
Coccolithophore Distribution in the Western Black Sea in the Summer of 2016


 , and
, and
Abstract
:1. Introduction
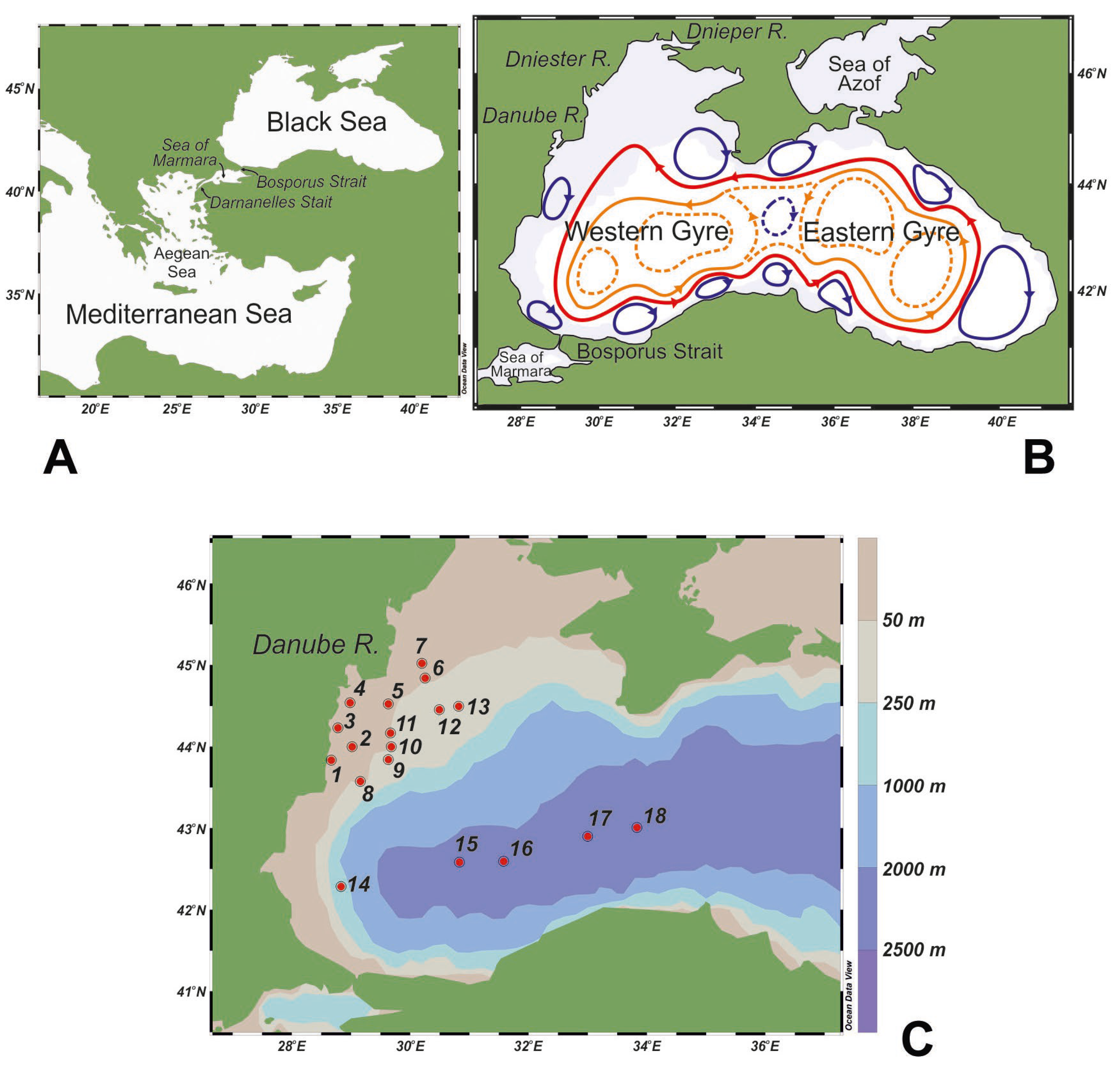
2. Study Area
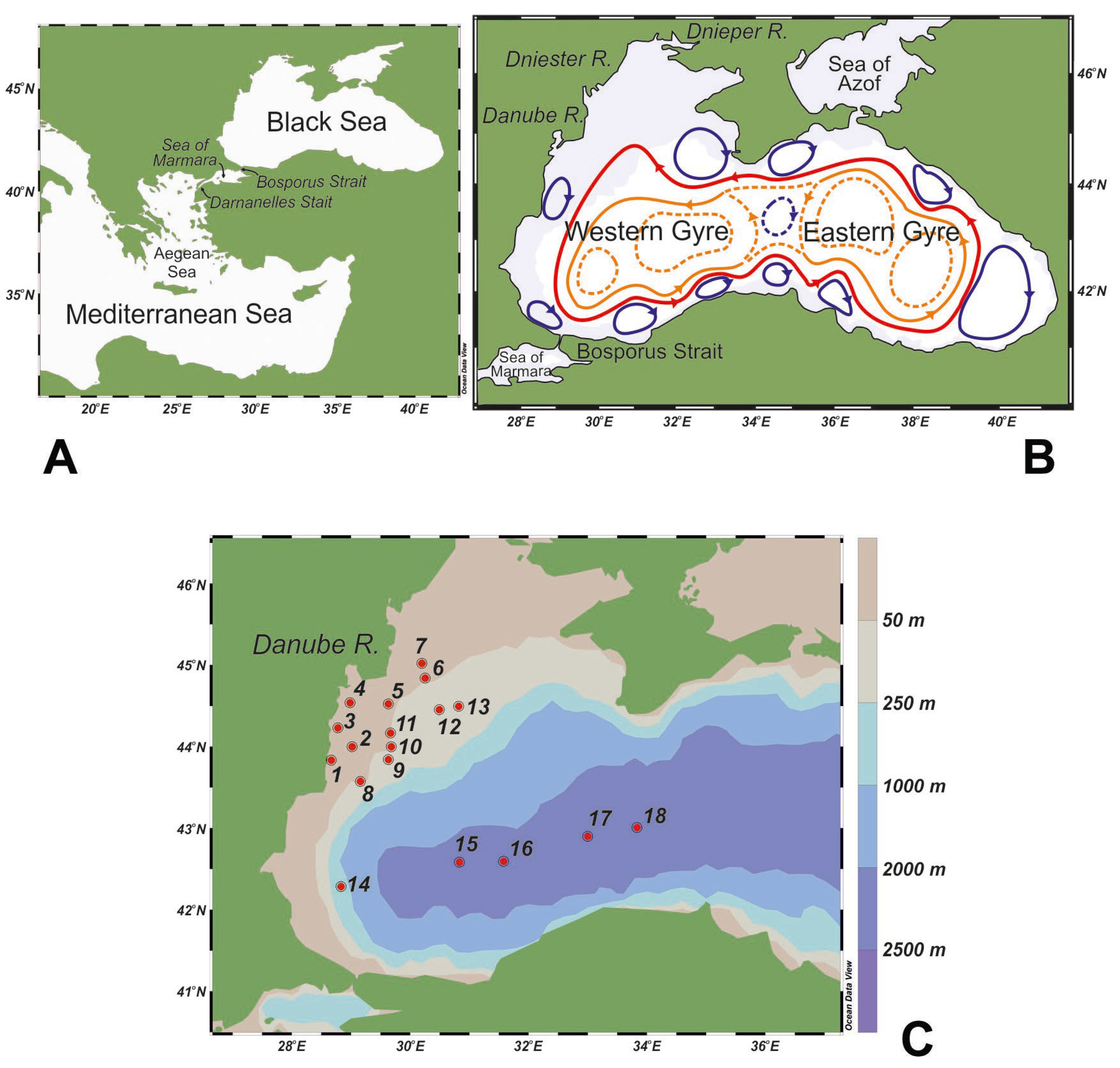
3. Material and Methods
3.1. Sample Collection and Analysis
3.2. Statistical Analysis
4. Results
4.1. Environmental Conditions (CTD Measurements)
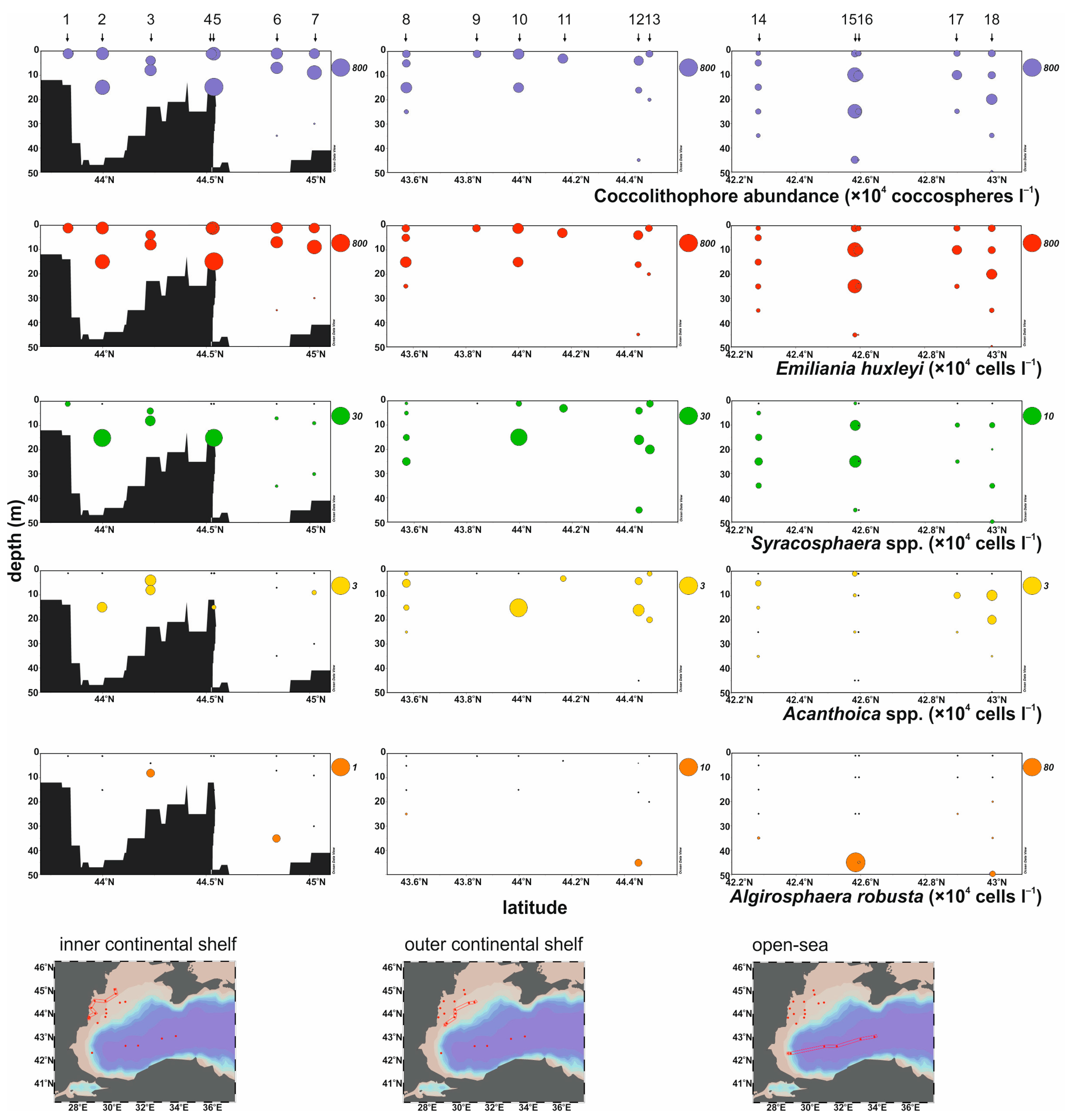
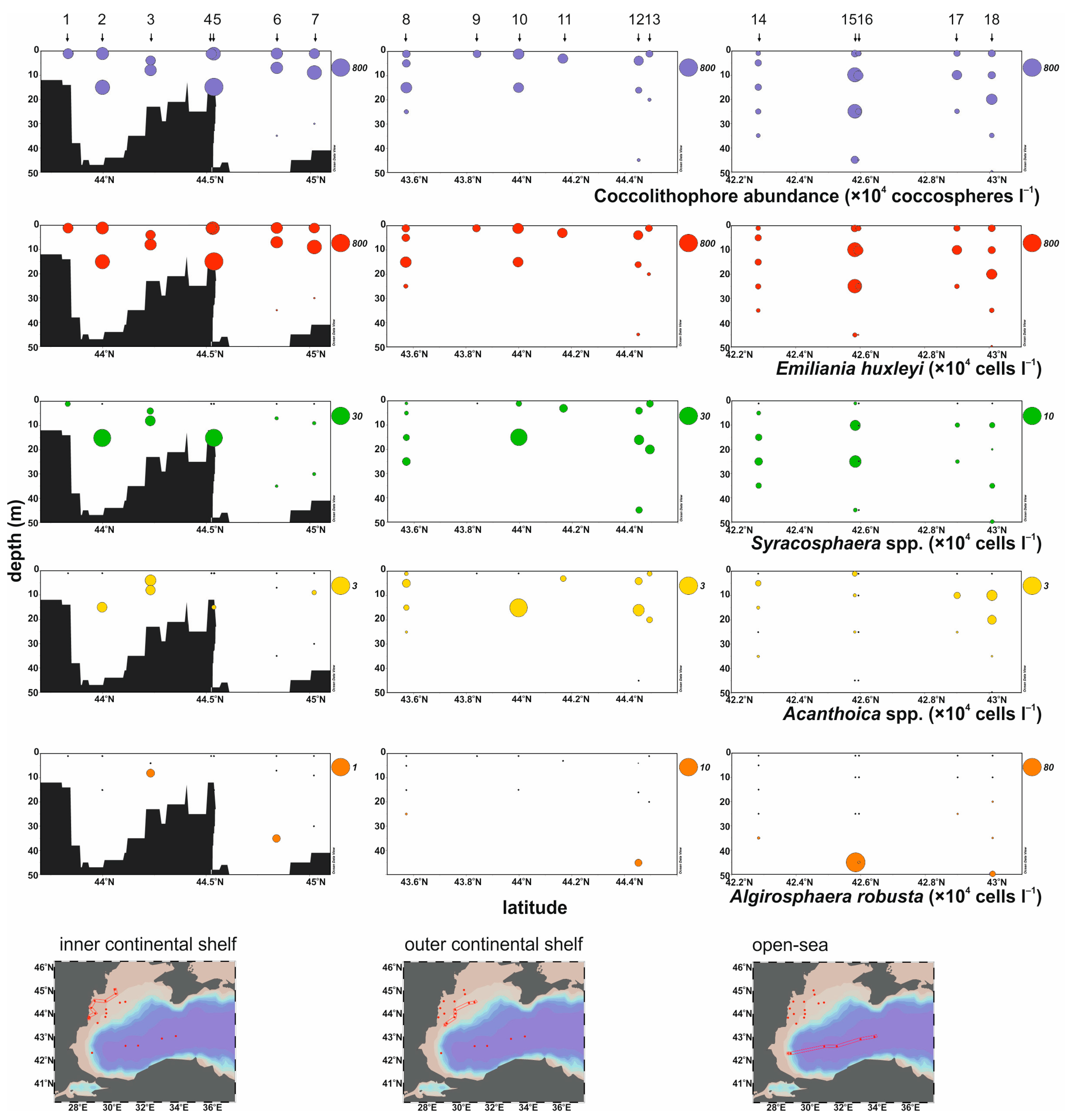
4.2. Coccolithophore Abundance and Distribution
4.3. Coccolithophore Taxonomy
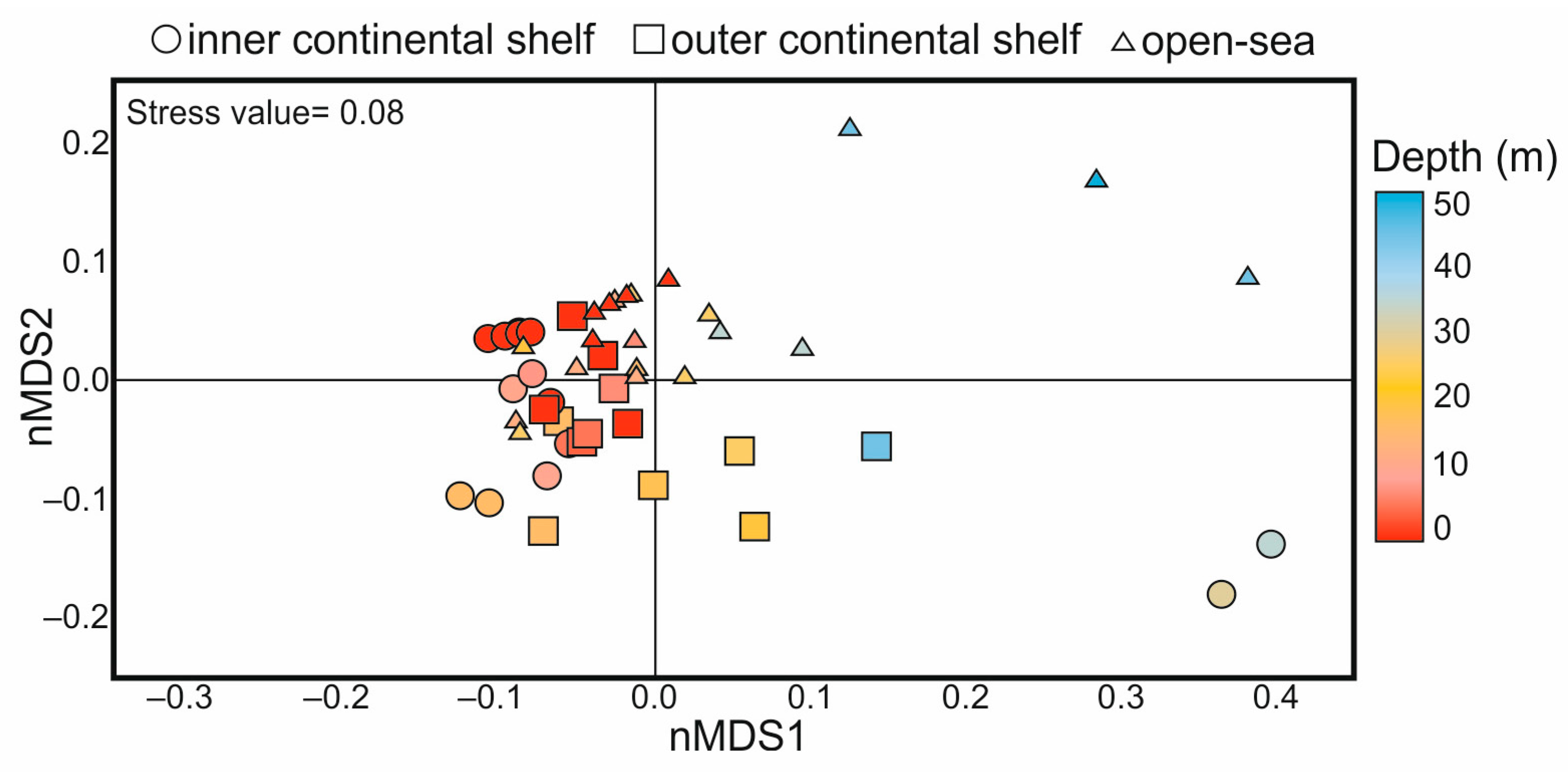
4.4. Statistical Analyses
5. Discussion
6. Conclusions
- Living coccolithophores showed high abundances, reaching a maximum of 763 × 104 coccospheres L−1 with the lightly calcified morphotype of E. huxleyi type A being the dominant species accounting for an average of 92%, confirming its typical dominance in summer coccolithophore assemblages of the Black Sea;
- Apart from E. huxleyi, and the other heterococcolithophores S. dilatata, S. molischii, A. acanthifera, and A. quattrospina, Algirosphaera robusta, and two holococcolithophore forms S. arethusae HOL and H. cornifera, contributed to the coccolithophore assemblages, indicating their ability to thrive in the low salinity conditions of the Black Sea;
- The vertical distribution of coccolithophore species appears to depend on the temperature and irradiance levels. Emiliania huxleyi, Syracosphaera spp., and Acanthoica spp. were mainly distributed in the upper water layers, within the thermocline. High abundances of A. robusta occurred below the thermocline, indicating its preference for low light and temperature conditions;
- The highest abundances of E. huxleyi (max. 737 × 104 cells L−1) and Syracosphaera (max. 26 × 104 cells L−1) were found over the northwestern inner shelf region in Danube-influenced waters, associated with increased freshwater and nutrient inputs;
- Our results indicate that the seasonal thermocline is the main factor regulating the distribution of living coccolithophores, whereas significant spatial variations in the assemblages are related to the influence of freshwater input.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Young, J.R.; Geisen, M.; Cros, L.; Kleijne, A.; Sprengel, C.; Probert, I.; Østergaard, J. A guide to extant coccolithophore taxonomy. J. Nannoplankton Res. 2003, 1, 1–125. [Google Scholar] [CrossRef]
- Jordan, R.W.; Chamberlain, A.H.L. Biodiversity among haptophyte algae. Biodivers. Conserv. 1997, 6, 131–152. [Google Scholar] [CrossRef]
- Brown, C.W.; Yoder, J.A. Coccolithophorid blooms in the global ocean. J. Geophys. Res. Ocean 1994, 99, 7467–7482. [Google Scholar] [CrossRef]
- Iglesias-Rodriguez, M.D.; Garcia Saez, A.; Groben, R.; Edwards, K.J.; Batley, J.; Medlin, L.K.; Hayes, P.K. Polymorphic microsatellite loci in global populations of the marine coccolithophorid Emiliania huxleyi. Mol. Ecol. Notes 2002, 2, 495–497. [Google Scholar] [CrossRef]
- Tyrrell, T.; Merico, A. Emiliania huxleyi: Bloom observations and the conditions that induce them. In Coccolithophores; Springer: Berlin/Heidelberg, Germany, 2004; pp. 75–97. [Google Scholar] [CrossRef]
- Holligan, P.M.; Fernandéz, E.; Aiken, J.; Balch, W.M.; Boyd, P.; Burkill, P.H.; Finc, M.; Groom, S.B.; Malin, G.; Müller, K.; et al. A biogeohemical study of the coccolithophore, Emiliania huxleyi, in the North Atlantic. Glob. Biogeochem. Cycles 1993, 7, 879–900. [Google Scholar] [CrossRef]
- Poulton, A.J.; Adey, T.R.; Balch, W.M.; Holligan, P.M. Relating coccolithophore calcification rates to phytoplankton community dynamics: Regional differences and implications for carbon export. Deep-Sea Res. II 2007, 54, 538–557. [Google Scholar] [CrossRef]
- Zondervan, I.; Zeebe, R.E.; Rost, B.; Riebesell, U. Decreasing marine biogenic calcification: A negative feedback on rising atmospheric pCO2. Glob. Biogeochem. Cycles 2001, 15, 507–516. [Google Scholar] [CrossRef]
- Rost, B.; Riebesell, U. Coccolithophores and the biological pump: Responses to environmental changes. In Coccolithophores: From Molecular Processes to Global Impact; Thierstein, H.R., Young, J.R., Eds.; Springer: Berlin/Heidelberg, Germany, 2004; pp. 99–125. [Google Scholar] [CrossRef]
- Taylor, A.R.; Brownlee, C.; Wheeler, G. Coccolithophore cell biology: Chalking up progress. Ann. Rev. Mar. Sci. 2017, 9, 283–310. [Google Scholar] [CrossRef]
- Riebesell, U.; Zondervan, I.; Rost, B.; Tortell, P.D.; Zeebe, R.E.; Morel, F.M.M. Reduced calcification of marine plankton in response to increased atmospheric CO2. Nature 2000, 407, 364–367. [Google Scholar] [CrossRef]
- Doney, S.C.; Fabry, V.J.; Feely, R.A.; Kleypas, J. Ocean acidification: The other CO2 problem. Ann. Rev. Mar. Sci. 2009, 1, 169–192. [Google Scholar] [CrossRef]
- Bach, L.T.; Riebesell, U.; Gutowska, M.A.; Federwisch, L.; Schulz, K.G. A unifying concept of coccolithophore sensitivity to changing carbonate chemistry embedded in an ecological framework. Prog. Oceanogr. 2015, 135, 125–138. [Google Scholar] [CrossRef]
- Oguz, T. Long-term impacts of anthropogenic forcing on the Black Sea ecosystem. Oceanography 2005, 18, 104–113. [Google Scholar] [CrossRef]
- Oguz, T.; Dippner, J.W.; Kaymaz, Z. Climatic regulation of the Black Sea hydro-meteorological and ecological properties at interannual-to-decadal time scales. J. Mar. Syst. 2006, 60, 235–254. [Google Scholar] [CrossRef]
- Mikaelyan, A.S.; Silkin, V.A.; Pautova, L.A. Coccolithophorids in the Black Sea: Their interannual and long-term changes. Oceanology 2011, 51, 39–48. [Google Scholar] [CrossRef]
- Mikaelyan, A.S.; Pautova, L.A.; Chasovnikov, V.K.; Mosharov, S.A.; Silkin, V.A. Alternation of diatoms and coccolithophores in the north-eastern Black Sea: A response to nutrient changes. Hydrobiologia 2015, 755, 89–105. [Google Scholar] [CrossRef]
- Eker-Develi, E.; Kideys, A.E. Distribution of phytoplankton in the southern Black Sea in summer 1996, spring and autumn 1998. J. Mar. Syst. 2003, 39, 203–211. [Google Scholar] [CrossRef]
- Pautova, L.A.; Mikaelyan, A.S.; Silkin, V.A. Structure of plankton phytocenoses in the shelf waters of the Northeastern Black Sea during the Emiliania huxleyi bloom in 2002–2005. Oceanology 2007, 47, 477–480. [Google Scholar] [CrossRef]
- Eker-Develi, E.; Konucu, M.; Psarra, S.; Slabakova, N.; Triantaphyllou, M.; Dimiza, M.; Karageorgis, A.P.; Uysal, Z.; Berthon, J.-F. Phytoplankton and pigment composition during an Emiliania huxleyi bloom in the Black Sea. Reg. Stud. Mar. Sci. 2023, 65, 103070. [Google Scholar] [CrossRef]
- Cokacar, T.; Oguz, T.; Kubilay, N. Satellite-detected early summer coccolithophore blooms and their interannual variability in the Black Sea. Deep Sea Res. Part 1 Oceanogr. Res. Pap. 2004, 51, 1017–1031. [Google Scholar] [CrossRef]
- Mikaelyan, A.S.; Pautova, L.A.; Pogosyan, S.I.; Sukhanova, I.N. Summer bloom of coccolithophorids in the northeastern Black Sea. Oceanology 2005, 45 (Suppl. 1), S127–S138. [Google Scholar]
- Oguz, T.; Merico, A. Factors controlling the summer Emiliania huxleyi bloom in the Black Sea: A modeling study. J. Mar. Syst. 2006, 59, 173–188. [Google Scholar] [CrossRef]
- Yasakova, O.N.; Okolodkov, Y.B.; Chasovnikov, V.K. Increasing contribution of coccolithophorids to the phytoplankton in the northeastern Black Sea. Mar. Pollut. Bull. 2017, 124, 526–534. [Google Scholar] [CrossRef] [PubMed]
- Kubryakov, A.A.; Mikaelyan, A.S.; Stanichny, S.V. Summer and winter coccolithophore blooms in the Black Sea and their impact on production of dissolved organic matter from Bio-Argo data. J. Mar. Syst. 2019, 199, 103220. [Google Scholar] [CrossRef]
- Kubryakov, A.A.; Mikaelyan, A.S.; Stanichny, S.V. Extremely strong coccolithophore blooms in the Black Sea: The decisive role of winter vertical entrainment of deep water. Deep Sea Res. Part 1 Oceanogr. Res. Pap. 2021, 173, 103554. [Google Scholar] [CrossRef]
- Silkin, V.; Mikaelyan, A.S.; Pautova, L.; Fedorov, A. Annual Dynamics of Phytoplankton in the Black Sea in Relation to Wind Exposure. J. Mar. Sci. Eng. 2021, 9, 1435. [Google Scholar] [CrossRef]
- Vostokov, S.V.; Vostokova, A.S.; Vazyulya, S.V. Seasonal and Long-Term Variability of Coccolithophores in the Black Sea According to Remote Sensing Data and the Results of Field Investigations. J. Mar. Sci. Eng. 2022, 10, 97. [Google Scholar] [CrossRef]
- Mikaelyan, A.S.; Kubryakov, A.A.; Silkin, V.A.; Pautova, L.A.; Chasovnikov, V.K. Regional climate and patterns of phytoplankton annual succession in the open waters of the Black Sea. Deep Sea Res. Part 1 Oceanogr. Res. Pap. 2018, 142, 44–57. [Google Scholar] [CrossRef]
- Mikaelyan, A.S.; Pautova, L.A.; Fedorov, A.V. Seasonal evolution of deep phytoplankton assemblages in the Black Sea. J. Sea Res. 2021, 178, 102125. [Google Scholar] [CrossRef]
- Osawa, M.; Takahashi, K.; Hay, B.J. Shell-bearing plankton fluxes in the central Black sea, 1989–1991. Deep Sea Res. Part 1 Oceanogr. Res. Pap. 2005, 52, 1677–1698. [Google Scholar] [CrossRef]
- Skampa, E.; Triantaphyllou, M.V.; Dimiza, M.D.; Gogou, A.; Malinverno, E.; Stavrakakis, S.; Parinos, C.; Panagiotopoulos, I.P.; Tselenti, D.; Archontikis, O.; et al. Coccolithophore export in three deep sea sites of the Aegean and Ionian Seas (Eastern Mediterranean): Biogeographical patterns and biogenic carbonate fluxes. Deep-Sea Res. II 2020, 171, 104690. [Google Scholar] [CrossRef]
- Özsoy, E.; Ünlüata, Ü. Oceanography of the Black Sea: A review of some recent results. Earth-Sci. Rev. 1997, 42, 231–272. [Google Scholar] [CrossRef]
- Stanev, E.V. On the mechanisms of the Black-Sea circulation. Earth-Sci. Rev. 1990, 28, 285–319. [Google Scholar] [CrossRef]
- Oguz, T.; Latun, V.S.; Latif, M.A.; Vladimirov, V.V.; Sur, H.I.; Markov, A.A.; Özsoy, E.; Kotovshchikov, B.B.; Eremeev, V.V.; Ünlüata, Ü. Circulation in the surface and intermediate layers of the Black-Sea. Deep Sea Res. Part 1 Oceanogr. Res. Pap. 1993, 40, 1597–1612. [Google Scholar] [CrossRef]
- Korotaev, G.; Oguz, T.; Nikiforov, A.; Koblinsky, C. Seasonal, interannual, and mesoscale variability of the Black Sea upper layer circulation derived from altimeter data. J. Geophys. Res. Ocean 2003, 108, 3122. [Google Scholar] [CrossRef]
- Barale, V. The European Marginal and Enclosed Seas: An Overview. In Remote Sensing of the European Seas; Barale, V., Gade, M., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 3–22. [Google Scholar] [CrossRef]
- Talley, L.D.; Pickard, G.L.; Emery, W.L.; Swift, J.H. Gravity waves, tides, and coastal oceanography. Chapter 8. In Descriptive Physical Oceanography, 6th ed.; Swift, J.H., Ed.; Academic Press: Cambridge, MA, USA, 2011; pp. 1–31. [Google Scholar] [CrossRef]
- Stewart, K.; Kassakian, S.; Krynytzky, M.; DiJulio, D.; Murray, J.W. Oxic, suboxic, and anoxic conditions in the Black Sea. In The Black Sea Flood Question: Changes in Coastline, Climate, and Human Settlement; Yanko-Hombach, V., Gilbert, A.S., Panin, N., Dolukhanov, P.M., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 1–21. [Google Scholar] [CrossRef]
- Murray, J.; Jannasch, H.; Honjo, S.; Anderson, R.F.; Reeburgh, W.S.; Top, Z.; Friederich, G.E.; Codispoti, L.A.; Izdar, E. Unexpected changes in the oxic/anoxic interface in the Black Sea. Nature 1989, 338, 411–413. [Google Scholar] [CrossRef]
- Fuchsman, C.A.; Staley, J.T.; Oakley, B.B.; Kirkpatrick, J.B.; Murray, J.W. Free-living and aggregate-associated Planctomycetes in the Black Sea. FEMS Microbiol. Ecol. 2012, 80, 402–416. [Google Scholar] [CrossRef] [PubMed]
- Miladinova, S.; Stips, A.; Garcia-Gorriz, E.; Macias Moy, D. Formation and changes of the Black Sea cold intermediate layer. Prog. Oceanogr. 2018, 167, 11–23. [Google Scholar] [CrossRef]
- Murray, J.W.; Top, Z.; Ozsoy, E. Hydrographic properties and ventilation of the Black Sea. Deep Sea Res. Part 1 Oceanogr. Res. Pap. 1991, 38, S663–S689. [Google Scholar] [CrossRef]
- Bollmann, J.; Cortés, M.Y.; Haidar, A.T.; Brabec, B.; Close, A.; Hofmann, R.; Palma, S.; Tupas, L.; Thierstein, H.R. Techniques for quantitative analyses of calcareous marine phytoplankton. Mar. Micropaleontol. 2002, 44, 163–185. [Google Scholar] [CrossRef]
- Cros, L.; Fortuño, J.M. Atlas of Northwestern Mediterranean Coccolithophores. Sci. Mar. 2002, 66, 1–182. [Google Scholar] [CrossRef]
- Young, J.R.; Bown, P.R.; Lees, J.A. Nannotax3 Website. International Nannoplankton Association. 2022. Available online: http://www.mikrotax.org/Nannotax3 (accessed on 21 April 2022).
- Schlitzer, R. Ocean Data View. 2023. Available online: https://odv.awi.de (accessed on 25 October 2023).
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Clarke, K.R. Non-parametric multivariate analysis of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Vedernikov, V.I.; Demidov, A.B. Vertical distribution of primary production and chlorophyll during different seasons in deep regions of the Black Sea. Oceanology 1997, 37, 376–384. [Google Scholar]
- Yunev, O.; Carstensen, J.; Stelmakh, L.; Belokopytov, V.; Suslin, V. Temporal changes of phytoplankton biomass in the western Black Sea shelf waters: Evaluation by satellite data (1998–2018). Estuar. Coast. Shelf Sci. 2022, 271, 107865. [Google Scholar] [CrossRef]
- Paasche, E. A review of the coccolithophorid Emiliania huxleyi (Prymnesiophyceae), with particular reference to growth, coccolith formation, and calcification-photosynthesis interactions. Phycologia 2002, 40, 503–529. [Google Scholar] [CrossRef]
- Cook, S.S.; Whittock, L.; Wright, S.W.; Hallegraeff, G.M. Photosynthetic pigment and genetic differences between two Southern Ocean morphotypes of Emiliania huxleyi (Haptophyta). J. Phycol. 2011, 47, 615–626. [Google Scholar] [CrossRef]
- Rosas-Navarro, A.; Langer, G.; Ziveri, P. Temperature affects the morphology and calcification of Emiliania huxleyi strains. Biogeosciences 2016, 13, 2913–2926. [Google Scholar] [CrossRef]
- Balch, W.M.; Kilpatric, K.A.; Trees, C.C. The 1991 coccolithophore bloom in the central North Atlantic. 1. Optical properties and factors affecting their distribution. Limnol. Oceanogr. 1996, 41, 1669–1683. [Google Scholar] [CrossRef]
- Paasche, E.; Brubak, S.; Skatiebpl, S.; Young, J.R.; Green, J.C. Growth and calcification in the coccolithophorid Emiliania huxleyi (Haptophyceae) at low salinities. Phycologia 1996, 35, 394–403. [Google Scholar] [CrossRef]
- Bollmann, J.; Herrle, H.O. Morphological variation of Emiliania huxleyi and sea surface salinity. Earth Planet. Sci. Lett. 2007, 255, 273–288. [Google Scholar] [CrossRef]
- Gebühr, C.; Sheward, R.M.; Herrle, J.O.; Bollmann, J. Strain-specific morphological response of the dominant calcifying phytoplankton species Emiliania huxleyi to salinity change. PLoS ONE 2021, 16, e0246745. [Google Scholar] [CrossRef] [PubMed]
- Paasche, E. Roles of nitrogen and phosphorus in coccolith formation in Emiliania huxleyi (Prymnesiophyceae). Eur. J. Phycol. 1998, 33, 33–42. [Google Scholar] [CrossRef]
- Triantaphyllou, M.; Dimiza, M.; Krasakopoulou, E.; Malinverno, E.; Lianou, V.; Souvermezoglou, E. Seasonal variation in Emiliania huxleyi coccolith morphology and calcification in the Aegean Sea (Eastern Mediterranean). Geobios 2010, 43, 99–110. [Google Scholar] [CrossRef]
- D’Amario, B.; Ziveri, P.; Grelaud, M.; Oviedo, A. Emiliania huxleyi coccolith calcite mass modulation by morphological changes and ecology in the Mediterranean Sea. PLoS ONE 2018, 13, e0201161. [Google Scholar] [CrossRef] [PubMed]
- Karatsolis, B.-T.; Triantaphyllou, M.V.; Dimiza, M.D.; Malinverno, E.; Lagaria, A.; Mara, P.; Archontikis, O.; Psarra, S. Coccolithophore assemblage response to Black Sea water inflow into the north Aegean Sea (NE Mediterranean). Cont. Shelf Res. 2017, 149, 138–150. [Google Scholar] [CrossRef]
- Dimiza, M.D.; Koukousioura, O.; Michailidis, I.; Dimou, V.-G.; Navrozidou, V.; Aligizaki, K.; Seferlis, M. Seasonal living coccolithophore distribution in the enclosed coastal environments of the Thessaloniki Bay (Thermaikos Gulf, NW Aegean Sea). Rev. Micropaleontol. 2020, 69, 100449. [Google Scholar] [CrossRef]
- Daniels, C.J.; Sheward, R.M.; Poulton, A.J. Biogeochemical implications of comparative growth rates of Emiliania huxleyi and Coccolithus species. Biogeosciences 2014, 11, 6915–6925. [Google Scholar] [CrossRef]
- Andruleit, H.; Rogalla, U. Coccolithophores in surface sediment of Arabian Sea in relation to environmental gradients in surface waters. Mar. Geol. 2002, 186, 505–526. [Google Scholar] [CrossRef]
- Triantaphyllou, M.V.; Ziveri, P.; Tselepides, A. Coccolithophore export production and response to seasonal surface water variability in the oligotrophic Cretan Sea (NE Mediterranean). Micropaleontology 2004, 50 (Suppl. 1), 127–144. [Google Scholar] [CrossRef]
- Dimiza, M.D.; Triantaphyllou, M.V.; Dermitzakis, M.D. Seasonality and ecology of living coccolithophores in Eastern Mediterranean coastal environments (Andros Island, Middle Aegean Sea). Micropaleontology 2008, 54, 159–175. [Google Scholar] [CrossRef]
- Dimiza, M.D.; Triantaphyllou, M.V.; Malinverno, E.; Psarra, E.; Karatsolis, B.-T.; Mara, P.; Lagaria, A.; Gogou, A. The composition and distribution of living coccolithophores in the Aegean Sea (NE Mediterranean). Micropaleontology 2015, 61, 521–540. [Google Scholar] [CrossRef]
- Cros, L. Planktonic Coccolithophores of the NW Mediterranean. Ph.D. Thesis, Universitat de Barcelona, Barcelona, Spain, 2001; 181p. [Google Scholar]
- Malinverno, E.; Triantaphyllou, M.V.; Stavrakakis, S.; Ziveri, P.; Lykousis, V. Seasonal and spatial variability of coccolithophore export production at the southwestern margin of Crete (eastern Mediterranean). Mar. Micropaleontol. 2009, 71, 131–147. [Google Scholar] [CrossRef]
- Oviedo, A.; Ziveri, P.; Álvarez, M.; Tanhua, T. Is coccolithophore distribution in the Mediterranean Sea related to seawater carbonate chemistry? Ocean Sci. 2015, 11, 13–32. [Google Scholar] [CrossRef]
- Skejić, S.; Arapov, J.; Kovačević, V.; Bužancić, M.; Bensi, M.; Giani, M.; Bakrač, A.; Mihanović, H.; Ninčević Gladan, Ž.; Urbini, L.; et al. Coccolithophore diversity in open waters of the middle Adriatic Sea in pre- and post-winter periods. Mar. Micropaleontol. 2018, 143, 30–45. [Google Scholar] [CrossRef]
- Weeks, S.J.; Pitcher, G.C.; Bernard, S. Satellite monitoring of the evolution of a coccolithophorid bloom in the Southern Benguela upwelling system. Oceanography 2003, 17, 83–89. [Google Scholar] [CrossRef]
- Daniels, C.J.; Tyrrell, T.; Poulton, A.J.; Young, J.R. A mixed life-cycle stage bloom of Syracosphaera bannockii (Borsetti and Cati, 1976) Cros et al. 2000 (Bay of Biscay, April 2010). J. Nannoplankton Res. 2014, 34, 31–35. [Google Scholar] [CrossRef]
- Skejić, S.; Arapov, J.; Bužancić, M.; Ninčević-Gladan, Ž.; Bakrač, A.; Straka, M.; Mandić, J. First evidence of an intensive bloom of the coccolithophore Syracosphaera halldalii in a highly variable estuarine environment (Krka River, Adriatic sea). Mar. Ecol. 2021, 42, e12641. [Google Scholar] [CrossRef]
- Kleijne, A. Morphology, Taxonomy and Distribution of Extant Coccolithophorids (Calcareous Nannoplankton). Ph.D. Thesis, Vrije Universiteit Amsterdam, Enschede, The Netherlands, 1993; 321p. [Google Scholar]
- Cerino, F.; Malinverno, E.; Fornasaro, D.; Kralj, M.; Cabrini, M. Coccolithophore diversity and dynamics at a coastal site in the Gulf of Trieste (northern Adriatic Sea). Estuar. Coast. Shelf Sci. 2017, 196, 331–345. [Google Scholar] [CrossRef]
- Šupraha, L.; Ljubešić, Z.; Mihanović, H.; Henderiks, J. Coccolithophore life-cycle dynamics in a coastal Mediterranean ecosystem: Seasonality and species-specific patterns. J. Plankton Res. 2016, 38, 1178–1193. [Google Scholar] [CrossRef]
- Jordan, R.W.; Winter, A. Assemblages of coccolithophorids and other living microplankton off the coast of Puerto Rico during January–May 1995. Mar. Micropaleontol. 2000, 39, 113–130. [Google Scholar] [CrossRef]
- D’Amario, B.; Ziveri, P.; Grelaud, M.; Oviedo, A.; Kralj, M. Coccolithophore haploid and diploid distribution patterns in the Mediterranean Sea: Can a haplo-diploid life cycle be advantageous under climate change? J. Plankton Res. 2017, 39, 781–794. [Google Scholar] [CrossRef]
- Godrijan, J.; Young, J.R.; Marić Pfannkuchen, D.; Precali, R.; Pfannkuchen, M. Coastal zones as important habitats of coccolithophores: A study of species diversity, succession, and life-cycle phases. Limnol. Oceanogr. 2018, 63, 1692–1710. [Google Scholar] [CrossRef]
- Keuter, S.; Silverman, J.; Krom, M.D.; Sisma-Ventura, G.; Yu, J.; Tsemel, A.; Ben-Ezra, T.; Sher, D.; Reich, T.; Koplovitz, G.; et al. Seasonal patterns of coccolithophores in the ultra-oligotrophic South-East Levantine Basin, Eastern Mediterranean Sea. Mar. Micropaleontol. 2022, 175, 102153. [Google Scholar] [CrossRef]
- Moncheva, S.; Gotsis-Skretas, O.; Pagou, K.; Krastev, A. Phytoplankton Blooms in Black Sea and Mediterranean Coastal Ecosystems Subjected to Anthropogenic Eutrophication: Similarities and Differences. Estuar. Coast. Shelf Sci. 2001, 53, 281–295. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Station | Bottom Depth (m) | Sampling Depth (m) | E. huxleyi (×104 cells L−1) | Syracosphaera (×104 cells L−1) | Acanthoica (×104 cells L−1) | A. robusta (×104 cells L−1) | |
|---|---|---|---|---|---|---|---|
| inner shelf | 1 | 33 | 1 | 257.23 | 2.16 | 0.00 | 0.00 |
| 2 | 48 | 1 | 371.28 | 0.00 | 0.00 | 0.00 | |
| 2 | 15 | 474.05 | 26.02 | 0.77 | 0.00 | ||
| 3 | 26 | 4 | 214.25 | 3.42 | 1.04 | 0.00 | |
| 3 | 8 | 282.68 | 8.93 | 0.77 | 0.15 | ||
| 4 | 22 | 1 | 299.65 | 0.00 | 0.00 | 0.00 | |
| 5 | 50 | 1 | 470.21 | 0.00 | 0.00 | 0.00 | |
| 5 | 15 | 737.64 | 25.40 | 0.15 | 0.00 | ||
| 6 | 50 | 1 | 302.24 | 0.00 | 0.00 | 0.00 | |
| 6 | 7 | 316.83 | 0.86 | 0.00 | 0.00 | ||
| 6 | 35 | 1.73 | 0.58 | 0.00 | 0.14 | ||
| 7 | 42 | 1 | 258.72 | 0.00 | 0.00 | 0.00 | |
| 7 | 9 | 442.05 | 1.11 | 0.14 | 0.00 | ||
| 7 | 30 | 1.78 | 0.89 | 0.00 | 0.00 | ||
| outer shelf | 8 | 68 | 1 | 134.49 | 0.59 | 0.13 | 0.00 |
| 8 | 5 | 130.30 | 1.21 | 0.57 | 0.00 | ||
| 8 | 15 | 242.94 | 3.03 | 0.25 | 0.00 | ||
| 8 | 25 | 31.44 | 4.81 | 0.03 | 0.10 | ||
| 9 | 75 | 1 | 143.98 | 0.00 | 0.00 | 0.00 | |
| 10 | 67 | 1 | 271.95 | 2.61 | 0.00 | 0.00 | |
| 10 | 15 | 187.17 | 23.29 | 3.02 | 0.00 | ||
| 11 | 65 | 3 | 188.95 | 5.01 | 0.28 | 0.00 | |
| 12 | 85 | 4 | 174.34 | 3.90 | 0.42 | 0.00 | |
| 12 | 16 | 66.82 | 7.70 | 1.08 | 0.00 | ||
| 12 | 45 | 13.51 | 3.31 | 0.00 | 1.29 | ||
| 13 | 101 | 1 | 105.68 | 3.83 | 0.17 | 0.00 | |
| 13 | 20 | 15.96 | 7.04 | 0.29 | 0.00 | ||
| open-sea | 14 | 1353 | 1 | 45.84 | 0.00 | 0.00 | 0.00 |
| 14 | 5 | 87.80 | 0.39 | 0.24 | 0.00 | ||
| 14 | 15 | 95.20 | 1.17 | 0.08 | 0.00 | ||
| 14 | 25 | 60.10 | 1.62 | 0.00 | 0.02 | ||
| 14 | 35 | 30.10 | 0.85 | 0.04 | 1.10 | ||
| 15 | 2165 | 1 | 132.79 | 0.23 | 0.18 | 0.00 | |
| 15 | 10 | 445.38 | 2.75 | 0.08 | 0.00 | ||
| 15 | 25 | 419.37 | 3.95 | 0.04 | 0.00 | ||
| 15 | 45 | 35.90 | 0.35 | 0.00 | 79.45 | ||
| 16 | 2150 | 1 | 72.51 | 0.00 | 0.00 | 0.00 | |
| 16 | 10 | 83.10 | 0.00 | 0.00 | 0.00 | ||
| 16 | 25 | 68.50 | 0.00 | 0.00 | 0.00 | ||
| 16 | 45 | 3.80 | 0.00 | 0.00 | 1.60 | ||
| 17 | 2204 | 1 | 89.06 | 0.00 | 0.00 | 0.00 | |
| 17 | 10 | 184.00 | 0.54 | 0.31 | 0.04 | ||
| 17 | 25 | 45.00 | 0.37 | 0.03 | 0.18 | ||
| 18 | 2205 | 1 | 111.88 | 0.00 | 0.00 | 0.00 | |
| 18 | 10 | 110.00 | 0.70 | 0.91 | 0.00 | ||
| 18 | 20 | 241.00 | 0.06 | 0.59 | 0.12 | ||
| 18 | 35 | 42.20 | 0.61 | 0.02 | 0.16 | ||
| 18 | 50 | 6.82 | 0.63 | 0.00 | 9.91 |
| E. huxleyi | Syracosphaera | Acanthoica | A. robusta | |
|---|---|---|---|---|
| temperature | 0.46 | −0.15 | 0.11 | −0.65 |
| salinity | −0.43 | −0.17 | −0.12 | 0.53 |
| chl-a | 0.57 | 0.25 | 0.19 | −0.26 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dimiza, M.D.; Triantaphyllou, M.V.; Ravani, A.; Malinverno, E.; Karatsolis, B.T.; Psarra, S.; Karageorgis, A.P. Coccolithophore Distribution in the Western Black Sea in the Summer of 2016. Diversity 2023, 15, 1194. https://doi.org/10.3390/d15121194
Dimiza MD, Triantaphyllou MV, Ravani A, Malinverno E, Karatsolis BT, Psarra S, Karageorgis AP. Coccolithophore Distribution in the Western Black Sea in the Summer of 2016. Diversity. 2023; 15(12):1194. https://doi.org/10.3390/d15121194
Chicago/Turabian StyleDimiza, Margarita D., Maria V. Triantaphyllou, Alexandra Ravani, Elisa Malinverno, Boris T. Karatsolis, Stella Psarra, and Aristomenis P. Karageorgis. 2023. "Coccolithophore Distribution in the Western Black Sea in the Summer of 2016" Diversity 15, no. 12: 1194. https://doi.org/10.3390/d15121194