Molecular and Morphological Phylogenetic Analysis of Naupactus Dejean (Curculionidae: Entiminae) and Allied Genera: The Dilemma of Classification


Abstract
:1. Introduction
- To recognize different groups of Naupactus and to analyze the relationships among them and with other genera of the Pantomorus-Naupactus complex: Aramigus Horn, Atrichonotus Buchanan, Eurymetopus Schoenherr, Floresianus Hustache, Galapaganus Lanteri, Hoplopactus Chevrolat, Lanterius Alonso-Zarazaga and Lyal, Pantomorus Schoenherr, Parapantomorus Emden, Phacepholis Horn, Symmathetes Schoenherr and Teratopactus Heller.
2. Materials and Methods
2.1. Taxon Sampling and Morphological Characters
2.2. Molecular Data: DNA Assay and Sequencing
2.3. Phylogenetic Analyses
3. Results
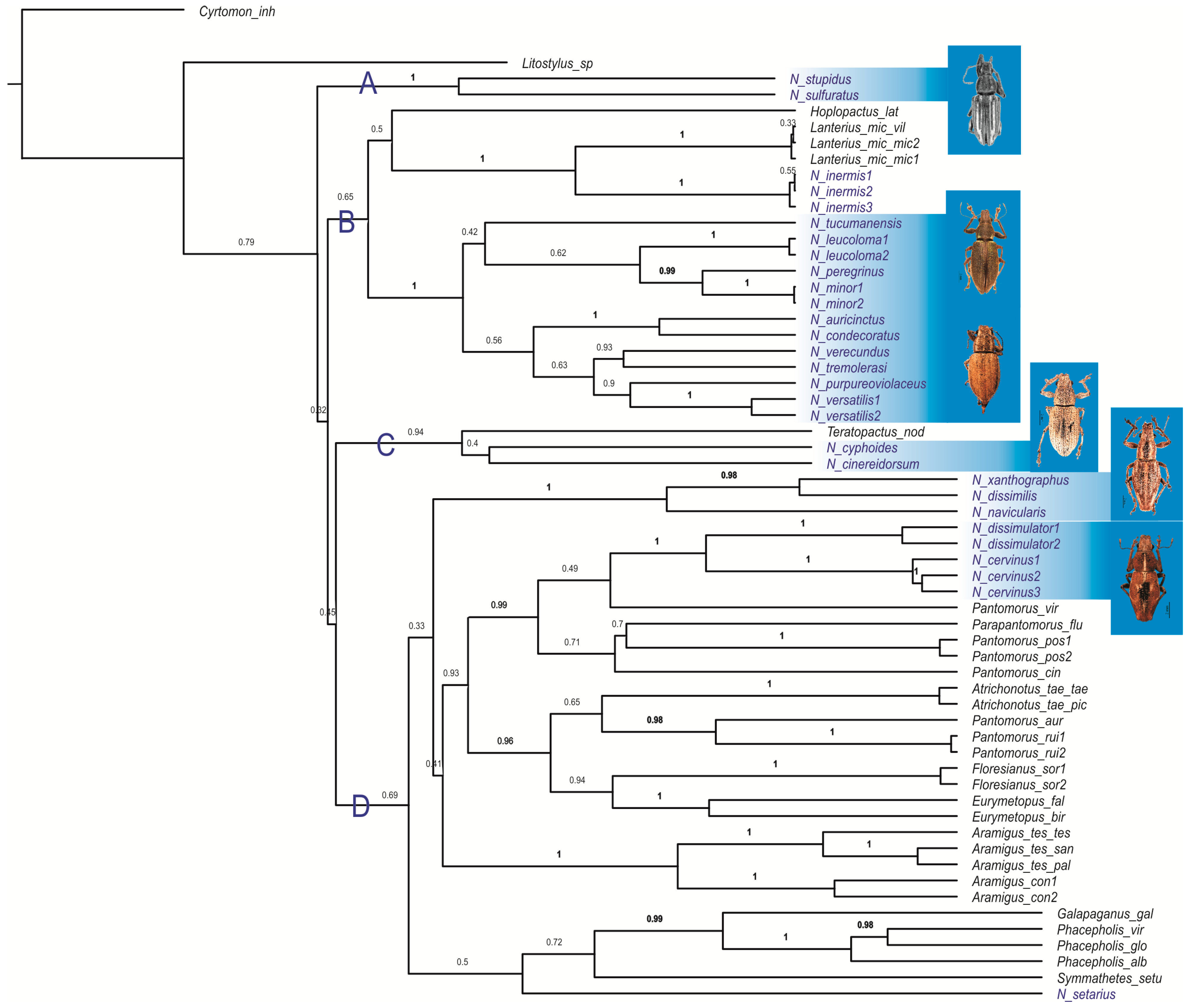
3.1. Bayesian Analysis
3.2. Parsimony Analysis
4. Discussion
4.1. Taxonomic Implications of Phylogenetic Analyses
4.2. Taxonomic Amendments
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Del Río, M.G.; Morrone, J.J.; Lanteri, A.A. Evolutionary biogeography of South American weevils of the tribe Naupactini (Coleoptera: Curculionidae). J. Biogeogr. 2015, 42, 1293–1304. [Google Scholar] [CrossRef]
- Lanteri, A.A.; del Río, M.G. Phylogeny of the tribe Naupactini (Coleoptera: Curculionidae) based on morphological characters. Syst. Entomol. 2017, 42, 429–447. [Google Scholar] [CrossRef]
- Alonso-Zarazaga, M.A.; Lyal, C.H.C. A World Catalogue of Families and Genera of Curculionoidea: (Insecta: Coleoptera) Excepting Scolytidae and Platypodidae; Entomopraxis, D.C.P.: Barcelona, Spain, 1999; ISBN 978-84-605-9994-4. [Google Scholar]
- Lanteri, A.A. Systematic revision and cladistic analysis of Phacepholis Horn (Coleoptera: Curculionidae). Southwest. Entomol. 1990, 15, 179–204. [Google Scholar]
- Lanteri, A.A. Systematic revision of Ericydeus Pascoe (Coleoptera: Curculionidae). Entomol. Scand. 1995, 26, 393–424. [Google Scholar] [CrossRef]
- Schoenherr, C.J. Genera et Species Curculionidum, cum Synonymia hujus Famili; Roret: Paris, France, 1833; Volume 1, pp. I–XV, 1–381, 383–681. [Google Scholar]
- Schoenherr, C.J. Genera et Species Curculionidum, cum Synonymia hujus Famili; Roret: Paris, France, 1840; Volume 6, pp. I, 1–474. [Google Scholar]
- Sharp, D.; Champion, G.C. Biologia Centrali-Americana; Taylor & Francis: London, UK, 1889–1911; Volume IV, part 3; p. vi-354. [Google Scholar]
- Hustache, A. Naupactini de l’Argentine et des régions limitrophes (Col. Curculion.). Rev. Soc. Entomol. Argent. 1947, 13, 3–146. [Google Scholar]
- Lanteri, A.A.; Marvaldi, A.E. Graphognathus Buchanan, a new synonym of Naupactus Dejean, and systematics of the N. leucoloma species group (Coleoptera: Curculionidae). Coleopt. Bull. 1995, 49, 206–228. [Google Scholar]
- Bordón, C. El género Naupactus Dejean (Coleoptera: Curculionidae) en Venezuela. Acta Biológica Venezuelica 1997, 17, 11–51. [Google Scholar]
- Morrone, J.J. The species of Entiminae ranged in America south of the United States. Anales del Instituto de Biología Serie Zoología 1999, 70, 99–168. [Google Scholar]
- Wibmer, G.J.; O’Brien, C.W. Annotated checklist of the weevils (Curculionidae sensu lato) of South America (Coleoptera: Curculionoidea). Mem. Am. Entomol. Inst. 1986, 39, 1–563. [Google Scholar]
- O’Brien, C.W.; Wibmer, G.J. Annotated checklist of the weevils (Curculionidae sensu lato) of North America, Central America and the West Indies (Coleoptera: Curculionoidea). Mem. Am. Entomol. Inst. 1982, 34, 1–382. [Google Scholar]
- Scataglini, M.A.; Lanteri, A.A.; Confalonieri, V.A. Phylogeny of the Pantomorus-Naupactus complex based on morphological and molecular data (Coleoptera: Curculionidae). Cladistics 2005, 21, 131–142. [Google Scholar] [CrossRef]
- Rosas-Echeverría, M.V.; Morrone, J.J.; del Río, M.G.; Lanteri, A.A. Phylogenetic analysis of the Pantomorus-Naupactus complex (Coleoptera: Curculionidae: Entiminae) from North and Central America. Zootaxa 2011, 2780, 1–19. [Google Scholar]
- Buchanan, L.L. The Species of Pantomorus of America North of Mexico; United States Department of Agriculture: Washington, DC, USA, 1939; Volume 34, pp. 1–39. [Google Scholar]
- Lanteri, A.A.; O’Brien, C.W. Taxonomic revision and cladistic analysis of Atrichonotus Buchanan (Coleoptera: Curculionidae). Trans. Am. Entomol. Soc. 1990, 116, 697–725. [Google Scholar]
- Lanteri, A.A.; del Río, M.G. Caracteres genitales de la hembra en la clasificación y filogenia de la tribu Naupactini (Coleoptera: Curculionidae). In Contribuciones Taxonómicas en Órdenes de Insectos Hiperdiversos; Llorente Bousquets, J., Lanteri, A.A., Eds.; UNAM-RIBES-CYTED: Mexico city, Mexico, 2008; pp. 159–176. [Google Scholar]
- Rosas, M.V.; del Río, M.G.; Lanteri, A.A.; Morrone, J.J. Track analysis of the North and Central American species of the Pantomorus-Naupactus complex (Coleoptera: Curculionidae). J. Zool. Syst. Evol. Res. 2011, 49, 309–314. [Google Scholar] [CrossRef]
- Lanteri, A.A.; del Río, M.G. Naupactus xanthographus (Germar) species group (Curculionidae: Entiminae: Naupactini): A comprehensive taxonomic treatment. J. Nat. Hist. 2017, 51, 1557–1587. [Google Scholar]
- Marvaldi, A.E.; Lanteri, A.A.; del Río, M.G.; Oberprieler, R.G. 3.7.5. Entiminae Schoenherr, 1823. In Handbook of Zoology. Coleoptera, Beetles. Morphology and Systematics, Vol. 3; Leschen, R.A., Beutel, R.G., Eds.; De Gruyter: Berlin, Germany; Boston, MI, USA, 2014; Volume 3, pp. 503–522. [Google Scholar]
- Bold Systems v4. Available online: http://www.boldsystems.org/ (accessed on 4 May 2018).
- Marvaldi, A.E.; del Río, M.G.; Pereyra, V.A.; Rocamundi, N.; Lanteri, A.A. A combined molecular and morphological approach to explore higher phylogenetics of entimine weevils with special reference to South American taxa. Diversity 2018. under review. [Google Scholar]
- Sequeira, A.; Lanteri, A.A.; Scataglini, M.A.; Confalonieri, V.A.; Farrell, B. Are flightless Galapaganus weevils older than the Galápagos Islands they inhabit? Heredity 2000, 85, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Rodriguero, M.S.; Lanteri, A.A.; Confalonieri, V.A. Speciation in the asexual realm: Is the parthenogenetic weevil Naupactus cervinus a complex of species in statu nascendi? Mol. Phylogenet. Evol. 2013, 68, 644–656. [Google Scholar] [CrossRef] [PubMed]
- Elias-Costa, A.J.; Confalonieri, V.A.; Lanteri, A.A.; Rodriguero, M.S. Game of Clones: Is Wolbachia inducing speciation in a weevil with a mixed reproductive mode? Mol. Phylogenet. Evol. 2018. under review. [Google Scholar]
- Sunnucks, P.; Hales, D.F. Numerous transposed sequences of mitochondrial cytochrome oxidase I-II in aphids of the genus Sitobion (Hemiptera: Aphididae). Mol. Biol. Evol. 1996, 13, 510–524. [Google Scholar] [CrossRef] [PubMed]
- Normark, B.B. Phylogeny and Evolution of Parthenogenesis in the Aramigus tessellatus Complex (Coleoptera: Curculionidae). Ph.D. Thesis, Cornell University, Ithaca, NY, USA, 1994. [Google Scholar]
- Bensansson, D.; Zhang, D.; Hewitt, G.M. Frequent assimilation of mitochondrial DNA by grasshopper nuclear genomes. Mol. Biol. Evol. 2000, 17, 406–415. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Lunt, D.H.; Zhand, D.X.; Szymura, J.M.; Hewitt, G.M. The insect cytochrome oxidase I gene evolutionary patterns and conserved primers for phylogenetic studies. Insect Mol. Biol. 1996, 5, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Sorenson, M.D.; Fleischer, R.C. Multiple independent transposition of mitochondrial DNA control region sequences to the nucleus. Proc. Natl. Acad. Sci. USA 1996, 93, 15239–15243. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.X.; Hewitt, G.M. Nuclear integrations: Challenges for mitochondrial DNA markers. Trends Ecol. Evol. (Amst.) 1996, 11, 247–251. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Goloboff, P.A.; Catalano, S. TNT version 1.5, including full implementation of phylogenetic morphometrics. Cladistics 2016, 32, 221–238. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Kluge, A.G.; Farris, J.S. Quantitative phyletics and the evolution of anurans. Syst. Biol. 1969, 18, 1–32. [Google Scholar] [CrossRef]
- Farris, J.S. The retention index and the rescaled consistency index. Cladistics 1989, 5, 417–419. [Google Scholar] [CrossRef]
- Bouckaert, R.; Heled, J.; Kühnert, D.; Vaughan, T.; Wu, C.H.; Xie, D.; Suchard, M.A.; Drummond, A.J. BEAST 2: A Software Platform for Bayesian Evolutionary Analysis. PLoS Comput. Biol. 2014, 10, e1002537. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Pfeiffe, W.; Schwartz, T. Creating the CIPRES Science Gate way for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; Towns, J., Ed.; IEEE: New Orleans, LA, USA, 2010. [Google Scholar]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach; Springer Science & Business Media: New York, NY, USA, 2002. [Google Scholar]
- Drumond, A.J.; Rambaut, A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol. Biol. 2007, 7, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A. FigTree v. 1.3.1. Computer Program and Documentation Distributed by the Author, 2006–2012. Available online: http://tree.bio.ed.ac.uk/software/ (accessed on 21 December 2017).
- Lanteri, A.A. Revisión de las especies argentinas del género Macrostylus Boheman, subgénero Mimographus Schoenherr (Coleoptera: Curculionidae). CIPFE-CED Orione Contribuciones en Biología 1985, 12, 1–6. [Google Scholar]
- Normark, B.B.; Lanteri, A.A. Incongruence between morphological and mitochondrial DNA characters suggests hybrid origins of parthenogenetic weevil lineages (genus Aramigus). Syst. Biol. 1998, 47, 475–494. [Google Scholar] [CrossRef] [PubMed]
- Lanteri, A.A.; del Río, M.G.; Rodriguero, M.; Confalonieri, V. Weevils of the Pantomorus-Naupactus complex: Cladistics and generic classification. Cladistics 2010, 26, 214. [Google Scholar]
- Del Río, M.G.; Lanteri, A.A.; Guedes, J.V.C. Taxonomic revision and cladistic analysis of Teratopactus Heller (Coleoptera: Curculionidae). Invertebr. Syst. 2006, 20, 585–602. [Google Scholar] [CrossRef]
- Lanteri, A.A. Revisión del género Asynonychus Crotch (Coleoptera: Curculionidae). Rev. Asoc. Cs. Nat. Litoral. 1986, 2, 161–174. [Google Scholar] [CrossRef]
- Blackwelder, R.E. Checklist of the Coleopterous insects of Mexico, Central America, the West Indies and South America. Part 5. Bull. U. S. Natl. Mus. 1947, 185, 765–925. [Google Scholar] [CrossRef]
- Lanteri, A.A.; Díaz, N.B. Systematic study and cladistic analysis of the genus Aramigus Horn (Coleoptera: Curculionidae). Trans. Am. Entomol. Soc. 1994, 120, 113–144. [Google Scholar]
- Lanteri, A.A. Systematics, cladistics and biogeography of a new weevil genus Galapaganus (Coleoptera: Curculionidae) from the Galápagos Islands, and coasts of Ecuador and Perú. Trans. Am. Entomol. Soc. 1992, 118, 227–267. [Google Scholar]
- Lanteri, A.A. New Taxonomic and Biogeographic Information on Galapaganus femoratus species group (Coleoptera: Curculionidae: Entiminae). Trans. Am. Entomol. Soc. 2004, 130, 177–192. [Google Scholar]
- Lanteri, A.A.; Morrone, J.J. Cladistics of the Naupactus leucoloma species group, Atrichonotus, and Eurymetopus (Coleoptera: Curculionidae). Revista de la Sociedad Entomológica Argentina 1995, 54, 99–112. [Google Scholar]
- Lanteri, A.A.; Normark, B.B. Parthenogenesis in the tribe Naupactini (Coleoptera: Curculionidae). Ann. Entomol. Soc. Am. 1995, 88, 722–731. [Google Scholar] [CrossRef]
- Lanteri, A.A. Revisión sistemática del género Eurymetopus Schoenherr (Coleoptera: Curculionidae) mediante la aplicación de técnicas numéricas. Revista de la Sociedad Entomológica Argentina 1984, 43, 247–281. [Google Scholar]
- Sequeira, A.; Sijapati, M.; Lanteri, A.; Roque Albelo, L. Nuclear and mitochondrial sequences confirm complex colonization patterns and clear species boundaries for flightless weevils in the Galapagos archipelago. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2008, 363, 3439–3451. [Google Scholar] [CrossRef] [PubMed]
- Sequeira, A.S.; Lanteri, A.A.; Albelo, L.R.; Bhattacharya, S.; Sijapati, M. Colonization history, ecological shifts and diversification in the evolution of endemic Galápagos weevils: Colonization and diversification of Galápagos weevils. Mol. Ecol. 2008, 17, 1089–1107. [Google Scholar] [CrossRef] [PubMed]
- Mayr, E. Principles of Systematic Zoology; McGraw-Hill: New York, NY, USA, 1969. [Google Scholar]
- Hennig, W. Phylogenetic Systematics; University of Illinois Press: Urbana, IL, USA, 1966. [Google Scholar]
- Marvaldi, A.E.; Sequeira, A.S.; O’Brien, C.W.; Farrell, B.D. Molecular and Morphological Phylogenetics of Weevils (Coleoptera, Curculionoidea): Do Niche Shifts Accompany Diversification? Syst. Biol. 2002, 51, 761–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jordal, B.H.; Sequeira, A.S.; Cognato, A.I. The age and phylogeny of wood boring weevils and the origin of subsociality. Mol. Phylogenet. Evol. 2011, 59, 708–724. [Google Scholar] [CrossRef] [PubMed]
- Gunter, N.L.; Oberprieler, R.G.; Cameron, S.L. Molecular phylogenetics of Australian weevils (Coleoptera: Curculionoidea): Exploring relationships in a hyperdiverse lineage through comparison of independent analyses. Austral Entomol. 2016, 55, 217–233. [Google Scholar] [CrossRef]
- Shin, S.; Clarke, D.J.; Lemmon, A.R.; Moriarty Lemmon, E.; Aitken, A.L.; Haddad, S.; Farrell, B.D.; Marvaldi, A.E.; Oberprieler, R.G.; McKenna, D.D. Phylogenomic Data Yield New and Robust Insights into the Phylogeny and Evolution of Weevils. Mol. Biol. Evol. 2018, 35, 823–836. [Google Scholar] [CrossRef] [PubMed]
- Franz, N.M. On the lack of good scientific reasons for the growing phylogeny/classification gap. Cladistics 2005, 21, 495–500. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Species | Acronyms | Location | Accession Numbers | Source |
|---|---|---|---|---|
| Cyrtomon inhalatus (Germar) | Cyrtomon_inh | AR, Entre Ríos, Victoria | MH537926 | |
| Litostylus sp. | Litostylus_sp | Dominica Island, Warner, Caribbean | HQ891471.1 | |
| Aramigus tessellatus Say, morph. tessellatus | Aramigus_tes_tes | AR, Buenos Aires, Otamendi | MH537929 | |
| Aramigus tessellatus Say, morph. pallidus | Aramigus_tes_pal | AR, Buenos Aires, Punta Lara | MH537928 | |
| Aramigus tessellatus Say, morph. santafecinus | Aramigus_tes_san | AR, Buenos Aires, La Plata | MH537927 | |
| Aramigus conirostris (Hustache) | Aramigus_con1 | UR, San José, Libertad | MH537930 | |
| Aramigus_con2 | AR, Entre Ríos, Paraná | U25295 | [23] | |
| Atrichonotus taeniatulus (Berg), morph. taeniatulus | Atrichonotus_tae_tae | AR, Mendoza, Guaymallén | MH537919 | [23] |
| Atrichonotus taeniatulus (Berg), morph. pictipennis | Atrichonotus_tae_pic | AR, Buenos Aires, Arrecifes | MH537931 | |
| Eurymetopus birabeni Kuschel | Eurymetopus_bir | AR, Buenos Aires | AY790877 | [23] |
| Eurymetopus fallax Boheman | Eurymetopus_fal | AR, Buenos Aires | AY790878 | [23] |
| Floresianus sordidus Hustache | Floresianus_sor1 | AR, Misiones | MH537932 | |
| Floresianus_sor2 | BR, RG do Sul, Santa María | MH537933 | ||
| Galapaganus galapagoensis (Linell) | Galapaganus_gal | EC, Galápagos, San Cristobal | AF015914 | [23] |
| Hoplopactus lateralis Arrow | Hoplopactus_lat | BR, São Paulo | MH537920 | [23] |
| * Lanterius inermis (Hustache) | N_inermis1 | AR, Misiones, Urugua-í | MH537908 | IBOL MLPCU0411 |
| N_inermis2 | AR, Misiones, Urugua-í | MH537909 | IBOL MLPCU0412 | |
| N_inermis3 | AR, Misiones, PP Moconá | MH537910 | IBOL MLPCU0407 | |
| Lanterius micaceus (Hustache), morph. micaceus | Lanterius_mic_mic1 | AR, Misiones, Urugua-í | MH537911 | IBOL MLPCU0420 |
| Lanterius_mic_mic2 | AR, Misiones, Urugua-í | MH537912 | IBOL MLPCU0418 | |
| Lanterius micaceus (Hustache), morph. villosipennis | Lanterius_mic_vil | AR, Misiones, Urugua-í | MH537913 | IBOL MLPCU0427 |
| Naupactus auricinctus Boheman | N_auricinctus | BR, São Paulo | MH537921 | [23] |
| Naupactus cervinus Boheman | N_cervinus1 | AR, Misiones, Cerro Azul | JX440490.1 | [23] |
| N_cervinus2 | BR, Misiones, Oberá | GQ406843.1 | [23] | |
| N_cervinus3 | AR, Córdoba, Río Cuarto | GQ406828.1 | [23] | |
| Naupactus cinereidorsum Hustache | N_cinereidorsum | AR, Córdoba | AY770388 | [23] |
| Naupactus condecoratus Boheman | N_condecoratus | AR, Misiones, PP Moconá | MH537914 | IBOL MLPCU00406 |
| Naupactus cyphoides (Heller) | N_cyphoides | AR, Misiones, San Ignacio | MH537942 | |
| Naupactus dissimilis Hustache | N_dissimilis | AR, Misiones, Yacutinga | MH537940 | |
| Naupactus dissimulator Boheman | N_dissimulator1 | AR, Misiones, PP Las Araucarias | MH537915 | IBOL MLPCU0041 |
| N_dissimulator2 | AR, Buenos Aires, Punta Lara | JX440494 | [23] | |
| Naupactus leucoloma Boheman | N_leucoloma1 | AR, Mendoza | MH537922 | [23] |
| N_leucoloma2 | AR, Entre Ríos, Victoria | MH537934 | ||
| Naupactus minor (Buchanan) | N_minor1 | AR, Entre Ríos | AY790881 | [23] |
| N_minor2 | AR, Buenos Aires | EU264960 | [23] | |
| Naupactus navicularis Boheman | N_navicularis | BR, São Paulo | AY790882 | [23] |
| Naupactus peregrinus (Boheman) | N_peregrinus | AR, Entre Ríos, Concordia | MH537935 | |
| Naupactus purpureoviolaceus Hustache | N_purpureoviolaceus | AR, Entre Ríos, Concordia | MH537936 | |
| Naupactus stupidus Boheman | N_stupidus | ME, Oaxaca, Salina Cruz | GU565274 | [23] |
| Naupactus sulfuratus Champion | N_sulfuratus | ME, Oaxaca, Salina Cruz | GU565270 | [23] |
| Naupactus tremolerasi Hustache | N_tremolerasi | BR, RG do Sul, Santa María | MH537937 | |
| Naupactus tucumanensis Hustache | N_tucumanensis | AR, Tucumán | MH537938 | |
| Naupactus verecundus Hustache | N_verecundus | AR, La Pampa, Santa Rosa | AF211490 | [23] |
| Naupactus versatilis Hustache | N_versatilis1 | AR, CABA | MH537939 | |
| N_versatilis2 | AR, Misiones, Teyú Cuaré | MH537916 | IBOL MLPCU0117 | |
| Naupactus xanthographus (Germar) | N_xanthographus | AR, Buenos Aires, Punta Lara | AY790880.1 | [23] |
| Pantomorus auripes Hustache | Pantomorus_aur | AR, Córdoba | AY770383 | [23] |
| Pantomorus cinerosus (Boheman) | Pantomorus_cin | AR, Córdoba | AY770384 | [23] |
| Pantomorus postfasciatus (Hustache) (misidentified as N. ambiguus [23]) | Pantomorus_pos1 | AR, Chaco, Resistencia | MH537917 | |
| Pantomorus_pos2 | BR, RG de Sul, Santa Maria | MH537918 | ||
| Pantomorus ruizi (Brèthes) | Pantomorus_rui1 | AR, Chubut, Trelew | MH537925 | [23] |
| Pantomorus_rui2 | AR, La Pampa | AY770385 | [23] | |
| Pantomorus viridisquamosus (Boheman) | Pantomorus_vir | AR, Buenos Aires | AY770386 | [23] |
| Parapantomorus fluctuosus (Boheman) | Parapantomorus_flu | BR, São Paulo | MH537941 | |
| Phacepholis albicans (Sharp) | Phacepholis_alb | ME, Gerrero, Tecpan | GU565278 | [23] |
| Phacepholis globicollis (Pascoe) | Phacepholis_glo | ME, Oaxaca, Salina Cruz | GU565273 | [23] |
| Phacepholis viridicans (Sharp) | Phacepholis_vir | ME, Jalisco, Chamela | GU565277 | [23] |
| *Symmathetes setarius (Boheman) | N_setarius | BR, Mato Grosso | MH537923 | [24] |
| Symmathetes setulosus Hustache | Symmathetes_setu | AR, Catamarca, Las Esquinas | MH537924 | [24] |
| Teratopactus nodicollis (Boheman) | Teratopactus_nod | BR, São Paulo | AY770387 | [15] |
|
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Del Río, M.G.; Rodriguero, M.S.; Confalonieri, V.A.; Lanteri, A.A. Molecular and Morphological Phylogenetic Analysis of Naupactus Dejean (Curculionidae: Entiminae) and Allied Genera: The Dilemma of Classification. Diversity 2018, 10, 59. https://doi.org/10.3390/d10030059
Del Río MG, Rodriguero MS, Confalonieri VA, Lanteri AA. Molecular and Morphological Phylogenetic Analysis of Naupactus Dejean (Curculionidae: Entiminae) and Allied Genera: The Dilemma of Classification. Diversity. 2018; 10(3):59. https://doi.org/10.3390/d10030059
Chicago/Turabian StyleDel Río, Maria G., Marcela S. Rodriguero, Viviana A. Confalonieri, and Analía A. Lanteri. 2018. "Molecular and Morphological Phylogenetic Analysis of Naupactus Dejean (Curculionidae: Entiminae) and Allied Genera: The Dilemma of Classification" Diversity 10, no. 3: 59. https://doi.org/10.3390/d10030059
APA StyleDel Río, M. G., Rodriguero, M. S., Confalonieri, V. A., & Lanteri, A. A. (2018). Molecular and Morphological Phylogenetic Analysis of Naupactus Dejean (Curculionidae: Entiminae) and Allied Genera: The Dilemma of Classification. Diversity, 10(3), 59. https://doi.org/10.3390/d10030059