

Changing Trends in Cetacean Strandings in the East China Sea: Identifying Relevant Variables and Implications for Conservation and Management

1
College of Marine Sciences, Shanghai Ocean University, Shanghai 201306, China
2
East China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Yangpu District, Shanghai 200090, China
3
Key Laboratory of East China Sea Fishery Resources Exploitation, Ministry of Agriculture and Rural Affairs, Shanghai 200090, China
*
Author to whom correspondence should be addressed.
Diversity 2023, 15(10), 1082; https://doi.org/10.3390/d15101082
Submission received: 14 September 2023
/
Revised: 9 October 2023
/
Accepted: 11 October 2023
/
Published: 13 October 2023
Abstract
:The frequency of cetacean strandings is increasing, with multiple causes identified. We examined temporal and spatial trends in stranding numbers along the coastal areas of the East China Sea from 1990 to 2021. Using structural equation models, we analyzed the relationships between strandings and climate, oceanic properties, and human activities. Our findings revealed time-dependent interannual variability in strandings but no linear trend, indicating relative stability in cetacean strandings. Seasonal patterns were observed only in narrow-ridged finless porpoises (Neophocaena asiaeorientalis), with significant spring increases (March to May). Clustering of strandings occurred in central Fujian and northern/southern Taiwan, according to spatial analysis. The impact of variables on cetacean strandings varied across time periods. Storm surges, coastal fishing, and the Pacific decadal oscillation were associated with stranding events from 1990 to 2006. However, from 2006 to 2021, the influence of storm surges weakened, while the climate index only indirectly affected strandings through sea surface temperature and salinity, with an increased intensity in the effects of winter sea surface temperature and salinity. Structural equation models unveiled the cascading effects of environmental changes on strandings. This study reports changing trends in cetacean strandings and identifies relevant variables. Although not exhaustive, understanding the reasons behind strandings enhances our comprehension of cetacean responses to environmental changes, supporting targeted conservation and management efforts.
1. Introduction
Cetaceans serve as essential sentinel species for evaluating the health of marine ecosystems due to their long lifespan, high trophic level, and significant ecological roles, including their impact on prey population dynamics, nutrient cycling, and ecosystem structure [1,2]. As top predators in marine ecosystems, cetaceans affect the population status and evolutionary dynamics of prey organisms. Additionally, their extensive feeding on marine organisms contributes to the bioaccumulation of pollutants, reflecting changes in the ecological environment [3,4,5]. This is particularly relevant for species inhabiting the continental shelf regions, as they are directly impacted by human activities, and their abundance and distribution serve as ecological indicators in environmental assessments [6]. Consequently, cetacean conservation often constitutes a fundamental objective in establishing marine protected areas [7,8]. Understanding the habitat utilization patterns of cetaceans is crucial for effective conservation measures [9] and provides insights into prey availability and environmental changes within marine ecosystems [10,11,12]. It is difficult to conduct comprehensive research on the majority of cetaceans due to their widespread distribution, extensive migrations, low population densities, and prolonged periods spent below the ocean’s surface. Furthermore, the lengthy lifespan of cetaceans necessitates long-term, continuous monitoring to observe population changes throughout their entire life cycles. This requires substantial financial support and policy attention, which are frequently deficient in developing nations and regions. Common monitoring techniques include visual and acoustic surveys, both of which have limitations such as high survey costs, demanding sea conditions, and effectiveness only for specific species [13,14]. For instance, visual surveys are particularly useful for documenting visually conspicuous and easily observed species such as bottlenose dolphins (Tursiops truncatus). Widespread on a global scale, strandings have been used for a long time as a relatively low-cost method to monitor marine mammal populations [15,16], providing fundamental information on species composition and relative abundance [17,18,19]. Since the last century, long-term monitoring and investigations of cetacean strandings have been conducted in countries such as the United Kingdom [20], Australia [21,22], the United States [23], and New Zealand [16]. For instance, the cetacean strandings investigation program in the United Kingdom has documented nearly 20,000 stranding events, establishing one of the world’s most extensive and long-term datasets on cetacean strandings and enabling valuable insights into population trends, stranding causes, and conservation strategies.
Cetacean strandings are the result of the combined effects of multiple factors [24]. Cetacean stranding patterns vary depending on species composition and population abundance, resulting in species-specific distinctions in stranding frequency and spatial-temporal distribution [25,26,27]. Moreover, climate change influences the utilization of cetacean habitats, prompting some species to depart from their native habitats and resulting in an increase in stranding rates in particular regions [28,29,30]. The magnetic sensitivity of cetaceans plays a vital role in their migration behavior, and disturbances in the Earth’s magnetic field caused by solar storms can disorient cetaceans, hindering their timely departure from hazardous areas and increasing the likelihood of stranding events [31,32]. Post-mortem examinations have revealed that a substantial portion of stranded cetaceans are the result of anthropogenic causes such as ship collisions, fisheries interactions, marine pollution, and naval exercises [33,34,35,36,37].
The East China Sea, rich in resources and characterized by a developed coastal economy, has experienced significant impacts on its marine ecosystem in recent decades due to intensive human activities and climate change [38]. Chinese waters are home to approximately 39% of the global cetacean population, and the East China Sea harbors a substantial proportion of China’s cetacean species [39,40]. Following notable revisions to the wildlife protection law of the People’s Republic of China in 2018, subsequent revisions to the list of key protected wild animals in China have resulted in more cetaceans being listed as the Grade I National Key Protected Animal Species in China [41]. In addition, the Ministry of Agriculture and Rural Affairs of the People’s Republic of China issued a notice in 2021 emphasizing the critical significance of proactive cetacean conservation management [42]. The investigation of cetacean strandings holds particular relevance for countries and regions that are relatively new to this field of study [43]. While regional (e.g., Hainan, Hong Kong, Taiwan) and national stranding databases exist, as well as studies on the spatio-temporal distribution patterns of strandings, there remains a dearth of research and analysis concerning the underlying causal factors driving cetacean stranding events [40,44,45]. The study seeks to uncover the interannual and seasonal variations and spatial patterns of cetacean strandings along the eastern coast of China, as well as to analyze the drivers of spatio-temporal changes and the disturbance patterns of various variables on strandings. This research contributes to the improvement of cetacean conservation policies and a greater comprehension of coastal ecosystems through its findings.
2. Materials and Methods
2.1. Study Area and Stranding Data
The study area included two parts of China’s waters, including the East China Sea. This article documents stranding events in five administrative regions: Jiangsu (JS), Shanghai (SH), Zhejiang (ZJ), Fujian (FJ), and Taiwan (TW) (Figure 1). We compiled records of cetacean strandings from 1990 to 2021, primarily from news articles, research papers, and other pertinent publications. Each stranding record consists of the date of stranding, location, and species. We initiated our data collection process by conducting systematic searches using keywords such as “whale”, “cetacean”, “dolphin”, and “porpoise” across various search engines, online English literature databases, as well as online Chinese literature databases. Subsequently, we refined our search results by incorporating additional keywords such as “stranding” to specifically target stranding records. These records were then compiled and aggregated, focusing on the coastal areas of the East China Sea. Furthermore, we complemented our dataset by extracting pertinent information from books and reports, as well as incorporating insights derived from reports submitted by residents and local communities. Based on search results, a rigorous selection process was applied to all available records, followed by meticulous manual examination. We conducted a comprehensive validation of the collected cetacean species records, with a primary focus on assessing the reliability of their sources and verifying the consistency of reported species occurrences with existing research findings. Records were scrutinized for factors such as stranding dates, locations, and cetacean species. Non-specialized, duplicate, irrelevant, and problematic entries were systematically excluded from the final dataset.
2.2. Environmental Data
In our analysis, we considered various factors associated with cetacean strandings (Table 1). As a key climatic indicator, the present study employs the Pacific decadal oscillation (PDO) index, which represents the average monthly value throughout the year. Another significant climatic variable considered is the East Asian summer monsoon index (EASMi), which quantifies the strength of the summer monsoon in the East Asian region by calculating the annual deviation from the mean monsoon index within the specified area. In terms of the fishing industry, the analysis focuses on the landed quantities from Jiangsu, Zhejiang, Shanghai, Fujian, and Taiwan. Trawls, purse seines, gillnets, and trap nets are the four principal fishing equipment used in the region.
2.2.1. The Influence of Climate on Cetacean Strandings
It has been documented that climate factors influence the distribution and abundance of prey organisms for cetaceans [46]. These factors contribute to the alteration of cetacean distribution and the increase of strandings [47]. Coastal storm surges have direct detrimental effects on whales, causing physical harm and disorientation. Another factor to consider is that storm surges can significantly impact whether a deceased or weakened cetacean ends up stranded on the shoreline. Additionally, similar climatic phenomena disrupt the availability of prey organisms, consequently impacting cetaceans [48,49]. The PDO, recognized as one of the primary mechanisms of decadal-scale climate variability, exerts substantial effects on global climate and ecosystems. Its effects are primarily observed through variations in sea surface temperature, salinity, and nutrient concentrations [46]. Notably, studies have demonstrated a significant contemporaneous negative correlation between the catch of economically important fish species in the East China Sea and PDO variations [50]. These findings emphasize the significance of incorporating the PDO into fisheries management and stranding analyses.
2.2.2. The Influence of Oceanic Properties on Cetacean Strandings
The temperature, salinity, and other oceanic properties of seawater have direct or indirect effects on the foraging, reproduction, and migration of cetaceans, resulting in alterations to their distribution [51] and consequently affecting stranding spatial patterns. Significant increases in precipitation or environmental pollution can lead to a sustained decrease in seawater salinity, resulting in higher disease prevalence among cetacean populations. This elevated disease occurrence, in turn, indirectly contributes to the incidence of strandings [52,53,54]. Moreover, rising sea temperatures contribute to the extended persistence of deleterious algal blooms, highlighting the significance of this ecological relationship [55].
2.2.3. The Influence of Human Activities on Cetacean Strandings
Cetacean distribution is intricately correlated with the availability of prey organisms. Examining stranded whale carcasses has revealed that food scarcity and the accumulation of seawater contaminants contribute to cetacean strandings [56]. The depletion of prey organisms for cetaceans as a result of overfishing has had detrimental effects on their survival and well-being [57]. Furthermore, the power of fishing vessels serves as the most direct measure of fishing effort, providing a more immediate reflection of the impact of fishing vessels on cetaceans beyond their fishing activities. For instance, during navigation, fishing vessels may accidentally collide with cetaceans or generate noise that affects them. Additionally, an increase in the power of fishing vessels can also lead to a higher number of abandoned fishing gears, posing a direct threat to cetaceans and leading to mortalities [58]. In our study, we use the power of fishing vessels and the yearly fishing catch of fishing vessels as indicators of human activities, allowing us to evaluate the level of disturbance caused to cetaceans.
2.3. Data Analysis
2.3.1. Temporal and Spatial Analyses
In order to examine the temporal patterns of cetacean strandings, seasonal and annual trends were analyzed based on the recorded counts of all stranding events. In order to verify the stationarity of the cetacean stranding data and that the mean and variance of the time series are constant over time, we conducted the augmented Dickey–Fuller test using the ‘tseries’ package [59] in the statistical software R [60]. In addition, the presence of monotonic trends in strandings throughout the 31-year time series (1990–2021) was assessed using the Mann–Kendall trend test implemented in the ‘trend’ package [61]. Furthermore, analyses of the seasonal difference were investigated with the Kruskal–Wallis [62] test using the software R for the entire dataset and the two most commonly reported families, Balaenopteridae and Phocoenidae. The seasons were defined as follows: spring (March to May), summer (June to August), autumn (September to November), and winter (December to February).
We used Moran’s I index to compute spatial autocorrelation [63]. This approach assesses whether stranding locations were dispersed, random, or clustered. Once the stranding patterns were determined, Gi* hotspot analysis was used to identify stranding hotspots [64]. On the basis of the calculated p-values and z-scores, the spatial clustering locations of high- and low-value factors can be determined. All spatial analyses were undertaken in ArcGIS 10.8.
2.3.2. Structural Equation Model
To examine the factors contributing to variations in cetacean strandings, we used structural equation models (SEM) to examine the correlation between key factors and interannual changes in strandings. A structural equation represents the causal relationships among variables, with a basic example being a univariate linear equation. SEM, as an analytical method, combines path analysis and confirmatory factor analysis to display the results of structural equations [65]. By integrating these methods, SEM allows for the examination of relationships between endogenous variables and exogenous variables [66], thereby revealing direct and indirect causal connections among multiple variables.
The implementation of the structural equation model in this study was based on the “piecewise SEM” package in the statistical software R, developed by Lefcheck in 2015. Unlike conventional SEM, this package employs local estimation rather than global estimation, enabling the analysis of each path individually [67]. The SEM was used to examine the impact of interannual changes in various variables on cetacean strandings [68]. Initially, an a priori model was developed based on previous research findings. The Fisher’s C test [67] was used for model identification to assess potential issues of overfitting or inadequate fit. Additionally, the directed separation test [69] was used to determine the conditional independence of variables. If the model fit was inadequate, significant missing paths (i.e., p < 0.05) with the smallest p-values were incorporated into the model via directed separation, and this procedure was repeated until an adequate model fit was achieved [67]. In cases where multiple models passed the Fish’s C test, the final model was selected based on the comparison of AIC values.
To account for the extensive temporal span of our study and the observable increase in fish catch in the eastern coastal waters of China from the 1990s to the early 21st century, followed by a subsequent plateau, the dataset was divided into two distinct time periods, using 2006 as the dividing line. For each period, separate structural equation models were developed to assure a comprehensive analysis. Through a series of iterations and refinements, we derived the final model, which exhibited a satisfactory level of goodness-of-fit. Specifically, considering that the annual average temperature may not adequately capture temperature variations, we compared the goodness of fit of the average temperature of the highest monthly average temperature and the average daily temperature during the winter months (December to February) within the model. Notably, the average daily temperature during the winter demonstrated the strongest performance. Recognizing the time-lagged impact of the PDO on marine ecosystems [70,71], we discovered that the PDOwin 2 yr-lag had stronger explanatory power after fitting several models with various delays of years. As a result, we incorporated the two-year-lagged winter PDO into the revised model. In an a priori model, the relationship between fish catch and strandings of cetaceans was not statistically significant. By disaggregating fish catch into power of fishing vessel and yearly fishing catch, we were able to identify a significant positive effect of catch on stranding events. To address the issue of collinearity between fishing vessel power and cetacean strandings, a direct relationship was established between the two variables. Ultimately, the final model demonstrated a favorable fit (p = 0.74). Furthermore, despite our initial expectation that PDO would not exert a direct influence on stranding occurrences, the results of the direct separation test revealed that this relationship was not conditionally independent, as predicted by an a priori model. Consequently, we integrated a direct effect of PDO into the structural equation model encompassing 1990 to 2006.
3. Results
During the period from 1990 to 2021, 239 cetacean stranding records were obtained and involved 27 cetacean species belonging to seven families [39]. The majority of the strandings belonged to three prominent families, namely Balaenopteridae, Delphinidae, and Phocoenidae, accounting for a substantial proportion of the overall strandings, specifically 46, 96, and 46 events, respectively, which collectively accounted for approximately 77% of cetacean stranding events (Table 2). The narrow-ridged finless porpoise (Neophocaena asiaeorientalis) was the most frequently recorded species, accounting for approximately 16% of cetacean stranding events. Intriguingly, an overwhelming majority of these porpoise strandings, approximately 70%, were recorded in Zhejiang Province (Figure 2). Temporally, cetacean strandings were observed annually throughout the entirety of the study period, with a notable peak in stranding events documented in 2008 and the highest number of cetacean stranding individuals observed in 2021 (Figure 2). Geographically, Taiwan had the highest number of stranding records, accounting for approximately 42% of cetacean stranding events, followed by Zhejiang (21%), and Fujian (21%), which also made significant contributions to the cetacean stranding events.
3.1. Temporal Patterns
The occurrence of whale strandings exhibited a discernible upward trend between 2000 and 2008, with the highest number of strandings recorded in 2008 (Figure 2). The Augmented Dickey–Fuller test was conducted to evaluate the stationarity of the time series, and the results indicated non-significance (p = 0.67), suggesting a non-stationary pattern. The Mann–Kendall trend test did not detect any significant upward or downward trend in the annual occurrence of strandings (p = 0.93). And the Kruskal–Wallis test revealed no significant seasonal differences (Kruskal–Wallis chi-squared = 1.9721, df = 3, p-value = 0.5782). Subsequently, a more concentrated investigation was conducted on families with an increased number of stranding records. Due to the considerable species diversity and ecological variations within the family Delphinidae [72], seasonal differences within this family were not examined (Figure 3). There were no significant seasonal differences in strandings within Family Balaenopteridae (Kruskal–Wallis chi-squared = 2.3792, df = 3, p-value = 0.4975). In contrast, significant seasonal differences were identified within Family Phocoenidae (Kruskal–Wallis chi-squared = 10.909, df = 3, p-value = 0.01223), with significant spring increases.
3.2. Spatial Patterns
Observations of cetacean strandings along the coasts of Jiangsu (JS), Shanghai (SH), Zhejiang (ZJ), Fujian (FJ), and Taiwan (TW) between 1990 and 2021 indicate a pervasive occurrence of stranding events in these regions. Notably, the frequency of stranding was highest in the northern and southern regions of Taiwan, whereas Jiangsu had considerably fewer records (Figure 4). We analyzed the spatial distribution pattern of cetacean strandings using the Global Moran’s I index, which indicates a statistically significant clustered pattern. (Z-score = 48.42 and p-value < 0.001). The Gi* hotspot analysis provided a statistical way to identify stranding clusters with significance levels. The central regions of Fujian and the northern and southern regions of Taiwan exhibited a high level of significance at the 99% confidence level, indicating the presence of significant stranding hotspots. In addition, the coastal areas of central Fujian and the southern end of Zhejiang were identified as significant hotspot regions for cetacean strandings at the 95% confidence level (Figure 5).
3.3. Factors Driving Patterns of Cetacean Strandings
Piecewise SEM was utilized to further elucidate the direct and indirect pathways of factors influencing cetacean strandings. The variables incorporated in the model collectively explained a large proportion of the variation in cetacean strandings. Moreover, the analysis of the graph revealed discernible variations in the strength of external influences on strandings across different temporal intervals. However, it is noteworthy that sea level pressure did not exhibit a significant influence on storms during the entire study period, consequently lacking an indirect impact on cetacean strandings. All of these variables explained 59% and 65%, respectively, of the variations in strandings during the two time periods (Figure 6). Specifically, from 1990 to 2006, the incidence of coastal storm surges, fishery capture, and the PDOwin 2 yr-lag were found to have direct and statistically significant effects on strandings, as indicated by standardized coefficients of 0.59, 0.6861, and −0.5933, respectively. Furthermore, the EASMi factor was observed to indirectly affect strandings by influencing the frequency of coastal storm surges. In addition, the rising total power of fishing vessels contributed to an increase in fishing pressure, thereby promoting strandings.
In the subsequent timeframe of 2007 to 2021, salinity and winter sea surface temperature emerged as significant factors influencing cetacean strandings, with standardized path coefficients of −0.4345 and −0.5635, respectively. In spite of the absence of direct effects of the PDOwin 2-year lag on strandings during this period, it was found to impart indirect influences via its association with sea surface temperature and salinity. Notably, the PDOwin 2 yr-lag exhibited a positive effect on temperature but a negative effect on salinity. By contrasting the cumulative effects of the two pathways through meticulous calculations, it is possible to conclude that the PDOwin 2 yr-lag had an overall negative impact on the incidence of strandings.
4. Discussion
4.1. Temporal and Spatial Trends
The temporal and spatial variations of cetacean strandings are reflective of the status of species, seasonal distribution, and population changes to a significant degree [73]. Analyzing stranding events is crucial for cetacean research. Temporal analyses of stranding events in this study have revealed interannual fluctuations but no linear trend. In the present study, the significant increase in stranding events between 2000 and 2008 is likely attributable, in part, to increased media coverage and public awareness. The seasonal analysis reveals that the overall distribution of cetacean strandings, as well as those within the Balaenopteridae family, exhibits a statistically insignificant pattern of seasonal uniformity. Nevertheless, this discovery contradicts existing perspectives. Due to the relatively limited sample size obtained in this study, all cetacean species were pooled together for seasonal analysis to examine the overall seasonality of cetacean strandings. Therefore, the difficulty in detecting seasonality across all cetacean species may be attributable to the wide variety of included species, each of which possesses distinct ecological characteristics. For instance, the family Phocoenidae experiences a surge in strandings during the spring in Indonesia [62], whereas certain species within the family Delphinidae experience a peak in strandings during the months of March and April in the UK [58]. In contrast, strandings of the family Ziphiidae are more common in autumn in China, Australia, and the Northeast Pacific [40,74,75]. Consequently, an overarching seasonal trend is not discernible. Due to its limited presence along the Chinese coast, the Balaenopteridae family displays a relatively uniform distribution of strandings throughout the year. Another reason is the presence of six species within the Balaenopteridae family in the East China Sea (Table 2). Due to differences in their ecological behaviors, it is challenging to monitor their seasonality. Thus, the sample size derived from strandings is small, and the occurrence appears to be random. Population migrations [22,76], temporal variations in prey distribution [77], climate change [78], and seasonal coastal human activities such as recreational pursuits and fishing operations [58,79] may contribute to significant seasonality in strandings. Notably, the narrow-ridged finless porpoise (Neophocaena asiaeorientalis) exhibits statistically significant seasonality in strandings during spring. This species is commonly found along the Chinese coast [80], and its reproductive season in the East China Sea occurs from April to May [81]. During this time, the cetacean population increases and moves closer to the coast, increasing the likelihood of strandings. Additionally, the finless porpoise demonstrates nurturing behavior, with young calves often tightly clinging to their mother’s dorsal fins in nearshore coastal areas, prolonging their proximity to the shore [82]. Therefore, another reason may be due to increased calves stranding [30]. The peak period of porpoise strandings coincides with the increased incidence of fishing and incidental injury events involving this species in the study area [83], indicating that fishing may also contribute to the seasonality of strandings. Nevertheless, thorough necropsies are required to corroborate the cause of death.
Based on the records, there are sporadic reports of cetacean strandings along the coasts of the study region, and the hotspots for stranding are situated on both sides of the Taiwan Strait. Similar to interannual variations in strandings, the differential attention given to these events by different governments and stranding organizations is a factor in the formation of spatial hotspots [84]. Regions with well-established networks tend to report more strandings due to efficient data collection, faster response times, and increased research efforts. In contrast, areas lacking such networks may have fewer reported cases, contributing to apparent spatial variations in cetacean strandings. In addition, numerous factors, such as species resources, climate conditions, fisheries, and maritime traffic in different regions, may have direct or indirect effects on strandings. The study area, which is characterized by numerous river estuaries, dense islands and reefs, and multiple cold and warm coastal currents, not only sustains an abundance of fishery resources [85], but also a great diversity of cetacean species. Therefore, the natural conditions in the Taiwan Strait make it a suitable habitat for cetaceans. It is noteworthy that roughly half of the cetacean species documented in Chinese waters are located within the Taiwan Strait [86]. This region’s elevated species diversity contributes to the frequency of cetacean strandings [22]. In addition, the Taiwan Strait is a vital passage connecting the East China Sea and the South China Sea, facilitating maritime commerce between East Asia, Southeast Asia, and the Indian Ocean’s coasts. The resulting dense and complex maritime traffic poses a direct threat to the survival and growth of cetaceans. According to necropsy results, fishing and marine traffic are causing a significant number of whale strandings in Taiwan’s waters [87]. In addition, a growing body of evidence suggests a link between cetacean strandings and human activities [12,88].
4.2. Correlates of Strandings through Time
To investigate variations in strandings in greater depth, we analyzed multiple variables and divided the study period into two distinct phases: 1990–2006 and 2007–2021. The significance of the influencing elements varied significantly between the two models. In the 1990–2006 model, storm frequency, the PDOwin 2-yr-lag, and fishery catch had a discernible effect on strandings, whereas sea surface temperature and salinity did not manifest a statistically significant effect. However, a subsequent analysis for the period 2007 to 2021 revealed a correlation between strandings and both sea surface temperature and salinity, whereas storm frequency and fishery catch exhibited no significant association. These findings suggest that the factors influencing strandings have changed over time. The following sections will conduct a thorough analysis of each individual factor.
The disruptive forces of strong storm surges and tumultuous oceanic conditions, which can affect the likelihood of cetacean strandings, are known to play a significant role in facilitating strandings during storm events. Consequently, a large number of cetacean strandings are frequently recorded during storms [23,89]. Notably, smaller coastal cetaceans are especially susceptible to the negative effects of cyclones because their limited mobility makes evading turbulent conditions difficult and makes them more susceptible to disturbance [47]. Intriguingly, our study reveals that the family Delphinidae comprises the most commonly stranded species, predominantly inhabiting sloping areas and coastal margins, which corresponds to the observed stranding patterns. In addition, cyclones and surges can contribute to the presence of whale carcasses near the coast, thereby increasing the number of recordings [89,90]. In addition, storm surges can influence the availability of prey by modifying various physicochemical properties of the water column, such as disturbances within the mixing layer and temperature and salinity fluctuations. Consequently, such alterations can have a significant impact on the distribution and abundance of phytoplankton and zooplankton, influencing the food chain and decreasing the availability of food resources for cetaceans [91,92]. These findings highlight the importance of storm surges as key environmental factors that influence the occurrence of cetacean strandings. Consequently, it is essential to closely monitor and investigate trends in storm surges in order to facilitate the development of more effective conservation and management strategies for cetaceans.
Our analysis reveals a direct relationship between fish catch and strandings during the first time period. It was discovered that the magnitude of fishing activities correlates with the frequency of cetacean strandings. However, the subsequent analysis for the period 2007–2021 did not reveal any statistically significant association between fish catch and strandings. This disparity may be attributed to the relatively stable levels of inshore landings during the latter time period. Another plausible explanation could be the variation in prey organisms among various cetacean species, and the documented fish catch incorporates the totality of nearshore fishing and is therefore incapable of capturing the interannual variations in prey organisms targeted by cetaceans [57]. To conduct an in-depth investigation into the effect of exploitation on cetacean strandings, the scope of the study must be restricted to cetacean species with comparable ecological behaviors. In addition, it is crucial to select primary onshore prey landing data in order to improve the accuracy and dependability of the results.
Based on stranding reports in the Atlantic and Mediterranean, malnutrition is the most common cause of cetacean strandings, and the increasing fish catch has a direct effect on the availability of food resources [57,93]. The depletion of resources forces whales to seek alternative food sources, causing some species to migrate to shallower water areas with greater resource abundance, thereby increasing the stranding risk [94]. In addition, the increased difficulty in procuring sustenance necessitates a greater frequency of foraging, thereby increasing the likelihood of vessel collisions [95]. Bycatch and entanglement continue to pose significant hazards to the survival of cetaceans [96]. Seventy-five percent of odontoceti whales and sixty-four percent of baleen whales were accidentally captured in gillnets globally between 1990 and 2011 [97]. Certain gillnet fisheries in New Zealand and the United States have alarmingly high bycatch rates, resulting in a precipitous decline of small cetaceans [98,99]. A total of 36% of fishermen in the Kuching Bay region of Malaysia unintentionally caught cetaceans, but only 50% of them had the ability to release the animals unharmed, according to an investigation [100]. Additionally, abandoned fishing gear poses a threat to cetaceans, as individuals may become entangled, resulting in injury or death [101,102,103]. Indirectly increasing the risks associated with cetacean bycatch and entanglement, the expansion of fishing activities and rapid technological advancements from 1990 to 2006 rendered fishing tools and equipment more efficient, thereby increasing the likelihood of cetacean bycatch and entanglement [35,104]. Large-scale fishing operations may also contribute to habitat degradation, forcing cetaceans to seek out new habitats or alter their migration routes, thereby increasing their likelihood of interacting with shallow water areas [105]. In addition, fishing activities are frequently accompanied by vessel noise and the emission of sonar signals, which can interfere with the acoustic navigation and communication systems of cetaceans [106,107], resulting in disorientation or disrupted migration patterns. In conclusion, the intensification of coastal fish catch can increase interactions between cetaceans and fishing vessels, making cetaceans more vulnerable to the effects of fishing gear, resource competition, and acoustic disturbances, and thus increasing the likelihood of strandings. To reduce the number of cetacean strandings, sustainable fisheries management measures must be implemented to reduce the negative impacts of fishing on cetaceans and their habitats while ensuring the coexistence of fishing and cetacean conservation efforts.
A higher value of the PDOwin 2 yr-lag correlates with a larger number of cetacean strandings, and the model also indicates the existence of both direct and indirect effects. During the positive phase of the PDO, the East Asian Subtropical Western Jet Stream strengthens and shifts southward [108]. As the majority of recorded cetacean species inhabit slope and continental shelf regions, a positive PDO is likely to result in an increase in cetacean strandings. When large-scale oceanic climate changes occur, cetaceans may abandon their habitats, resulting in a decrease in species abundance and a decrease in the likelihood of cetacean strandings [109]. The absence of this phenomenon during the positive phase of the PDO can be attributed to the diverse species composition and varying adaptive abilities of various cetaceans. In addition to influencing the local climate, the PDO can indirectly affect cetacean strandings by affecting the distribution and abundance of their prey. During the negative phase of the PDO, there is a warming of seawater and an increase in primary productivity in the coastal waters of China, leading to an increase in prey for cetaceans and subsequently altering their distribution. This indirect effect of the PDO can contribute to variations in strandings.
In addition, the 2007–2021 model reveals a significant correlation between winter sea surface temperature and stranding records, suggesting that higher temperatures are associated with fewer strandings. The influence of the marine environment on cetaceans is predominantly determined by their ecological habits and environmental adaptability, resulting in variations in stranding patterns among various cetacean species [110]. In the western waters of Scotland, for instance, the relative strandings of white-beaked dolphins (a cold-water species) and common dolphins (a warm-water species) respond differently to variations in water temperature [111]. The majority of cetaceans recorded in this study are warm-water species, and as water temperatures rise, they are likely to migrate to polar regions, except for some particular species with high-site fidelity [112], thereby reducing their activity range. Therefore, if cetaceans migrate northward in years with higher temperatures, the abundance in the study area decreases, resulting in fewer strandings. The effect of temperature on the metabolism of cetaceans is another factor. Higher temperatures can reduce the availability of prey, making it difficult for cetaceans to obtain the necessary energy for growth and development, thereby delaying sexual maturation and lowering progeny survival rates [113,114]. Consequently, variations in temperature have a significant impact on reproductive rates and reproductive success, resulting in decreased population sizes and stranding rates [25,29]. The model demonstrates a significant negative correlation between salinity and strandings, a relationship that has been substantiated by previous research [52,54]. In instances of stranding events, a notable decline in seawater salinity has been detected. Cetaceans exposed to prolonged periods of low salinity conditions have been documented to develop dermatological abnormalities, including vesiculation, erosion, and the formation of intra-epithelial pustules, ultimately resulting in ulceration and hypodermal necrosis [53]. These pathological changes adversely affect the health of cetaceans, thereby increasing the occurrence of cetacean strandings. Stratification and circulation can influence the distribution of light, temperature, oxygen, and nutrients in the water, thereby influencing the availability of prey [115,116].
Although we have explained some of the causes of cetacean strandings, there are still unquantifiable factors that cannot be accounted for in the model. In several areas, vessel collisions are one of the leading anthropogenic causes of cetacean mortality [117,118,119]. In addition to fishing vessels, cruise ships, cargo ships, and sailboats are also capable of colliding with cetaceans. As marine fisheries, transportation, and tourism continue to develop, collisions with cetaceans are on the rise [120]. To reduce collision incidents, it is necessary to enhance collision records and conduct research on high-collision-rate regions and species. Additionally, it is difficult to acquire data on the distance from the shore and oceanic conditions at the time of cetacean fatalities [121]. In this study, we assumed that all dead whales would be observed and recorded, ignoring the possibility that individuals may sink, be scavenged, or decompose prior to stranding and that small cetaceans may be impeded from reaching the shore by various environmental factors [122]. Additionally, coastlines with higher human activity are more likely to detect strandings, thereby increasing the probability of inadvertent stranding records. To minimize the impact of these factors on research conclusions, it is essential to pay close attention to stranding details, such as the cause of mortality and decomposition status, and to increase stranding monitoring and reporting.
In order to use stranding data to support the research and conservation of cetaceans, it is essential to consider the trends and key influencing factors. Studies have revealed the spatiotemporal characteristics of strandings, which may reflect seasonal and interannual fluctuations in cetacean abundance and their regional and seasonal resource utilization patterns. We analyzed the direct and indirect impacts of various variables on cetacean strandings using structural equation models, revealing causal relationships between strandings and human activities, environmental variables, and prey distribution. Both the indirect and direct effects of the PDO’s impact on temperature emphasize the relationship between climate change and the stranding of cetaceans. Understanding how cetaceans respond to climate change can result in more effective protection measures and fewer stranding incidents. The information gathered from strandings can be used to guide stranding response efforts and the allocation of conservation resources, which are essential for conservation measures and management work, even though there may not be an immediate decline in the number of strandings due to an increasing focus on cetaceans and growing anthropogenic threats.
4.3. The Implications for Conservation and Management
This study provides insights into the historical patterns of cetacean strandings along the East China Sea coast and can be used to inform the management of marine resources. Firstly, the identification of cetacean stranding hotspots is a key outcome, allowing local authorities and conservation organizations to focus their efforts on monitoring and implementing protective measures in these specific areas [123]. By concentrating resources on these hotspots, conservation efforts can be more effective in safeguarding local cetacean populations. Secondly, the analysis of temporal trends and seasonal characteristics of cetacean strandings is valuable for local conservation. It equips them with the necessary information to allocate resources efficiently and implement timely response measures when there is an increase in stranding events [124,125]. Understanding the timing of strandings facilitates better planning and preparedness [126]. Additionally, it provides recommendations for policies related to fisheries, environmental protection, and other areas, thereby preventing indirect harm to cetaceans and improving overall management efficiency. Looking forward, future measures should be implemented to enhance local cetacean conservation and management. These measures may include increasing the importance of stranding data recording, enhancing environmental protection initiatives, and managing fishing activities. Such actions are essential for protecting and managing local cetacean populations while promoting the sustainability of marine ecosystems.
On a global scale, this study reinforces the significance of cetacean strandings in the context of cetacean conservation and management. Stranding data, as easily accessible information, plays a crucial role in identifying and addressing urgent issues related to cetacean protection [20]. Furthermore, the research underscores the global importance of the East China Sea due to its rich cetacean biodiversity. Given the migratory nature of cetaceans, the information obtained in this study can facilitate international cooperation among neighboring countries and international organizations. This cooperation can contribute to the development of regional protection strategies and the establishment of scientifically sound protected areas. Additionally, the cetacean stranding information obtained in this study can be used for comparative analysis in other global regions [126]. Researchers worldwide can utilize these data as reference points to enhance their understanding of cetacean strandings on a global scale. Moreover, the structural equation model employed in this study, with its ability to reveal cascading effects between factors, provides a valuable analytical method reference for other cetacean researchers globally. This methodology can aid in investigating complex relationships among environmental variables and cetacean strandings [126,127]. The findings of this study are not limited to the East China Sea region. They have broader applicability for global conservation efforts. This research on cetacean stranding factors contributes not only to the protection and management of cetaceans in the East China Sea but also to addressing similar issues in other regions globally.
In the realm of future research directions, there are several critical avenues that merit exploration. Firstly, a focus on species-specific research is paramount. It is imperative that future studies delve deeper into the intricacies of specific cetacean species, particularly those that exhibit pronounced seasonal strandings, exemplified by the narrow-ridged finless porpoise. Such investigations will provide the foundation for the development of targeted strategies aimed at reducing stranding probabilities. These measures may encompass heightened vigilance during peak stranding seasons and the prompt initiation of rescue operations [128]. Continued monitoring and a steadfast emphasis on stranding hotspots represent a second pivotal research trajectory. The formulation of effective cetacean stranding response plans, aimed at enhancing the success rate of rescue operations and ensuring a comprehensive acquisition of stranding information, holds the promise of yielding invaluable insights into the evolving stranding patterns and the adaptive responses of cetaceans to external fluctuations [44]. These findings hold immense significance in the formulation of dynamic management strategies that can better safeguard these marine species. Furthermore, the enhancement of our comprehension regarding cetacean interactions with other marine species constitutes a third avenue of exploration. Delving into the intricacies of how cetaceans engage with their prey and other marine organisms can culminate in a more comprehensive grasp of cetacean ecology and behavior [123,129]. This deeper insight is essential for the holistic understanding of these marine mammals within their ecosystem. Lastly, the knowledge garnered from this study should be actively harnessed for policy development. The information acquired has the potential to underpin the creation of protective measures that mitigate the impacts of various factors, such as fisheries and local environmental changes, on cetaceans. Policymakers must take these research findings into account when formulating decisions related to the conservation and management of cetaceans. In summation, these research directions not only hold the promise of benefiting local cetacean populations but also bear relevance to the global conservation and management endeavors surrounding these majestic marine mammals.
Author Contributions
Conceptualization, Z.L., S.L. and S.Y.; methodology, Z.L. and S.Y.; investigation, S.Y. and Z.L.; writing—original draft preparation, S.Y. and Z.L.; project administration, Z.L., Y.J. and S.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Key Research and Development Project (2020YFD0900804) and special funds for a survey of habitats for rare and endangered aquatic species by the Ministry of Agriculture and Rural Affairs (125C0505).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors are cordially thankful to the East China Sea Fisheries Research Institute for the possibility of research.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Roman, J.; Estes, J.A.; Morissette, L.; Smith, C.; Costa, D.; McCarthy, J.; Nation, J.; Nicol, S.; Pershing, A.; Smetacek, V. Whales as marine ecosystem engineers. Front. Ecol. Environ. 2014, 12, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Hazen, E.L.; Abrahms, B.; Brodie, S.; Carroll, G.; Jacox, M.G.; Savoca, M.S.; Scales, K.L.; Sydeman, W.J.; Bograd, S.J. Marine top predators as climate and ecosystem sentinels. Front. Ecol. Environ. 2019, 17, 565–574. [Google Scholar] [CrossRef]
- Wells, R.S.; Rhinehart, H.L.; Hansen, L.J.; Sweeney, J.C.; Townsend, F.I.; Stone, R.; Casper, D.R.; Scott, M.D.; Hohn, A.A.; Rowles, T.K. Bottlenose dolphins as marine ecosystem sentinels: Developing a health monitoring system. EcoHealth 2004, 1, 246–254. [Google Scholar] [CrossRef]
- Smith, L.V.; McMinn, A.; Martin, A.; Nicol, S.; Bowie, A.R.; Lannuzel, D.; van der Merwe, P. Preliminary investigation into the stimulation of phytoplankton photophysiology and growth by whale faeces. J. Exp. Mar. Biol. Ecol. 2013, 446, 1–9. [Google Scholar] [CrossRef]
- Berge, J.; Gabrielsen, T.M.; Moline, M.; Renaud, P.E. Evolution of the Arctic Calanus complex: An Arctic marine avocado? J. Plankton Res. 2012, 34, 191–195. [Google Scholar] [CrossRef]
- Parsons, K.M.; Everett, M.; Dahlheim, M.; Park, L. Water, water everywhere: Environmental DNA can unlock population structure in elusive marine species. Roy. Soc. Open. Sci. 2018, 5, 180537. [Google Scholar] [CrossRef]
- Santos, M.C.d.O.; Bressem, M.-F.V. Cetaceans using the marine protected area of “Parque Estadual Marinho da Laje de Santos”, Southeastern Brazil. Braz. J. Oceanogr. 2017, 65, 605–613. [Google Scholar] [CrossRef]
- Bailey, H.; Thompson, P.M. Using marine mammal habitat modelling to identify priority conservation zones within a marine protected area. Mar. Ecol. Prog. Ser. 2009, 378, 279–287. [Google Scholar] [CrossRef]
- Sequeira, A.M.M.; Hays, G.C.; Sims, D.W.; Eguíluz, V.M.; Rodríguez, J.P.; Heupel, M.R.; Harcourt, R.; Calich, H.; Queiroz, N.; Costa, D.P. Overhauling ocean spatial planning to improve marine megafauna conservation. Front. Mar. Sci. 2019, 6, 639. [Google Scholar] [CrossRef]
- Aguirre, A.A.; Tabor, G.M. Introduction: Marine vertebrates as sentinels of marine ecosystem health. EcoHealth 2004, 1, 236–238. [Google Scholar] [CrossRef]
- Friedlaender, A.S.; Halpin, P.N.; Qian, S.S.; Lawson, G.L.; Wiebe, P.H.; Thiele, D.; Read, A.J. Whale distribution in relation to prey abundance and oceanographic processes in shelf waters of the Western Antarctic Peninsula. Mar. Ecol. Prog. Ser. 2006, 317, 297–310. [Google Scholar] [CrossRef]
- Bossart, G.D. Marine mammals as sentinel species for oceans and human health. Vet. Pathol. 2011, 48, 676–690. [Google Scholar] [CrossRef]
- Evans, P.G.; Hammond, P.S. Monitoring cetaceans in European waters. Mammal. Rev. 2004, 34, 131–156. [Google Scholar] [CrossRef]
- Hammond, P.S.; Francis, T.B.; Heinemann, D.; Long, K.J.; Moore, J.E.; Punt, A.E.; Reeves, R.R.; Sepúlveda, M.; Sigurðsson, G.M.; Siple, M.C. Estimating the abundance of marine mammal populations. Front. Mar. Sci. 2021, 8, 1316. [Google Scholar] [CrossRef]
- IJsseldijk, L.L.; ten Doeschate, M.T.; Brownlow, A.; Davison, N.J.; Deaville, R.; Galatius, A.; Gilles, A.; Haelters, J.; Jepson, P.D.; Keijl, G.O. Spatiotemporal mortality and demographic trends in a small cetacean: Strandings to inform conservation management. Biol. Conserv. 2020, 249, 108733. [Google Scholar] [CrossRef]
- Thompson, K.F.; Millar, C.D.; Scott Baker, C.; Dalebout, M.; Steel, D.; van Helden, A.L.; Constantine, R. A novel conservation approach provides insights into the management of rare cetaceans. Biol. Conserv. 2013, 157, 331–340. [Google Scholar] [CrossRef]
- Zhao, L.; Zhong, M.; Wu, F.; Dai, Y.; Aierken, R.; Chen, M.; Wang, X. First Record of Omura’s Whale (Balaenoptera omurai) in the Beibu Gulf, China. Aquat. Mamm. 2020, 46, 301–306. [Google Scholar] [CrossRef]
- Pyenson, N.D. The high fidelity of the cetacean stranding record: Insights into measuring diversity by integrating taphonomy and macroecology. Proc. R. Soc. B 2011, 278, 3608–3616. [Google Scholar] [CrossRef]
- Quaggiotto, M.-M.; Sánchez-Zapata, J.A.; Bailey, D.M.; Payo-Payo, A.; Navarro, J.; Brownlow, A.; Deaville, R.; Lambertucci, S.A.; Selva, N.; Cortés-Avizanda, A. Past, present and future of the ecosystem services provided by cetacean carcasses. Ecosyst. Serv. 2022, 54, 101406. [Google Scholar] [CrossRef]
- Coombs, E.J.; Deaville, R.; Sabin, R.C.; Allan, L.; O’Connell, M.; Berrow, S.; Smith, B.; Brownlow, A.; Doeschate, M.T.; Penrose, R.; et al. What can cetacean stranding records tell us? A study of UK and Irish cetacean diversity over the past 100 years. Mar. Mamm. Sci. 2019, 35, 1527–1555. [Google Scholar] [CrossRef]
- Kemper, C.; Flaherty, A.; Gibbs, S.; Hill, M.; Long, M.; Byard, R. Cetacean captures, strandings and mortalities in South Australia 1881–2000, with special reference to human interactions. Aust. Mammal. 2005, 27, 37–47. [Google Scholar] [CrossRef]
- Foord, C.S.; Rowe, K.M.; Robb, K. Cetacean biodiversity, spatial and temporal trends based on stranding records (1920–2016), Victoria, Australia. PLoS ONE 2019, 14, e0223712. [Google Scholar] [CrossRef] [PubMed]
- Evans, K.; Thresher, R.; Warneke, R.; Bradshaw, C.J.; Pook, M.; Thiele, D.; Hindell, M.A. Periodic variability in caetacean strandings: Links to large-scale climate events. Biol. Lett. 2005, 1, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Cordes, D. The causes of whale strandings. N. Z. Vet. J. 1982, 30, 21–24. [Google Scholar] [CrossRef]
- Ortiz-Wolford, J.; Corona-Figueroa, M.F.; Dávila, V.; Cabrera, A.A. Cetacean stranding records along the Pacific coastline of Guatemala, 2007–2021: Implications for management, conservation and research. Mar. Policy. 2021, 134, 104827. [Google Scholar] [CrossRef]
- Liu, M.; Lin, M.; Zhang, P.; Xue, T.; Li, S. An overview of cetacean stranding around Hainan Island in the South China Sea, 1978–2016: Implications for research, conservation and management. Mar. Policy. 2019, 101, 147–153. [Google Scholar] [CrossRef]
- De Weerdt, J.; Ramos, E.A.; Pouplard, E.; Kochzius, M.; Clapham, P. Cetacean strandings along the Pacific and Caribbean coasts of Nicaragua from 2014 to 2021. Mar. Biodivers. Rec. 2021, 14, 13. [Google Scholar] [CrossRef]
- Mannino, M.A.; Talamo, S.; Tagliacozzo, A.; Fiore, I.; Nehlich, O.; Piperno, M.; Tusa, S.; Collina, C.; Di Salvo, R.; Schimmenti, V. Climate-driven environmental changes around 8200 years ago favoured increases in cetacean strandings and Mediterranean hunter-gatherers exploited them. Sci. Rep. 2015, 5, 16288. [Google Scholar] [CrossRef]
- Kebke, A.; Samarra, F.; Derous, D. Climate change and cetacean health: Impacts and future directions. Philos. T. R. Soc. B. 2022, 377, 20210249. [Google Scholar] [CrossRef]
- Russell, M.; Bloodgood, J.; Carmichael, R. Spatial, temporal and demographic patterns of cetacean strandings in the northcentral Gulf of Mexico. J. Cetacean Res. Manag. 2022, 23, 171–182. [Google Scholar] [CrossRef]
- Heinrich Vanselow, K.; Ricklefs, K.; Colijn, F. Solar driven geomagnetic anomalies and sperm whale (Physeter macrocephalus) strandings around the North Sea: An analysis of long term datasets. Open Mar. Biol. J. 2009, 3, 89–94. [Google Scholar] [CrossRef]
- Vanselow, K.H.; Jacobsen, S.; Hall, C.; Garthe, S. Solar storms may trigger sperm whale strandings: Explanation approaches for multiple strandings in the North Sea in 2016. Int. J. Astrobiol. 2018, 17, 336–344. [Google Scholar] [CrossRef]
- Lusher, A.L.; Hernandez-Milian, G.; Berrow, S.; Rogan, E.; O’Connor, I. Incidence of marine debris in cetaceans stranded and bycaught in Ireland: Recent findings and a review of historical knowledge. Environ. Pollut. 2018, 232, 467–476. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Delgado, J.; Fernández, A.; Sierra, E.; Sacchini, S.; Andrada, M.; Vela, A.I.; Quesada-Canales, Ó.; Paz, Y.; Zucca, D.; Groch, K. Pathologic findings and causes of death of stranded cetaceans in the Canary Islands (2006–2012). PLoS ONE 2018, 13, e0204444. [Google Scholar] [CrossRef] [PubMed]
- Peltier, H.; Authier, M.; Deaville, R.; Dabin, W.; Jepson, P.D.; van Canneyt, O.; Daniel, P.; Ridoux, V. Small cetacean bycatch as estimated from stranding schemes: The common dolphin case in the northeast Atlantic. Environ. Sci. Policy. 2016, 63, 7–18. [Google Scholar] [CrossRef]
- Walker, W.A.; Coe, J.M. Survey of marine debris ingestion by odontocete cetaceans. In Proceedings of the second international conference on marine debris, Honolulu, HI, USA, 2–7 April 1989; pp. 2–7. [Google Scholar]
- Parsons, E.; Dolman, S.J.; Wright, A.J.; Rose, N.A.; Burns, W. Navy sonar and cetaceans: Just how much does the gun need to smoke before we act? Mar. Pollut. Bull. 2008, 56, 1248–1257. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; He, M.; Meng, W.; Zhang, Y.; Yun, H.; Lu, Y.; Huang, Z.; Mo, X.; Hu, B.; Liu, B.; et al. Temporal-spatial change of China’s coastal ecosystems health and driving factors analysis. Sci. Total Environ. 2022, 845, 157319. [Google Scholar] [CrossRef] [PubMed]
- Committee on Taxonomy. List of Marine Mammal Species and Subspecies. Society for Marine Mammalogy. Available online: www.marinemammalscience.org (accessed on 1 September 2023).
- Liu, M.; Lin, M.; Li, S. Species diversity and spatiotemporal patterns based on cetacean stranding records in China, 1950–2018. Sci. Total Environ. 2022, 822, 153651. [Google Scholar] [CrossRef]
- Wei, G.; Zhang, S.; Yi, Y.; Liu, s.; Wang, L.; Ding, F.; Fu, Y.; Wu, M. Define wildlife protection and promote economic animal production—Understanding of new list of key protected wild animals in china. J. Econ. Anim. 2021, 4, 25. [Google Scholar] [CrossRef]
- Li, Y. The Ministry of Agriculture and Rural Affairs of the People’s Republic of China issued a document to strengthen the protection of marine mammals. Ocean. Fish. 2021, 5, 32–33. [Google Scholar] [CrossRef]
- Aragones, L.V.; Jefferson, T.A.; Marsh, H. Marine mammal survey techniques applicable in developing countries. Asian Mar. Biol. 1997, 14, 15–39. [Google Scholar] [CrossRef]
- Zhao, L.; Zhu, Q.; Miao, X.; Xu, M.; Wu, F.; Dai, Y.; Tao, C.; Mou, J.; Wang, X. An overview of cetacean strandings, bycatches and rescues along the western coast of the Taiwan Strait, China: 2010–2015. Acta Oceanolog. Sin. 2017, 36, 31–36. [Google Scholar] [CrossRef]
- Parsons, E. Strandings of small cetaceans in Hong Kong territorial waters. J. Mar. Biol. Assoc. UK 1998, 78, 1039–1042. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, D.; Geng, T. Research on the sustainable development strategy of China’s deep blue fishery economy under climate change. J. Shandong Univ. Sci. Technol. (Soc. Sci.) 2018, 6, 121–129. [Google Scholar]
- Schumann, N.; Gales, N.J.; Harcourt, R.G.; Arnould, J.P. Impacts of climate change on Australian marine mammals. Aust. J. Zool. 2013, 61, 146–159. [Google Scholar] [CrossRef]
- Santora, J.A.; Hazen, E.L.; Schroeder, I.D.; Bograd, S.J.; Sakuma, K.M.; Field, J.C. Impacts of ocean climate variability on biodiversity of pelagic forage species in an upwelling ecosystem. Mar. Ecol. Prog. Ser. 2017, 580. [Google Scholar] [CrossRef]
- Oviatt, C.; Smith, L.; McManus, M.C.; Hyde, K. Decadal Patterns of Westerly Winds, Temperatures, Ocean Gyre Circulations and Fish Abundance: A Review. Climate 2015, 3, 833–857. [Google Scholar] [CrossRef]
- Xu, C.; Zhang, L.; Yu, J.; Zhang, Y. Impact of climate change on China’soffshore fishing: Taking the pacific decadal oscillation as an example. Resour. Sci. 2022, 44, 386–400. [Google Scholar] [CrossRef]
- Learmonth, J.A.; MacLeod, C.D.; Santos, M.B.; Pierce, G.J.; Crick, H.; Robinson, R. Potential effects of climate change on marine mammals. Oceanogr. Mar. Biol. 2006, 44, 431. [Google Scholar] [CrossRef]
- Duignan, P.J.; Stephens, N.S.; Robb, K. Fresh water skin disease in dolphins: A case definition based on pathology and environmental factors in Australia. Sci. Rep. 2020, 10, 21979. [Google Scholar] [CrossRef]
- Bloodgood, J.C.G.; Deming, A.C.; Colegrove, K.M.; Russell, M.L.; Díaz Clark, C.; Carmichael, R.H. Causes of death and pathogen prevalence in bottlenose dolphins Tursiops truncatus stranded in Alabama, USA, between 2015 and 2020, following the Deepwater Horizon oil spill. Dis. Aquat. Org. 2023, 155, 87–102. [Google Scholar] [CrossRef] [PubMed]
- Toms, C.N.; Stone, T.; Och, T. Skin lesion and mortality rate estimates for common bottlenose dolphin (Tursiops truncatus) in the Florida Panhandle following a historic flood. PLoS ONE 2021, 16, e0257526. [Google Scholar] [CrossRef] [PubMed]
- Gulland, F.M.; Baker, J.; Howe, M.; LaBrecque, E.; Leach, L.; Moore, S.E.; Reeves, R.R.; Thomas, P.O. A Review of Climate Change Effects on Marine Mammals in United States Waters: Past Predictions, Observed Impacts, Current Research and Conservation Imperatives. Clim. Change. Ecol. 2022, 3, 100054. [Google Scholar] [CrossRef]
- Law, R.J.; Barry, J.; Barber, J.L.; Bersuder, P.; Deaville, R.; Reid, R.J.; Brownlow, A.; Penrose, R.; Barnett, J.; Loveridge, J. Contaminants in cetaceans from UK waters: Status as assessed within the Cetacean Strandings Investigation Programme from 1990 to 2008. Mar. Pollut. Bull. 2012, 64, 1485–1494. [Google Scholar] [CrossRef]
- Bearzi, G.; Politi, E.; Agazzi, S.; Azzellino, A. Prey depletion caused by overfishing and the decline of marine megafauna in eastern Ionian Sea coastal waters (central Mediterranean). Biol. Conserv. 2006, 127, 373–382. [Google Scholar] [CrossRef]
- Leeney, R.H.; Amies, R.; Broderick, A.C.; Witt, M.J.; Loveridge, J.; Doyle, J.; Godley, B.J. Spatio-temporal analysis of cetacean strandings and bycatch in a UK fisheries hotspot. Biodivers. Conserv. 2008, 17, 2323–2338. [Google Scholar] [CrossRef]
- Trapletti, A.; Hornik, K.; LeBaron, B. Time Series Analysis and Computational Finance. 2015. Available online: http://cran.r-project.org/web/packages/tseries/index.html (accessed on 3 September 2023).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015; Available online: http://www.R-project.org (accessed on 3 September 2023).
- Pohlert, T. Non-parametric trend tests and change-point detection. CC BY-ND 2016, 4, 1–18. [Google Scholar]
- Mustika, P.L.K.; High, K.K.; Putra, M.I.H.; Sahri, A.; Ratha, I.M.J.; Prinanda, M.O.; Agung, F.; Purnomo, F.S.; Kreb, D. When and Where Did They Strand? The Spatio-Temporal Hotspot Patterns of Cetacean Stranding Events in Indonesia. Oceans 2022, 3, 509–526. [Google Scholar] [CrossRef]
- Carpenter, T.E. Methods to investigate spatial and temporal clustering in veterinary epidemiology. Prev. Vet. Med. 2001, 48, 303–320. [Google Scholar] [CrossRef]
- Getis, A.; Ord, J.K. The analysis of spatial association by use of distance statistics. Geogr. Anal. 1992, 24, 189–206. [Google Scholar] [CrossRef]
- Grace, J.B. Structural Equation Modeling and Natural Systems; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Kline, R.B. Principles and Practice of Structural Equation Modeling; Guilford Publications: New York, NY, USA, 2015. [Google Scholar]
- Lefcheck, J.S. piecewiseSEM: Piecewise structural equation modelling in R for ecology, evolution, and systematics. Methods. Ecol. Evol. 2016, 7, 573–579. [Google Scholar] [CrossRef]
- Katz, M.H. Multivariable Analysis: A Practical Guide for Clinicians and PublicHealth Researchers; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Shipley, B. Confirmatory path analysis in a generalized multilevel context. Ecology 2009, 90, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Vijverberg, S.; Coumou, D. The role of the Pacific Decadal Oscillation and ocean-atmosphere interactions in driving US temperature predictability. npj Clim. Atmos. Sci. 2022, 5, 18. [Google Scholar] [CrossRef]
- Matsumura, S.; Horinouchi, T. Pacific Ocean decadal forcing of long-term changes in the western Pacific subtropical high. Sci. Rep. 2016, 6, 37765. [Google Scholar] [CrossRef]
- Wursig, B.; Perrin, W.F. Encyclopedia of Marine Mammals; Academic Press: Cambridge, MA, USA, 2009. [Google Scholar]
- Truchon, M.-H.; Measures, L.; L’Hérault, V.; Brethes, J.-C.; Galbraith, P.S.; Harvey, M.; Lessard, S.; Starr, M.; Lecomte, N. Marine mammal strandings and environmental changes: A 15-year study in the St. Lawrence ecosystem. PLoS ONE 2013, 8, e59311. [Google Scholar] [CrossRef]
- Groom, C.J.; Coughran, D.K.; Smith, H.C. Records of beaked whales (family Ziphiidae) in Western Australian waters. Mar. Biodivers. Rec. 2014, 7, e50. [Google Scholar] [CrossRef]
- Mitchell, E. Northeast Pacific stranding distribution and seasonality of Cuvier’s beaked whale Ziphius cavirostris. Can. J. Zool. 1968, 46, 265–279. [Google Scholar] [CrossRef]
- Owen, K.; Kavanagh, A.S.; Warren, J.D.; Noad, M.J.; Donnelly, D.; Goldizen, A.W.; Dunlop, R.A. Potential energy gain by whales outside of the Antarctic: Prey preferences and consumption rates of migrating humpback whales (Megaptera novaeangliae). Polar Biol. 2017, 40, 277–289. [Google Scholar] [CrossRef]
- Norman, S.A.; Huggins, J.; Carpenter, T.E.; Case, J.T.; Lambourn, D.M.; Rice, J.; Calambokidis, J.; Gaydos, J.K.; Hanson, M.B.; Duffield, D.A. The application of GIS and spatiotemporal analyses to investigations of unusual marine mammal strandings and mortality events. Mar. Mamm. Sci. 2012, 28, E251–E266. [Google Scholar] [CrossRef]
- Norman, S.; Bowlby, C.; Brancato, M.; Calambokidis, J.; Duffield, D.; Gearin, P.; Gornall, T.; Gosho, M.; Hanson, B.; Hodder, J. Cetacean strandings in Oregon and Washington between 1930 and 2002. J. Cetacean Res. Manag. 2004, 6, 87–100. [Google Scholar] [CrossRef]
- Liu, M.; Lin, M.; Turvey, S.; Li, S. Fishers’ knowledge as an information source to investigate bycatch of marine mammals in the South China Sea. Anim. Conserv. 2017, 20, 182–192. [Google Scholar] [CrossRef]
- Kaiya, Z.; Leatherwood, S.; Jefferson, T.A. Records of small cetaceans in Chinese waters: A review. Asian Mar. Biol. 1995, 12, 119–139. [Google Scholar]
- Dong, J.; Wang, G.; Xiao, Z. Migration and population difference of the finless porpoise in China. Mar. Sci. 1993, 5, 42–45. [Google Scholar]
- Wang, P. A research of Odontoceti cetaceans in the Yellow Sea and Bohai Sea. Chin. J. Zool. 1979, 2, 31–34. [Google Scholar]
- Wang, Y.; Li, W.; Van Waerebeek, K. Strandings, bycatches and injuries of aquatic mammals in China, 2000–2006, as reviewed from official documents: A compelling argument for a nationwide strandings programme. Mar. Policy. 2015, 51, 242–250. [Google Scholar] [CrossRef]
- Nemiroff, L.; Wimmer, T.; Daoust, P.-Y.; McAlpine, D.F. Cetacean strandings in the Canadian Maritime provinces, 1990–2008. Can. Field. Nat. 2010, 124, 32–44. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, J.; Song, P.; Zhang, R.; Li, Y.; Zhong, Z.; Lin, L. Composition of the Taiwan Strait fish fauna. Biodivers. Sci. 2014, 22, 525. [Google Scholar] [CrossRef]
- Li, S. Studies on sea mammals and its distribution from Fujian coastal waters. J. Oceanogr. Taiwan Strait 1997, 16, 485–488. [Google Scholar]
- Li, W.-T.; Chou, L.-S.; Chiou, H.-Y.; Chen, I.-H.; Yang, W.-C. Analyzing 13 years of cetacean strandings: Multiple stressors to cetaceans in Taiwanese waters and their implications for conservation and future research. Front. Mar. Sci. 2021, 8, 606722. [Google Scholar] [CrossRef]
- Aniceto, A.S.; Tassara, L.; Rikardsen, A.; Blévin, P. Mass strandings of seven toothed and baleen whale species in Northern Norway in March 2020 call for further investigation. Polar Biol. 2021, 44, 1457–1461. [Google Scholar] [CrossRef]
- Geraci, J.R.; Lounsbury, V.J. Marine Mammals Ashore: A Field Guide for Strandings; National Aquarium in Baltimore: Baltimore, MD, USA, 2005. [Google Scholar]
- Moore, M.J.; Mitchell, G.H.; Rowles, T.K.; Early, G. Dead cetacean? Beach, bloat, float, sink. Front. Mar. Sci. 2020, 7, 333. [Google Scholar] [CrossRef]
- Murthy, C.; Sinha, P.; Rao, Y.; Dube, S.; Rao, A.; Sinha, P.; Chittibabu, P. Storm surges: Worst coastal marine hazard. In Modelling and Monitoring of Coastal Marine Processes; Capital Publishing Company: New Delhi, India, 2008; pp. 125–140. [Google Scholar] [CrossRef]
- Lawler, I.R.; Parra, G.; Noad, M. Vulnerability of marine mammals in the Great Barrier Reef to climate change. In Climate Change and the Great Barrier Reef: A Vulnerability Assessment; The Great Barrier Reef Marine Park Authority: Townsville City, Australia, 2007. [Google Scholar] [CrossRef]
- Weir, C.R.; Pierce, G.J. A review of the human activities impacting cetaceans in the eastern tropical Atlantic. Mammal. Rev. 2013, 43, 258–274. [Google Scholar] [CrossRef]
- Giralt Paradell, O.; Methion, S.; Rogan, E.; Díaz López, B. Modelling ecosystem dynamics to assess the effect of coastal fisheries on cetacean species. J. Environ. Manag. 2021, 285, 112175. [Google Scholar] [CrossRef]
- Pennino, M.G.; Rufener, M.-C.; Giménez, J.; Berlinguer, F.; Bollo, E.; Appino, S.; Zucca, D.; Chessa, G.; Rotta, A. Understanding the causes of mortality and contaminant loads of stranded cetacean species in Sardinian waters (Italy) using Bayesian Hierarchical Models. J. Sea Res. 2022, 181, 102170. [Google Scholar] [CrossRef]
- Read, A.J. The looming crisis: Interactions between marine mammals and fisheries. J. Mammal. 2008, 89, 541–548. [Google Scholar] [CrossRef]
- Reeves, R.R.; McClellan, K.; Werner, T.B. Marine mammal bycatch in gillnet and other entangling net fisheries, 1990 to 2011. Endanger. Species. Res. 2013, 20, 71–97. [Google Scholar] [CrossRef]
- Hamner, R.M.; Constantine, R.; Oremus, M.; Stanley, M.; Brown, P.; Scott Baker, C. Long-range movement by Hector’s dolphins provides potential genetic enhancement for critically endangered Maui’s dolphin. Mar. Mamm. Sci. 2014, 30, 139–153. [Google Scholar] [CrossRef]
- Jaramillo-Legorreta, A.M.; Cardenas-Hinojosa, G.; Nieto-Garcia, E.; Rojas-Bracho, L.; Thomas, L.; Ver Hoef, J.M.; Moore, J.; Taylor, B.; Barlow, J.; Tregenza, N. Decline towards extinction of Mexico’s vaquita porpoise (Phocoena sinus). Roy. Soc. Open. Sci. 2019, 6, 190598. [Google Scholar] [CrossRef]
- Ambie, S.; Peter, C.; Minton, G.; Ngeian, J.; Zulkifli Poh, A.N.; Mujahid, A.; Tuen, A.A. Utilizing interview-based data to measure interactions of artisanal fishing communities and cetacean populations in Kuching Bay, Sarawak, East Malaysia. Ocean. Coast. Manag. 2023, 239, 106592. [Google Scholar] [CrossRef]
- Benaka, L.; Bullock, D.; Davis, J.; Seney, E.; Winarsoo, H. US National Bycatch Report (Update 1); US Department of Commerce: Washington, DC, USA, 2013. [Google Scholar]
- Gilman, E.; Brothers, N.; McPherson, G.; Dalzell, P. A review of cetacean interactions with longline gear. J. Cetacean Res. Manag. 2007, 8, 215. [Google Scholar] [CrossRef]
- Tulloch, V.; Pirotta, V.; Grech, A.; Crocetti, S.; Double, M.; How, J.; Kemper, C.; Meager, J.; Peddemors, V.; Waples, K. Long-term trends and a risk analysis of cetacean entanglements and bycatch in fisheries gear in Australian waters. Biodivers. Conserv. 2020, 29, 251–282. [Google Scholar] [CrossRef]
- de Quirós, Y.B.; Hartwick, M.; Rotstein, D.S.; Garner, M.M.; Bogomolni, A.; Greer, W.; Niemeyer, M.E.; Early, G.; Wenzel, F.; Moore, M. Discrimination between bycatch and other causes of cetacean and pinniped stranding. Dis. Aquat. Org. 2018, 127, 83–95. [Google Scholar] [CrossRef]
- Díaz López, B.; Methion, S.; Giralt Paradell, O. Living on the edge: Overlap between a marine predator’s habitat use and fisheries in the Northeast Atlantic waters (NW Spain). Prog. Oceanogr. 2019, 175, 115–123. [Google Scholar] [CrossRef]
- Evans, P.G.H. Chapter 5—Conservation threats. In European Whales, Dolphins, and Porpoises; Evans, P.G.H., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 159–202. [Google Scholar]
- Board, O.S.; Council, N.R. Marine Mammal Populations and Ocean Noise: Determining When Noise Causes Biologically Significant Effects; National Academies Press: Washington, DC, USA, 2005. [Google Scholar]
- Zhang, W.; Dong, X.; Xue, F. Intraseasonal variations of the East Asian Summer Monsoon in El Niño developing years and La Niña years under different phases of the Pacific Decadal Oscillation. Chin. J. Atmos. Sci. 2020, 44, 390–406. [Google Scholar]
- Saavedra, C.; Pierce, G.J.; Gago, J.; Jusufovski, D.; Cabrero, Á.; Cerviño, S.; López, A.; Martínez-Cedeira, J.A.; Santos, M.B. Factors driving patterns and trends in strandings of small cetaceans. Mar. Biol. 2017, 164, 165. [Google Scholar] [CrossRef]
- Warlick, A.J.; Huggins, J.L.; Lambourn, D.M.; Duffield, D.A.; D’Alessandro, D.N.; Rice, J.M.; Calambokidis, J.; Hanson, M.B.; Gaydos, J.K.; Jeffries, S.J.; et al. Cetacean strandings in the US Pacific northwest 2000–2019 reveal potential linkages to oceanographic variability. Front. Mar. Sci. 2022, 9, 758812. [Google Scholar] [CrossRef]
- MacLeod, C.D.; Bannon, S.M.; Pierce, G.J.; Schweder, C.; Learmonth, J.A.; Herman, J.S.; Reid, R.J. Climate change and the cetacean community of north-west Scotland. Biol. Conserv. 2005, 124, 477–483. [Google Scholar] [CrossRef]
- MacLeod, C.D. Global climate change, range changes and potential implications for the conservation of marine cetaceans: A review and synthesis. Endanger. Species Res. 2009, 7, 125–136. [Google Scholar] [CrossRef]
- Wild, S.; Krützen, M.; Rankin, R.W.; Hoppitt, W.J.; Gerber, L.; Allen, S.J. Long-term decline in survival and reproduction of dolphins following a marine heatwave. Curr. Biol. 2019, 29, R239–R240. [Google Scholar] [CrossRef]
- IJsseldijk, L.L.; Hessing, S.; Mairo, A.; Ten Doeschate, M.T.; Treep, J.; van den Broek, J.; Keijl, G.O.; Siebert, U.; Heesterbeek, H.; Gröne, A. Nutritional status and prey energy density govern reproductive success in a small cetacean. Sci. Rep. 2021, 11, 19201. [Google Scholar] [CrossRef]
- Ullah, A.; Bano, A.; Khan, N. Climate change and salinity effects on crops and chemical communication between plants and plant growth-promoting microorganisms under stress. Front. Sustain. Food. Syst. 2021, 5, 618092. [Google Scholar] [CrossRef]
- Velasco, J.; Gutiérrez-Cánovas, C.; Botella-Cruz, M.; Sánchez-Fernández, D.; Arribas, P.; Carbonell, J.A.; Millán, A.; Pallarés, S. Effects of salinity changes on aquatic organisms in a multiple stressor context. Philos. T. R. Soc. B. 2019, 374, 20180011. [Google Scholar] [CrossRef] [PubMed]
- Cates, K.; DeMaster, D.; Brownell Jr, R.; Silber, G.; Gende, S.; Leaper, R. Strategic Plan to Mitigate the Impacts of Ship Strikes on Cetacean Populations: 2017–2020, IWC Strategic Plan to Mitigate Ship Strikes. Available online: https://iwc.int/ship-strikes (accessed on 6 September 2023).
- Laist, D.W.; Knowlton, A.R.; Mead, J.G.; Collet, A.S.; Podesta, M. Collisions between ships and whales. Mar. Mamm. Sci. 2001, 17, 35–75. [Google Scholar] [CrossRef]
- Peltier, H.; Beaufils, A.; Cesarini, C.; Dabin, W.; Dars, C.; Demaret, F.; Dhermain, F.; Doremus, G.; Labach, H.; Van Canneyt, O. Monitoring of marine mammal strandings along French coasts reveals the importance of ship strikes on large cetaceans: A challenge for the European Marine Strategy Framework Directive. Front. Mar. Sci. 2019, 6, 486. [Google Scholar] [CrossRef]
- Douglas, A.B.; Calambokidis, J.; Raverty, S.; Jeffries, S.J.; Lambourn, D.M.; Norman, S.A. Incidence of ship strikes of large whales in Washington State. J. Mar. Biol. Assoc. UK 2008, 88, 1121–1132. [Google Scholar] [CrossRef]
- Martínez-Cedeira, J.; Morales, X.; Garcia, J.; Parada, A.; Covelo, P.; López, A. How many strand? Offshore marking and coastal recapture of cetacean carcasses. In Proceedings of the Abstract Book—25th Conference of the European Cetacean Society, Cadiz, Spain, 21–23 March 2011. [Google Scholar]
- Peltier, H.; Dabin, W.; Daniel, P.; Van Canneyt, O.; Dorémus, G.; Huon, M.; Ridoux, V. The significance of stranding data as indicators of cetacean populations at sea: Modelling the drift of cetacean carcasses. Ecol. Indic. 2012, 18, 278–290. [Google Scholar] [CrossRef]
- Betty, E.L.; Bollard, B.; Murphy, S.; Ogle, M.; Hendriks, H.; Orams, M.B.; Stockin, K.A. Using emerging hot spot analysis of stranding records to inform conservation management of a data-poor cetacean species. Biodivers. Conserv. 2020, 29, 643–665. [Google Scholar] [CrossRef]
- Pradip Na Thalang, P.; Thongratsakul, S.; Poolkhet, C. Spatial, Temporal, and Geographical Factors Associated with Stranded Marine Endangered Species in Thailand during 2006–2015. Biology 2023, 12, 448. [Google Scholar] [CrossRef]
- Dudhat, S.; Pande, A.; Nair, A.; Mondal, I.; Srinivasan, M.; Sivakumar, K. Spatio-temporal analysis identifies marine mammal stranding hotspots along the Indian coastline. Sci. Rep. 2022, 12, 4128. [Google Scholar] [CrossRef]
- Peltier, H.; Ridoux, V. Marine megavertebrates adrift: A framework for the interpretation of stranding data in perspective of the European Marine Strategy Framework Directive and other regional agreements. Environ. Sci. Policy. 2015, 54, 240–247. [Google Scholar] [CrossRef]
- Ganley, L.C.; Byrnes, J.; Pendleton, D.E.; Mayo, C.A.; Friedland, K.D.; Redfern, J.V.; Turner, J.T.; Brault, S. Effects of changing temperature phenology on the abundance of a critically endangered baleen whale. Glob. Ecol. Conserv. 2022, 38, e02193. [Google Scholar] [CrossRef]
- Prado, J.H.F.; Mattos, P.H.; Silva, K.G.; Secchi, E.R. Long-Term Seasonal and Interannual Patterns of Marine Mammal Strandings in Subtropical Western South Atlantic. PLoS ONE 2016, 11, e0146339. [Google Scholar] [CrossRef] [PubMed]
- Spitz, J.; Richard, E.; Meynier, L.; Pusineri, C.; Ridoux, V. Dietary plasticity of the oceanic striped dolphin, Stenella coeruleoalba, in the neritic waters of the Bay of Biscay. J. Sea Res. 2006, 55, 309–320. [Google Scholar] [CrossRef]
Figure 1.
The red box is map of study and the area where the local variables are gathered (Jiangsu (JS), Shanghai (SH), Zhejiang (ZJ), Fujian (FJ), and Taiwan (TW)).
Figure 1.
The red box is map of study and the area where the local variables are gathered (Jiangsu (JS), Shanghai (SH), Zhejiang (ZJ), Fujian (FJ), and Taiwan (TW)).
Figure 2.
The annual trend of stranded cetacean individuals and events in JS, SH, ZJ, FJ and TW between 1990 and 2021.
Figure 2.
The annual trend of stranded cetacean individuals and events in JS, SH, ZJ, FJ and TW between 1990 and 2021.
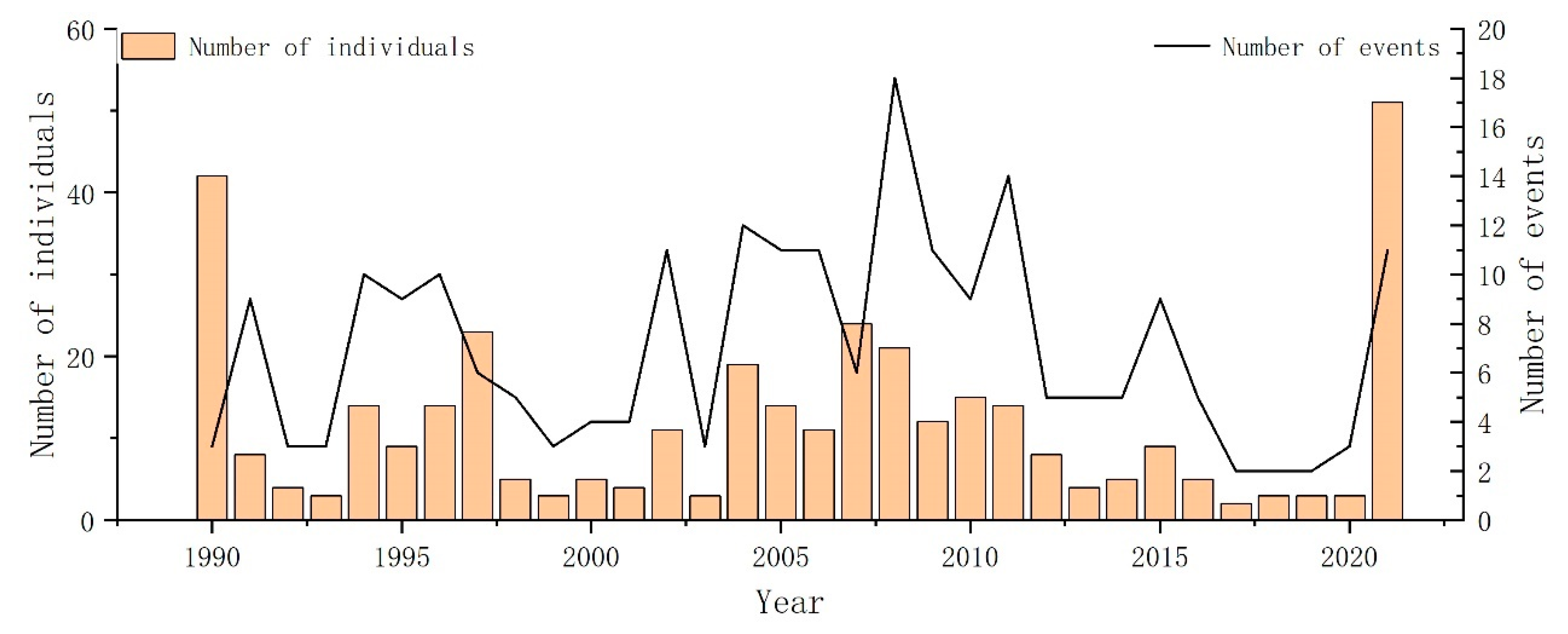
Figure 3.
The total number of cetacean stranding events in JS, SH, ZJ, FJ, and TW between 1990 and 2021, separated by month within each season.
Figure 3.
The total number of cetacean stranding events in JS, SH, ZJ, FJ, and TW between 1990 and 2021, separated by month within each season.

Figure 4.
50-km × 50-km grid-cell counts Jiangsu (JS), Shanghai (SH), Zhejiang (ZJ), Fujian (FJ), and Taiwan (TW).
Figure 4.
50-km × 50-km grid-cell counts Jiangsu (JS), Shanghai (SH), Zhejiang (ZJ), Fujian (FJ), and Taiwan (TW).
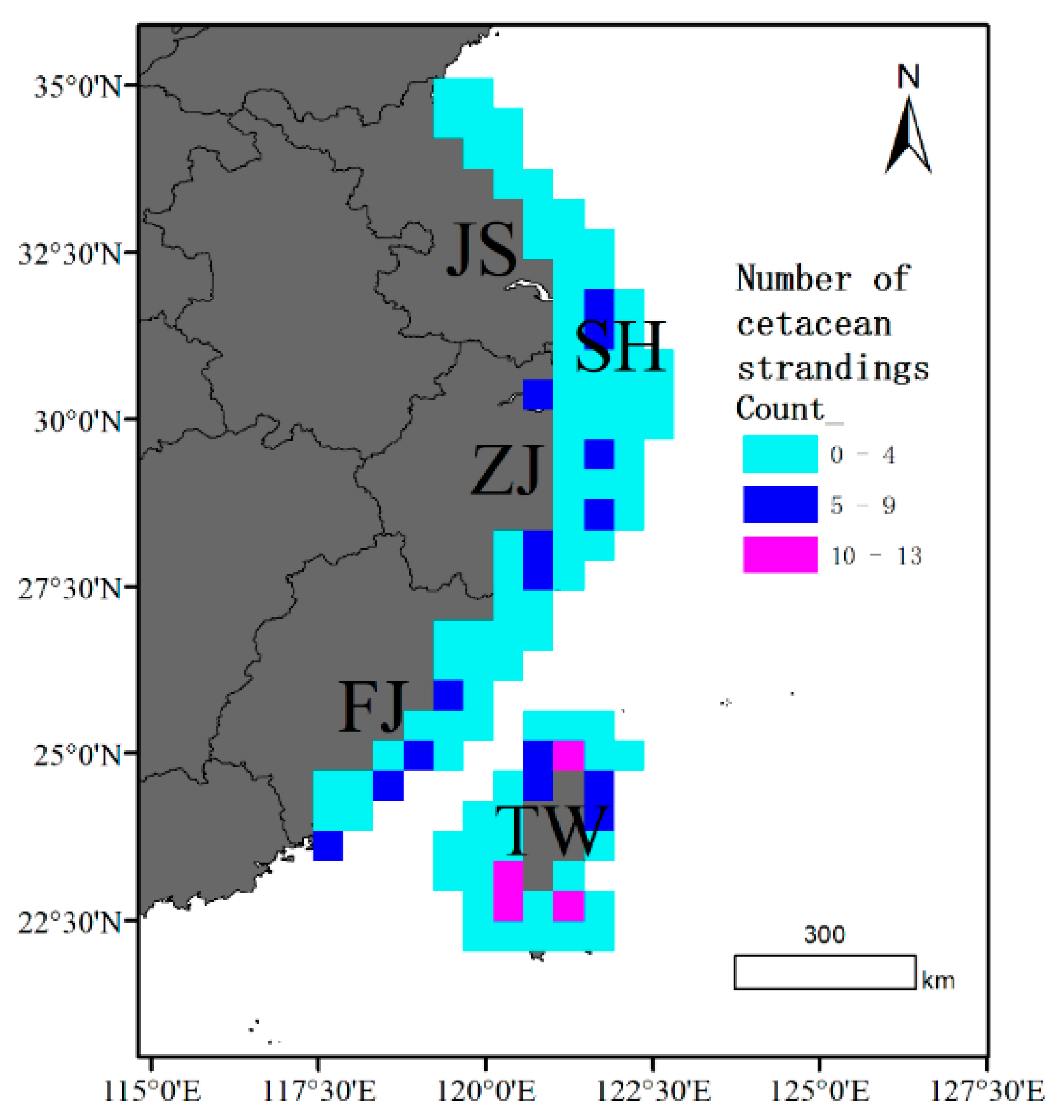
Figure 5.
Hotspot analysis of stranding events, 1990–2021.
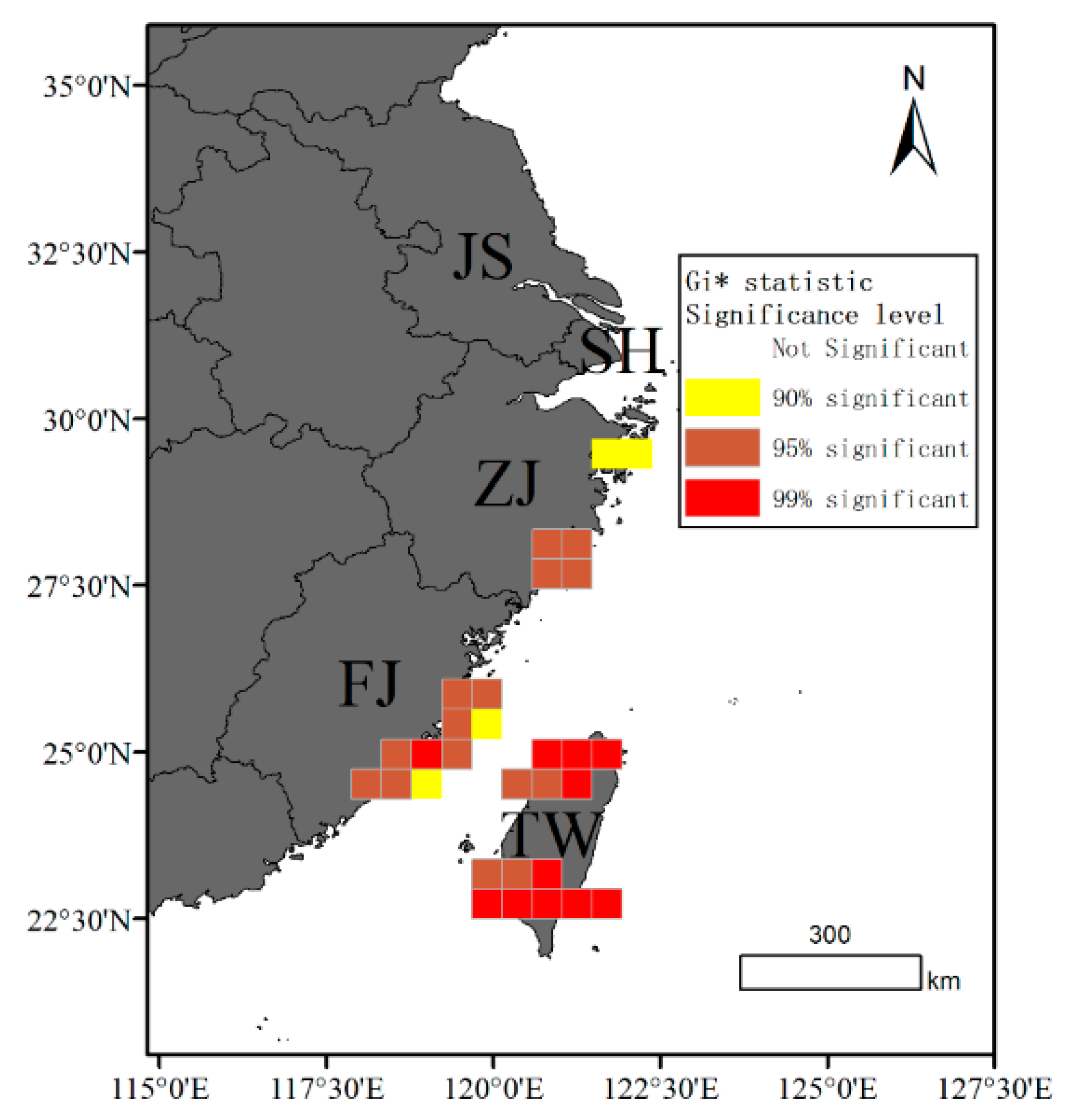
Figure 6.
Piecewise structural equation modeling (SEM) describing the effects of predictor variables on cetacean strandings in two periods. The first period was from 1990 to 2006 (A), and the second period was from 2006 to 2021 (B). *: p < 0.05, **: p < 0.01, ***: p < 0.001. Significant paths (p < 0.05) are presented in blue (positive effects) and in red (negative effects). Non-significant paths (p > 0.05) are shown in dashed gray lines. Numbers adjacent to arrows indicate path coefficient estimates. The variance explained by the model (R2) is shown next to each response variable.
Figure 6.
Piecewise structural equation modeling (SEM) describing the effects of predictor variables on cetacean strandings in two periods. The first period was from 1990 to 2006 (A), and the second period was from 2006 to 2021 (B). *: p < 0.05, **: p < 0.01, ***: p < 0.001. Significant paths (p < 0.05) are presented in blue (positive effects) and in red (negative effects). Non-significant paths (p > 0.05) are shown in dashed gray lines. Numbers adjacent to arrows indicate path coefficient estimates. The variance explained by the model (R2) is shown next to each response variable.
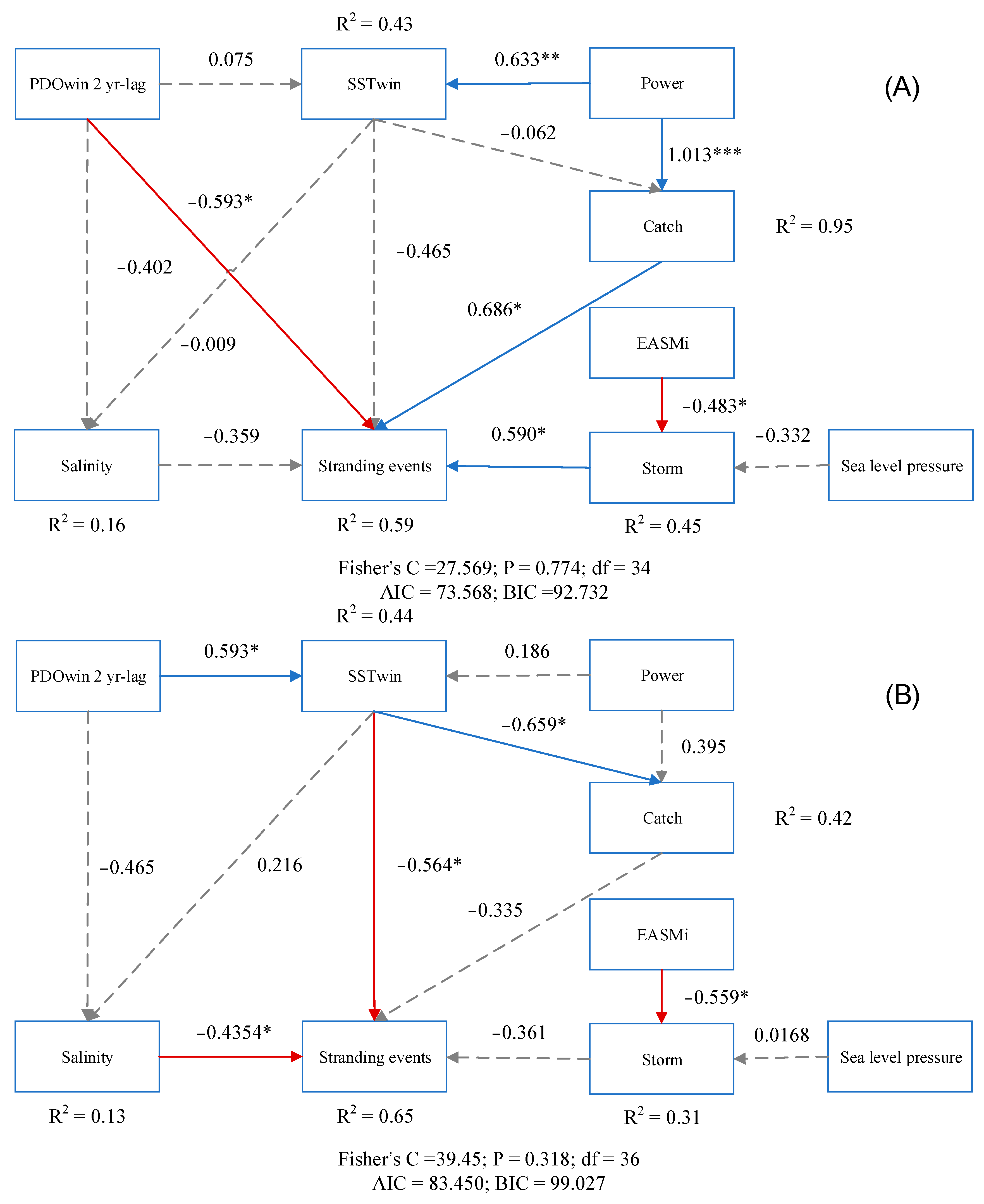

{kind=link}
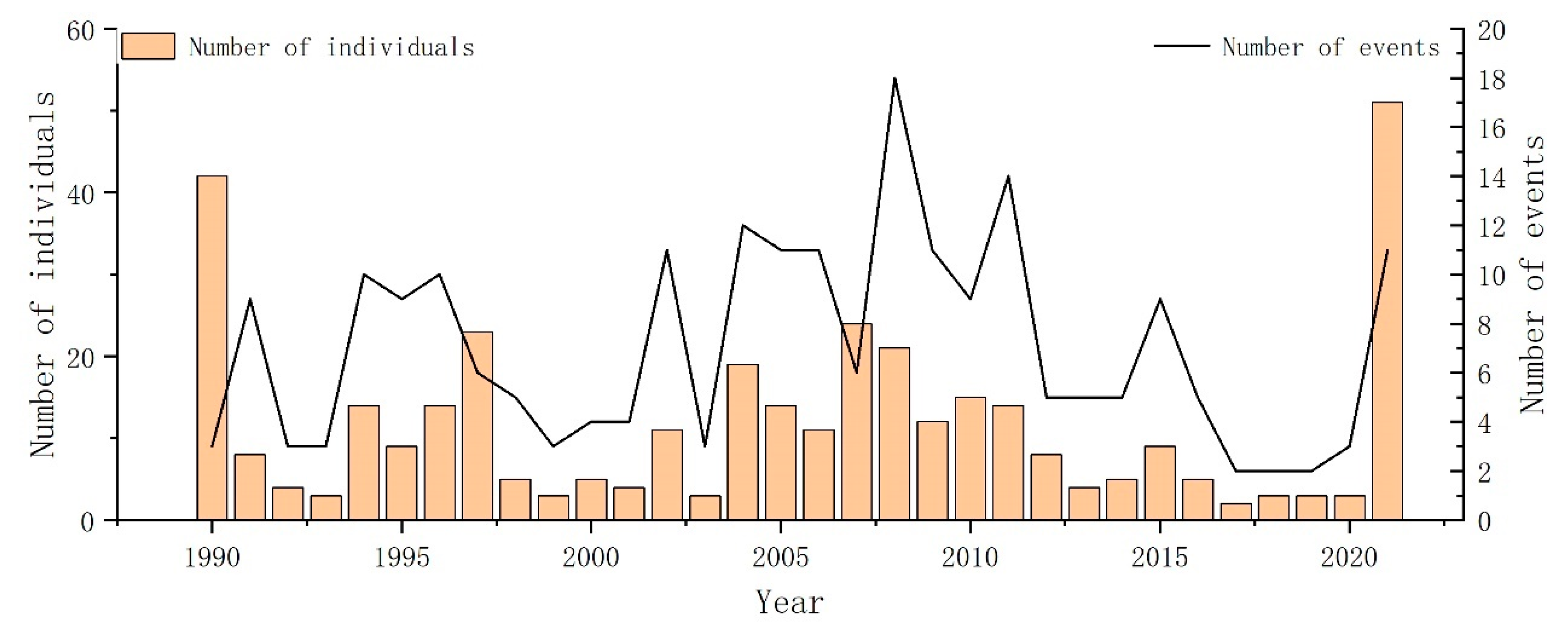{kind=link}
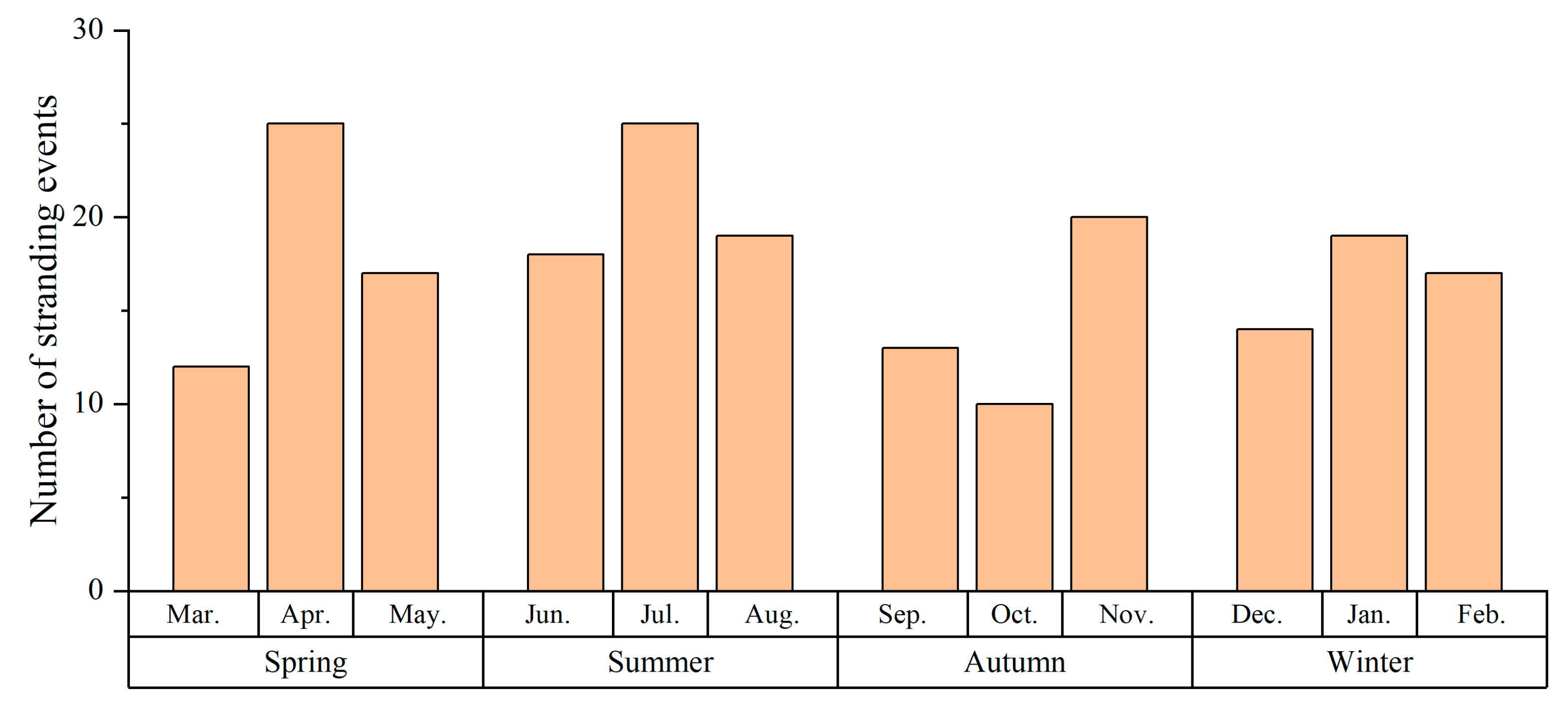{kind=link}
{kind=link}
{kind=link}
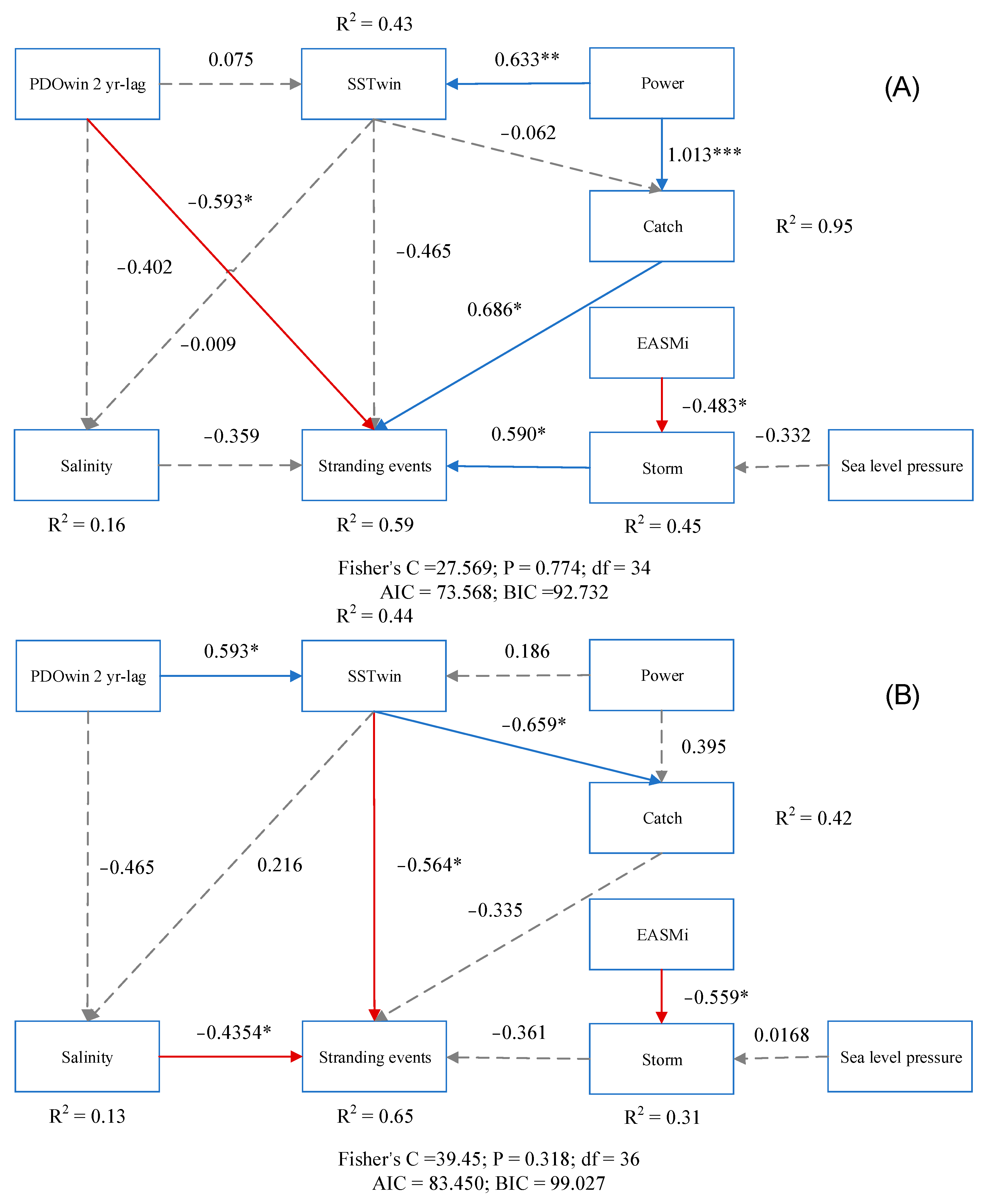{kind=link}
Table 1.
Environmental variables used in our structural equation model.
| Variable | Data | Sources * |
|---|---|---|
| Pacific Decadal Oscillation | A pattern of climate variability in the Pacific Ocean characterized by long-term changes in sea surface temperatures and atmospheric conditions | Climate Prediction Center, NOAA |
| The East Asian summer monsoon index | A measure of the strength and activity of the summer monsoon in East Asia | National Tibetan Plateau Data Center |
| Sea Level Pressure | The atmospheric pressure at sea level | The Fleet Numerical Meteorology and Oceanography Center (FNMOC), NOAA |
| Storm surge | Count of annual storm surge occurrences | Bulletin of China marine disaster |
| Sea surface temperature | The measurement of ocean surface temperature | Met Office: HadISST |
| Sea surface Salinity | The measurement of salt content in the uppermost layer of the ocean’s surface | Naval Oceanographic Office, NOAA NCEI |
| Power of fishing vessel | Annual total power of fishing vessels | China Fisheries Yearbook from 1990–2021 |
| Yearly fishing catch | Total yearly catch data for 40 fish species | China Fisheries Yearbook from 1990–2021 |
* NOAA: National Oceanic and Atmospheric Administration, NCEI: National Centers for Environmental Information.
Table 2.
Stranded cetaceans from 1990 to 2021 along coasts of East China Sea.
| Common Name | Scientific Name | Family | Number of Events |
|---|---|---|---|
| Short-finned pilot whale | Globicephala macrorhynchus Gray, 1846 | Delphinidae | 27 |
| Risso’s dolphin | Grampus griseus G. Cuvier, 1812 | Delphinidae | 14 |
| Pygmy killer whale | Feresa attenuata Gray, 1874 | Delphinidae | 9 |
| Indo-Pacific humpback dolphin | Sousa chinensis Osbeck, 1765 | Delphinidae | 9 |
| Pantropical spotted dolphin | Stenella attenuata Gray, 1846 | Delphinidae | 9 |
| Melon-headed whale | Peponocephala electra Gray, 1846 | Delphinidae | 7 |
| Fraser’s dolphin | Lagenodelphis hosei Fraser, 1956 | Delphinidae | 6 |
| Rough-toothed dolphin | Steno bredanensis G. Cuvier in Lesson, 1828 | Delphinidae | 5 |
| Common bottlenose dolphin | Tursiops truncatus Montagu, 1821 | Delphinidae | 5 |
| False killer whale | Pseudorca crassidens Owen, 1846 | Delphinidae | 3 |
| Spinner dolphin | Stenella longirostris Gray, 1828 | Delphinidae | 2 |
| Common minke whale | Balaenoptera acutorostrata Lacépède, 1804 | Balaenopteridae | 13 |
| Omura‘s whale | Balaenoptera omurai Wada, Oishi & Yamada, 2003 | Balaenopteridae | 12 |
| Bryde’s whale | Balaenoptera edeni Anderson, 1878 | Balaenopteridae | 11 |
| Fin whale | Balaenoptera physalus Linnaeus, 1758 | Balaenopteridae | 4 |
| Sei whale | Balaenoptera borealis Lesson, 1828 | Balaenopteridae | 3 |
| Humpback whale | Megaptera novaeangliae Borowski, 1781 | Balaenopteridae | 3 |
| Cuvier’s beaked whale | Ziphius cavirostris Cuvier, 1823 | Ziphiidae | 10 |
| Blainville’s beaked whale | Mesoplodon densirostris de Blainville, 1817 | Ziphiidae | 9 |
| Longman’s beaked whale | Mesoplodon pacificus Longman, 1926 | Ziphiidae | 2 |
| Ginkgo-toothed beaked whale | Mesoplodon ginkgodens Baker & van Helden, 1999 | Ziphiidae | 1 |
| Narrow-ridged finless porpoise | Neophocaena asiaeorientalis Pilleri and Gihr, 1972 | Phocoenidae | 40 |
| Indo-Pacific finless porpoise | Neophocaena phocaenoides Cuvier, 1829 | Phocoenidae | 2 |
| Dwarf sperm whale | Kogia sima Owen, 1866 | Kogiidae | 13 |
| Pygmy sperm whale | Kogia breviceps de Blainville, 1838 | Kogiidae | 9 |
| Sperm whale | Physeter macrocephalus Linnaeus, 1758 | Physeteridae | 9 |
| North Pacific right whale | Eschrichtius robustus Lilljeborg, 1861 | Eschrichtiidae | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yang, S.; Li, S.; Jin, Y.; Liu, Z. Changing Trends in Cetacean Strandings in the East China Sea: Identifying Relevant Variables and Implications for Conservation and Management. Diversity 2023, 15, 1082. https://doi.org/10.3390/d15101082
AMA Style
Yang S, Li S, Jin Y, Liu Z. Changing Trends in Cetacean Strandings in the East China Sea: Identifying Relevant Variables and Implications for Conservation and Management. Diversity. 2023; 15(10):1082. https://doi.org/10.3390/d15101082
Chicago/Turabian StyleYang, Shaobo, Shengfa Li, Yan Jin, and Zunlei Liu. 2023. "Changing Trends in Cetacean Strandings in the East China Sea: Identifying Relevant Variables and Implications for Conservation and Management" Diversity 15, no. 10: 1082. https://doi.org/10.3390/d15101082
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.