
The Denticle Multiverse: Morphological Diversity of Placoid Scales across Ontogeny in the Portuguese Dogfish, Centroscymnus coelolepis, and Its Systematic Implications

Abstract
:1. Introduction
2. Materials and Methods
2.1. Examined Specimens and Skin Samples
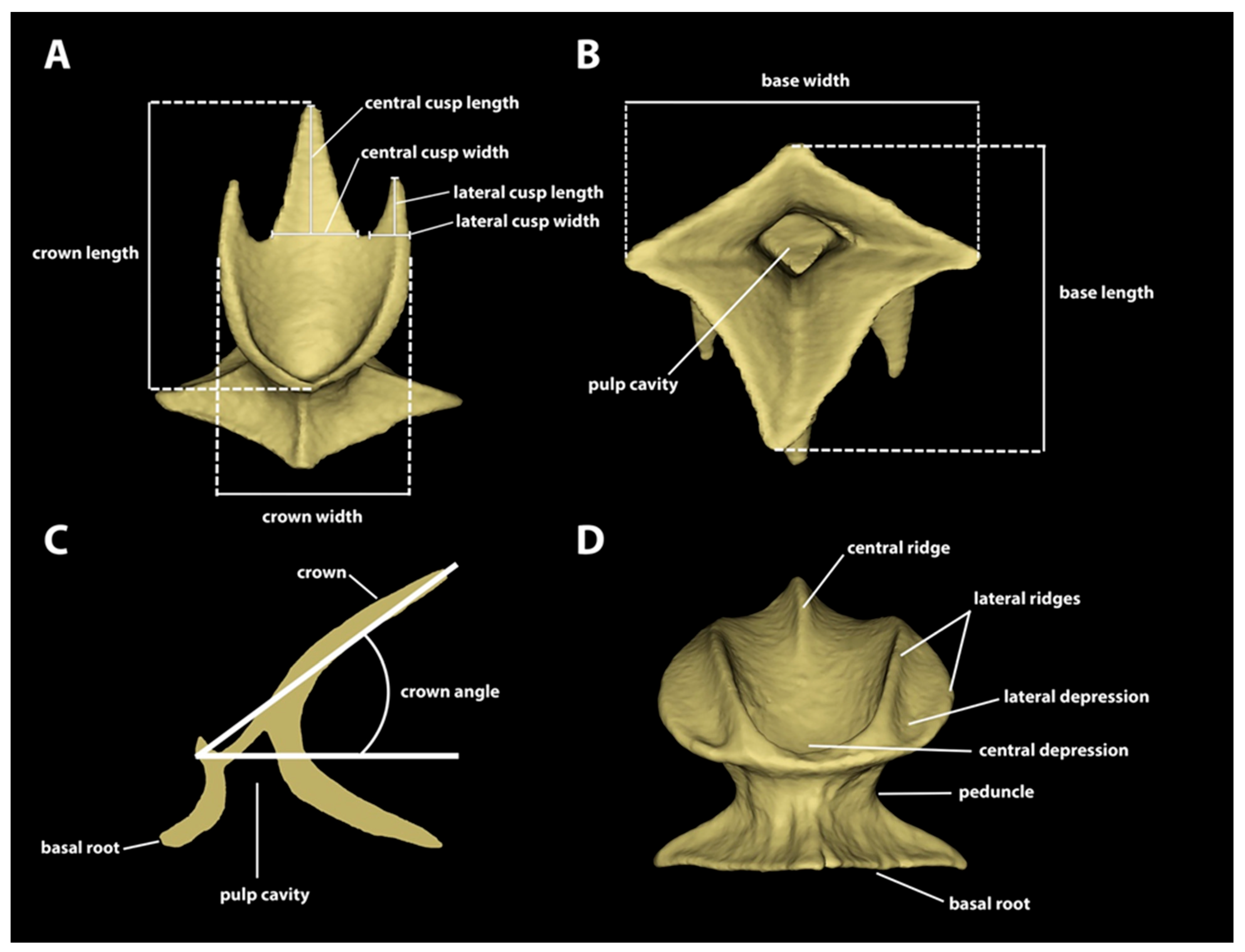
2.2. µCT Scanning Analysis and Dermal-Denticle Measurements
2.3. Statistics
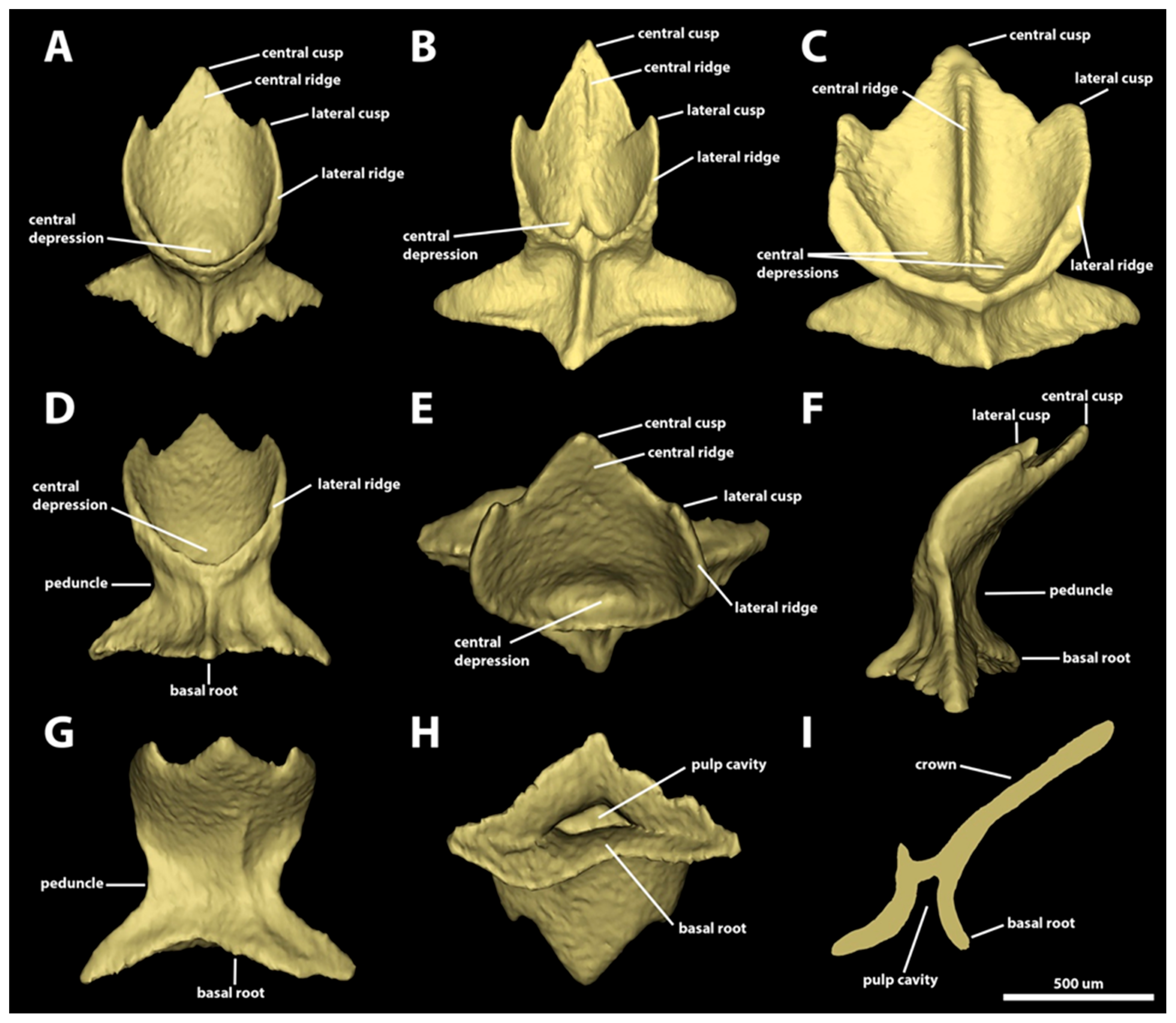
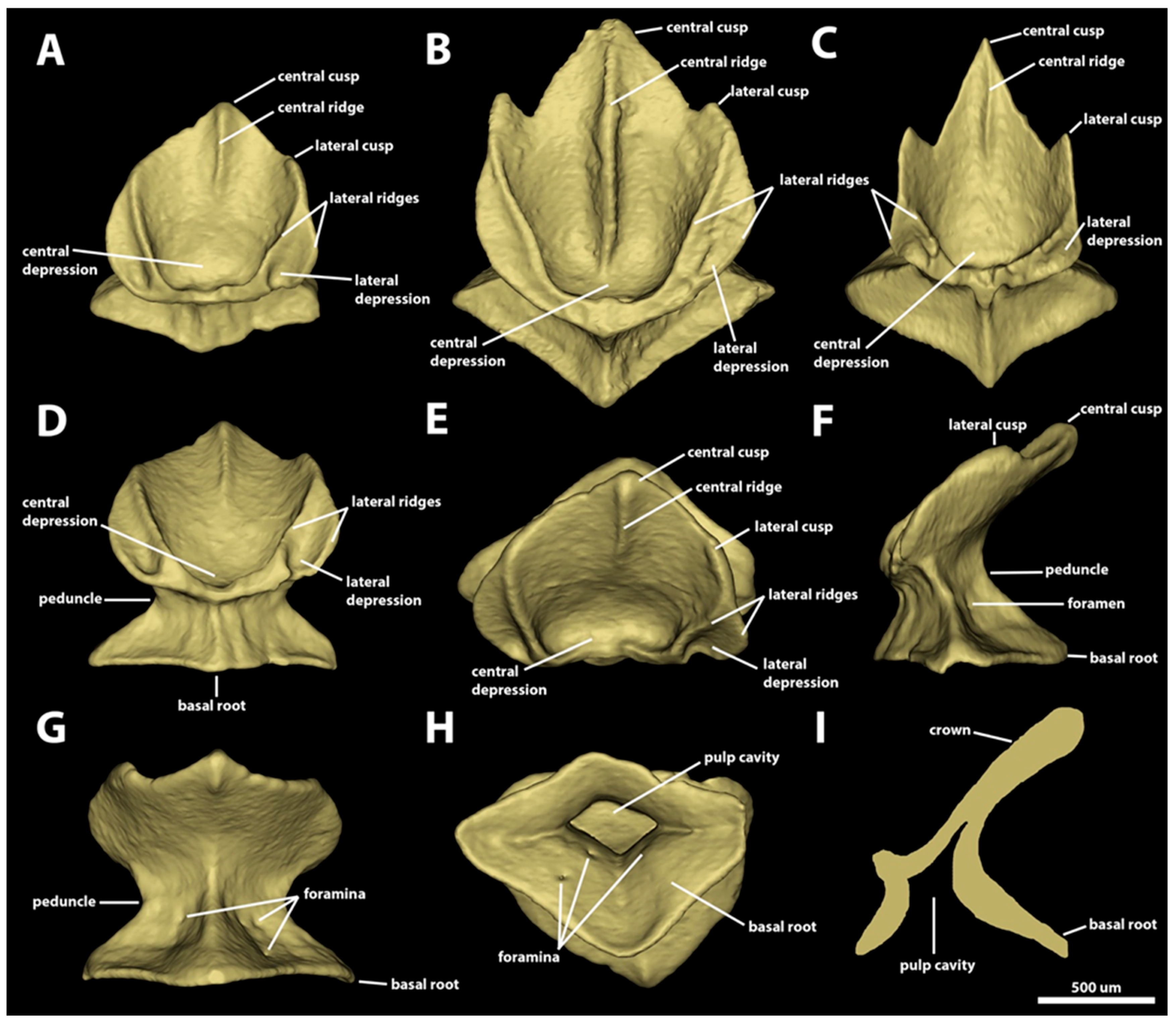
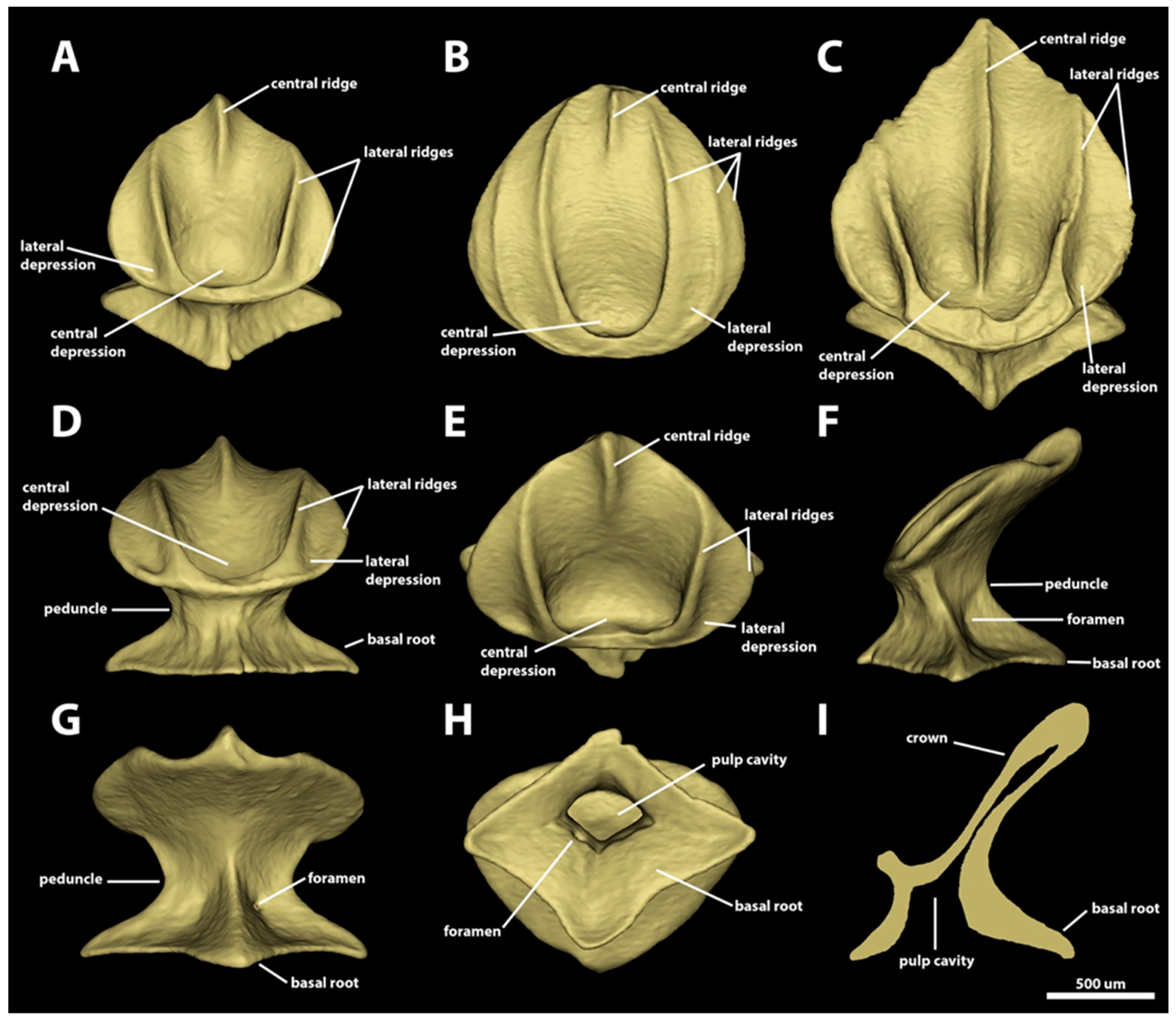
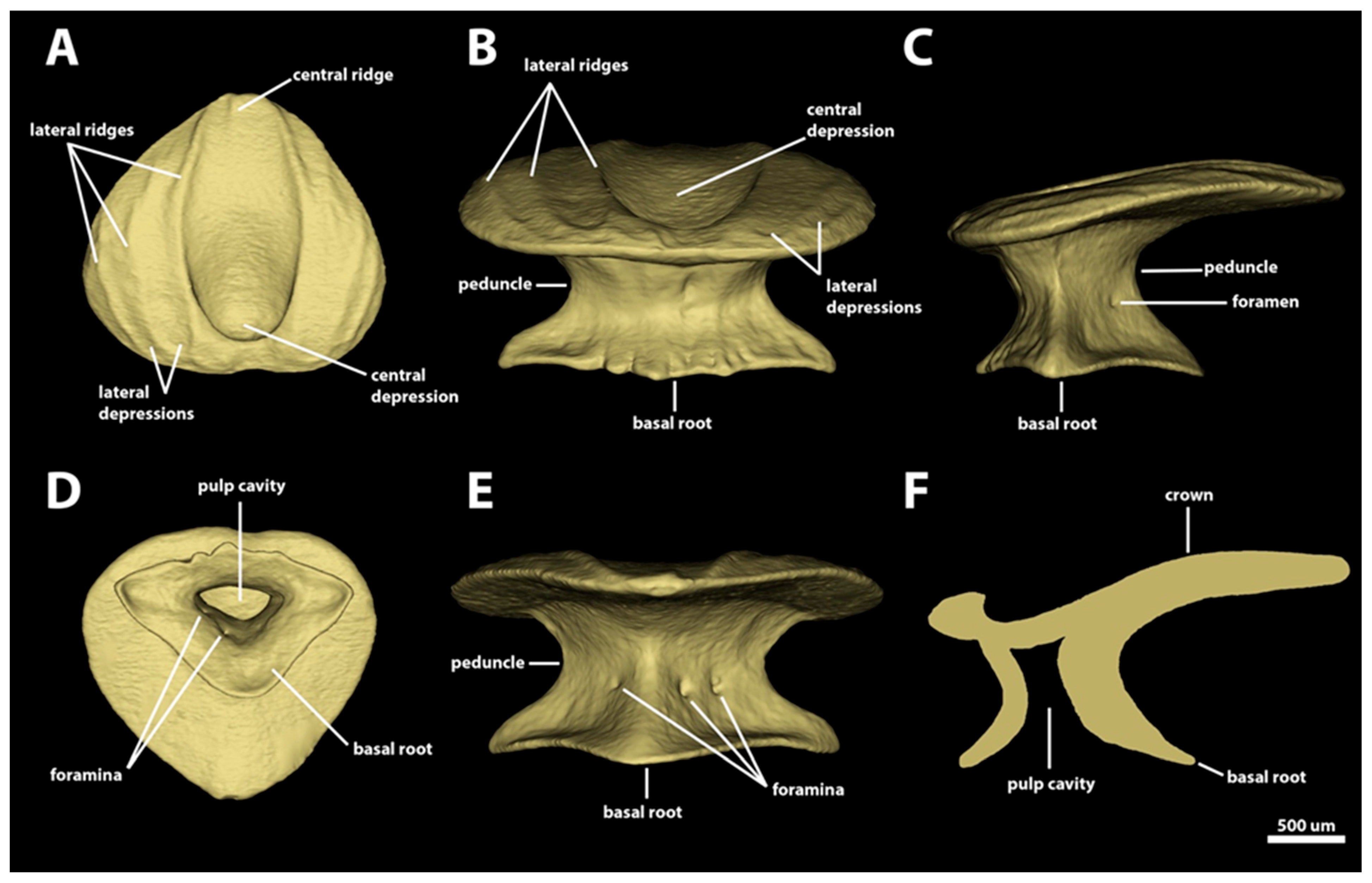
3. Results
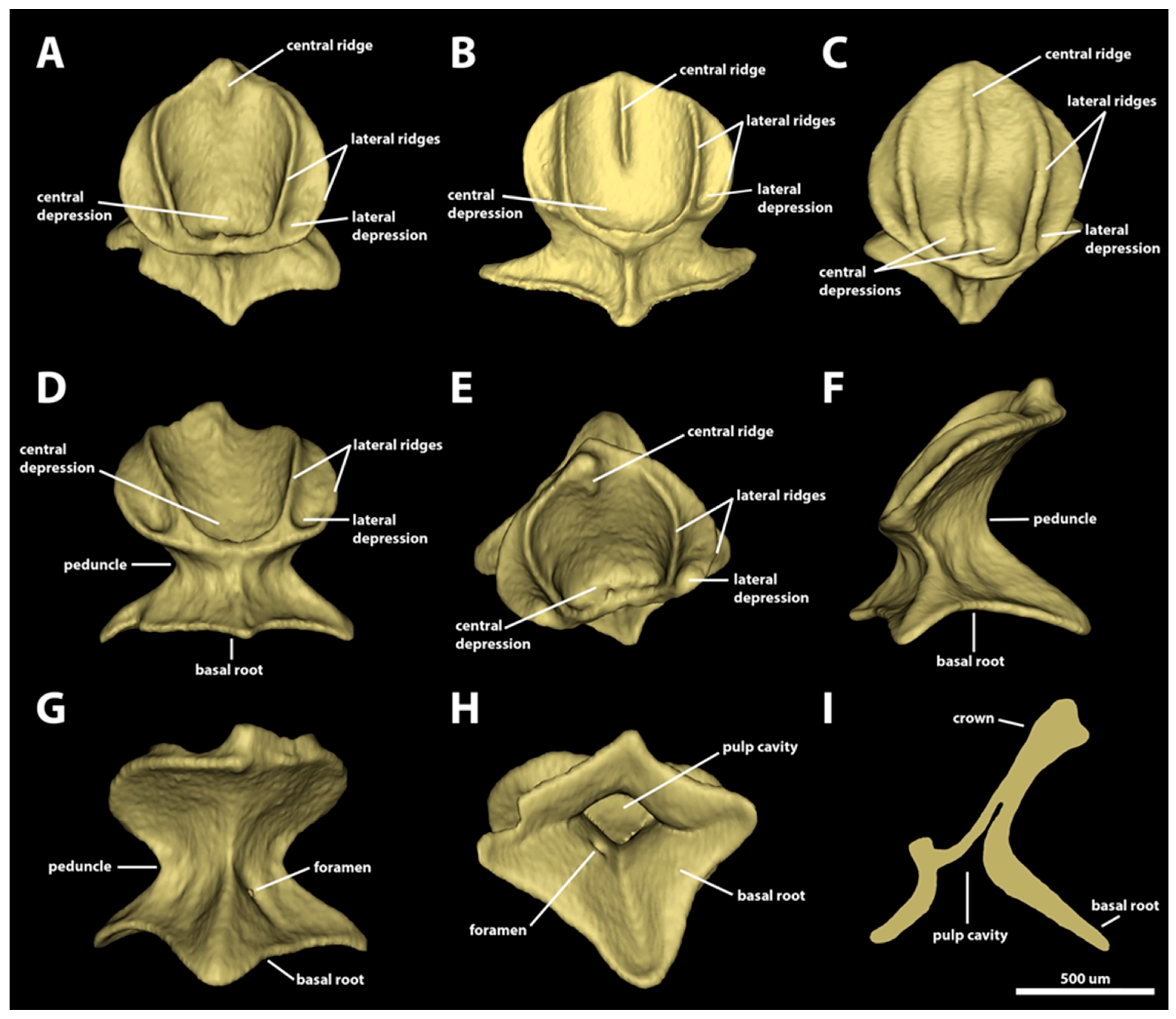
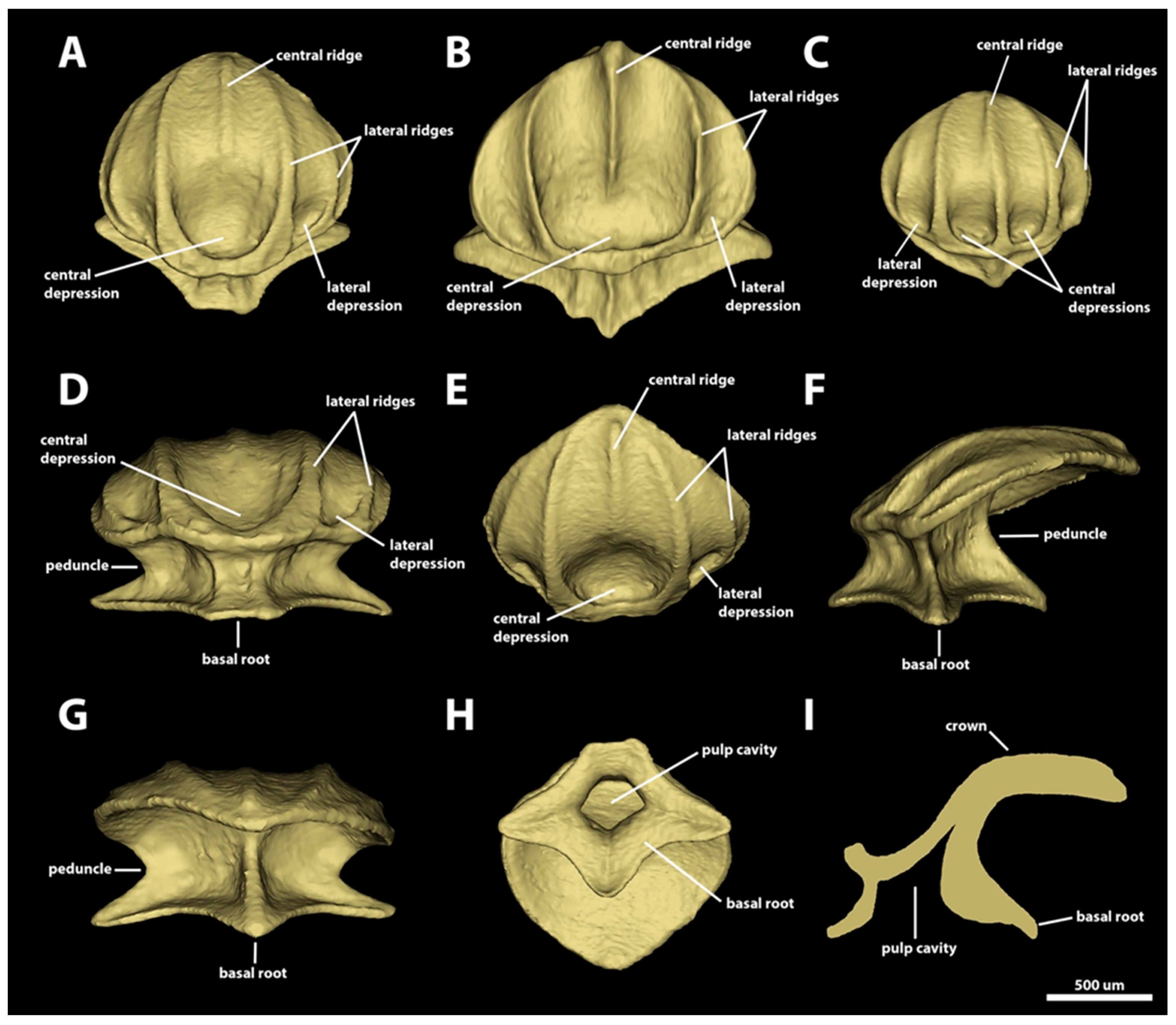
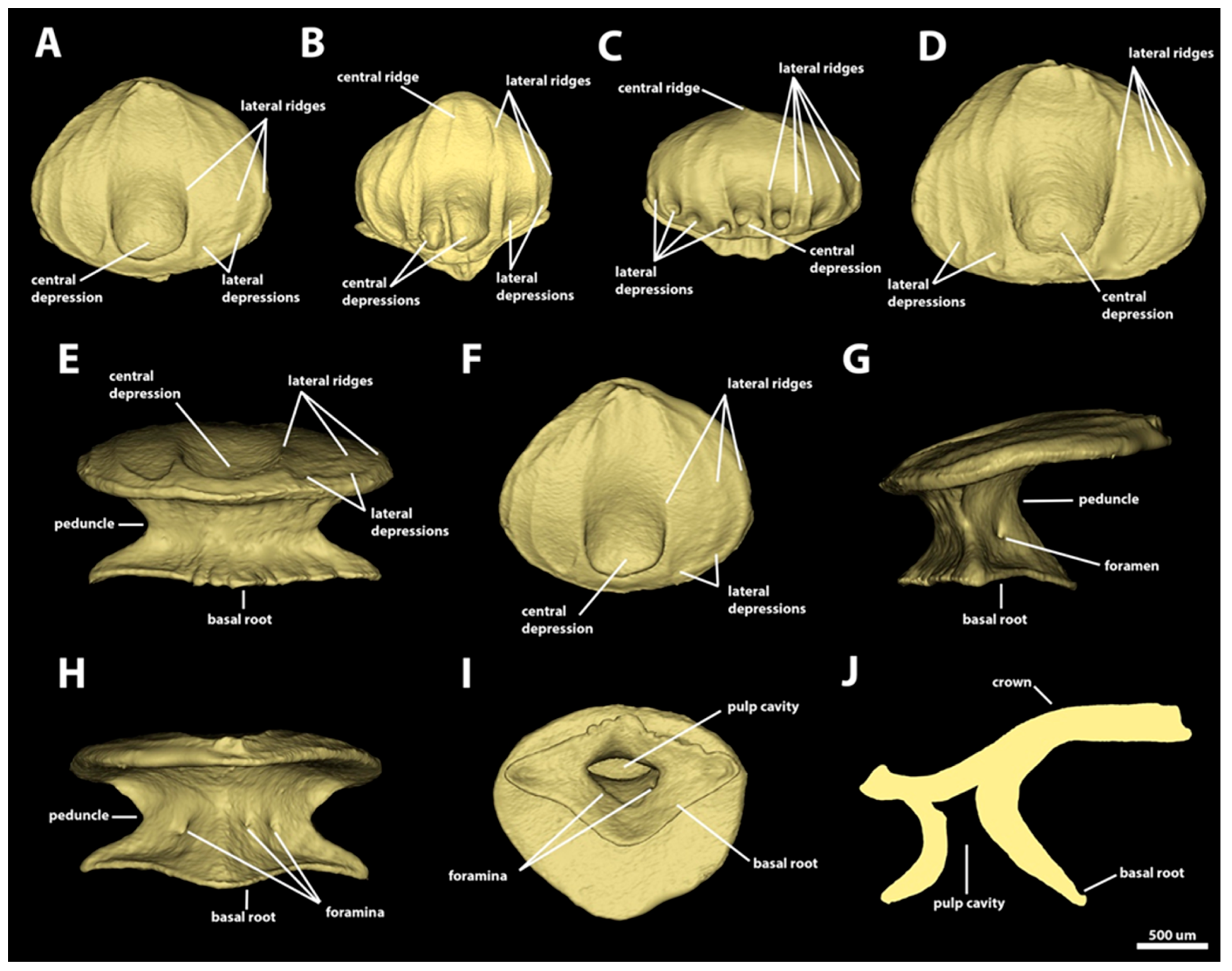
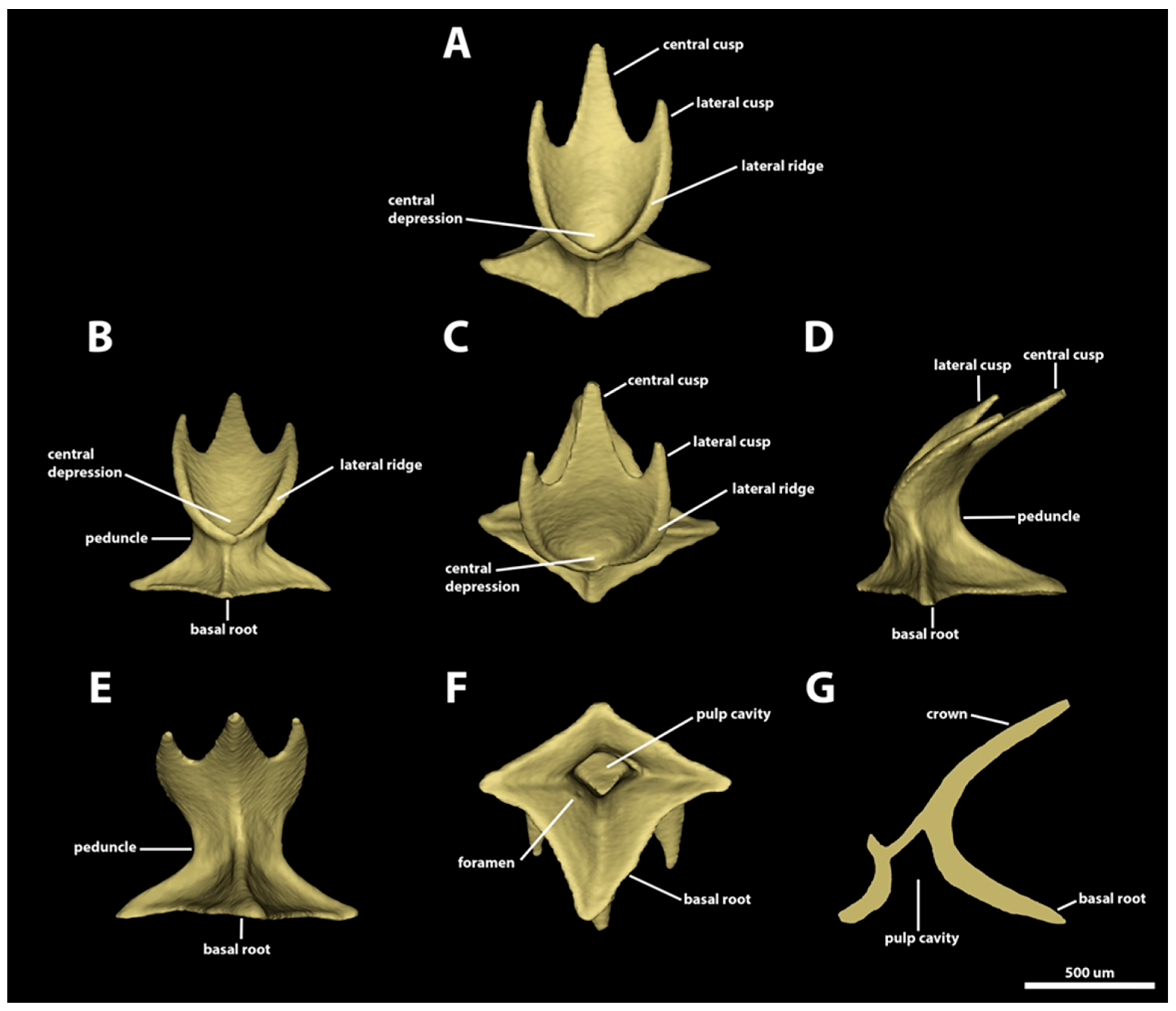
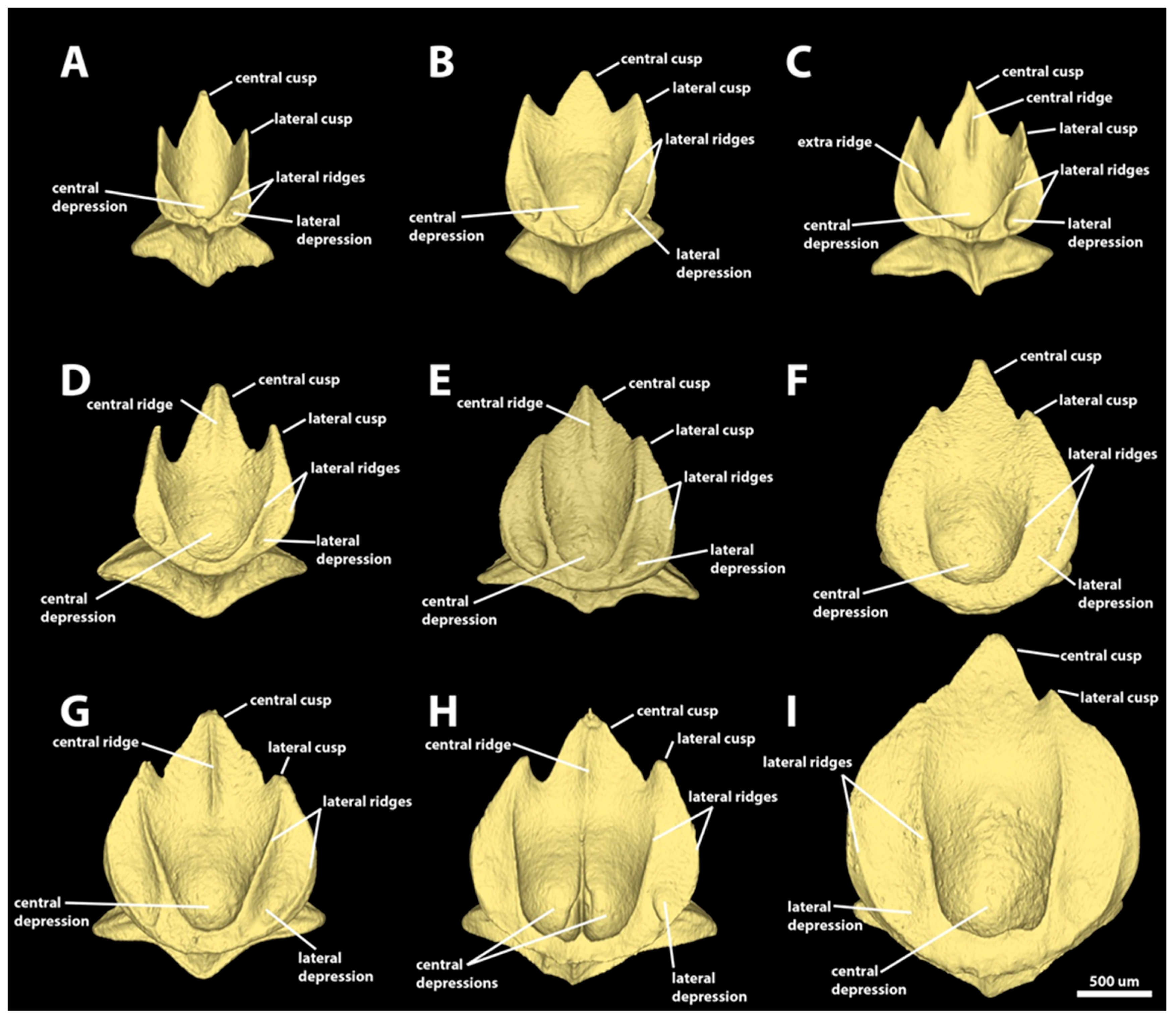
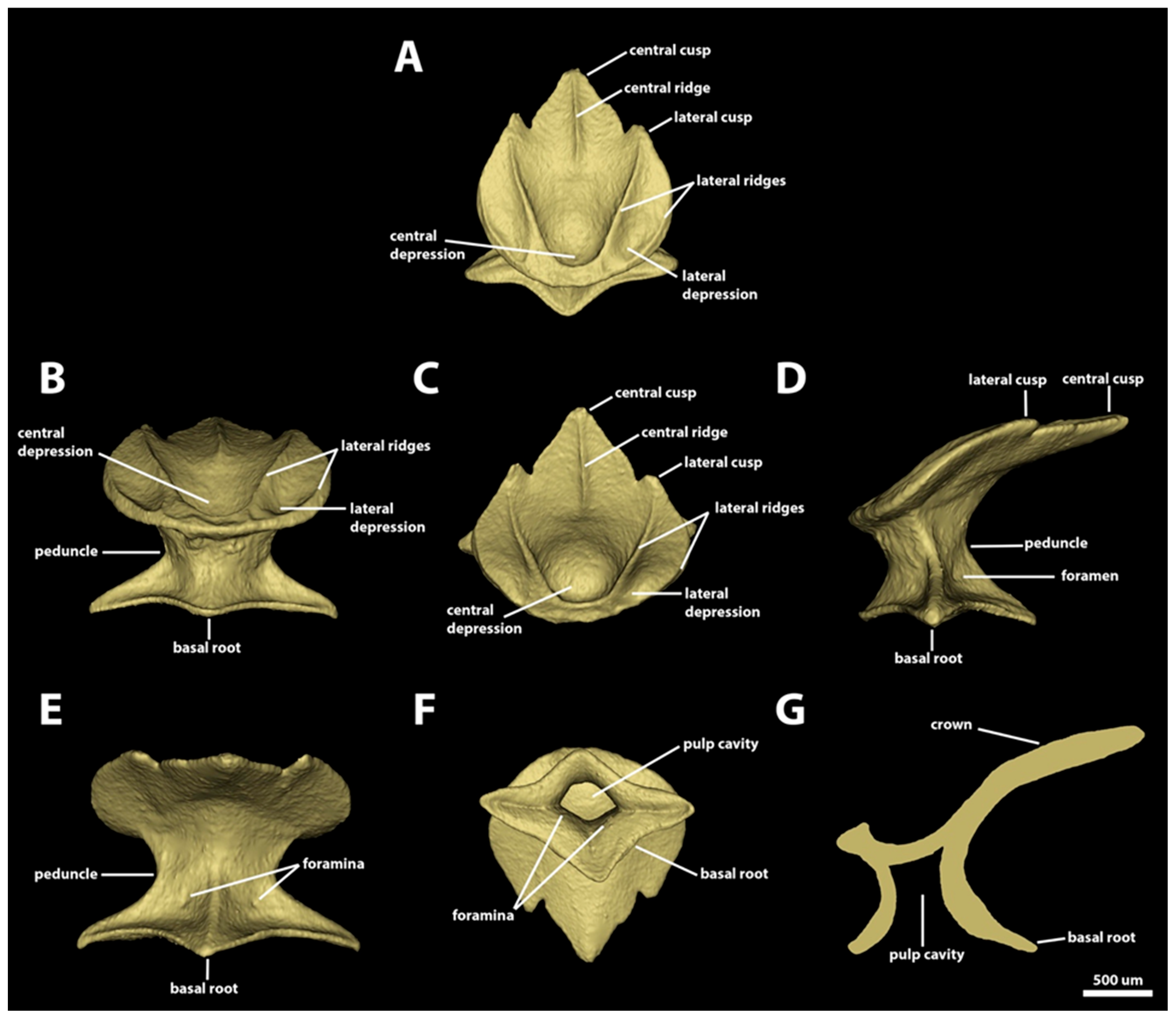
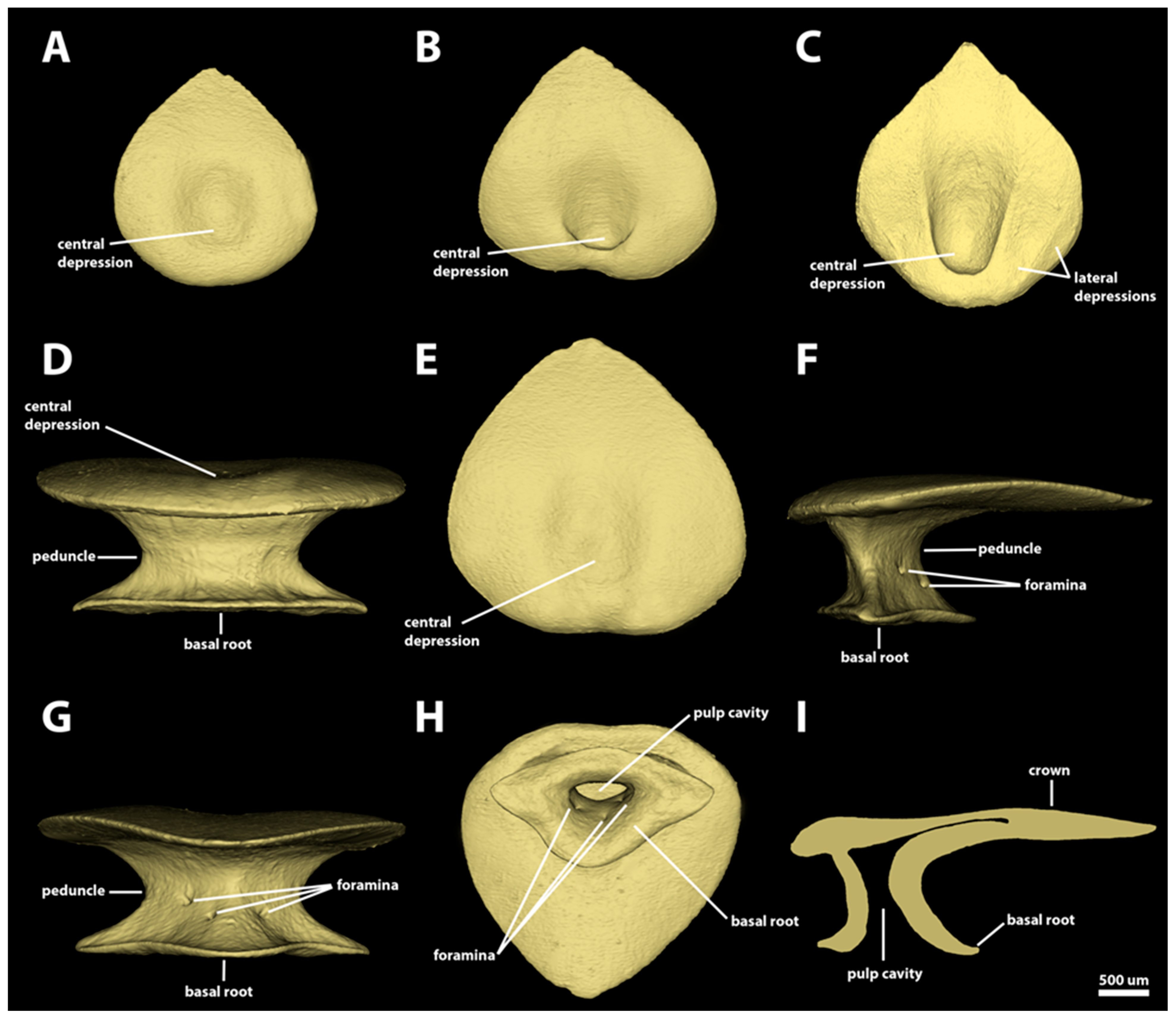
3.1. Denticle Morphotypes
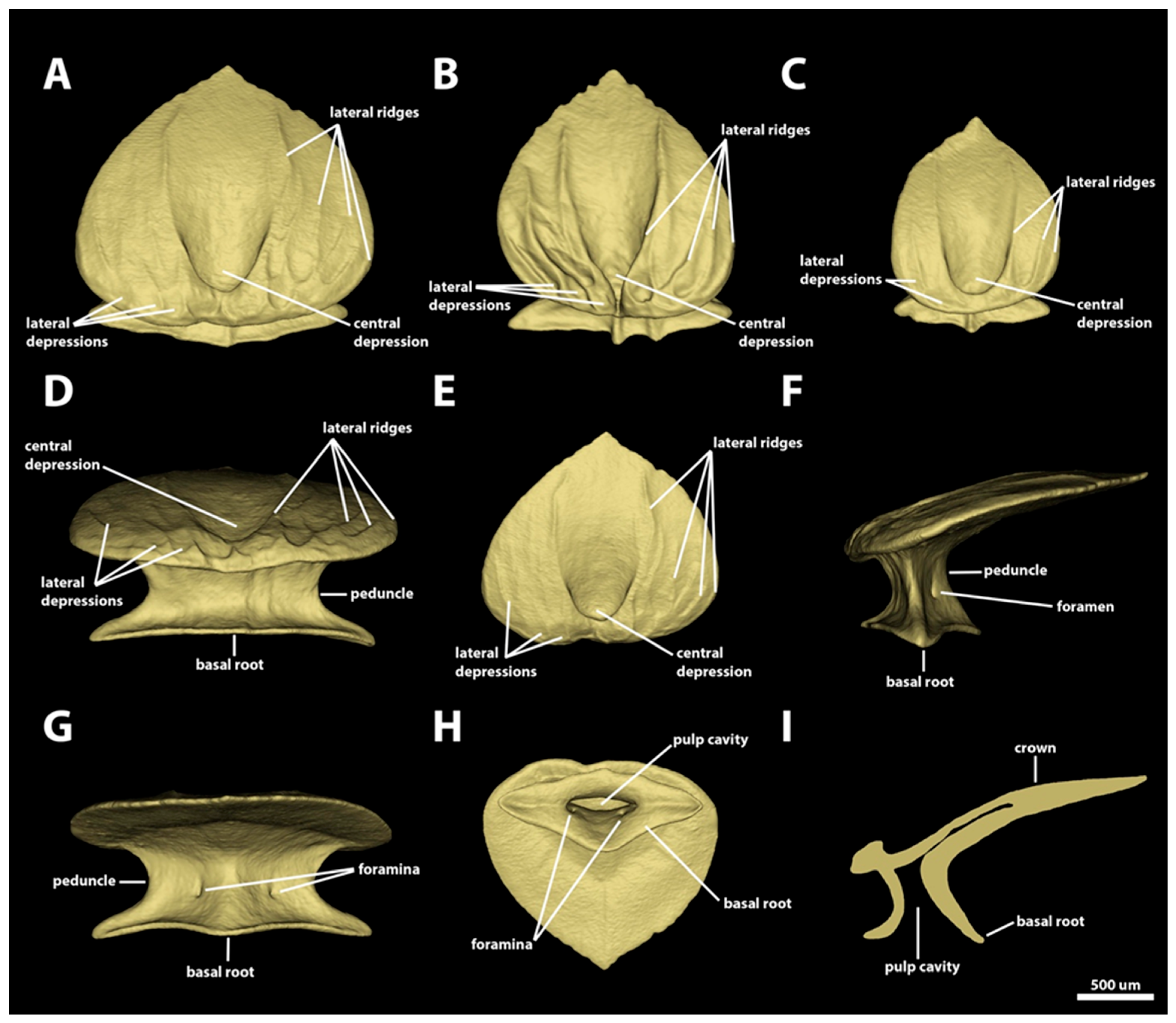
3.1.1. Snout
3.1.2. Spiracular Region
3.1.3. Branchial, Trunk, and Tail Regions
3.2. Denticle Replacement
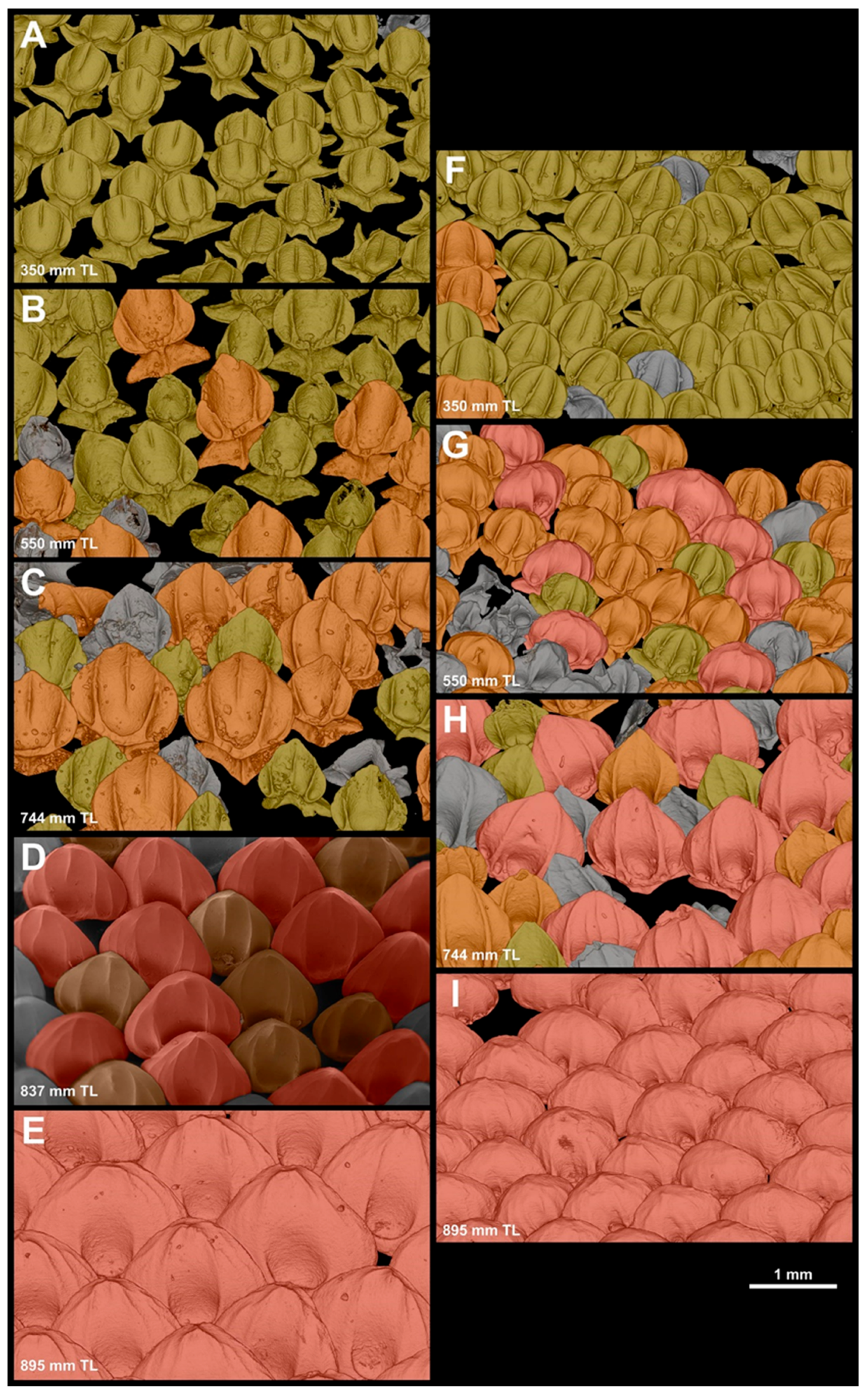
3.2.1. Denticle Replacement in the Snout
3.2.2. Denticle Replacement in the Spiracular Region
3.2.3. Denticle-Replacement Patterns in the Branchial, Abdominal, and Caudal Regions
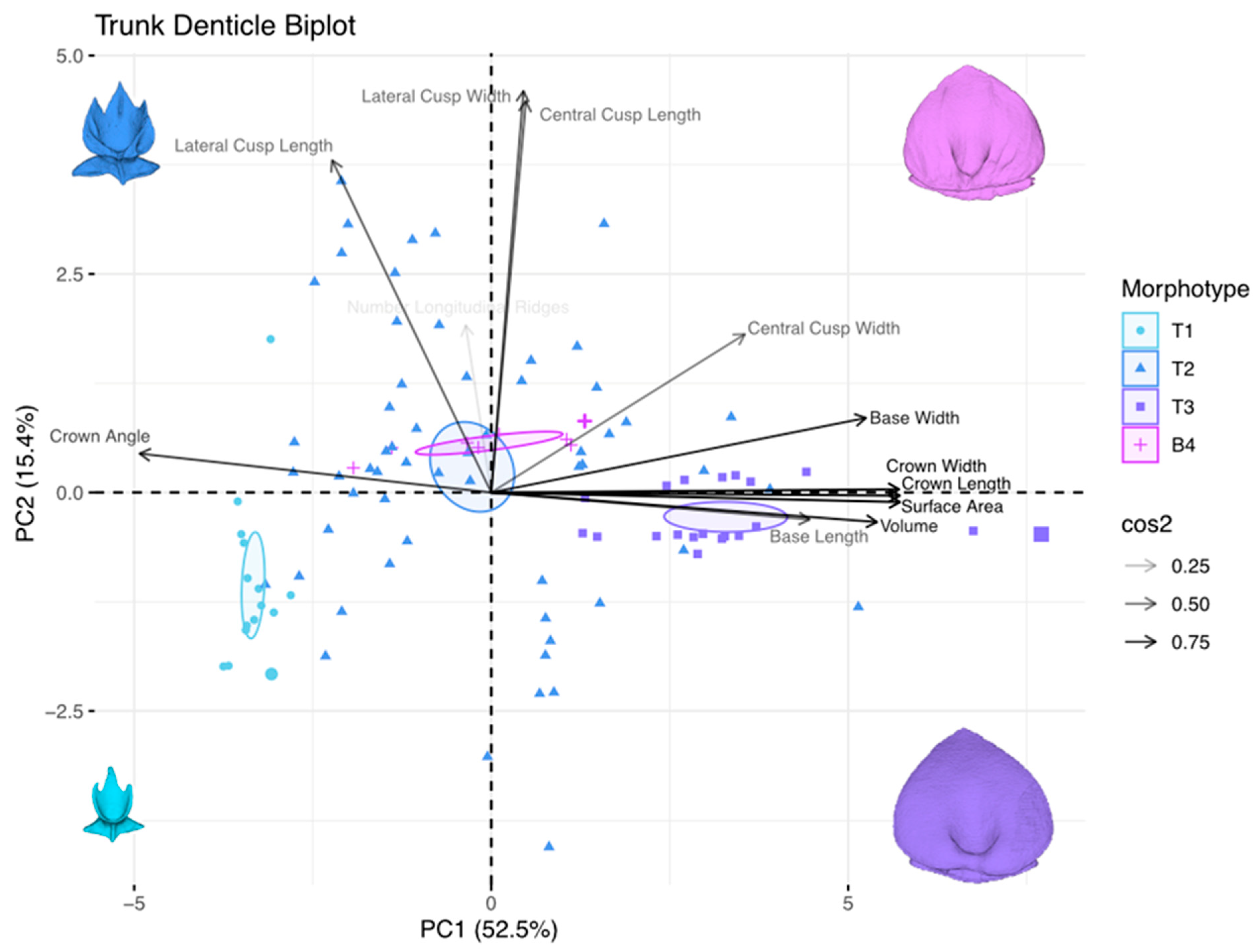
3.3. Size Variation within Each Dermal-Denticle Morphotype
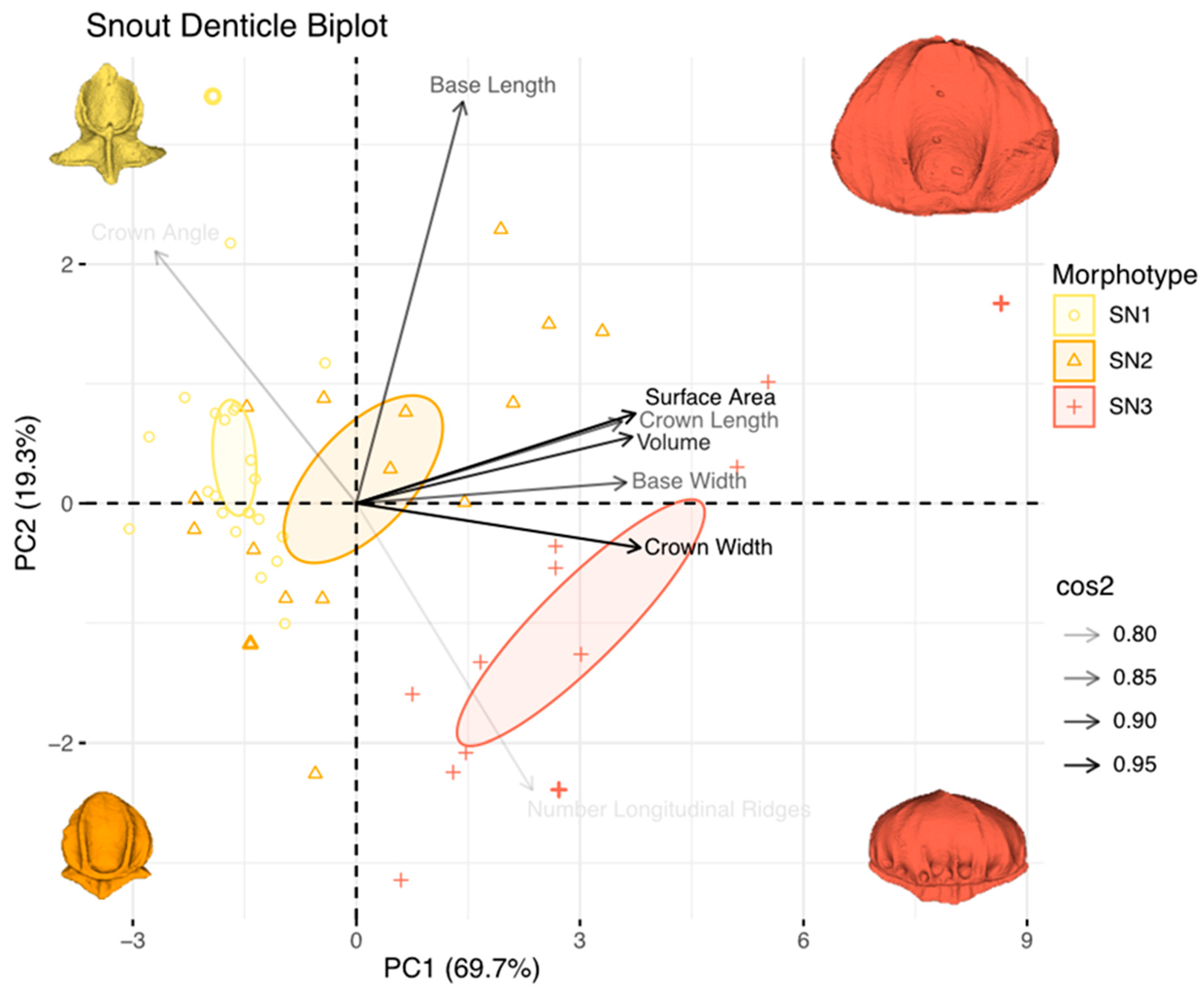
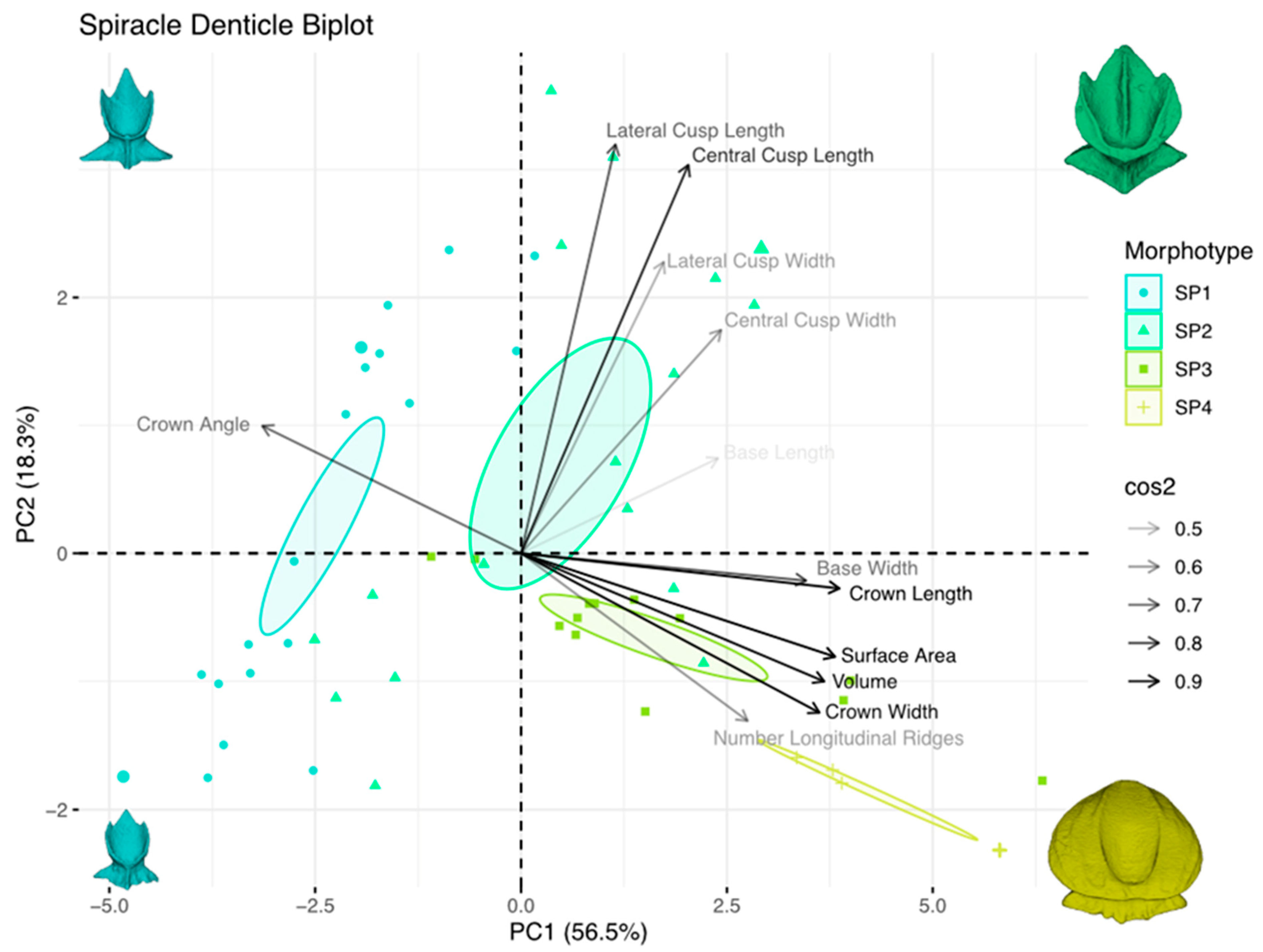
3.4. Principal Component Analysis of Dermal-Denticle Morphotypes
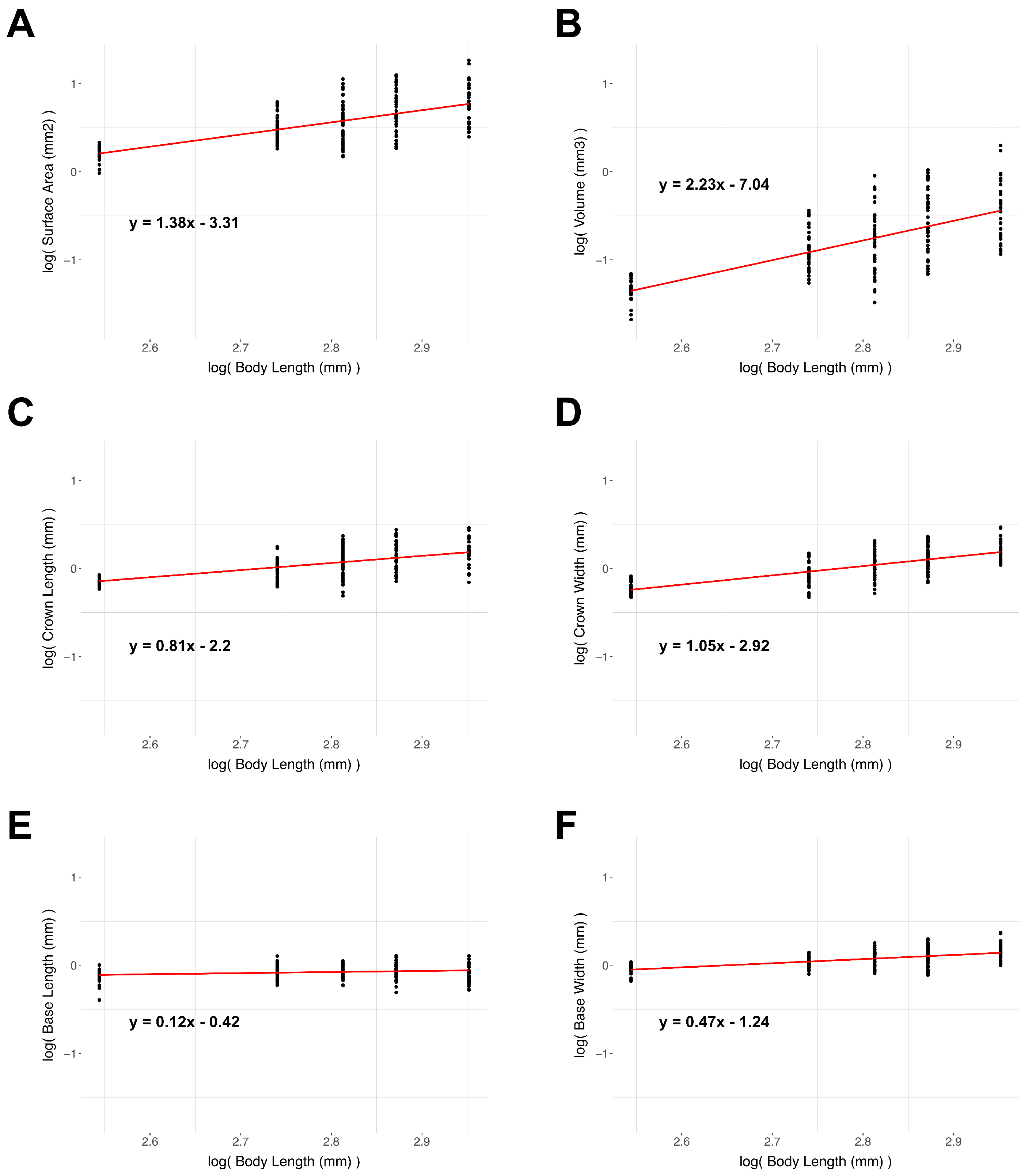
3.5. Denticle Scaling
4. Discussion
4.1. Denticle Morphology in Centroscymnus coelolepis
4.2. Denticles and the Taxonomy of Centroscymnus coelolepis and C. owstonii
4.3. Monophyly of Centroscymnus: Additional Support from Dermal Denticles
4.4. Ontogenetic Variation and Phylogenetic Inference within the Family Somniosidae
4.5. Denticle Replacement and Function
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Complete List of Examined Specimens
References
- Naylor, G.J.; Caira, J.N.; Jensen, K.; Rosana, K.A.; Straube, N.; Lakner, C. Elasmobranch phylogeny: A mitochondrial estimate based on 595 species. In The Biology of Sharks and Their Relatives; Carrier, J.C., Musick, J.A., Heithaus, M.R., Eds.; CRC Press: Boca Raton, FL, USA, 2012; pp. 31–56. [Google Scholar]
- Straube, N.; Li, C.; Claes, J.M.; Corrigan, S.; Naylor, G.J.P. Molecular phylogeny of Squaliformes and first occurrence of bioluminescence in sharks. BMC Evol. Biol. 2015, 15, 162. [Google Scholar] [CrossRef]
- Ebert, D. Deep-sea cartilaginous fishes of the Indian Ocean. Volume 1. Sharks. FAO Species Cat. Fish. Purp. 2013. [Google Scholar]
- Ebert, D.A.; Dando, M.; Fowler, S. Sharks of the World: A Complete Guide; Princeton University Press: Princeton, NJ, USA, 2021. [Google Scholar]
- White, W.T.; Vaz, D.F.B.; Ho, H.; Ebert, D.A.; de Carvalho, M.R.; Corrigan, S.; Rochel, E.; de Carvalho, M.; Tanaka, S.; Naylor, G.J.P. Redescription of Scymnodon ichiharai Yano and Tanaka 1984 (Squaliformes: Somniosidae) from the western North Pacific, with comments on the definition of somniosid genera. Ichthyol. Res. 2015, 62, 213–229. [Google Scholar] [CrossRef]
- Weigmann, S.; Vaz, D.F.B.; White, W.T.; de Carvalho, M.R.; Thiel, R. Distribution and comments on the morphology of Centroscymnus owstonii Garman, 1906 (Squaliformes: Somniosidae), with focus on its occurrence in the Indian Ocean. Mar. Biodivers. 2016, 46, 641–653. [Google Scholar] [CrossRef]
- Vaz, D.F.B. Scymnodon plunketi (Waite, 1910): A junior synonym of Scymnodon macracanthus (Regan, 1906) (Somniosidae: Elasmobranchii). J. Fish Biol. 2021, 99, 472–494. [Google Scholar] [CrossRef] [PubMed]
- Vaz, D.F.B. Revisão Taxonômica e Morfológica do Gênero Centroscymnus Barboza du Bocage & Britto-Capello, 1864, com Comentários no Arranjo Genérico da Família Somniosidae (Chondrichthyes: Squaliformes); Universidade de São Paulo: São Paulo, Brazil, 2015. [Google Scholar]
- Yano, K.; Tanaka, S. Portuguese shark, Centroscymnus coelolepis from Japan, with notes on C. owstoni. Jpn. J. Ichthyol. 1983, 30, 208–216. [Google Scholar]
- Yano, K.; Tanaka, S. Review of the deep sea squaloid shark genus Scymnodon of Japan, with a description of a new species. Jpn. J. Ichthyol. 1984, 30, 341–360. [Google Scholar]
- Compagno, L.J.V.; Dando, M.; Fowler, S.A. Field Guide to the Sharks of the World; Harper Collins Publishers Ltd.: London, UK, 2005. [Google Scholar]
- Didier, D.; Kemper, J.M.; Ebert, D.A. Phylogeny, Biology, and Classification of Extant Holocephalans. In Biology of Sharks and Their Relatives, 2nd ed.; Carrier, J.C., Musick, J.A., Heithaus, M.R., Eds.; CRC Press: Boca Raton, FL, USA, 2012; pp. 97–122. [Google Scholar]
- Last, P.R.; White, W.T.; de Carvalho, M.R.; Séret, B.; Stehmann, M.F.W.; Naylor, G.J.P. Rays of the World; CSIRO Publishing: Clayton South, Australia, 2016. [Google Scholar]
- Donoghue, P.C.; Rücklin, M. The ins and outs of the evolutionary origin of teeth. Evol. Dev. 2016, 18, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Ankhelyi, M.; Wainwright, D.K.; Lauder, G.V. Diversity of dermal denticle structure in sharks: Skin surface roughness and three-dimensional morphology. J. Morphol. 2018, 279, 1132–1154. [Google Scholar] [CrossRef]
- Sibert, E.C.; Rubin, L.D. An early Miocene extinction in pelagic sharks. Science 2021, 372, 1105. [Google Scholar] [CrossRef] [PubMed]
- Reif, W.-E. Squamation and Ecology of Sharks. Cour. Forsch.-Inst. Senckenberg 1985, 78, 1–255. [Google Scholar]
- Taniuchi, T.; Garrick, J.A.F. A new species of Scymnodalatias from southern oceans, and comments on other Squaliform sharks. Jpn. J. Ichthyol. 1986, 33, 119–134. [Google Scholar] [CrossRef]
- Deynat, P.P.; Séret, B. Le revêtement cutané des raies (Chondrichthyes, Elasmobranchii, Batoidea). Morphologie et arrangement des denticules cutanés. Ann. Des Sci. Nat. Zool. 1996, 17, 65–83. [Google Scholar]
- Poscai, A.N.; Silva, J.P.C.B.; Casas, A.L.; Lenktaitis, P.; Gadig, O.B.F. Morphological study of the oral denticles of the porbeagle shark Lamna nasus. J. Fish Biol. 2022, 101, 226–235. [Google Scholar] [CrossRef]
- Reif, W.-E. Morphology and hydrodynamic effects of the scales of fast swimming sharks. Fortschr. Zool. 1985, 30, 483–485. [Google Scholar]
- Oeffner, J.; Lauder, G.V. The hydrodynamic function of shark skin and two biomimetic applications. J. Exp. Biol. 2012, 215, 785–795. [Google Scholar] [CrossRef] [PubMed]
- Lauder, G.V.; Wainwright, D.K.; Domel, A.G.; Weaver, J.; Wen, L.; Bertoldi, K. Structure, biomimetics, and fluid dynamics of fish skin surfaces. Phys. Rev. Fluids 2016, 1, 060502. [Google Scholar] [CrossRef]
- Raschi, W.; Tabit, C. Functional aspects of placoid scales: A review and update. Aust. J. Mar. Fresh. Res. 1992, 43, 123–147. [Google Scholar] [CrossRef]
- Reif, W. Protective and hydrodynamic function of the dermal skeleton of elasmobranchs. Neues Jahrb. Für Geol. Und Paläontologie Abh. 1978, 157, 131–141. [Google Scholar]
- Reif, W.-E. Wound healing in sharks. Zoomorphologie 1978, 90, 101–111. [Google Scholar] [CrossRef]
- Dillon, E.M.; Bagla, A.; Plioplys, K.D.; McCauley, D.J.; Lafferty, K.D.; O’Dea, A. Dermal denticle shedding rates vary between two captive shark species. Mar. Ecol. Prog. Ser. 2022, 682, 153–167. [Google Scholar] [CrossRef]
- Vaz, D.F.; De Carvalho, M.R. Morphological and taxonomic revision of species of Squatina from the Southwestern Atlantic Ocean (Chondrichthyes: Squatiniformes: Squatinidae). Zootaxa 2013, 3695, 1–81. [Google Scholar] [CrossRef] [PubMed]
- Bigelow, H.B.; Schroeder, W.C. Deep water elasmobranchs and chimaeroids from north-western Atlantic slope. Bull. Mus. Comp. Zool. 1954, 112, 37–87. [Google Scholar]
- Garrick, J.A.F. The systematic position of Centroscymnus waitei (Thompson, 1930), Selachii. Trans. R. Soc. New Zealand 1955, 83, 227–239. [Google Scholar]
- Garrick, J.A.F. Scymnodon plunketi (Waite, 1910), an abundant deep-water shark of New Zealand waters. Trans. R. Soc. New Zealand 1959, 87, 271–282. [Google Scholar]
- Cadenat, J.; Blache, J. Requins de Mediterranée et d’ Atlantique. Faune Tropicale. ORSTOM 1981, 21, 1–330. [Google Scholar]
- White, W.T.; Ebert, D.A.; Naylor, G.J.P.; Ho, H.C.; Clerkin, P.; Verissimo, A.; Cotton, C.F. Revision of the genus Centrophorus (Squaliformes: Centrophoridae): Part 1-Redescription of Centrophorus granulosus (Bloch & Schneider), a senior synonym of C. acus Garman and C. niaukang Teng. Zootaxa 2013, 3752, 35–72. [Google Scholar] [PubMed]
- White, W.T.; Ebert, D.A.; Naylor, G.J.P. Revision of the genus Centrophorus (Squaliformes: Centrophoridae): Part 2—Description of two new species of Centrophorus and clarification of the status of Centrophorus lusitanicus Barbosa du Bocage & de Brito Capello, 1864. Zootaxa 2019, 4344, 86–114. [Google Scholar]
- White, W.T.; Guallart, J.; Ebert, D.A.; Naylor, G.J.P.; Verissimo, A.; Cotton, C.F.; Harris, M.; Serena, F.; Iglesias, S.P. Revision of the genus Centrophorus (Squaliformes: Centrophoridae): Part 3—Redescription of Centrophorus uyato (Rafinesque) with a discussion of its complicated nomenclatural history. Zootaxa 2022, 5155, 1–51. [Google Scholar] [CrossRef]
- Garman, S. The Plagiostomia (sharks, skates, and rays). Mem. Mus. Comp. Zool. 1913, 36, 1–515. [Google Scholar]
- Bigelow, H.B.; Schroeder, W.C. Sharks. In Fishes of the Western North Atlantic; Tee-Van, J., Breder, C.M., Hildebrand, S.F., Parr, A.E., Schroeder, W.C., Eds.; Sears Foundation for Marine Research, Yale University: New Haven, CT, USA, 1948; pp. 59–546. [Google Scholar]
- Bigelow, H.B.; Schroeder, W.C. A study of the sharks of the suborder Squaloidea. Bull. Mus. Comp. Zool. 1957, 117, 1–150. [Google Scholar]
- Castro, J.I. The Sharks of North America; Oxford Univ. Press: Oxford, UK, 2011. [Google Scholar]
- Garrick, J.A.F. Studies on New Zealand Elasmobranchii. Part V. Scymnodalatias n. g. Based on Scymnodon sherwoodi Archey, 1921 (Selachii). Trans. R. Soc. N. Z. 1956, 83, 555–571. [Google Scholar]
- Garrick, J.A.F. Two northern hemisphere species of Centroscymnus in New Zealand Waters. Trans. R. Soc. N. Z. 1959, 87, 75–89. [Google Scholar]
- Barbosa du Bocage, J.V.; de Brito Capello, F. Sur quelque espèces inédites de Squalidae de la tribu Acanthiana, Gray, qui fréquentent les côtes du Portugal. Proc. Zool. Soc. Lond. B 1864, 1864, 260–263. [Google Scholar]
- Bigelow, H.B.; Schroeder, W.C.; Springer, S. New and little known sharks from the Atlantic and Gulf of Mexico. Bull. Mus. Comp. Zool. 1953, 109, 211–276. [Google Scholar]
- Atkinson, C.J.L.; Collin, S.P. Structure and topographic distribution of oral denticles in elasmobranch fishes. Biol. Bull. 2012, 222, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, C.J.L.; Martin, K.J.; Fraser, G.J.; Collin, S.P. Morphology and distribution of taste papillae and oral denticles in the developing oropharyngeal cavity of the bamboo shark, Chiloscyllium punctatum. Biol. Open 2017, 5, 1759–1769. [Google Scholar] [CrossRef] [PubMed]
- Sabaj, M.H. Standard Symbolic Codes for Institutional Resource Collections in Herpetology and Ichthyology: An Online Reference. Version 7.1 (21 March 2019). Am. Soc. Ichthyol. Herpetol. 2019. Available online: http://www.asih.org/ (accessed on 8 October 2023).
- Rincon, G.; Mota, R.; Mazzoleni, R.; Lessa, R.; de Moura, M.F.; Charvet, P. Dermal denticle variations on a newborn Roughskin Dogfish Centroscymnus owstonii (Chondrichthyes: Somniosidae) captured off northeastern Brazil with notes on ontogenetic differentiation. Reg. Stud. Mar. Sci. 2021, 44, 101761. [Google Scholar] [CrossRef]
- Reif, W. Morphogenesis and function of the squamation in sharks. Neues Jahrb. Für Geol. Und Paläontologie Abh. 1982, 164, 172–183. [Google Scholar] [CrossRef]
- Motta, P.; Habegger, M.L.; Lang, A.; Hueter, R.; Davis, J. Scale morphology and flexibility in the shortfin mako Isurus oxyrinchus and the blacktip shark Carcharhinus limbatus. J. Morphol. 2012, 273, 1096–1110. [Google Scholar] [CrossRef] [PubMed]
- Gabler-Smith, M.K.; Wainwright, D.K.; Wong, G.A.; Lauder, G.V. Dermal denticle diversity in sharks: Novel patterns on the interbranchial skin. Integr. Org. Biol. 2021, 3, obab034. [Google Scholar] [CrossRef] [PubMed]
- Popp, M.; White, C.F.; Bernal, L.P.; Wainwright, S.A.; Lauder George, V. The denticle surface of thresher shark tails: Three-dimensional structure and comparison to other pelagic species. J. Morphol. 2020, 281, 938–955. [Google Scholar] [CrossRef] [PubMed]
- Macías-Cuyare, M.; Oddone, M.C. Morphological pattern of the dermal denticles of the Southern sawtail catshark Galeus mincaronei Soto, 2001. J. Morphol. 2022, 283, 1120–1134. [Google Scholar] [CrossRef] [PubMed]
- Vaz, D.F.; de Carvalho, M.R. New Species of Squatina (Squatiniformes: Squatinidae) from Brazil, with comments on the taxonomy of angel sharks from the Central and Northwestern Atlantic. Copeia 2018, 106, 144–160. [Google Scholar] [CrossRef]
- Wen, L.; Weaver, J.C.; Lauder, G.V. Biomimetic shark skin: Design, fabrication, and hydrodynamic function. J. Exp. Biol. 2014, 217, 1656–1666. [Google Scholar] [CrossRef]
- Wen, L.; Weaver, J.C.; Thornycroft, P.J.M.; Lauder, G.V. Hydrodynamic function of biomimetic shark skin: Effect of denticle pattern and spacing. Bioinsp. Biomimet. 2015, 10, 1–13. [Google Scholar] [CrossRef]
- Domel, A.G.; Saadat, M.; Weaver, J.; Haj-Hariri, H.; Bertoldi, K.; Lauder, G.V. Shark denticle-inspired designs for improved aerodynamics. J. Roy. Soc. Inter. 2018, 15, 20170828. [Google Scholar] [CrossRef]
- Domel, A.G.; Domel, G.; Weaver, J.; Saadat, M.; Bertoldi, K.; Lauder, G.V. Hydrodynamic properties of biomimetic shark skin: Effect of denticle size and swimming speed. Bioinsp. Biomimet. 2018, 13, 056014. [Google Scholar] [CrossRef]
- Dillon, E.M.; Norris, R.D.; O’Dea, A. Dermal denticles as a tool to reconstruct shark communities. Mar. Ecol. Prog. Ser. 2017, 566, 117–134. [Google Scholar] [CrossRef]
- Dillon, E.M.; McCauley, D.J.; Morales-Saldaña, J.M.; Leonard, N.D.; Zhao, J.-X.; O’Dea, A. Fossil dermal denticles reveal the preexploitation baseline of a Caribbean coral reef shark community. Proc. Nat. Acad. Sci. USA 2021, 118, e2017735118. [Google Scholar] [CrossRef]
- Feichtinger, I.; Adnet, S.; Cuny, G.; Guinot, G.; Kriwet, J.; Neubauer, T.; Pollerspöck, J.; Shimada, K.; Straube, N.; Underwood, C. Comment on “An early Miocene extinction in pelagic sharks”. Science 2021, 374, eabk0632. [Google Scholar] [CrossRef]
- Naylor, G.J.P.; de Lima, A.; Castro, J.I.; Hubbell, G.; de Pinna, M.C.C. Comment on “An early Miocene extinction in pelagic sharks”. Science 2021, 374, eabj8723. [Google Scholar] [CrossRef]
- Tomita, T.; Murakumo, K.; Komoto, S.; Dove, A.; Kino, M.; Miyamoto, K.; Toda, M. Armored eyes of the whale shark. PLoS ONE 2020, 15, e0235342. [Google Scholar] [CrossRef]
- Nelson, G.J. Pharyngeal denticles (placoid scales) of sharks, with notes on the dermal skeleton of vertebrates. Amer. Mus. Nov. 1970, 2415, 1–26. [Google Scholar]
- Martin, K.J.; Rasch, L.J.; Cooper, R.L.; Metscher, B.D.; Johanson, Z.; Fraser, G.J. Sox2+ progenitors in sharks link taste development with the evolution of regenerative teeth from denticles. Proc. Nat. Acad. Sci. USA 2016, 113, 14769–14774. [Google Scholar] [CrossRef] [PubMed]
- Cooper, R.L.; Nicklin, E.F.; Rasch, L.J.; Fraser, G.J. Teeth outside the mouth: The evolution and development of shark denticles. Evol. Dev. 2022, 25, 54–72. [Google Scholar] [CrossRef] [PubMed]
- Tortonese, E. Studi sui Plagiostimi. Arch. Zool. Ital. 1952, 37, 383–398. [Google Scholar]
- Nakabo, T. Fishes of Japan with Pictorial Keys to the Species; Tokai University Press: Tokyo, Japan, 2002. [Google Scholar]
- Garman, S. New Plagiostomia. Bull. Mus. Comp. Zool. 1906, 11, 202–208. [Google Scholar]
- Compagno, L.J.V. Sharks of the world. An annotated and illustrated catalogue of sharks species known to date. FAO Fish. Synop. 1984, 125, 1–655. [Google Scholar]
- Compagno, L.J.V. Sharks. In The Living Marine Resources of the Western Central Atlantic. Vol. 1: Introduction, Molluscs, Crustaceans, Hagfishes, Sharks, Batoid Fishes and Chimaeras; Carpenter, K.E., Ed.; FAO: Rome, Italy, 2002; pp. 357–505. [Google Scholar]
- Last, P.R.; Stevens, J.D. Sharks and Rays of Australia, 2nd ed.; Fisheries Research & Development Corporation: Canberra, Australia, 2009. [Google Scholar]
- Ebert, D.A.; Fowler, S.L.; Compagno, L.J. Sharks of the World: A Fully Illustrated Guide; Wild Nature Press: Plymouth, UK, 2013. [Google Scholar]
- Weigmann, S. Annotated checklist of the living sharks, batoids and chimaeras (Chondrichthyes) of the world, with a focus on biogeographical diversity. J. Fish Biol. 2016, 88, 837–1037. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.P.C.B.; Vaz, D.F.B. Morphology and phylogenetic significance of the pelvic articular region in elasmobranchs (Chondrichthyes). Cladistics 2023, 39, 155–197. [Google Scholar] [CrossRef]
- Vaz, D.F.B.; Hilton, E.J. The caudal skeleton of Batrachoidiformes (Teleostei: Percomorphacea): A study of morphological diversity, intraspecific variation, and phylogenetic inferences. Zool. J. Linn. Soc. 2020, 189, 228–286. [Google Scholar] [CrossRef]
- Petean, F.d.; de Carvalho, M.R. Comparative morphology and systematics of the cookiecutter sharks, genus Isistius Gill (1864) (Chondrichthyes: Squaliformes: Dalatiidae). PLoS ONE 2018, 13, e0201913. [Google Scholar]
- Bechert, D.W.; Bartenwerfer, M.; Hoppe, G.; Reif, W.E. Drag reduction mechanisms derived from shark skin. AIAA 1986, 2, 1044–1068. [Google Scholar]
- Bixler, G.D.; Bhushan, B. Fluid Drag Reduction with Shark-Skin Riblet Inspired Microstructured Surfaces. Adv. Funct. Mater. 2013, 23, 4507–4528. [Google Scholar] [CrossRef]
- Dai, W.; Alkahtani, M.; Hemmer, P.R.; Liang, H. Drag-reduction of 3D printed shark-skin-like surfaces. Friction 2019, 7, 603–612. [Google Scholar] [CrossRef]
- Luo, Y.; Xu, X.; Li, D.; Song, W. Recent developments in fabricating drag reduction surfaces covering biological sharkskin morphology. Rev. Chem. Eng. 2015, 32, 93–113. [Google Scholar] [CrossRef]
- Gabler-Smith, M.K.; Lauder, G.V. Ridges and riblets: Shark skin surfaces versus biomimetic models. Front. Mar. Sci. 2022, 9, 975062. [Google Scholar] [CrossRef]
- Raayai-Ardakani, S.; McKinley, G.H. Drag reduction using wrinkled surfaces in high Reynolds number laminar boundary layer flows. Phys. Fluids 2017, 29, 093605. [Google Scholar] [CrossRef]
- Raayai-Ardakani, S.; McKinley, G.H. Geometric optimization of riblet-textured surfaces for drag reduction in laminar boundary layer flows. Phys. Fluids 2019, 31, 053601. [Google Scholar] [CrossRef]
- Irschick, D.J.; Hammerschlag, N. Morphological scaling of body form in four shark species differing in ecology and life history. Biol. J. Linn. Soc. 2014, 114, 126–135. [Google Scholar] [CrossRef]
- Irschick, D.J.; Fu, A.; Lauder, G.; Wilga, C.; Kuo, C.-Y.; Hammerschlag, N. A comparative morphological analysis of body and fin shape for eight shark species. Biol. J. Linn. Soc. 2017, 122, 589–604. [Google Scholar] [CrossRef]
- Gayford, J.H.; Godfrey, H.; Whitehead, D.A. Ontogenetic morphometry of the brown smoothhound shark Mustelus henlei with implications for ecology and evolution. J. Morphol. 2023, 284, e21608. [Google Scholar] [CrossRef] [PubMed]


























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species and Catalog Number | Total Length (mm) | Method |
|---|---|---|
| Centroscymnus coelolepis | ||
| MCZ 132510 | 350 | µCT |
| MCZ 38294 | 550 | µCT |
| MCZ 38297 | 650 | µCT |
| MCZ 57703 | 744 | µCT |
| MCZ 39621 | 895 | µCT |
| MZUSP 118097 | 328 | SEM |
| MNHN 2001 3226 | 681 | SEM |
| USNM 408586 | 837 | SEM |
| NSMT-P 32586 | 949 | SEM |
| Centroscymnus owstonii | ||
| MCZ 35038 | 280 | SEM |
| MNHN 1989 0654 | 525 | SEM |
| HUMZ 101723 | 627 | SEM |
| TMFE 577 | 659 | SEM |
| UERJ.AC 1460 | 950 | SEM |
| BMNH 1973.7.9.9 | 1071 | SEM |
| Centroselachus crepidater | ||
| AMNH 58099 | 283 | SEM |
| MNHN 1998 1299A | 768 | SEM |
| USNM 408581 | 525 | SEM |
| USNM 408582 | 426 | SEM |
| Scymnodon macracanthus | ||
| MNHN 2008 1916 | 582 | SEM |
| BMNH 1884.2.6.7 (holotype) | 667 | SEM |
| MNHN 2003 1708 | 691 | SEM |
| NSMT-P 32079 | 1179 | SEM |
| Scymnodon ichiharai | ||
| TMFE 1875 | 942 | SEM |
| Scymnodon ringens | ||
| BMNH 1991.7.9.690 | 317 | SEM |
| BMNH 1991.7.9.688 | 518 | SEM |
| BMNH 1987.1.21.86 | 674 | SEM |
| Scymnodalatias cf. garrick | ||
| ZMH 122774 | 417 | Steromicroscope |
| Scymnodalatias albicauda | ||
| HUMZ 129360 | 170 | SEM |
| Somniosus cf. microcephalus | ||
| UF 162500 | 1505 | SEM |
| Somniosus cf. rostratus | ||
| MZUSP uncatalogued | 1090 | SEM |
| Zameus squamulosus | ||
| USNM 220496 | 341 | SEM |
| USNM 220489 | 532 | SEM |
| Snout Morphotype | Spiracular Morphotype | Trunk Morphotype | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Character | 1 | 2 | 3 | 1 | 2 | 3 | 4 | 1 | 2 | 3 | 4 |
| Number of longitudinal lateral ridges | 2–4 | 4 | 6+ | 2 | 4 | 4 | 6 | 2 | 4 | 0 | 6+ |
| Development of external lateral ridges | when present, reduced | complete | complete | absent | reduced | reduced or complete | complete | absent | reduced | absent | complete |
| Posterior cusps | absent | absent | absent | present | present | absent | absent | present | present | absent | absent |
| Central cusp aspect ratio | n/a | n/a | n/a | 0.5–1.06 | 0.41–0.95 | n/a | n/a | 1.08–1.8 | 0.52–1.49 | n/a | n/a |
| Lateral depressions | present | present | present | absent | present | present | present | absent | present | absent | present |
| Relative position of anterolateral edge of the crown | n/a | n/a | n/a | aligned with lpc | external to lpc | n/a | n/a | aligned with lpc | external to lpc | n/a | n/a |
| Snout Morphotype | Spiracular Morphotype | Trunk Morphotype | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 1 | 2 | 3 | 4 | 1 | 2 | 3 | 4 | |
| Character | |||||||||||
| Surface Area (mm2) | 1.48–2.61 | 1.87–4.37 | 2.49–8.79 | 0.97–2.46 | 1.69–4.93 | 2.22–8.43 | 5.94–7.83 | 1.48–2.28 | 1.98–12.54 | 6.9–18.37 | 3.21–7.81 |
| Volume (mm3) | 0.03–0.1 | 0.07–0.21 | 0.12–0.60 | 0.02–0.11 | 0.06–0.26 | 0.08–0.63 | 0.39–0.55 | 0.04–0.09 | 0.08–1.03 | 0.43–1.98 | 0.13–0.47 |
| Crown Length (mm) | 0.49–0.9 | 0.75–1.32 | 0.7–1.62 | 0.53–0.93 | 0.7–1.35 | 0.84–1.94 | 1.5–1.72 | 0.71–1.0 | 0.91–2.43 | 1.88–2.89 | 1.29–1.78 |
| Crown Width (mm) | 0.58–0.90 | 0.8–1.27 | 1.13–2.11 | 0.47–0.63 | 0.64–1.22 | 0.9–1.77 | 1.65–1.93 | 0.49–0.71 | 0.62–2.23 | 1.61–2.94 | 1.09–1.96 |
| Crown Aspect Ratio (Length/Width) | 0.76–1.16 | 0.77–1.04 | 0.62–0.98 | 0.94–1.52 | 0.96–1.38 | 0.79–1.12 | 0.89–0.91 | 1.3–1.52 | 1.07–1.67 | 0.95–1.19 | 0.9–1.18 |
| Central Cusp Length (mm) | - | - | - | 0.11–0.4 | 0.18–0.44 | - | - | 0.29–0.43 | 0.27–0.53 | - | - |
| Central Cusp Width (mm) | - | - | - | 0.2–0.42 | 0.32–0.64 | - | - | 0.22–0.34 | 0.29–0.70 | - | - |
| Central Cusp Aspect Ratio | - | - | - | 0.5–1.06 | 0.41–0.95 | - | - | 1.08–1.8 | 0.52–1.49 | - | - |
| Lateral Cusp Length (mm) | - | - | - | 0.03–0.13 | 0.03–0.16 | - | - | 0.1–0.25 | 0.04–0.24 | - | - |
| Lateral Cusp Width (mm) | - | - | - | 0.07–0.18 | 0.09–0.24 | - | - | 0.11–0.23 | 0.08–0.29 | - | - |
| Lateral Cusp Aspect Ratio | - | - | - | 0.29–0.89 | 0.21–0.81 | - | - | 0.64–1.49 | 0.24–1.24 | - | - |
| Central Cusp/Lateral Cusp Length | - | - | - | 2.42–8.12 | 2.72–7.76 | - | - | 1.77–4.14 | 1.54–8.68 | - | - |
| Central Cusp/Lateral cusp Width | - | - | - | 1.89–5.6 | 1.79–5.56 | - | - | 1.26–2.5 | 1.37–5.94 | - | - |
| Basal Root Length (mm) | 0.64–1.08 | 0.49–1.01 | 0.53–1.01 | 0.4–1.06 | 0.63–1.14 | 0.66–1.28 | 0.87–1.02 | 0.68–0.89 | 0.59–1.28 | 0.88–1.28 | 0.52–0.81 |
| Basal Root Width (mm) | 0.77–1.12 | 0.81–1.42 | 1.13–1.8 | 0.68–1.21 | 0.84–1.38 | 0.90–1.75 | 1.33–1.52 | 0.85–1.06 | 0.79–1.91 | 1.06–2.36 | 1.01–1.76 |
| Basal Root Aspect Ratio | 0.69–0.98 | 0.55–0.92 | 0.46–0.63 | 0.59–1.03 | 0.67–1.0 | 0.63–0.88 | 0.65–067 | 0.68–0.94 | 0.57–1.6 | 0.51–1.06 | 0.4–0.61 |
| Number Longitudinal Ridges | 2–5 | 4–5 | 6–10 | 2–3 | 4–5 | 4–5 | 6 | 2–3 | 4–5 | 0 | 6–9 |
| Crown Angle (degrees) | 19.63–58.42 | 2.59–35.56 | 1.29–15.69 | 36.51–59.7 | 27.62–50.40 | 11.33–35.84 | 7.31–8.48 | 28.65–52.41 | 11.05–48.96 | 2.41–19.06 | 13.79–26.85 |
| Denticle Variable | Expected Isometric Slope | Slope | Standard Error of Slope | Adjusted R2 | Regression p-Value |
|---|---|---|---|---|---|
| Surface Area | 2 | 1.38 | 0.11 | 0.42 | <0.001 |
| Volume | 3 | 2.23 | 0.17 | 0.47 | <0.001 |
| Crown Length | 1 | 0.81 | 0.08 | 0.33 | <0.001 |
| Crown Width | 1 | 1.05 | 0.07 | 0.51 | <0.001 |
| Basal Root Length | 1 | 0.12 | 0.05 | 0.03 | 0.01 |
| Basal Root Width | 1 | 0.47 | 0.05 | 0.32 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vaz, D.F.B.; Avery, T.M.; Gabler-Smith, M.K.; Lauder, G.V. The Denticle Multiverse: Morphological Diversity of Placoid Scales across Ontogeny in the Portuguese Dogfish, Centroscymnus coelolepis, and Its Systematic Implications. Diversity 2023, 15, 1105. https://doi.org/10.3390/d15111105
Vaz DFB, Avery TM, Gabler-Smith MK, Lauder GV. The Denticle Multiverse: Morphological Diversity of Placoid Scales across Ontogeny in the Portuguese Dogfish, Centroscymnus coelolepis, and Its Systematic Implications. Diversity. 2023; 15(11):1105. https://doi.org/10.3390/d15111105
Chicago/Turabian StyleVaz, Diego F. B., Tess M. Avery, Molly K. Gabler-Smith, and George V. Lauder. 2023. "The Denticle Multiverse: Morphological Diversity of Placoid Scales across Ontogeny in the Portuguese Dogfish, Centroscymnus coelolepis, and Its Systematic Implications" Diversity 15, no. 11: 1105. https://doi.org/10.3390/d15111105