Microbial Community in Hyperalkaline Steel Slag-Fill Emulates Serpentinizing Springs


Abstract
:1. Introduction
2. Materials and Methods
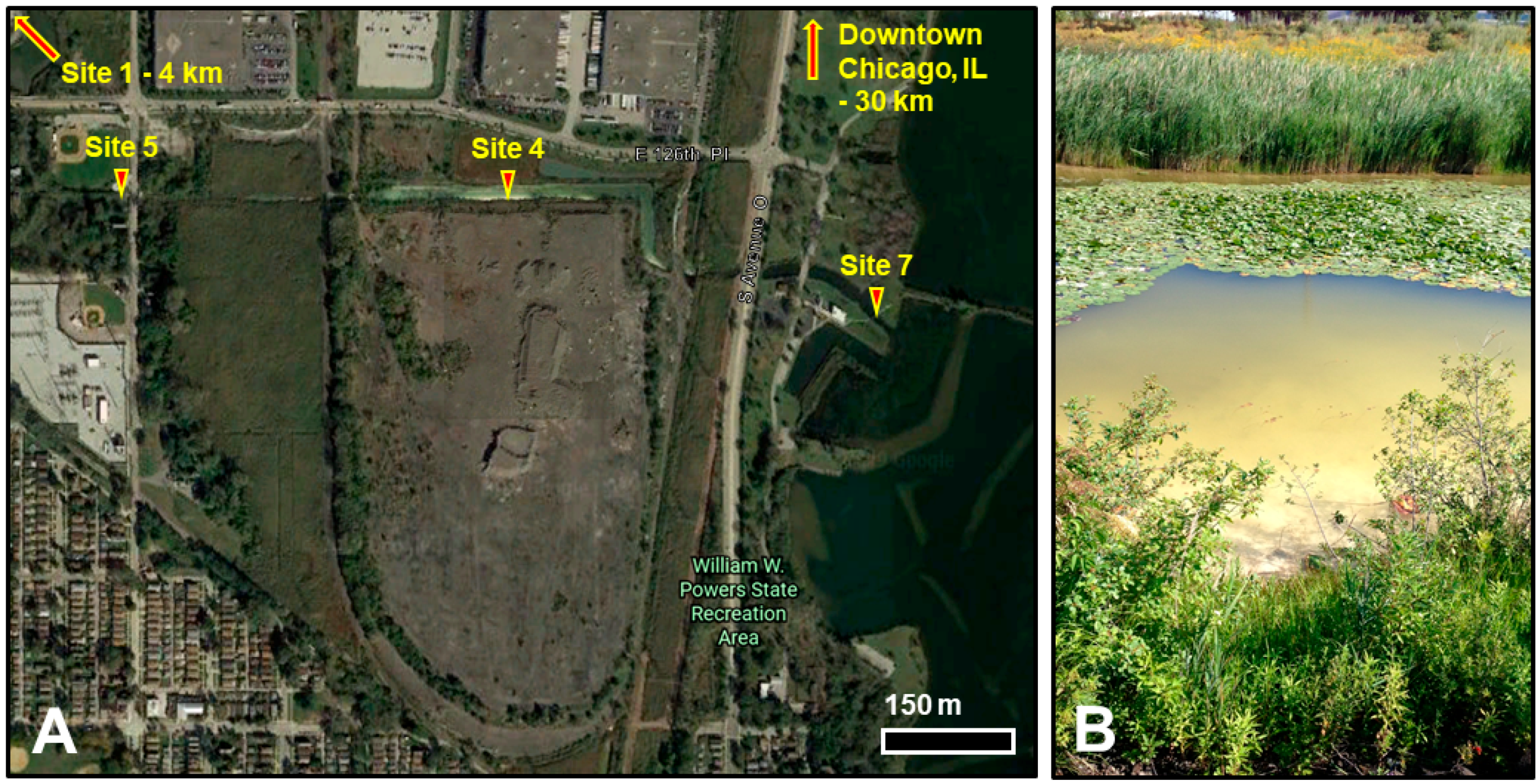
2.1. Site Description and Sampling
2.2. DNA Extraction and 16S rRNA Gene Amplification and Sequencing
2.3. 16S rRNA Gene Classification and Analysis
2.4. Metagenomic Sequencing, Assembly, and Analysis
3. Results
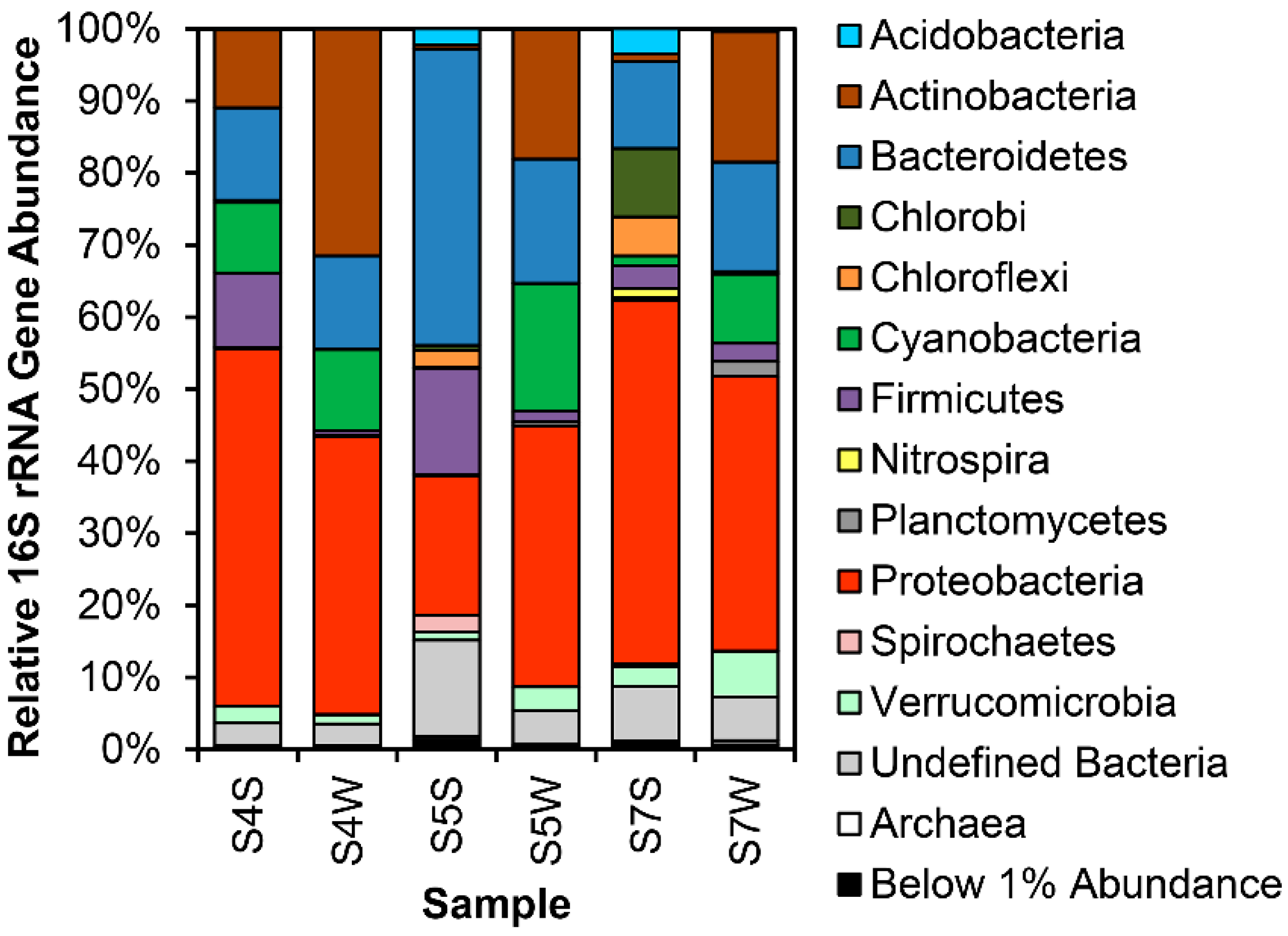
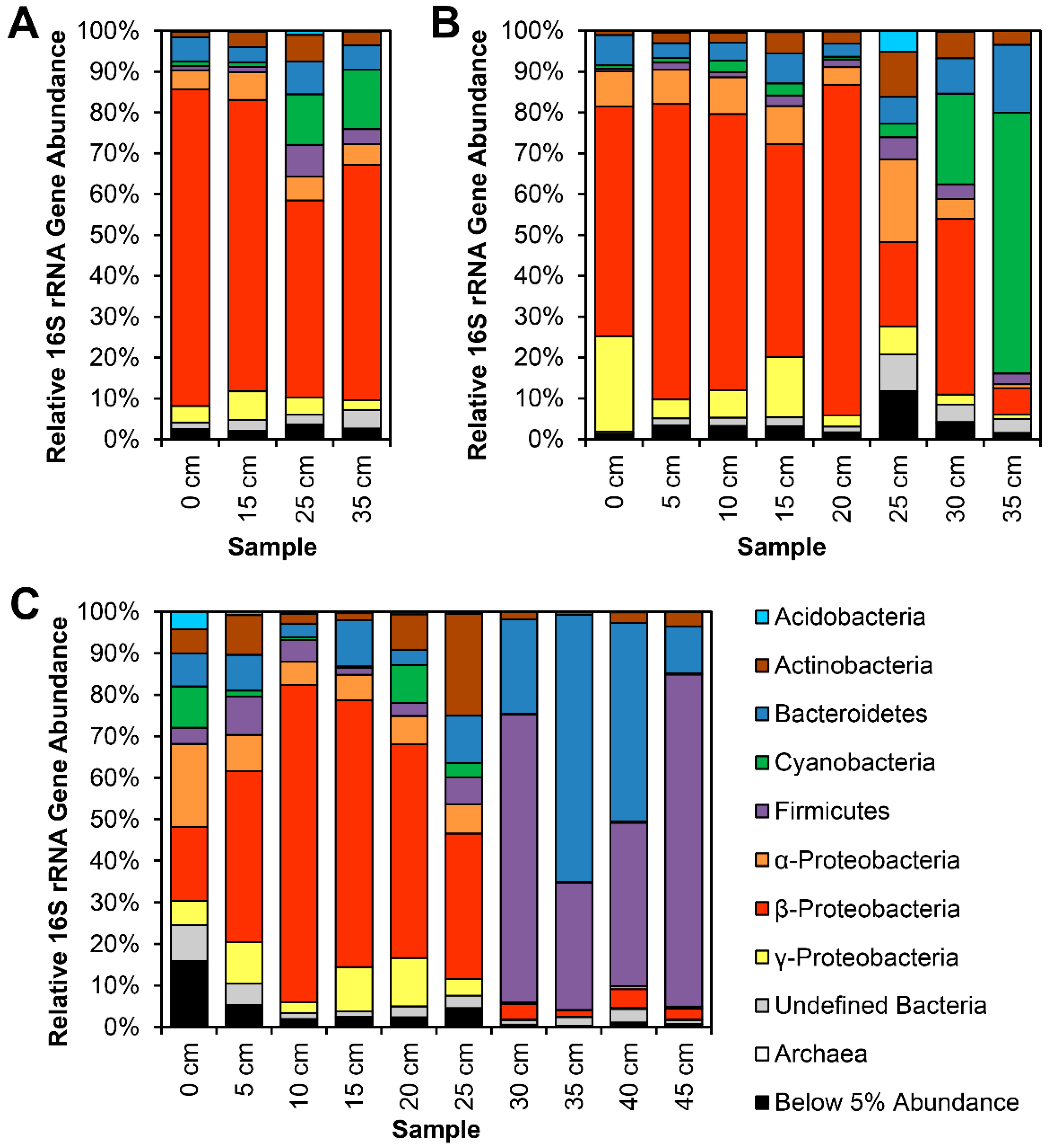
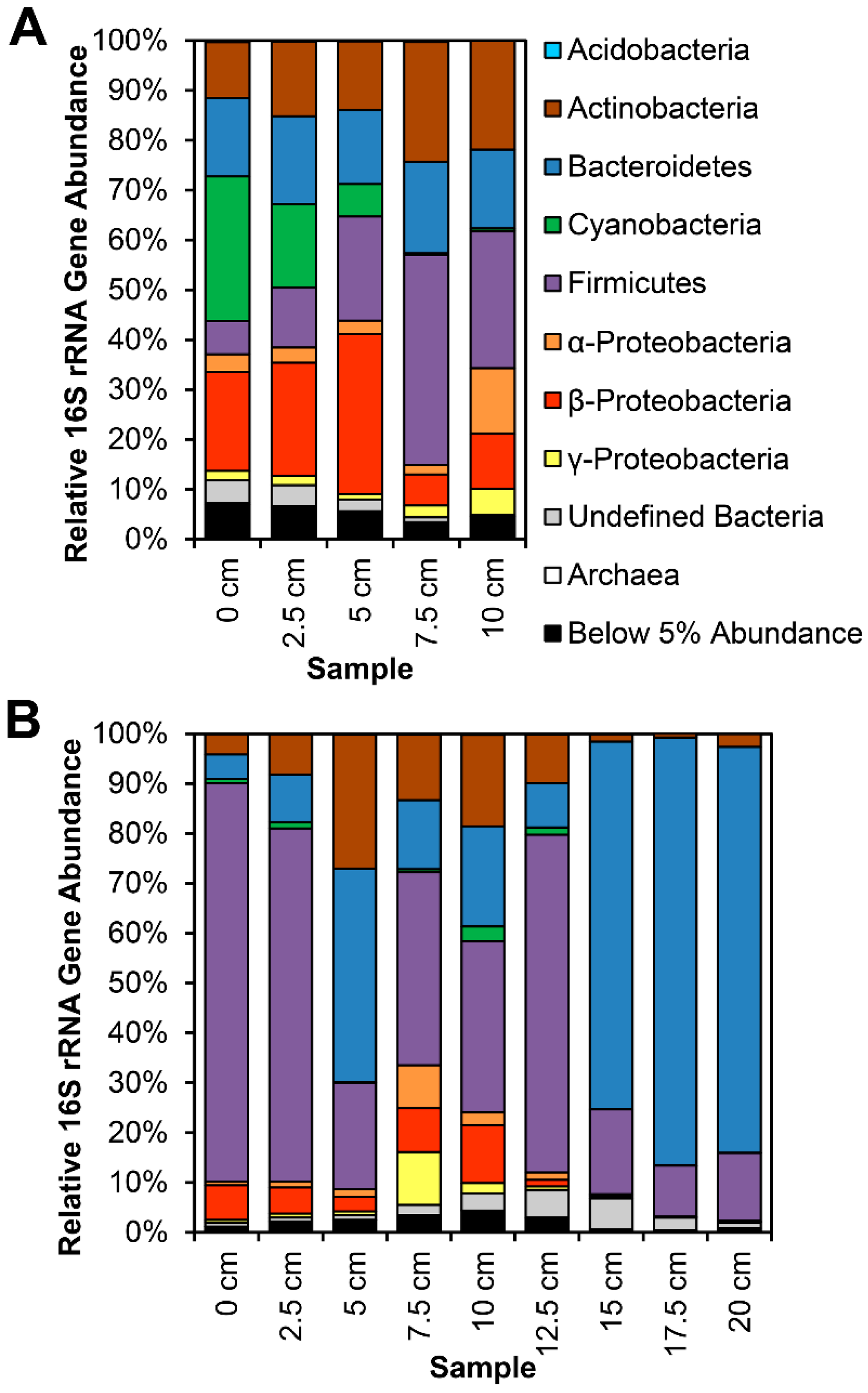
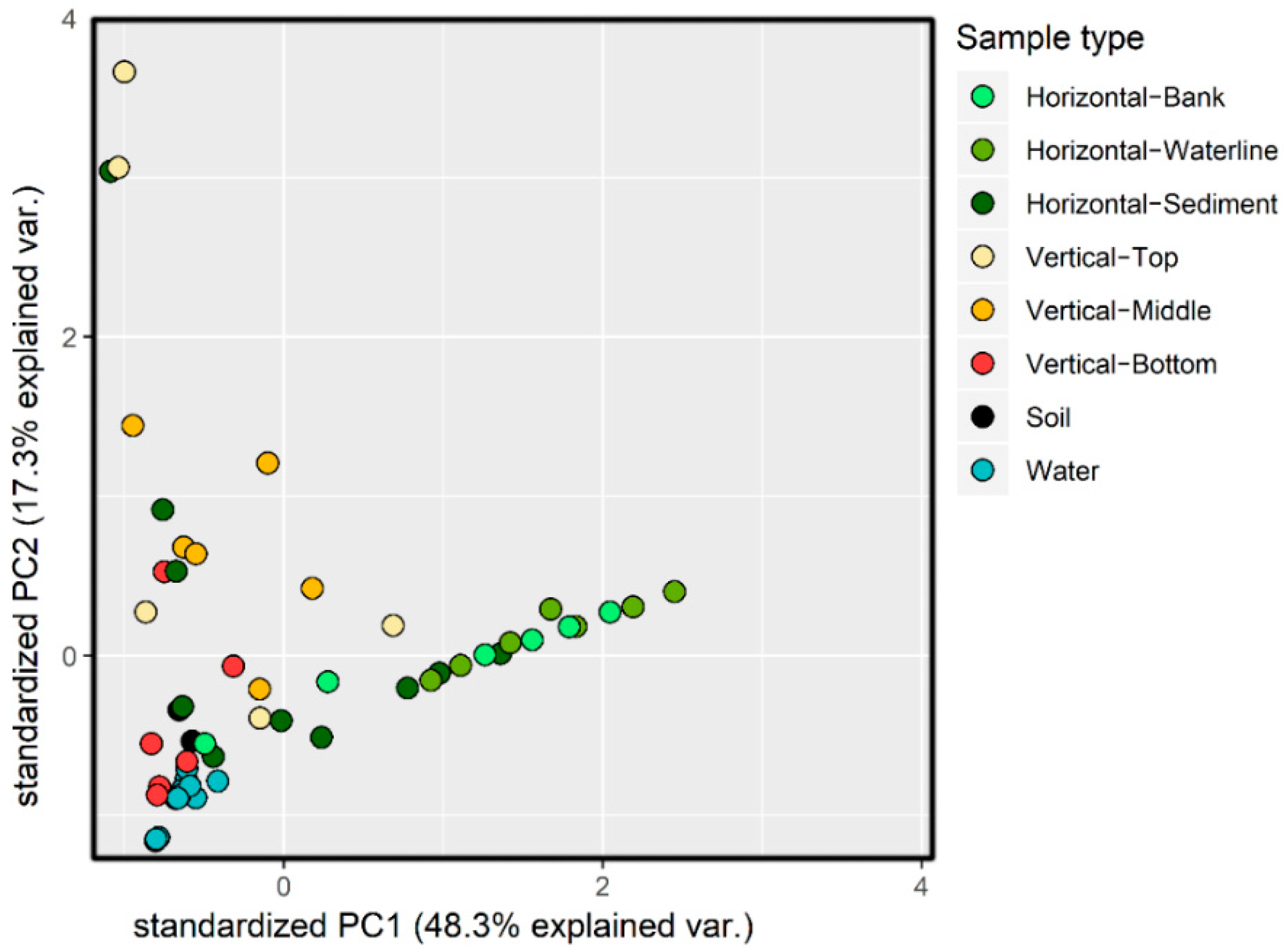
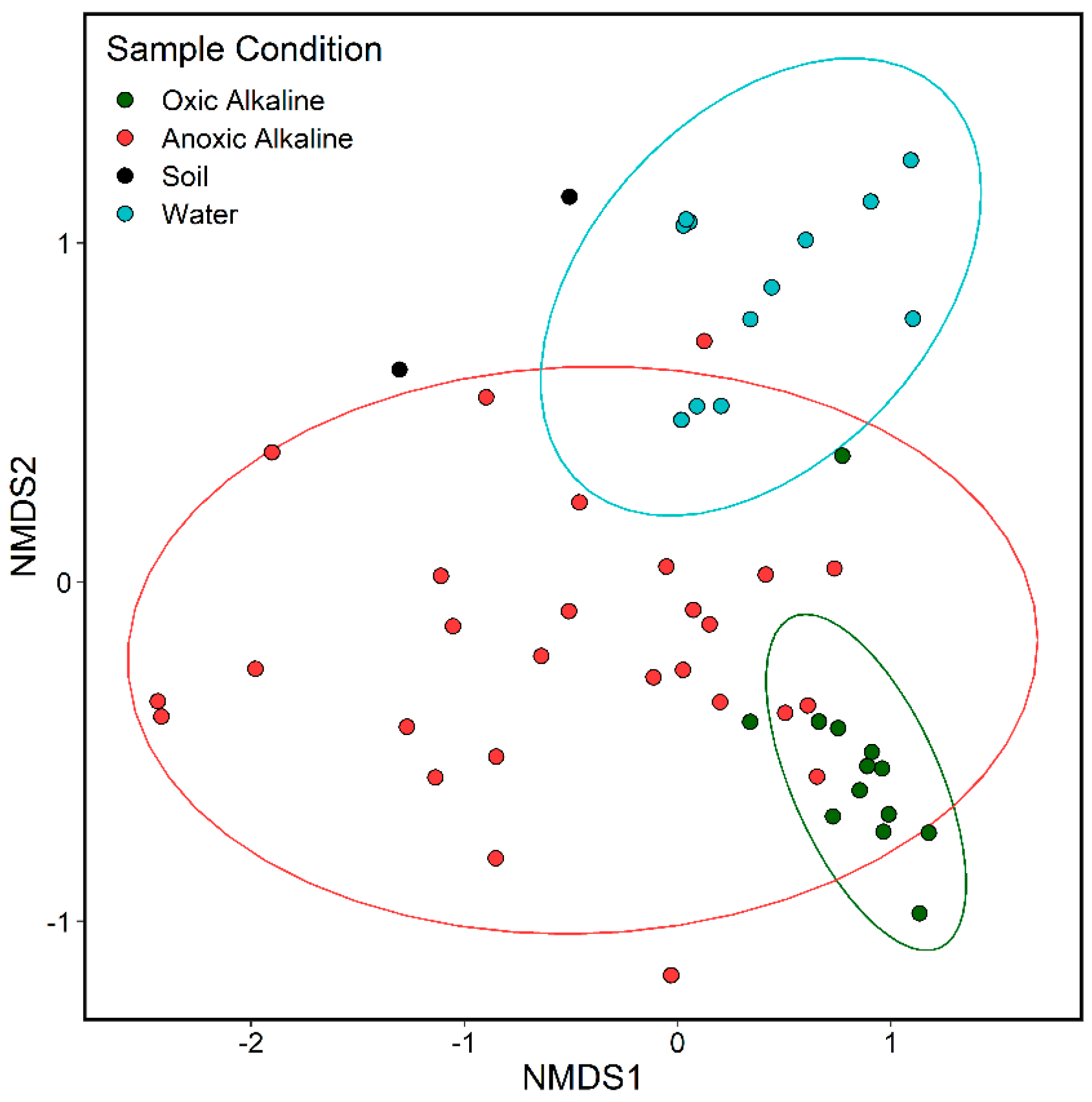
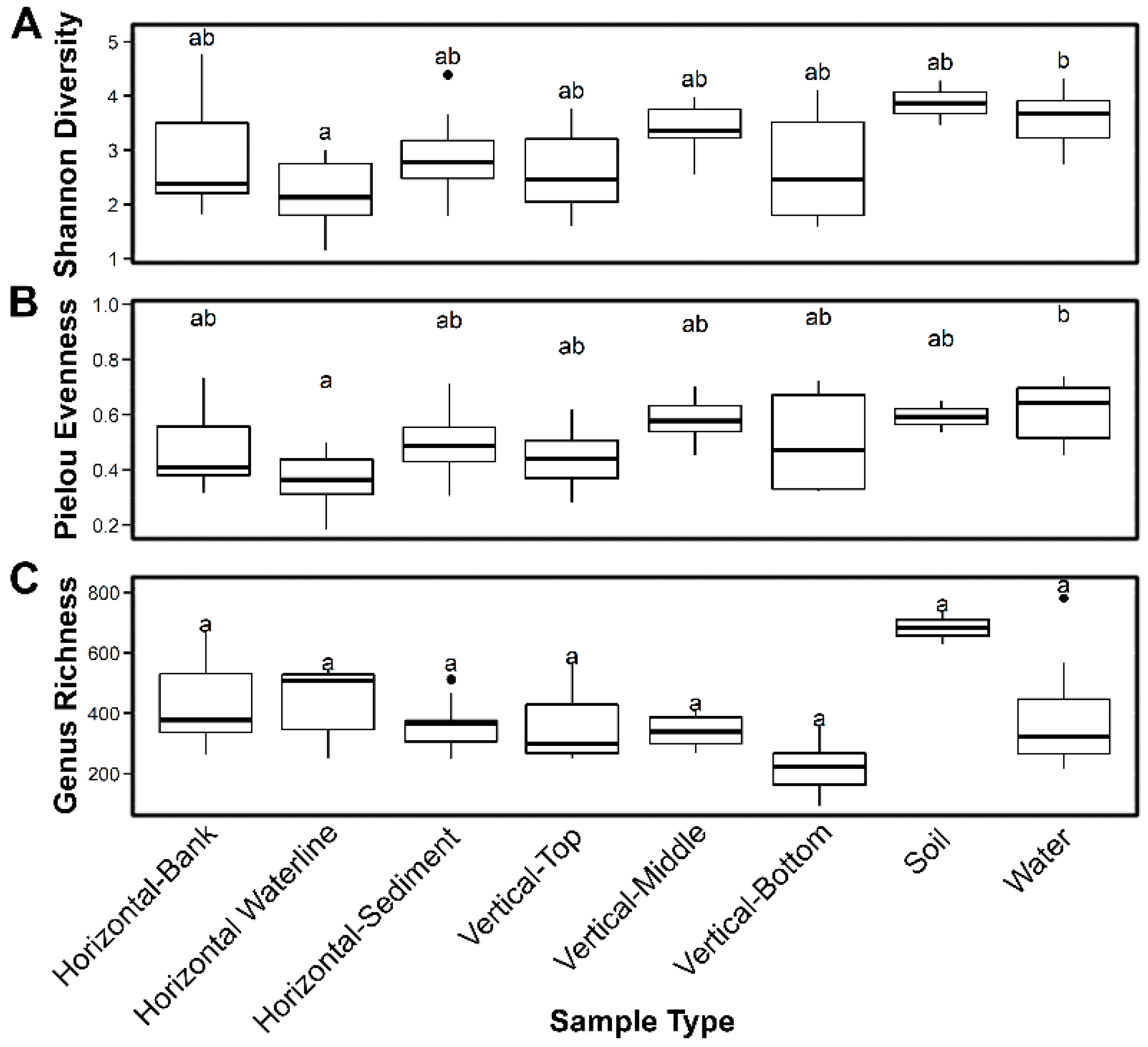
3.1. 16S rRNA Gene Analysis
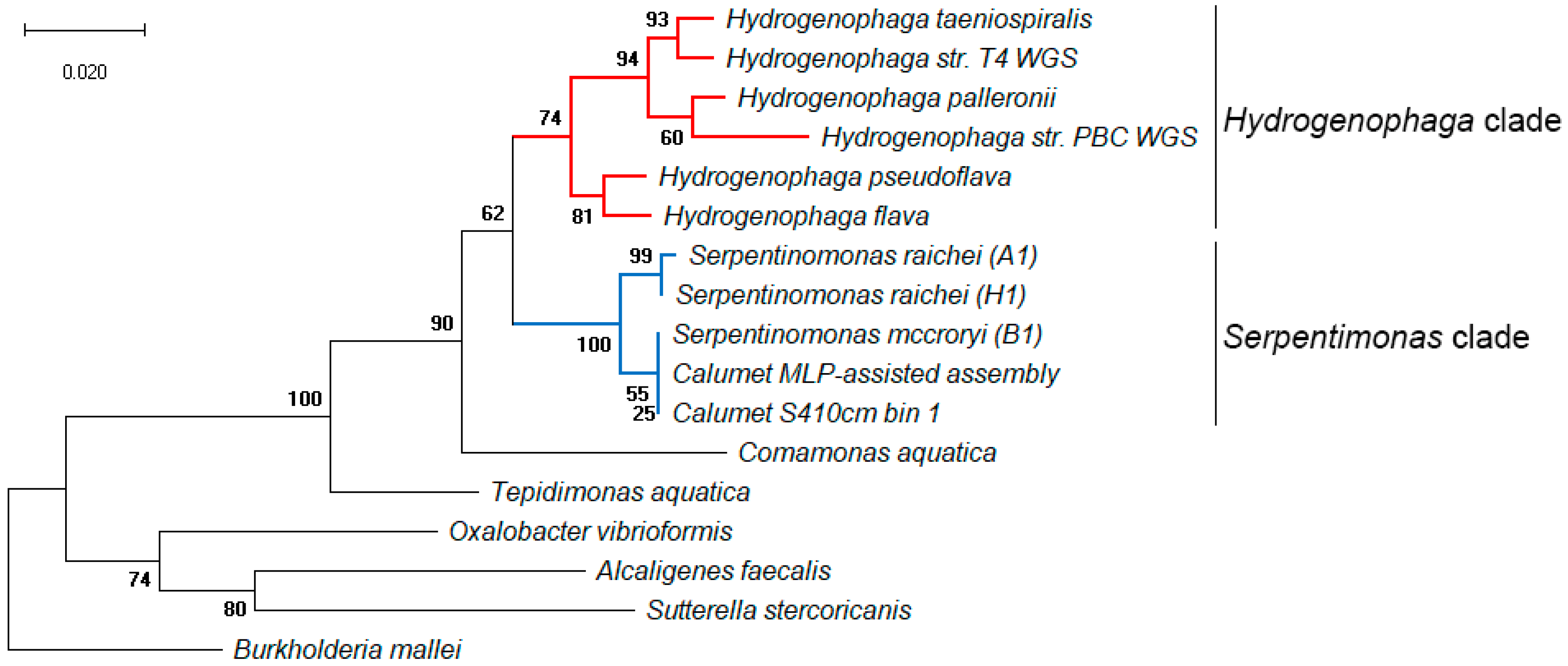
3.2. Metagenomic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rothschild, L.J.; Mancinelli, R.L. Life in extreme environments. Nature 2001, 409, 1092. [Google Scholar] [CrossRef] [PubMed]
- Pikuta, E.V.; Hoover, R.B.; Tang, J. Microbial Extremophiles at the Limits of Life. Crit. Rev. Microbiol. 2007, 33, 183–209. [Google Scholar] [CrossRef] [PubMed]
- Harrison, J.P.; Gheeraert, N.; Tsigelnitskiy, D.; Cockell, C.S. The limits for life under multiple extremes. Trends Microbiol. 2013, 21, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Bonilla, J.O.; Kurth, D.G.; Cid, F.D.; Ulacco, J.H.; Gil, R.A.; Villegas, L.B. Prokaryotic and eukaryotic community structure affected by the presence of an acid mine drainage from an abandoned gold mine. Extremophiles 2018, 22, 699–711. [Google Scholar] [CrossRef] [PubMed]
- Kalwasińska, A.; Felföldi, T.; Szabó, A.; Deja-Sikora, E.; Kosobucki, P.; Walczak, M. Microbial communities associated with the anthropogenic, highly alkaline environment of a saline soda lime, Poland. Antonie Van Leeuwenhoek 2017, 110, 945–962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freitas, D.B.; Lima-Bittencourt, C.I.; Reis, M.P.; Costa, P.S.; Assis, P.S.; Chartone-Souza, E.; Nascimento, A.M.A. Molecular characterization of early colonizer bacteria from wastes in a steel plant. Lett. Appl. Microbiol. 2008, 47, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Kay, R.T.; Duwelius, R.F.; Brown, T.A.; Micke, F.A.; Witt-Smith, C.A. Geohydrology, Water Levels and Directions of Flow, and Occurrence of Light-Nonaqueous-Phase Liquids on Ground Water in Northwestern Indiana and the Lake Calumet Area of Northeastern Illinois; Water-Resources Investigations Report; U.S. Geological Survey: Indianapolis, IN, USA, 1996; pp. 1–26/88.
- Colten, C.E. Industrial Wastes in the Calumet Area, 1869–1970 A Historical Geography; State Water Survey Division Report; Hazardous Waste Research and Information Center: Champaign, IL, USA; Available online: https://www.ideals.illinois.edu/handle/2142/1903 (accessed on 18 May 2019).
- Westphal, L.M.; Longoni, M.; LeBlanc, C.L.; Wali, A. Anglers’ Appraisals of the Risks of Eating Sport-Caught Fish from Industrial Areas: Lessons from Chicago’s Calumet Region. Hum. Ecol. Rev. 2008, 15, 46–62. [Google Scholar]
- Dourson, M.L.; Chinkin, L.R.; MacIntosh, D.L.; Finn, J.A.; Brown, K.W.; Reid, S.B.; Martinez, J.M. A case study of potential human health impacts from petroleum coke transfer facilities. J. Air Waste Manag. Assoc. 2016, 66, 1061–1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayless, E.R.; Greeman, T.K.; Harvey, C.C. Hydrology and Geochemistry of a Slag-Affected Aquifer and Chemical Characteristics of Slag-Affected Ground Water, Northwestern Indiana and Northeastern Illinois; Water-Resources Investigations Report; U.S. Geological Survey; Branch of Information Services [distributor]: Denver, CO, USA, 1998; pp. 1–4/67.
- Roadcap, G.S.; Kelly, W.R.; Bethke, C.M. Geochemistry of extremely alkaline (pH > 12) ground water in slag-fill aquifers. Ground Water 2005, 43, 806–816. [Google Scholar] [CrossRef] [PubMed]
- Roadcap, G.S.; Sanford, R.A.; Jin, Q.; Pardinas, J.R.; Bethke, C.M. Extremely alkaline (pH > 12) ground water hosts diverse microbial community. Ground Water 2006, 44, 511–517. [Google Scholar] [CrossRef]
- Suzuki, S.; Kuenen, J.G.; Schipper, K.; van der Velde, S.; Ishii, S.; Wu, A.; Sorokin, D.Y.; Tenney, A.; Meng, X.; Morrill, P.L.; et al. Physiological and genomic features of highly alkaliphilic hydrogen-utilizing Betaproteobacteria from a continental serpentinizing site. Nat. Commun. 2014, 5, 3900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woycheese, K.M.; Meyer-Dombard, D.R.; Cardace, D.; Argayosa, A.M.; Arcilla, C.A. Out of the dark: Transitional subsurface-to-surface microbial diversity in a terrestrial serpentinizing seep (Manleluag, Pangasinan, the Philippines). Front. Microbiol. 2015, 6, 44. [Google Scholar] [CrossRef] [PubMed]
- Meyer-Dombard, D.R.; Woycheese, K.M.; Yargıçoğlu, E.N.; Cardace, D.; Shock, E.L.; Güleçal-Pektas, Y.; Temel, M. High pH microbial ecosystems in a newly discovered, ephemeral, serpentinizing fluid seep at Yanartaş (Chimera), Turkey. Front. Microbiol. 2015, 5, 723. [Google Scholar] [CrossRef] [PubMed]
- Fones, E.M.; Colman, D.R.; Kraus, E.A.; Nothaft, D.B.; Poudel, S.; Rempfert, K.R.; Spear, J.R.; Templeton, A.S.; Boyd, E.S. Physiological adaptations to serpentinization in the Samail Ophiolite, Oman. ISME J. 2019, 1, 1750–1762. [Google Scholar] [CrossRef] [PubMed]
- Willems, A.; Busse, J.; Goor, M.; Pot, B.; Falsen, E.; Jantzen, E.; Hoste, B.; Gillis, M.; Kersters, K.; Auling, G.; et al. Hydrogenophaga, a New Genus of Hydrogen-Oxidizing Bacteria That Includes Hydrogenophaga flava comb. nov. (Formerly Pseudomonas flava), Hydrogenophaga palleronii (Formerly Pseudomonas palleronii), Hydrogenophaga pseudoflava (Formerly Pseudomonas pseudoflava and “Pseudomonas carboxydoflava”), and Hydrogenophaga taeniospiralis (Formerly Pseudomonas taeniospiralis). Int. J. Syst. Evol. Microbiol. 1989, 39, 319–333. [Google Scholar]
- Waska, K. Hyperalkaline Aquifers of Calumet Wetlands: Environmental Interactions between Geochemistry and Microbiology. Ph.D. Thesis, Northern Illinois University, DeKalb, IL, USA, 2014. [Google Scholar]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian Classifier for Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Lahti, L.; Shetty, S. Microbiome R Package. Available online: http://microbiome.github.com/microbiome (accessed on 16 June 2019).
- Vu, V.Q. Ggbiplot: A Ggplot2 Based Biplot. Available online: http://github.com/vqv/ggbiplot (accessed on 16 June 2019).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. Available online: https://CRAN.R-project.org/package=vegan (accessed on 15 June 2019).
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. J. Comput. Mol. Cell Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.-W.; Simmons, B.A.; Singer, S.W. MaxBin 2.0: An automated binning algorithm to recover genomes from multiple metagenomic datasets. Bioinformatics 2016, 32, 605–607. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. Barrnap v0. Available online: https://github.com/tseemann/barrnap (accessed on 22 January 2019).
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Frank, E.; Hall, M.; Holmes, G.; Kirkby, R.; Pfahringer, B.; Witten, I.H.; Trigg, L. Weka-A Machine Learning Workbench for Data Mining. Data Min. Knowl. Discov. Handb. 2009, 1269–1277. [Google Scholar]
- Becraft, E.D.; Dodsworth, J.A.; Murugapiran, S.K.; Ohlsson, J.I.; Briggs, B.R.; Kanbar, J.; De Vlaminck, I.; Quake, S.R.; Dong, H.; Hedlund, B.P.; et al. Single-Cell-Genomics-Facilitated Read Binning of Candidate Phylum EM19 Genomes from Geothermal Spring Metagenomes. Appl. Environ. Microbiol. 2016, 82, 992–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hierarchy Browser. Available online: http://rdp.cme.msu.edu/hierarchy/hb_intro.jsp (accessed on 27 June 2019).
- Home-Gene-NCBI. Available online: https://www.ncbi.nlm.nih.gov/gene/ (accessed on 27 June 2019).
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genomics 2008, 9, 75. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [PubMed]
- Baas-Becking, L.G.M. Geobiologie; of Inleiding tot de Milieukunde; WP Van Stockum & Zoon NV: Den Haag, The Netherlands, 1934. [Google Scholar]
- Schrenk, M.O.; Brazelton, W.J.; Lang, S.Q. Serpentinization, Carbon, and Deep Life. Rev. Mineral. Geochem. 2013, 75, 575–606. [Google Scholar] [CrossRef] [Green Version]
- Crespo-Medina, M.; Twing, K.I.; Sánchez-Murillo, R.; Brazelton, W.J.; McCollom, T.M.; Schrenk, M.O. Methane Dynamics in a Tropical Serpentinizing Environment: The Santa Elena Ophiolite, Costa Rica. Front. Microbiol. 2017, 8, 916. [Google Scholar] [CrossRef] [Green Version]
- Horikoshi, K. Alkaliphiles: Some Applications of Their Products for Biotechnology. Microbiol. Mol. Biol. Rev. 1999, 63, 735–750. [Google Scholar] [Green Version]
- Banciu, H.L.; Muntyan, M.S. Adaptive strategies in the double-extremophilic prokaryotes inhabiting soda lakes. Curr. Opin. Microbiol. 2015, 25, 73–79. [Google Scholar] [CrossRef]
- Detkova, E.N.; Boltyanskaya, Y.V. Osmoadaptation of haloalkaliphilic bacteria: Role of osmoregulators and their possible practical application. Microbiology 2007, 76, 511–522. [Google Scholar] [CrossRef]
- Brazelton, W.J.; Morrill, P.L.; Szponar, N.; Schrenk, M.O. Bacterial Communities Associated with Subsurface Geochemical Processes in Continental Serpentinite Springs. Appl. Environ. Microbiol. 2013, 79, 3906–3916. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Read Set | Total Sequence Input (bp) | Total Scaffold Length (bp) | Number of Scaffolds | Maximum Contig Length (bp) | N50 (bp) | L50 (bp) |
|---|---|---|---|---|---|---|
| S1S | 4,819,674,420 | 9,869,266 | 8842 | 70,488 | 4776 | 457 |
| S1W | 1,942,236,264 | 96,016,654 | 189,034 | 131,675 | 896 | 15,108 |
| S40cm | 6,321,349,632 | 27,714,568 | 7294 | 288,607 | 10,449 | 641 |
| S410cm | 1,138,541,340 | 4,063,552 | 2257 | 62,774 | 11,177 | 86 |
| S420cm | 8,551,723,860 | 33,109,460 | 39,264 | 137,610 | 3111 | 2415 |
| S435cm | 7,864,380,132 | 31,171,314 | 20,943 | 138,650 | 17,104 | 437 |
| S4S | 8,901,634,656 | 17,907,967 | 13,735 | 135,896 | 7433 | 522 |
| S4verticalbottom1 | 3,488,100,120 | 45,790,429 | 148,148 | 32,268 | 293 | 44,271 |
| S4verticaltop1 | 5,195,853,276 | 11,790,385 | 21,636 | 99,847 | 6486 | 372 |
| S4W | 781,642,320 | 91,790,252 | 237,098 | 42,452 | 363 | 58,467 |
| S5S | 7,256,817,336 | 766,453,974 | 1,650,212 | 594,813 | 488 | 338,228 |
| S5W | 1,891,476,984 | 216,488,361 | 534,113 | 123,446 | 404 | 124,669 |
| S7S | 3,027,277,992 | 293,538,298 | 692,250 | 139,485 | 450 | 164,345 |
| S7W | 1,202,217,492 | 4,759,255 | 17,534 | 36,970 | 259 | 6,864 |
| Sum | 62,382,925,824 | 1,650,463,735 | 3,582,360 | 2,034,981 | ||
| Average | 4,455,923,273.14 | 117,890,266.79 | 255,882.86 | 145,355.79 | 4549.21 | 54,063.00 |
| Read Set | Scaffold Bins | Detected 16S rRNA Genes | Chimeric Bins | Average Completeness | Average Genome Size | Average GC Content |
|---|---|---|---|---|---|---|
| S1S | 2 | 4 | 1 | 64.1% | 3,945,426 | 63.5% |
| S1W | 6 | 11 | 2 | 75.2% | 3,808,967 | 55.3% |
| S40cm | 4 | 12 | 1 | 36.5% | 3,072,671 | 59.7% |
| S410cm | 2 | 2 | 0 | 49.1% | 1,787,619 | 62.7% |
| S420cm | 3 | 19 | 1 | 50.2% | 4,217,172 | 56.3% |
| S435cm | 5 | 22 | 3 | 81.3% | 4,979,437 | 45.9% |
| S4S | 4 | 21 | 3 | 46.5% | 3,947,249 | 51.6% |
| S4verticalbottom1 | 2 | 7 | 2 | 62.6% | 1,663,564 | 50.2% |
| S4verticaltop1 | 2 | 10 | 1 | 74.3% | 4,420,534 | 54.1% |
| S4W | 4 | 8 | 3 | 68.2% | 2,177,976 | 48.3% |
| S5S | 23 | 42 | 4 | 72.9% | 3,830,319 | 53.9% |
| S5W | 6 | 10 | 1 | 58.6% | 2,675,264 | 46.0% |
| S7S | 4 | 9 | 2 | 75.0% | 4,072,426 | 46.4% |
| Sum | 67 | 177 | 24 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ohlsson, J.I.; Osvatic, J.T.; Becraft, E.D.; Swingley, W.D. Microbial Community in Hyperalkaline Steel Slag-Fill Emulates Serpentinizing Springs. Diversity 2019, 11, 103. https://doi.org/10.3390/d11070103
Ohlsson JI, Osvatic JT, Becraft ED, Swingley WD. Microbial Community in Hyperalkaline Steel Slag-Fill Emulates Serpentinizing Springs. Diversity. 2019; 11(7):103. https://doi.org/10.3390/d11070103
Chicago/Turabian StyleOhlsson, J. Ingemar, Jay T. Osvatic, Eric D. Becraft, and Wesley D. Swingley. 2019. "Microbial Community in Hyperalkaline Steel Slag-Fill Emulates Serpentinizing Springs" Diversity 11, no. 7: 103. https://doi.org/10.3390/d11070103
APA StyleOhlsson, J. I., Osvatic, J. T., Becraft, E. D., & Swingley, W. D. (2019). Microbial Community in Hyperalkaline Steel Slag-Fill Emulates Serpentinizing Springs. Diversity, 11(7), 103. https://doi.org/10.3390/d11070103