp Macrocystis pyrifera (phaeophyceae, laminariales) at different depths as estimated using pam fluorometry. Aquat. Bot. 2010, 92, 119–128. [Google Scholar] [CrossRef]
- Tokinaga, H.; Tanimoto, Y.; Xie, S.-P. Sst-induced surface wind variations over the brazil–malvinas confluence: Satellite and in situ observations. J. Clim. 2005, 18, 3470–3482. [Google Scholar] [CrossRef]
- Cataldi, M.; Caro, L.D.; Schifanella, C. Emerging topic detection on twitter based on temporal and social terms evaluation. In Proceedings of the Tenth International Workshop on Multimedia Data Mining, Washington, DC, USA, 25 July 2010; pp. 1–10. [Google Scholar]
- De Souza, R.B.; Mata, M.M.; Garcia, C.A.; Kampel, M.; Oliveira, E.N.; Lorenzzetti, J.A. Multi-sensor satellite and in situ measurements of a warm core ocean eddy south of the brazil-malvinas confluence region. Remote Sens. Environ. 2006, 100, 52–66. [Google Scholar] [CrossRef]
- Rothäusler, E.; Gómez, I.; Karsten, U.; Tala, F.; Thiel, M. Physiological acclimation of floating macrocystis pyrifera to temperature and irradiance ensures long-term persistence at the sea surface at mid-latitudes. J. Exp. Mar. Biol. Ecol. 2011, 405, 33–41. [Google Scholar] [CrossRef]
- Clendenning, K. Organic productivity in kelp areas. Nova Hedwigia 1971, 32, 259–263. [Google Scholar]
- Colombo-Pallotta, M.F.; García-Mendoza, E.; Ladah, L.B. Photosynthetic performance, light absorption, and pigment composition of Macrocystis pyrifera (laminariales, phaeophyceae) blades from different depths. J. Phycol. 2006, 42, 1225–1234. [Google Scholar] [CrossRef]
- Cheung, W.W.L.; Lam, V.W.Y.; Sarmiento, J.L.; Kearney, K.; Watson, R.; Pauly, D. Projecting global marine biodiversity impacts under climate change scenarios. Fish Fish. 2009, 10, 235–251. [Google Scholar] [CrossRef]
- Reed, D.; Washburn, L.; Rassweiler, A.; Miller, R.; Bell, T.; Harrer, S. Extreme warming challenges sentinel status of kelp forests as indicators of climate change. Nat. Commun. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Ladah Lydia, B.; Zertuche-González José, A. Giant kelp (Macrocystis pyrifera) survival in deep water (25–40 m) during el niño of 1997–1998 in baja california, mexico. Bot. Mar. 2004, 47, 367–372. [Google Scholar] [CrossRef]
- Pehlke, C.; Bartsch, I. Changes in depth distribution and biomass of sublittoral seaweeds at Helgoland (North Sea) between 1970 and 2005. Clim. Res. 2008, 37, 135–147. [Google Scholar] [CrossRef]
- Yoneshigue-Valentin, Y. In the Life Cycle of Laminaria abyssalis (Laminariales, phaeophyta) in Culture. In Proceedings of the Thirteenth International Seaweed Symposium, Vancouver, BC, Canada, 13–18 August 1989; Springer: Berlin, Germany, 1990; pp. 461–466. [Google Scholar]
- Yoneshigue-Valentin, Y.; Mitchell, G.J.; Gurgel, C.F.D. Quelques observations préliminaires sur les macroalgues de la plateforme continentale du sudest brésilien. Acta Bot. Gall. 1995, 142, 161–165. [Google Scholar] [CrossRef]
- O’Hara, T.D.; Consalvey, M.; Lavrado, H.P.; Stocks, K.I. Environmental predictors and turnover of biota along a seamount chain. Mar. Ecol. 2010, 31, 84–94. [Google Scholar] [CrossRef]
- Williams, S.L.; Smith, J.E. A global review of the distribution, taxonomy, and impacts of introduced seaweeds. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 327–359. [Google Scholar] [CrossRef]
- Azevedo, C.A.A.D.; Cassano, V.; Júnior, P.A.H.; Batista, M.B.; Oliveira, M.C.D. Detecting the non-native Grateloupia turuturu (halymeniales, rhodophyta) in southern Brazil. Phycologia 2015, 54, 451–454. [Google Scholar] [CrossRef]
- Edwards, M.; Hernandez-Carmona, G. Delayed recovery of giant kelp near its southern range limit in the north pacific following el niño. Mar. Biol. 2005, 147, 273–279. [Google Scholar] [CrossRef]
- Nyberg, C.D.; Wallentinus, I. Can species traits be used to predict marine macroalgal introductions? Biol. Invasions 2005, 7, 265–279. [Google Scholar] [CrossRef]
- South, P.M.; Lilley, S.A.; Tait, L.W.; Alestra, T.; Hickford, M.J.H.; Thomsen, M.S.; Schiel, D.R. Transient effects of an invasive kelp on the community structure and primary productivity of an intertidal assemblage. Mar. Freshw. Res. 2015, 67, 103–112. [Google Scholar] [CrossRef]
- Thompson, G.A.; Schiel, D.R. Resistance and facilitation by native algal communities in the invasion success of Undaria pinnatifida. Mar. Ecol. Prog. Ser. 2012, 468, 95–105. [Google Scholar] [CrossRef]
- Valentine, J.P.; Johnson, C.R. Establishment of the introduced kelp Undaria pinnatifida in Tasmania depends on disturbance to native algal assemblages. J. Exp. Mar. Biol. Ecol. 2003, 295, 63–90. [Google Scholar] [CrossRef]
- Bernhardt, J.R.; Leslie, H.M. Resilience to climate change in coastal marine ecosystems. Annu. Rev. Mar. Sci. 2013, 5, 371–392. [Google Scholar] [CrossRef] [PubMed]
- Challinor, A.J.; Adger, W.N.; Benton, T.G. Climate risks across borders and scales. Nat. Clim. Chang. 2017, 7, 621–623. [Google Scholar] [CrossRef]
- Barry, J.P.; Baxter, C.H.; Sagarin, R.D.; Gilman, S.E. Climate-related, long-term faunal changes in a California rocky intertidal community. Science 1995, 267, 672–675. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, W.N. Effect of boundary layer transport on the fixation of carbon by the giant kelp Macrocystis pyrifera. Mar. Biol. 1980, 56, 103–110. [Google Scholar] [CrossRef]


| Samples | Alpha | EK | rETRmax | Fv/Fm |
|---|---|---|---|---|
| Rafts | 0.183–0.302 | 212.54–304.00 | 50.16–88.07 | 0.559–0.661 |
| Chilean population/January | - | - | - | 0.584–0.611 |
| Chilean population/July | - | - | - | 0.711–0.722 |
| Chilean population/August | - | - | - | 0.656–0.681 |
| American Population * | 0.3–0.926 | 34.4–178.5 | 14.7–147.4 | 0.4–0.75 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bernardes Batista, M.; Batista Anderson, A.; Franzan Sanches, P.; Simionatto Polito, P.; Lima Silveira, T.C.; Velez-Rubio, G.M.; Scarabino, F.; Camacho, O.; Schmitz, C.; Martinez, A.; et al. Kelps’ Long-Distance Dispersal: Role of Ecological/Oceanographic Processes and Implications to Marine Forest Conservation. Diversity 2018, 10, 11. https://doi.org/10.3390/d10010011
Bernardes Batista M, Batista Anderson A, Franzan Sanches P, Simionatto Polito P, Lima Silveira TC, Velez-Rubio GM, Scarabino F, Camacho O, Schmitz C, Martinez A, et al. Kelps’ Long-Distance Dispersal: Role of Ecological/Oceanographic Processes and Implications to Marine Forest Conservation. Diversity. 2018; 10(1):11. https://doi.org/10.3390/d10010011
Chicago/Turabian StyleBernardes Batista, Manuela, Antônio Batista Anderson, Paola Franzan Sanches, Paulo Simionatto Polito, Thiago Cesar Lima Silveira, Gabriela M. Velez-Rubio, Fabrizio Scarabino, Olga Camacho, Caroline Schmitz, Ana Martinez, and et al. 2018. "Kelps’ Long-Distance Dispersal: Role of Ecological/Oceanographic Processes and Implications to Marine Forest Conservation" Diversity 10, no. 1: 11. https://doi.org/10.3390/d10010011