1. Introduction
Parasitic plants are a taxonomically diverse group of angiosperms that are widely described in ethno-pharmacology but remain largely unknown by modern science. Parasitic plants are mainly nonphotosynthetic, completely depending on host plant for nutrients [
1]. Thus, profound interactions with the ecosystem are usually observed. Climate has major impacts, especially on the pattern of secondary metabolites, which is extremely dependent on the weather conditions. In fact, reactive oxygen species (ROS) are more rapidly produced, and their products more prone to accumulate in plants subjected to arid climates. As a consequence, plants have developed several mechanisms to cope with drought and to counteract oxidative damage [
2], including the production of enzymatic and non-enzymatic antioxidant systems [
3]. Thus, the metabolic patterns of plants that belong to different bioclimate areas could accordingly differ.
Cynomorium coccineum is an edible, non-photosynthetic parasitic plant that is widely used in traditional medicine [
4]. It occurs in a large area along the Mediterranean Basin, including Spain, southern Italy, Malta, Corse, Crete, northern Africa, and the Arabian Peninsula [
5], where it is known by different names such as “Tarthuth” in Arabia, “Champignon de Malte” in Corsica and “Cagalloni strantaxiu” in Sardinia [
6]. This plant is a holo-parasite associated with arid coastal climates and saline soils, and it is characterized by a highly distinguishable form. In fact, its color (brown-reddish for the absence of photosynthetic pigments) and its prolate form are responsible for one of its common names: “Maltese mushroom” (although it is not a mushroom at all). Depending on the bioclimate,
C. coccineum can grow on different halophytic plants. For instance, in the Tataouine region (southeast Tunisia), which is characterized by a harsh continental climate due to its large distance from the Mediterranean Sea and the pronounced influence of the Sahara Desert, the most stable plant landscape consists of perennial plants that are drought-resistant, minimizing transpiration between periods when the soil water reserves enable growth. Here,
C. coccineum is found on plants that are characteristic of this plant cover, including
Haloxylon salicornicum,
Salsola vermiculata, and
Retama raetam. This is in contrast to the Mediterranean Basin where the bioclimate is less arid and is characterized by rainy winters and warm/dry summers,
C. coccineum is found to grow on different halophytic plants, particularly
Atriplex halimus,
Obione portulacoides, and
Inula crithmoides [
7].
The medicinal properties of
C. coccineum have been held in high regard from ancient times to the present day. In Arabian medical practice, it is considered the “treasure of drugs”. Recent studies have evaluated the antioxidant potential and nutraceutical potential of
C. coccineum in Algeria [
8] and Sardinia (Italy) [
5,
6,
9,
10]. Rached and his coworkers determined the antioxidant activity of specimens of
C. coccineum grown in Algeria using scavenge of the DPPH radical and β-carotene bleaching [
8]. A similar study was performed in specimens of
C. songaricum, an Asian species of the genus
Cynomorium, widely used in traditional Chinese medicine [
11,
12,
13]. Previous studies have showed that
Cynomorium plants contain condensed tannins, phenolic acids, steroids, anthocyanins, triterpenes, acidic heteropolysaccharides, butyl fructosides, flavanoids, lignan glycosides, alkaloids, and other compounds [
5,
13,
14].
Recently, the chemical composition and biological activity of the fixed oil isolated from Sardinian
C. coccineum were also studied [
4,
15]. The oil was obtained by supercritical fluid extraction with carbon dioxide (SFE-CO
2), a more sustainable separation technique that constitutes an industrial alternative to classical solvent extraction processes. The resulting oil contained mainly triacylglycerols and their derivates. The main fatty acids were oleic (18:1
n-9), linoleic (18:2
n-6), palmitic (16:0), and α-linolenic (18:3
n-3) acid.
The aim of this work was to compare the total phenolic and anthocyanin content and antioxidant activity of different extracts (in n-hexane, chloroform, ethyl acetate, acetone, methanol, and water) of C. coccineum from two different geographical areas: Tataouine (southeast Tunisia) and Sardinia (south Italy). The Tunisian C. coccineum is available only for a very small period of the year in Tunisia due to the arid climate, and to the best of our knowledge, it has thus far been very poorly studied. In contrast, in other regions of the Mediterranean basin with a cooler and more humid climate, such as Sardinia, the plant is available in greater quantities, but limited to few weeks during the spring. The composition of the water and methanol extracts and of the fixed oils obtained by CO2-SFE extraction were also characterized by 1H NMR spectroscopy and chromatographic techniques, respectively. The results are discussed in terms of the effect of environmental stress on the chemical composition and nutraceutical potential of C. coccineum.
2. Materials and Methods
2.1. Chemicals
Deuterium oxide (D2O, 99.9%), sodium 3-trimethylsilyl-propionate-2,2,3,3-d4 (TSP), fatty acid and fatty acid methyl ester standards, and Desferal (deferoxaminemesylate salt) were acquired from Sigma–Aldrich (Milan, Italy). Cyanidin 3-O-glucoside was obtained from Extrasynthese (Genay, France). Ultrapure water (18 mΩ) was obtained with a Milli-Q Advantage A10 System apparatus (Millipore, Milan, Italy). All the other reagents, standards and solvents were of HPLC grade and were purchased from Sigma-Aldrich and used without further purification.
2.2. Plant Materials
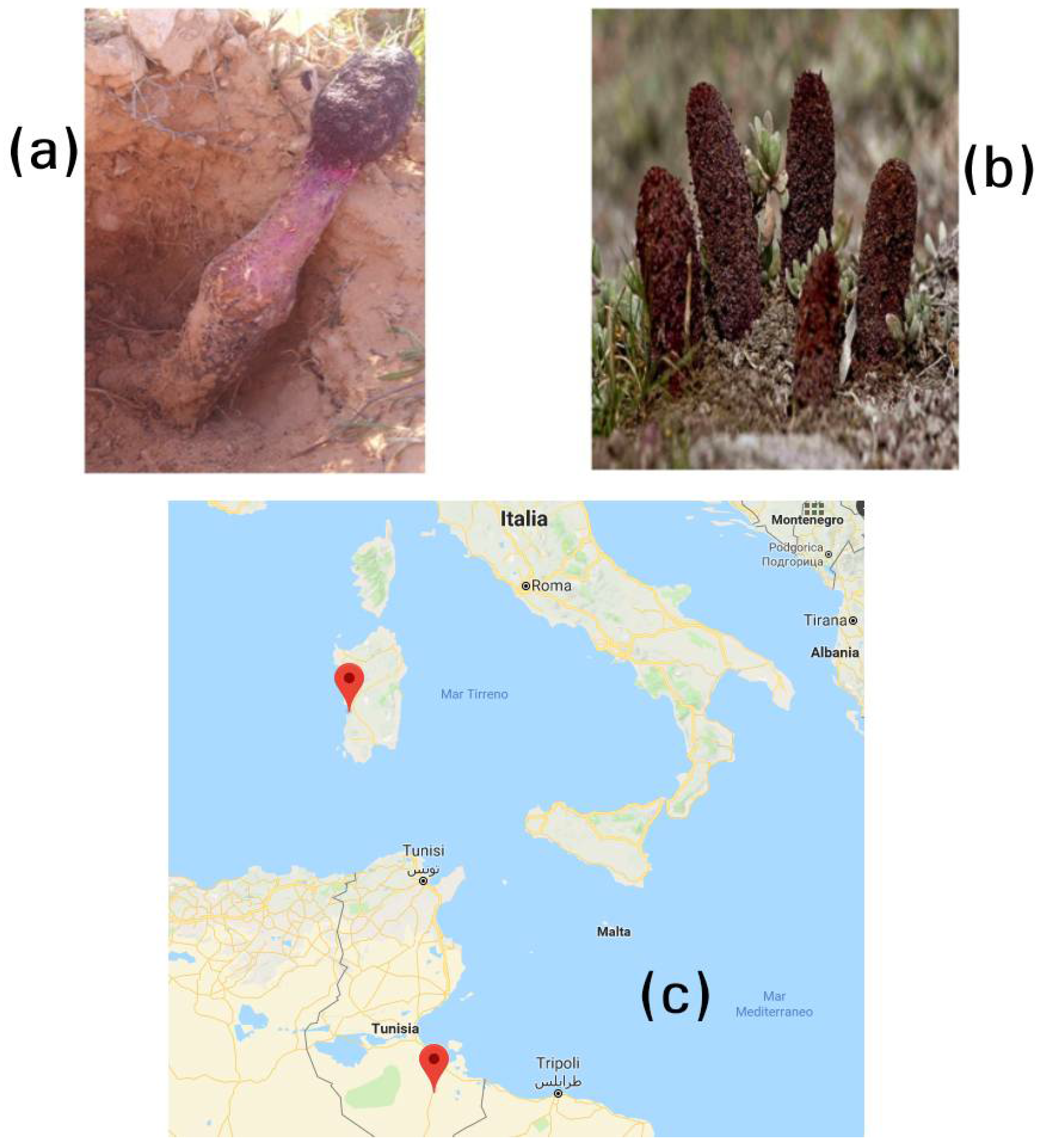
C. coccineum was collected from Tataouine, in southeast Tunisia, in March 2016 and from Arborea in southwestern Sardinia (Italy) in April 2016 (
Figure 1), during the flowering period (that usually lasts only few days depending on seasonal climate fluctuations). The collections of the Department of Life Science, University of Sfax and the Department of Biomedical Sciences, University of Cagliari were used to store reference materials of the both species. The specimens were kept at about 4 °C in a portable refrigerator (using dry ice to further control the temperature), and brought to the laboratory within one hour of harvest. About 4 kg (containing approximately 40 different samples) per collection, equally distributed in an area of about 4 ha, were collected to obtain a representative sample. The aerial part of the plants was then gently cleaned and the residual earth removed. The specimens were cut into slices approximately 0.5 cm thick and hung for freeze-drying, using a laboratory apparatus (6 kg/24 h) equipped with glass bell-jars and vacuum pump.
2.3. Preparation of Extracts
2.3.1. Supercritical Fluid Extraction
Supercritical CO
2 (purity 99%-Air Liquide Italia, Cagliari, Italy) extraction (SFE) was performed in a laboratory apparatus equipped with a 320 mL extraction vessel (
Figure 2). Extractions were carried out in a semibatch mode with batch charging of the vegetable matter and a continuous flow of solvent, adopting an experimental arrangement that omits the first separator. Approximately 250 g of plant sample was charged in each run at 250 bar and 40 °C in the extraction section and 20 bar and 15 °C in the second separator.
2.3.2. Maceration
Freeze-dried samples were extracted by maceration, using different solvents with increasing polarity. Briefly, 25 g of powder was treated with 150 mL of each solvent, viz.,
n-hexane, chloroform, ethyl acetate, acetone, methanol, and water, serially (
Figure 3). After 24 h at 25 °C in the dark, the supernatant was recovered by filtration. The process was repeated, and the solvent in the supernatant was evaporated in a vacuum rotary evaporator, to partially separate the bioactive compounds in the extracts.
2.4. Determination of Total Phenolics
Total phenolics were determined using Folin-Ciocalteu reagent, in accordance with previous studies [
16]. Gallic acid was used for the calibration (linearity range 0.05–0.6 mM), and the results were calculated as gallic acid equivalents per gram of dry material (mmol GAE/g).
2.5. Determination of Total Anthocyanins
Total anthocyanin determination was performed as already described [
6]. Differential pH absorbance at 510 nm and 700 nm were measured using 0.2 M HCl/KCl pH 1.0 and 1 M sodium acetate buffers pH 4.5. The final absorbance was calculated using Equation (1).
The total anthocyanin concentration was then obtained using molar extinction coefficient of cyanidin 3-O-glucoside (C3G) (29,300 M−1·cm−1) and Lambert–Beer equation.
2.6. DPPH (1,1-Diphenyl-2-picrylhydrazyl) Radical Scavenging Assay
The DPPH scavenging assay was performed using a well-known spectrophotometric assay, involving the dark purple DPPH radical [
17,
18]. Trolox was used as the standard (linearity range 5–50 μM), and the results were calculated as Trolox equivalents per gram of dry material (mmol TE/g).
2.7. Ferric Reducing Antioxidant Power (FRAP)
FRAP was determined using a spectrophotometric method [
18], using both Trolox equivalents per gram of dry material (mmol TE/g) and mmol of Fe(II) per gram of dry material (mmol Fe
II/g) as the reference for the calibration curves.
2.8. Trolox Equivalent Antioxidant Capacity (TEAC) Assay ABTS
The TEAC spectrophotometric assay was performed using the stable ABTS radical as the probe [
19]. The radical has been diluted with potassium phosphate buffet 75 mM pH 7.4, then 1 mL of radical was incubated for 6′ with 10 μL sample. Decrease in absorbance at 734 nm was finally detected. Trolox was used as the standard and the results were expressed as Trolox equivalents per gram of dry material (mmol TE/g).
2.9. ORAC-PYR (Oxygen Radical Absorbance Capacity-Pyrogallol Red) Determination
The hydrogen atom transfer (HAT) method ORAC-PYR was also used, in the presence of pyrogallol red as the probe, and 2,2′-azobis(2-amidinopropane) dihydrochloride (AAPH) as the hydroxyl radical releaser [
18,
20]. Trolox was used for the calibration curve, and the results were expressed as Trolox Equivalents per gram of dry material (mmol TE/g).
2.10. NMR Measurements
Aqueous and methanolic extracts were dried under vacuum. A 10 mg aliquot of each dried extracts was resuspended in 600 μL of D2O with the internal standard (TSP) added at a known concentration (1 mM) and vortexed. Then, 650 μL of the reconstituted solution was transferred to a 5 mm (o.d.) NMR tube.
1H NMR analysis were conducted on a Varian UNITY INOVA 500 spectrometer operating at 499.84 MHz. Spectra were acquired at 300 K from 256 scans collected over a spectral width of 6000 Hz. A 45° pulse, an acquisition time of 2.3 s and a relaxation delay of 4 s were used. Suppression of the water peak was achieved by applying a pre-saturation technique with low power radiofrequency irradiation for 1.5 s. A line broadening of 0.3 Hz was applied to all free induction decays (FIDs) prior to Fourier transformation. Spectra were manually phase- and baseline-corrected using MestReNova software (version 8.1.2, Mestrelab Research S.L., Santiago de Compostela, Spain) and 1H NMR chemical shifts referencing on TSP at δ 0.00 ppm was performed.
2D 1H−1H correlation spectroscopy (COSY) and 1H−1H total correlation spectroscopy (TOCSY) experiments were performed with a spectral width of 6000 Hz in both dimensions, an acquisition time of 0.171 s, a delay time of 1.0 s, number of scans of 128 and number of data points of 2048 (f2) and 256 (f1). TOCSY experiments were performed in phase-sensitive mode. 1H−13C heteronuclear single-quantum correlation (HSQC) and heteronuclear multiple-bond correlation (HMBC) spectra were acquired using spectral widths of 28,901 (f1) and 6000 Hz (f2), an acquisition time of 0.171 s, a delay time of 1.0 s, dummy scans of 32, 2048 data points in f2 and 256 increments in f1. The molecular composition of extracts, expressed as mol %, was estimated by measuring the area under the NMR peaks assigned to specific functional groups of molecules and normalizing integrals to the sum of total spectra area to compensate for possible differences in the extraction yields.
2.11. Fixed Oil Saponification and Fatty Acids Analysis
2.11.1. Oil Saponification
C. coccineum fixed oils (3 mg, in EtOH solution) were subjected to mild saponification as previously described [
4]. Dried saponifiable fractions, with free fatty acids, were injected into the HPLC system [
4]. A portion of dried fatty acids after saponification was methylated and fatty acid methyl esters were analyzed by GC-FID as previously reported [
4].
2.11.2. HPLC-DAD Analysis
According to the literature, the quali-quantitative composition of unsaturated fatty acids was obtained with a 1100 HPLC-DAD system (Agilent Technologies, Palo Alto, CA, USA) [
4]. UFAs, detected at 200 nm, were eluted with CH
3CN/H
2O/CH
3COOH (75/25/0.12,
v/
v/
v) as mobile phase at a flow rate of 2.3 mL/min, using an Agilent Technologies XDB-C
18 Eclipse (150 × 4.6 mm, 3.5 µm particle size) column (at 37 °C), equipped with a Zorbax XDB-C
18 Eclipse (12.5 × 4.6 mm, 5 µm particle size) guard column. Fatty acids were identified using standard compounds (linear calibration curves, correlation coefficients > 0.995) and the conventional UV spectra; the recording and integration of the chromatogram data were carried out with an Agilent OpenLAB Chromatography data system [
4].
2.11.3. GC-FID Analysis
HPLC data were combined and confirmed by GC-FID, to obtain an exhaustive characterization of the fatty acid profile of
C. coccineum oils. Fatty acid methyl esters were measured using a HP-6890 GC-FID, as previously reported [
4]. Fatty acid methyl esters were identified by comparing the retention times to those of standard compounds and quantified as a percentage of the total amount of fatty acids (g %).
2.12. HPLC Analyses of Free Fatty Acids in the Oil
Aliquots of
C. coccineum fixed oils, dissolved in CH
3CN/0.14% CH
3COOH (
v/
v) (2 mg/mL solutions), were injected into the HPLC-DAD system and the quali-quantitative composition of free unsaturated fatty acids (FUFA) was obtained as described in the
Section 2.11.2. [
4].
2.13. Statistical Analysis
GraFit 7 (Erithacus Software, London, UK), R 2.5.1 (R Foundation for Statistical Computing, Vienna, Austria), and GraphPad INSTAT software (GraphPad software, San Diego, CA, USA) were used for data analysis. The one-way analysis of variance (ANOVA) and the Bonferroni multiple comparisons test were used to evaluate the statistical significance of the differences.
3. Results and Discussion
3.1. Total Antioxidant Power and Phenolics Content
Five different extracts of
C. coccineum with increasing polarity were evaluated for their contents of phenolics and anthocyanins, using the residual material for subsequent extractions (
Figure 3), to partially separate the bioactive compounds, and allow a clearer comparison between samples. The total antioxidant capacity was also determined using both electron transfer (ET) methods (i.e., the DPPH assay, the TEAC-ABTS-based assay, and the FRAP method) and the hydrogen atom transfer (HAT) method (ORAC-PYR). The full results are summarized in
Table 1 and
Table 2.
Antioxidant activity was remarkable in all the tested extracts. There were differences depending on the solvent and the origin of the plant. The highest antioxidant capacity values were obtained with the HAT-based method, the ORAC-PYR assay, as expected. TEAC, FRAP, and DPPH (electron-transfer based methods) still showed significant activity, but the results confirmed that they could underestimate total antioxidant capacity, as reported in previous studies [
18,
21].
Since phenolics are the main component responsible for antioxidant activity [
22,
23,
24], this fraction was spectrophotometrically determined using the Folin–Ciocalteu assay. Anthocyanins (a subclass of phenolics) were also determined.
Phenolic compounds are polar compounds. In fact, the three more polar extracts (acetone, methanol and water) are richer than the less polar extracts. Thus, the extraction solvent affects the content of polyphenols in the extracts. Indeed, acetone, water and methanol, which are the most polar solvents, can extract large quantities of phenolic compounds [
25].
Anthocyanins are colored compounds (i.e., orange, purple or blue) that are widely involved in the coloration of petals but are also found in many other tissues. Their synthesis in the foliar organs is often activated by stress (cold, deficiencies, senescence).
Cyanidin-3-
O-glucoside (a widespread anthocyanin) has been identified as the main component of the external layer of
C. coccineum [
5,
6].
The analysis of these results shows that no significant difference was observed in the aqueous extracts. Meanwhile, the Tunisian acetone extracts were richer in phenolics (p < 0.05), whereas the Sardinian methanolic extracts were richer (p < 0.05), suggesting highly complex and different chemical compositions. On the whole, Sardinian Cynomorium is richer in polyphenols (approximately 30% more, p < 0.05).
Statistical analysis of the total anthocyanins content determined shows very significant differences in the various solvents used. Indeed, the Tunisian acetone extract and the Sardinian ethyl acetate extract of
C.
coccineum are the richest in anthocyanins, while only trace amounts are present in the hexane extracts from the two provenances. In the Sardinian samples, anthocyanins seem to be almost equally distributed in the ethyl acetate, acetone, methanol, and water extracts, whereas in the Tunisian samples, the majority of anthocyanins were extracted by acetone. On the whole, Tunisian
Cynomorium is richer in anthocyanins (
p < 0.05), confirming the role of these chemicals in the prevention of drought stress [
2]. In addition, the different extraction patterns suggest a different chemical composition in this phenolic sub-class.
Table 1 and
Table 2 show that hexane extracts exhibit very low antioxidant activity. In contrast, the four more polar extracts present significant power, as confirmed by both the ET and HAT assays.
The highest activity was observed in the acetone extracts, which were the richest in polyphenols, suggesting their important contribution to radical scavenging activity.
However, no clear pattern was observed between the two samples, which showed very similar biological activity overall in this context. For example, the Sardinian aqueous extract presented the highest values in the ORAC-PYR and DPPH assays, whereas TEAC-ABTS and FRAP assays found Tunisian aqueous samples to be the richest.
Overall, antioxidant activity seems to be quite similar in the two samples, despite a quite different phenolic pattern.
3.2. 1H NMR Spectroscopy
The chemical profiles of the aqueous and methanol extracts from Tunisian and Sardinian
C. coccineum were characterized by
1H NMR spectroscopy.
Figure 4 shows representative
1H NMR spectra of the samples under investigation. Spectral resonances of metabolites were assigned by combining 1D NMR spectra with data in the literature [
26], the available online Human Metabolome databases (HMDB) (
http://www.hmdb.ca), and the 500-MHz library from Chenomx NMR suite version 7.5 (Evaluation edition, Chenomx Inc., Edmonton, AB, Canada). Finally, 2D NMR experiments were also performed (
Supplementary Materials Figure S1).
The high-field region of the spectra (0–3.0 ppm) revealed the presence of amino acids (alanine, asparagine, glutamine, proline, threonine, and valine) and organic acids (acetate, citrate, formate, fumarate, malate, malonate, and succinate). Most of these metabolites identified in the 1D NMR spectrum of the extracts were confirmed by the cross-peaks in the 2D NMR spectra (
Supporting Information Figure S1). For instance, the presence of alanine was confirmed by
1H–
1H COSY cross peak between a doublet at δ
H 1.46 and a signal at δ
H 3.80. Similar correlations for the signals of asparagine at δ
H 2.87 and 2.97 with δ
H 4.02 were observed in the COSY spectrum. Acetic acid, formic acid, and succinate were assigned by the
13C chemical shifts obtained from the HSQC spectra (
Supplementary Materials Figure S1B). The middle-field spectral region of both extracts (3.0–5.5 ppm) was mainly dominated by signals from carbohydrates. Here, the anomeric peaks permitted the identification of β-glucose, α-glucose, fructose, and sucrose. Furthermore, betaine was also identified in this region, the singlets at δ
H 3.27 and 3.91 being heterocorrelated to the carbons at δ
C 56.21 and 68.92 in the HSQC experiment, respectively (
Supporting Information Figure S1B). The low-field region of the spectra (5.5–9 ppm) exhibited the signals with the weakest intensities, arising mainly from phenolic compounds.
A semi-quantitative evaluation of the compositional differences between sample extracts was performed by considering the easily integrated peaks. The results are presented in mol % in
Table 3. As shown, the relative molar content of carbohydrates was higher in both Sardinian sample extracts than in the Tunisian ones. In addition, fructose was the most abundant sugar in the Tunisian extracts, while sucrose predominated in the Sardinian ones. Compared to
C. coccineum growing in Sardinia, the Tunisian specimens had higher relative levels of total amino acids. Among the identified amino acids, asparagine was the most abundant in the Tunisian extracts, while alanine was predominant in the Sardinian samples. Seven organic acids were identified, and the major one in all samples was malic acid. Finally, the betaine content in samples from Tataouine was significantly higher than that in specimens from Sardinia (
p < 0.001).
Most of the metabolites detected here participate in various generic biochemical pathways (notably glycolysis, the tricarboxylic acid cycle, and amino acid metabolism) and some of them are known to accumulate in plants in response to abiotic stress. For instance, betaine is an organic osmolyte responsible for osmoregulation and osmoprotection, generally accumulated in plants to counteract environmental stresses such as salinity and extreme temperature [
27]. Soluble sugars play a dual role in plants, not only as metabolic resources and structural constituents of cells but also as signals regulating various processes associated with plant growth and development. Thus, given the different bioclimates that characterize the two geographical areas under investigation, it is likely that part of the diversity in the levels of these compounds between Tunisian and Sardinian extracts is due to a different metabolic adaptation of
C. coccineum to abiotic stress, stronger in the dry and hot bioclimate of Tunisia than the subtropical Mediterranean bioclimate of Sardinia.
3.3. Fatty Acids Profile of C. coccineum Fixed Oils
Quali-quantitative information on the individual fatty acids (FAs) that compose
C. coccineum fixed oils was obtained by GC-FID and HPLC-DAD analyses.
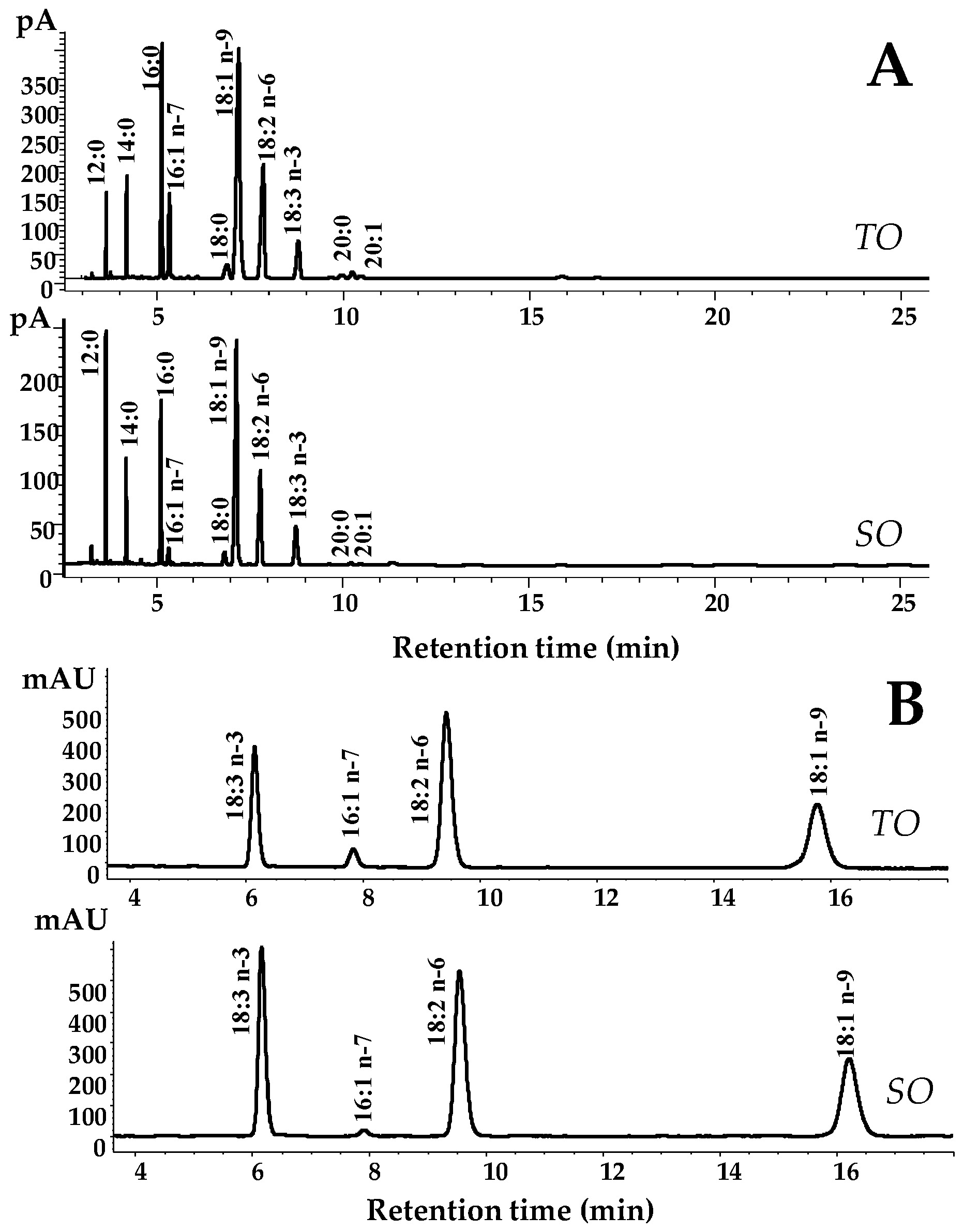
Figure 5 shows the fatty acid chromatographic profiles of Tunisian (TO) and Sardinian (SO) SFE oils obtained by GC-FID (
Figure 5A) and HPLC-DAD (
Figure 5B) analyses.
Table 4 shows the FAs composition (expressed as % of total FAs, g/100 g) of the fixed oils obtained by GC-FID analysis. The TO showed a concentration of approximately 23% saturated FAs (SFAs) (mainly lauric acid 14:0, palmitic acid 16:0, and stearic acid 18:0, 3, 15 and 3%, respectively), 48% monounsaturated FAs (MUFAs) (mainly oleic acid 18:1
n-9 and palmitoleic acid16:1
n-7, 40 and 5%), and 24% of polyunsaturated FAs (PUFAs), mainly constituted by the essential FAs linoleic acid (18:2
n-6) and α-linolenic acid (18:3
n-3), which represented 18 and 6% of the FAs content, respectively.
The absolute content of main unsaturated FAs (UFAs) (
Table 5) was detected by HPLC as follows: 459.7 mg/g, 205.7 mg/g and 76.07 mg/g of 18:1
n-9, 18:2
n-6, and 18:3
n-3, respectively. The TO showed a high content of free UFAs (FUFAs), and values of the free forms of all the UFAs > 60% were measured by HPLC. The SO was characterized by a similar FAs profile, with a high level of MUFAs (41% of 18:1
n-9), followed by SFA (31%) and PUFAs (24%). SO showed a significant higher level of SFA (
p < 0.05) and lower amount of MUFAs (
p < 0.001) than TO. In particular, differences were observed in the levels of 12:0 (9% and 2% for SO and TO, respectively;
p < 0.01) and 16:1
n-7 (1% and 5% for SO and TO, respectively;
p < 0.001). Significant higher absolute values of 16:1
n-7 (
p < 0.001), 18:1
n-9 (
p < 0.05), and 18:3
n-3 (
p < 0.001), were detected by HPLC analysis in TO (
Table 5). In addition, SO exhibited a significant lower amount of FUFAs (15%,
p < 0.001) than TO.
The FAs composition of TO and SO oils was similar to that of
C. coccineum fixed oil obtained from a Sardinian sample in a previous study [
4]. In particular, the major FAs were MUFAs (for the most part, 38% 18:1
n-9 and 2% 16:1
n-7), followed by 31% PUFAs and 27% SFAs (mainly 16:0). The previously studied oil showed a lower FUFAs content than TO but twice the quantity in SO.
Both SO and TO, like olive oil, palm oil, and sunflower oil [
4], contained mostly oleic acid (39–40%) but also exhibited a significantly increased content of the essential FAs 18:2
n-2 and 18:3
n-3 (total value 24%), with a high ratio of UFAs to SFAs. The typical FAs composition makes both
C. coccineum oils suitable for nutritional application, as they are a rich source of functional compounds with potential health benefits [
16]. TO was characterized by a higher value of the ratio MUFAs/SFAs (2.05) than of SO (1.34). This result could be ascribable to several factors, such as differences in the FA metabolism due to the impact of the harvesting location (climate, soil, and water availability) and the diversity of metabolic precursors derived from different host plants. The higher level of FUFAs measured in TO was likely due to greater lipase activity in Tunisian
C. coccineum, which resulted in enzymatic hydrolysis during the harvesting, handling, and processing of oil [
28].
4. Conclusions
C. coccineum has long been used in the folk medicine of many countries in North Africa and the Middle East. Unfortunately, the arid climate of these countries means that C. coccineum specimens are few and are only available for a very small period of the year. In other regions of the Mediterranean basin, which have a cooler and more humid climate, C. coccineum specimens are available in greater quantities, although limited to a very short time span during the spring period.
In this work, we studied chemical and biological differences in C. coccineum growing in Sardinia (Italy) and Tunisia and have reported the first phytochemical characterization of the specimens from Tunisia, including the apolar portion.
On the whole, the collected data show significant differences between specimens from the two geographical sites. The Tunisian samples are richer in anthocyanins, betaine, proline, and unsaturated fatty acids, whereas the Sardinian sample showed higher contents of total phenolics, and saturated fatty acids.
This different composition could possibly be related to some plant defense mechanisms against climate stress; however, in both cases, the C. coccineum specimens appear to represent an adequate source of plant material suitable for nutraceutical applications.

 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}