
Spatial Shifts in Species Richness in Response to Climate and Environmental Change: An Adaption of the EUROMOVE Model in the Czech Republic



Abstract
:1. Introduction
2. Materials and Methods
2.1. Species Data
2.2. Climate and Environmental Data
2.3. Spatial Models
2.4. Model Evaluation
2.5. Mean Stable Area Index (MSAi)
3. Results
3.1. Climate Indices
3.2. Model Performance
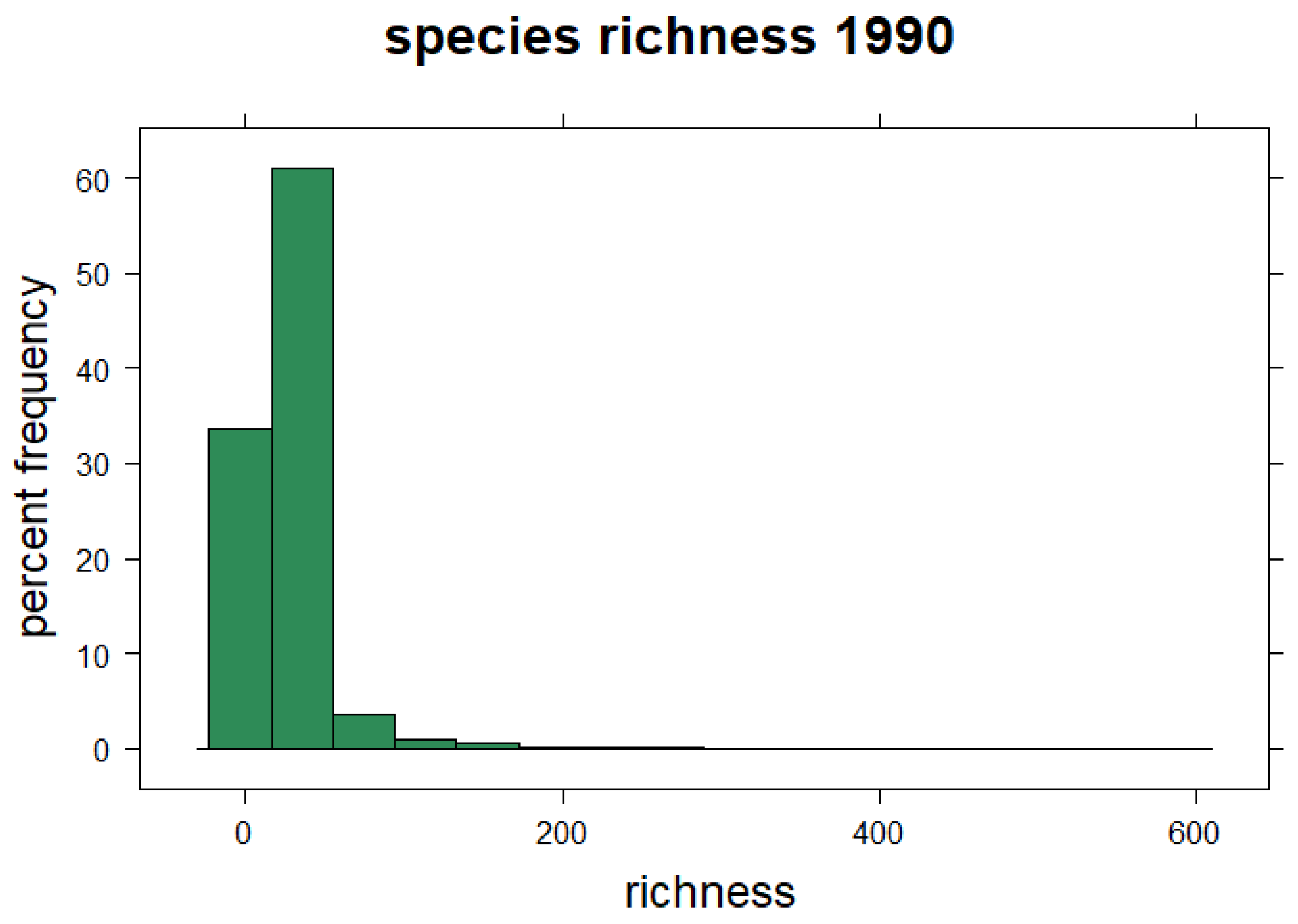
3.3. Species Richness and Distribution
3.4. Species Respond to Climate and Environmental Conditions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hlásny, T.; Holuša, J.; Štěpánek, P.; Turčáni, M.; Polčák, N. Expected impacts of climate change on forests Czech Republic as a case study. J. For. Sci. 2011, 57, 422–431. [Google Scholar] [CrossRef] [Green Version]
- Bakkenes, M.; Eickhout, B.; Alkemade, R. Impacts of different climate stabilisation scenarios on plant species in Europe. Glob. Environ. Chang. 2006, 16, 19–28. [Google Scholar] [CrossRef]
- Harrison, P.A.; Berry, P.M.; Butt, N.; New, M. Modelling climate change impacts on species’ distributions at the European scale: Implications for conservation policy. Environ. Sci. Policy 2006, 9, 116–128. [Google Scholar] [CrossRef]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.N.; Ferreira De Siqueira, M.; Grainger, A.; Hannah, L.; et al. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Bakkenes, M.; Alkemade, J.R.M.; Ihle, F.; Leemans, R.; Latour, J.B. Assessing effects of forecasted climate change on the diversity and distribution of European higher plants for 2050. Glob. Chang. Biol. 2002, 8, 390–407. [Google Scholar] [CrossRef]
- Pearson, R.G.; Dawson, T.P.; Berry, P.M.; Harrison, P.A. SPECIES: A spatial evaluation of climate impact on the envelope of species. Ecol. Model. 2002, 154, 289–300. [Google Scholar] [CrossRef]
- Huntley, B.; Berry, P.M.; Cramer, W.; McDonald, A.P. Modelling present and potential future ranges of some European higher plants using climate response surfaces. J. Biogeogr. 1995, 22, 967–1001. [Google Scholar] [CrossRef] [Green Version]
- Araújo, M.B.; Peterson, A.T. Uses and misuses of bioclimatic envelope modeling. Ecology 2012, 93, 1527–1539. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Pearson, R.G. Species’ Distribution Modeling for Conservation Educators and Practitioners. Synthesis. Am. Mus. Nat. Hist. 2007, 3, 1–50. [Google Scholar]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef] [PubMed]
- Guisan, A.; Zimmermann, N.E. Predictive habitat distribution models in ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Opdam, P.; Luque, S.; Jones, K.B. Changing landscapes to accommodate for climate change impacts: A call for landscape ecology. Landsc. Ecol. 2009, 24, 715–721. [Google Scholar] [CrossRef] [Green Version]
- Wilson, K.A.; Westphal, M.I.; Possingham, H.P.; Elith, J. Sensitivity of conservation planning to different approaches to using predicted species distribution data. Biol. Conserv. 2005, 122, 99–112. [Google Scholar] [CrossRef]
- Guisan, A.; Rahbek, C. SESAM—A new framework integrating macroecological and species distribution models for predicting spatio-temporal patterns of species assemblages. J. Biogeogr. 2011, 38, 1433–1444. [Google Scholar] [CrossRef] [Green Version]
- Ferrier, S.; Guisan, A. Spatial modelling of biodiversity at the community level. J. Appl. Ecol. 2006, 43, 393–404. [Google Scholar] [CrossRef]
- Barker, N.K.S.; Slattery, S.M.; Darveau, M.; Cumming, S.G. Modeling distribution and abundance of multiple species: Different pooling strategies produce similar results. Ecosphere 2014, 5, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.J.; Hijmans, R.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Maestre, F.T.; Quero, J.L.; Gotelli, N.J.; Escudero, A.; Delgado-baquerizo, M.; García-gómez, M.; Bowker, M.A.; Soliveres, S.; Escolar, C.; García-palacios, P.; et al. Europe PMC Funders Group Plant species richness and ecosystem multifunctionality in global drylands. Science 2012, 335, 214–218. [Google Scholar] [CrossRef] [Green Version]
- Tilman, D.; Reich, P.B.; Knops, J.M.H. Biodiversity and ecosystem stability in a decade-long grassland experiment. Nature 2006, 441, 629–632. [Google Scholar] [CrossRef]
- McCann, K.S. The diversity-stability. Nature 2000, 405, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Hooper, D.U.; Adair, E.C.; Cardinale, B.J.; Byrnes, J.E.K.; Hungate, B.A.; Matulich, K.L.; Gonzalez, A.; Duffy, J.E.; Gamfeldt, L.; Connor, M.I. A global synthesis reveals biodiversity loss as a major driver of ecosystem change. Nature 2012, 486, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Noss, R.F. Indicators for Monitoring Biodiversity: A Hierarchical Approach Published by: Wiley for Society for Conservation Biology Indicators for Monitoring Biodiversity: A Hierarchical Approach. Conserv. Biol. 1990, 4, 355–364. [Google Scholar] [CrossRef]
- Schipper, A.M.; Hilbers, J.P.; Meijer, J.R.; Antão, L.H.; Benítez-López, A.; de Jonge, M.M.J.; Leemans, L.H.; Scheper, E.; Alkemade, R.; Doelman, J.C.; et al. Projecting terrestrial biodiversity intactness with GLOBIO 4. Glob. Chang. Biol. 2020, 26, 760–771. [Google Scholar] [CrossRef] [Green Version]
- Alkemade, R.; Van Oorschot, M.; Miles, L.; Nellemann, C.; Bakkenes, M.; Ten Brink, B. GLOBIO3: A framework to investigate options for reducing global terrestrial biodiversity loss. Ecosystems 2009, 12, 374–390. [Google Scholar] [CrossRef] [Green Version]
- Scholes, R.J.; Biggs, R. A biodiversity intactness index. Nature 2005, 434, 45–49. [Google Scholar] [CrossRef]
- Loh, J.; Green, R.E.; Ricketts, T.; Lamoreux, J.; Jenkins, M.; Kapos, V.; Randers, J. The Living Planet Index: Using species population time series to track trends in biodiversity. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 289–295. [Google Scholar] [CrossRef] [Green Version]
- Ten Brink, B. Biodiversity Indicators for the OECD Environmental Outlook and Strategy; PBL Netherlands Environmental Assess-ment Agency: The Hague, The Netherlands, 2000; Volume 2004, pp. 22–23. [Google Scholar]
- Vačkář, D.; Ten Brink, B.; Loh, J.; Baillie, J.E.M.; Reyers, B. Review of multispecies indices for monitoring human impacts on biodiversity. Ecol. Indic. 2012, 17, 58–67. [Google Scholar] [CrossRef]
- Ten Brink, B.; Tekelenburg, T. Biodiversity: How Much Is Left? The Natural Capital Index Framework (NCI) 2002. Available online: https://unstats.un.org/unsd/envaccounting/seeaLES/egm/NKBio_bk2.pdf (accessed on 4 February 2022).
- EC Natura 2000. In European Commission DG Environment News Letter; European Commision: Brussels, Belgium, 2009; pp. 1–16.
- Čermák, P.; Mikita, T.; Trnka, M.; Štšpáneke, P.; Jurečka, F.; Kusbach, A.; Šebesta, J.A.N. Changes in climate characteristics of forest altitudinal zones within the Czech Republic and their possible consequences for forest species composition. Balt. For. 2018, 24, 234–248. [Google Scholar]
- Machar, I.; Vlckova, V.; Bucek, A.; Vozenilek, V.; Salek, L.; Jerabkova, L. Modelling of climate conditions in forest vegetation zones as a support tool for forest management strategy in European beech dominated forests. Forests 2017, 8, 82. [Google Scholar] [CrossRef] [Green Version]
- MacKu, J. Climatic characteristics of forest vegetation zones of the Czech Republic. J. Landsc. Ecol. 2015, 7, 39–48. [Google Scholar] [CrossRef] [Green Version]
- Araújo, M.B.; Alagador, D.; Cabeza, M.; Nogués-Bravo, D.; Thuiller, W. Climate change threatens European conservation areas. Ecol. Lett. 2011, 14, 484–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berry, P.M.; Dawson, T.P.; Harrison, P.A.; Pearson, R.; Butt, N. The sensitivity and vulnerability of terrestrial habitats and species in Britain and Ireland to climate change. J. Nat. Conserv. 2003, 11, 15–23. [Google Scholar] [CrossRef]
- Phillips, S.B.; Aneja, V.P.; Kang, D.; Arya, S.P. Modelling and analysis of the atmospheric nitrogen deposition in North Carolina. Int. J. Glob. Environ. Issues 2006, 6, 231–252. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S. “A Brief Tutorial on Maxent” in Species Distribution Modeling for Educators and Practitioners. Lessons Conserv. 2010, 3, 107–135. [Google Scholar]
- van Vuuren, D.P.; Edmonds, J.; Kainuma, M.; Riahi, K.; Thomson, A.; Hibbard, K.; Hurtt, G.C.; Kram, T.; Krey, V.; Lamarque, J.F.; et al. The representative concentration pathways: An overview. Clim. Chang. 2011, 109, 5–31. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Elith, J. [R Manual] Species distribution modeling with R Introduction. R Man. 2017, 71. [Google Scholar] [CrossRef] [Green Version]
- Cutler, D.R.; Edwards, T.C.; Beard, K.H.; Cutler, A.; Hess, K.T.; Gibson, J.; Lawler, J.J. Random forests for classification in ecology. Ecology 2007, 88, 2783–2792. [Google Scholar] [CrossRef]
- Breiman, L. Random forest. In Machine Learning; Statistics Department University of California: Berkeley, CA, USA, 2001; pp. 15–32. [Google Scholar]
- Guisan, A.; Edwards, T.C.; Hastie, T. Generalised linear and generalised additive models in studies of species distributions: Setting the scene. Ecol. Model. 2002, 157, 89–100. [Google Scholar] [CrossRef] [Green Version]
- McCullagh, P.; Nelder, J.A. Generalized Linear Models, 2nd ed.; Chapman & Hall: London, UK, 1989. [Google Scholar]
- Freeman, E.A.; Moisen, G.G. A comparison of the performance of threshold criteria for binary classification in terms of predicted prevalence and kappa. Ecol. Model. 2008, 217, 48–58. [Google Scholar] [CrossRef]
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Chytrý, M.; Kučera, T.; Kočí, M.; Grulich, V.; Lustyk, P. Habitat Catalogue of the Czech Republic (in Czech), 2nd ed.; Agentura Ochrany Přírody a Krajiny ČR: Prague, Czech Republic, 2010; ISBN 9788087457030. [Google Scholar]
- Lustyk, P. The Habitats Quality and Its Use for Assesment the Degree of Threat to Czech Flora Vascular Plant Species. Ph.D. Thesis, Mendel University in Brno, Brno, Czech Republic, 2021. (In Czech). [Google Scholar]
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Garcia-Gonzalo, J.; Seidl, R.; Delzon, S.; Corona, P.; Kolström, M.; et al. Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Raskin, P.D. Global scenarios: Background review for the Millennium Ecosystem Assessment. Ecosystems 2005, 8, 133–142. [Google Scholar] [CrossRef]
- Riahi, K.; Rao, S.; Krey, V.; Cho, C.; Chirkov, V.; Fischer, G.; Kindermann, G.; Nakicenovic, N.; Rafaj, P. RCP 8.5-A scenario of comparatively high greenhouse gas emissions. Clim. Chang. 2011, 109, 33–57. [Google Scholar] [CrossRef] [Green Version]
- Pearson, R.G.; Dawson, T.P.; Liu, C. Modelling species distributions in Britain: A hierarchical integration of climate and land-cover data. Ecography 2004, 27, 285–298. [Google Scholar] [CrossRef]
- Vellend, M.; Baeten, L.; Myers-Smith, I.H.; Elmendorf, S.C.; Beauséjour, R.; Brown, C.D.; De Frenne, P.; Verheyen, K.; Wipf, S. Global meta-analysis reveals no net change in local-scale plant biodiversity over time. Proc. Natl. Acad. Sci. USA 2013, 110, 19456–19459. [Google Scholar] [CrossRef] [Green Version]
- Pechanec, V.; Kilianová, H.; Tangwa, E.; Vondráková, A.; Machar, I. What is the development capacity for provision of ecosystem services in the Czech Republic? Sustainability 2019, 11, 4273. [Google Scholar] [CrossRef] [Green Version]
- Krkoška lorencová, E.; Harmáčková, Z.V.; Landová, L.; Pártl, A.; Vačkář, D. Assessing impact of land use and climate change on regulating ecosystem services in the czech republic. Ecosyst. Health Sustain. 2016, 2, e01210. [Google Scholar] [CrossRef]
- Pretzsch, H.; Schütze, G.; Uhl, E. Resistance of European tree species to drought stress in mixed versus pure forests: Evidence of stress release by inter-specific facilitation. Plant Biol. 2013, 15, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Coppola, E.; Nogherotto, R.; Ciarlo’, J.M.; Giorgi, F.; van Meijgaard, E.; Kadygrov, N.; Iles, C.; Corre, L.; Sandstad, M.; Somot, S.; et al. Assessment of the European Climate Projections as Simulated by the Large EURO-CORDEX Regional and Global Climate Model Ensemble. J. Geophys. Res. Atmos. 2021, 126, 1–20. [Google Scholar] [CrossRef]
- Leathwick, J.R.; Whitehead, D.; McLeod, M. Predicting changes in the composition of New Zealand’s indigenous forests in response to global warming: A modelling approach. Environ. Softw. 1996, 11, 81–90. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Source | Description | Original Scale |
|---|---|---|---|
| Species | Agency for Nature Conservation and Landscape Protection (OAPK) (http://www.ochranaprirody.cz/en/), accessed on (26 September 2019) | Raw survey data of over 3000 higher vascular plant species in the Czech Republic from 1960 to 1991 | 500 × 500 m |
| Climate 1 | Climate change in the Czech Republic (http://www.klimatickazmena.cz) accessed through Czechglobe (http://www.czechglobe.cz), on (1 May 2020) | Long-term means; 30 years for the historical climate and 20 years for the future climate. The model used is HadGEM2, scenario RCP 8.5 | 500 × 500 m |
| Topography (Slope) (Aspect) | The Czech Office for Surveying, Mapping, and Cadastre (https://ags.cuzk.cz/arcgis2/services/dmr5g/ImageServer) accessed on (15 June 2021) | 5 m | |
| Soil (Soil texture) (Soil depth) | Research Institute for Soil and Water Conservation + Forest Management Institute (2018) | Soil associations, on an agricultural soil map of rated soil-ecological units | 1:10,000 |
| Hydrology (drainage) | Research Institute for Soil and Water Conservation + Forest Management Institute (2018) | Combination of soil infiltration ability, group of forest types and maps of soil associations | 1:10,000 |
| Distance to rivers | Open street map (OSM) | 10 or 100 m distance from the river | |
| Vegetation quality (Plant cover) | CzechGlobe + ESA + Palacký University | 1:10,000 | |
| Geology | Czech Geological Survey (Geological maps) | Geological materials | 1:100,000 |
| Anrain | Tcold | Tempgs | Lenvegt | |||||
|---|---|---|---|---|---|---|---|---|
| Min | Max | Min | Max | Min | Max | Min | Max | |
| 1990 | 445 | 1345 | −8.0 | −2.9 | 9.1 | 13.8 | 151 | 240 |
| 2018 | 352 | 1713 | −8.3 | −3.3 | 9.6 | 15.0 | 116 | 281 |
| 2060 | 457 | 1541 | −6.8 | 0.7 | 10.4 | 15.0 | 154 | 283 |
| 2100 | 480 | 1617 | −4.1 | 3.6 | 12 | 15.8 | 173 | 324 |
| Min | Mean | Max | |
|---|---|---|---|
| AUC | 0.43 | 0.80 | 0.99 |
| TSS | 0.026 | 0.43 | 0.83 |
| Classification Methods | ||||
|---|---|---|---|---|
| Evaluation Metrics | Species | Maxent | Random Forest | GLM (Logistic) |
| Alnus sp., n = 544 sites, subspecies = 2 | ||||
| AUC | 0.80 | 0.79 | 0.70 | |
| TSS | 0.54 | 0.46 | 0.34 | |
| Fagus syvestica L., n = 884 sites, subspecies = 1 | ||||
| AUC | 0.79 | 0.82 | 0.80 | |
| TSS | 0.52 | 0.50 | 0.44 | |
| Festuca sp., n = 9031 sites, subspecies = 11 | ||||
| AUC | 0.64 | 0.78 | 0.62 | |
| TSS | 0.62 | 0.41 | 0.17 | |
| Picea abie, n = 16,301 sites, subspecies = 1 | ||||
| AUC | 0.73 | 0.75 | 0.73 | |
| TSS | 0.58 | 0.39 | 0.34 | |
| Poa sp., n = 6215 sites, subspecies = 6 | ||||
| AUC | 0.66 | 0.80 | 0.58 | |
| TSS | 0.59 | 0.44 | 0.13 | |
| Quercus sp., n = 4939 sites, subspecies = 3 | ||||
| AUC | 0.76 | 0.82 | 0.75 | |
| TSS | 0.57 | 0.50 | 0.38 | |
| Rubus sp., n = 16,899 sites, subspecies = 4 | ||||
| AUC | 0.69 | 0.80 | 0.69 | |
| TSS | 0.58 | 0.47 | 0.28 | |
| Salix sp., n = 617 sites, subspecies = 2 | ||||
| AUC | 0.76 | 0.79 | 0.71 | |
| TSS | 0.50 | 0.47 | 0.29 | |
| Modelling Period | Mean Area (km2) | Species Number | Species Lost | Estimated MSAi |
|---|---|---|---|---|
| 1990 | 22,194 | 686 | - | - |
| 2018 | 23,746 | 675 | 11 | ~1.0 |
| 2060 | 11,544 | 661 | 26 | 0.50 |
| 2100 | 12,021 | 548 | 140 | 0.43 |
| Species | Net Change (%) 2018 a –1990 |
|---|---|
| Alnus sp. | −2 |
| Fagus syvestica L. | +10 |
| Festuca. sp. | −1 |
| Picea abie | +42 |
| Poa sp. | +6 |
| Quercus sp. | +5 |
| Rubus sp. | +9 |
| Salix sp. | +26 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tangwa, E.; Pechanec, V.; Brus, J.; Vyvlecka, P. Spatial Shifts in Species Richness in Response to Climate and Environmental Change: An Adaption of the EUROMOVE Model in the Czech Republic. Diversity 2022, 14, 235. https://doi.org/10.3390/d14040235
Tangwa E, Pechanec V, Brus J, Vyvlecka P. Spatial Shifts in Species Richness in Response to Climate and Environmental Change: An Adaption of the EUROMOVE Model in the Czech Republic. Diversity. 2022; 14(4):235. https://doi.org/10.3390/d14040235
Chicago/Turabian StyleTangwa, Elvis, Vilem Pechanec, Jan Brus, and Pavel Vyvlecka. 2022. "Spatial Shifts in Species Richness in Response to Climate and Environmental Change: An Adaption of the EUROMOVE Model in the Czech Republic" Diversity 14, no. 4: 235. https://doi.org/10.3390/d14040235
APA StyleTangwa, E., Pechanec, V., Brus, J., & Vyvlecka, P. (2022). Spatial Shifts in Species Richness in Response to Climate and Environmental Change: An Adaption of the EUROMOVE Model in the Czech Republic. Diversity, 14(4), 235. https://doi.org/10.3390/d14040235