Figure 1.
Representative member of the genus Brevitrygon highlighting key features of the dorsal disc and tail, modified after Chandy (1957). Abbreviations: id = interorbital denticles, pc = pectoral fin, pv = pelvic fin, sd = secondary denticle band, sn = snout, sp = spiracle, spr = inner ridge of spiracular pit, ssd = scapular denticles, st1 and st2 = caudal stings, tt = tail thorns.
Figure 1.
Representative member of the genus Brevitrygon highlighting key features of the dorsal disc and tail, modified after Chandy (1957). Abbreviations: id = interorbital denticles, pc = pectoral fin, pv = pelvic fin, sd = secondary denticle band, sn = snout, sp = spiracle, spr = inner ridge of spiracular pit, ssd = scapular denticles, st1 and st2 = caudal stings, tt = tail thorns.
Figure 2.
Oronasal region of (a) Brevitrygon imbricata, non-type NTM S 13160-009, female 186 mm DW, Sri Lanka (preserved); (b) B. walga, non-type CSIRO H 8666-01, female 233 mm DW, Myanmar (preserved); (c) B. heterura, non-type CSIRO H 5584-07, female 200 mm DW, Sabah, Malaysia (preserved); (d) B. javaensis, holotype MZB 21461, female 164 mm DW, Cilacap, Indonesia (preserved); (e) B. manjajiae sp. nov., holotype USNM 222555, adult male 231 mm DW, Pakistan (preserved).
Figure 2.
Oronasal region of (a) Brevitrygon imbricata, non-type NTM S 13160-009, female 186 mm DW, Sri Lanka (preserved); (b) B. walga, non-type CSIRO H 8666-01, female 233 mm DW, Myanmar (preserved); (c) B. heterura, non-type CSIRO H 5584-07, female 200 mm DW, Sabah, Malaysia (preserved); (d) B. javaensis, holotype MZB 21461, female 164 mm DW, Cilacap, Indonesia (preserved); (e) B. manjajiae sp. nov., holotype USNM 222555, adult male 231 mm DW, Pakistan (preserved).
Figure 3.
Lateral (a,b,d,f,h) and dorsal (c,e,g,i) views of the post-sting tail of females of Brevitrygon species. (a) B. imbricata non-type BPBM 27597 (1 of 2), female 202 mm DW, Kerala, India (preserved) showing a shortened tail with obvious dorsal and ventral skin folds; (b,c) B. walga non-type CSIRO H 8666-01, female 233 mm DW, Nabule, Myanmar (preserved); (d,e) B. heterura non-type CSIRO H 5584-07, female 200 mm DW, Sabah, Malaysia; (f,g) B. javaensis holotype MZB 21461, female 164 mm DW, Cilacap, Indonesia; (h,i) B. manjajiae sp. nov. paratype LACM 38134-37, female 207 mm DW, Sindh, Pakistan (preserved).
Figure 3.
Lateral (a,b,d,f,h) and dorsal (c,e,g,i) views of the post-sting tail of females of Brevitrygon species. (a) B. imbricata non-type BPBM 27597 (1 of 2), female 202 mm DW, Kerala, India (preserved) showing a shortened tail with obvious dorsal and ventral skin folds; (b,c) B. walga non-type CSIRO H 8666-01, female 233 mm DW, Nabule, Myanmar (preserved); (d,e) B. heterura non-type CSIRO H 5584-07, female 200 mm DW, Sabah, Malaysia; (f,g) B. javaensis holotype MZB 21461, female 164 mm DW, Cilacap, Indonesia; (h,i) B. manjajiae sp. nov. paratype LACM 38134-37, female 207 mm DW, Sindh, Pakistan (preserved).
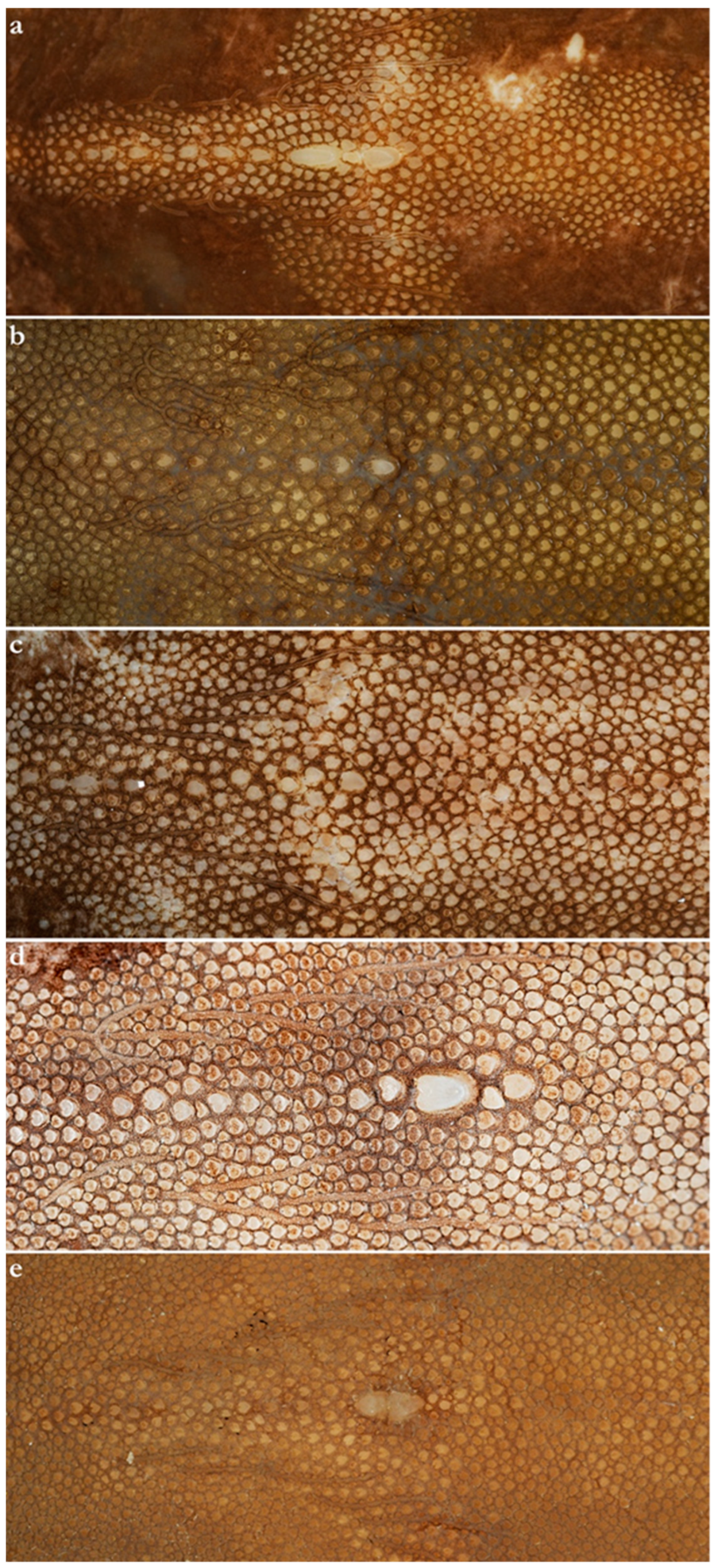
Figure 4.
Squamation of the central disc in the genus Brevitrygon—secondary denticle band. (a) B. imbricata, non-type NTM S 13160-009, female 186 mm DW, Sri Lanka (preserved); (b) B. walga, non-type CSIRO H 8666-01, female 233 mm DW, Myanmar (preserved); (c) B. heterura, non-type CSIRO H 5584-07, female 200 mm DW, Sabah, Malaysia (preserved); (d) B. javaensis, holotype MZB 21461, female 164 mm DW, Cilacap, Indonesia (preserved); (e) B. manjajiae sp. nov., holotype USNM 222555, adult male 231 mm DW, Pakistan (preserved).
Figure 4.
Squamation of the central disc in the genus Brevitrygon—secondary denticle band. (a) B. imbricata, non-type NTM S 13160-009, female 186 mm DW, Sri Lanka (preserved); (b) B. walga, non-type CSIRO H 8666-01, female 233 mm DW, Myanmar (preserved); (c) B. heterura, non-type CSIRO H 5584-07, female 200 mm DW, Sabah, Malaysia (preserved); (d) B. javaensis, holotype MZB 21461, female 164 mm DW, Cilacap, Indonesia (preserved); (e) B. manjajiae sp. nov., holotype USNM 222555, adult male 231 mm DW, Pakistan (preserved).
Figure 5.
Squamation in the genus Brevitrygon—dorsolateral view of pre-sting tail. (a) B. imbricata, non-type NTM S 13160-009, female 186 mm DW, Sri Lanka (preserved); (b) B. walga, non-type CSIRO H 8666-01, female 233 mm DW, Myanmar (preserved); (c) B. heterura, non-type CSIRO H 5584-07, female 200 mm DW, Sabah, Malaysia (preserved); (d) B. javaensis, holotype MZB 21461, female 164 mm DW, Cilacap, Indonesia (preserved); (e) B. manjajiae sp. nov., holotype USNM 222555, adult male 231 mm DW, Pakistan (preserved).
Figure 5.
Squamation in the genus Brevitrygon—dorsolateral view of pre-sting tail. (a) B. imbricata, non-type NTM S 13160-009, female 186 mm DW, Sri Lanka (preserved); (b) B. walga, non-type CSIRO H 8666-01, female 233 mm DW, Myanmar (preserved); (c) B. heterura, non-type CSIRO H 5584-07, female 200 mm DW, Sabah, Malaysia (preserved); (d) B. javaensis, holotype MZB 21461, female 164 mm DW, Cilacap, Indonesia (preserved); (e) B. manjajiae sp. nov., holotype USNM 222555, adult male 231 mm DW, Pakistan (preserved).
Figure 6.
Squamation in the genus Brevitrygon—scapular denticles. (a) B. imbricata, non-type NTM S 13160-009, female 186 mm DW, Sri Lanka (preserved); (b) B. walga, non-type CSIRO H 8666-01, female 233 mm DW, Myanmar (preserved); (c) B. heterura, non-type CSIRO H 5584-07, female 200 mm DW, Sabah, Malaysia (preserved); (d) B. javaensis, holotype MZB 21461, female 164 mm DW, Cilacap, Indonesia (preserved); (e) B. manjajiae sp. nov., holotype USNM 222555, adult male 231 mm DW, Pakistan (preserved).
Figure 6.
Squamation in the genus Brevitrygon—scapular denticles. (a) B. imbricata, non-type NTM S 13160-009, female 186 mm DW, Sri Lanka (preserved); (b) B. walga, non-type CSIRO H 8666-01, female 233 mm DW, Myanmar (preserved); (c) B. heterura, non-type CSIRO H 5584-07, female 200 mm DW, Sabah, Malaysia (preserved); (d) B. javaensis, holotype MZB 21461, female 164 mm DW, Cilacap, Indonesia (preserved); (e) B. manjajiae sp. nov., holotype USNM 222555, adult male 231 mm DW, Pakistan (preserved).
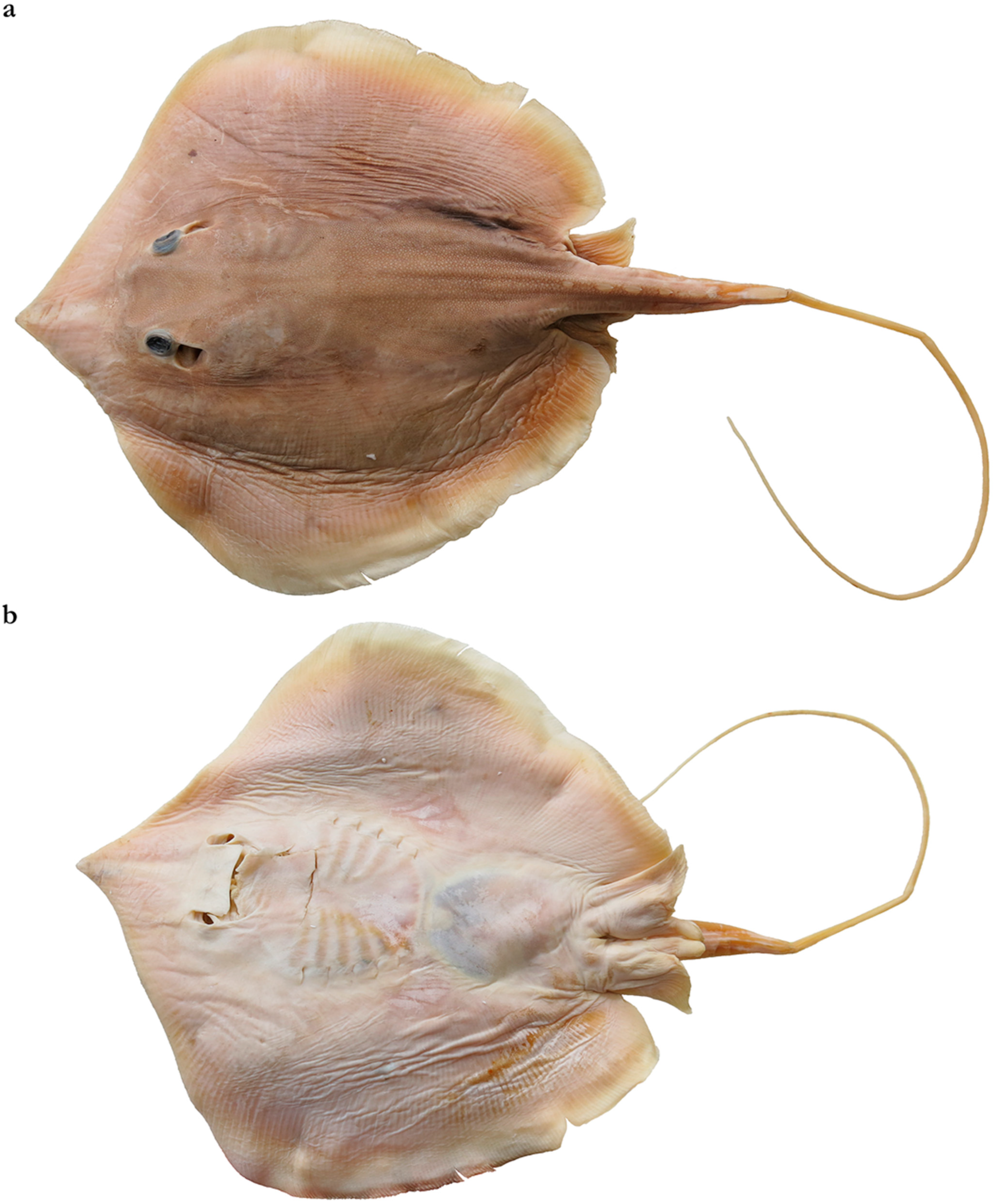
Figure 7.
Raja imbricata Bloch & Schneider (=B. imbricata), holotype ZMB 7585, adult male ~171 mm DW, Tharangambadi, India: (a) dorsal surface, dry mounted; (b) close up of the typically narrow secondary denticle band on the disc and anterior tail. Scale bar: 5 cm.
Figure 7.
Raja imbricata Bloch & Schneider (=B. imbricata), holotype ZMB 7585, adult male ~171 mm DW, Tharangambadi, India: (a) dorsal surface, dry mounted; (b) close up of the typically narrow secondary denticle band on the disc and anterior tail. Scale bar: 5 cm.
Figure 8.
Dorsal surfaces of (a) Trygon walga not Müller & Henle (=B. imbricata), syntype MNHN 0000-2439 (1 of 2), female 173 mm DW, Coromandel, India; (b) Trygon dadong Bleeker (=B. imbricata), holotype RMNH.PISC.7446, female 163 mm DW, Bitan Island, Indonesia.
Figure 8.
Dorsal surfaces of (a) Trygon walga not Müller & Henle (=B. imbricata), syntype MNHN 0000-2439 (1 of 2), female 173 mm DW, Coromandel, India; (b) Trygon dadong Bleeker (=B. imbricata), holotype RMNH.PISC.7446, female 163 mm DW, Bitan Island, Indonesia.
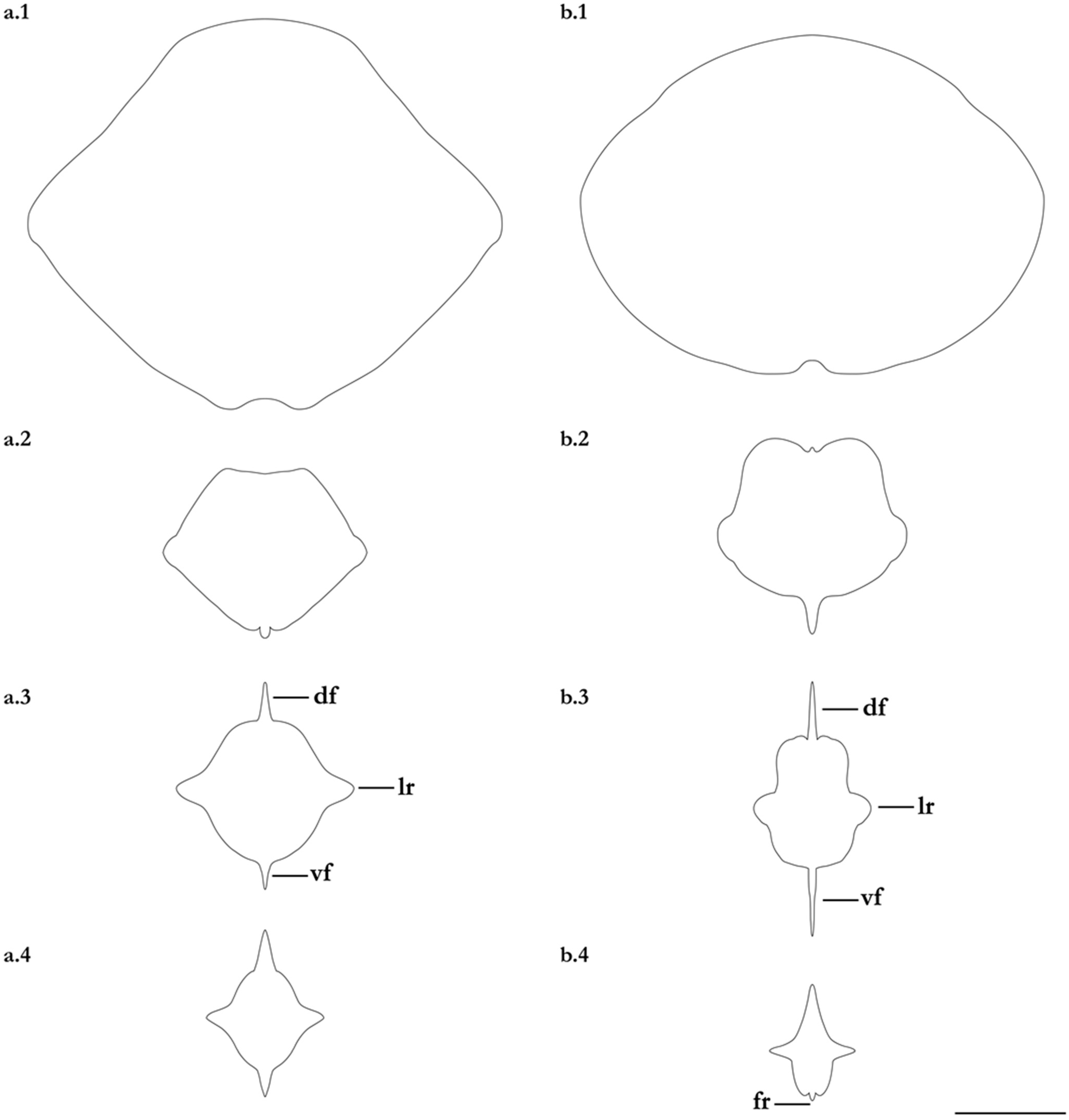
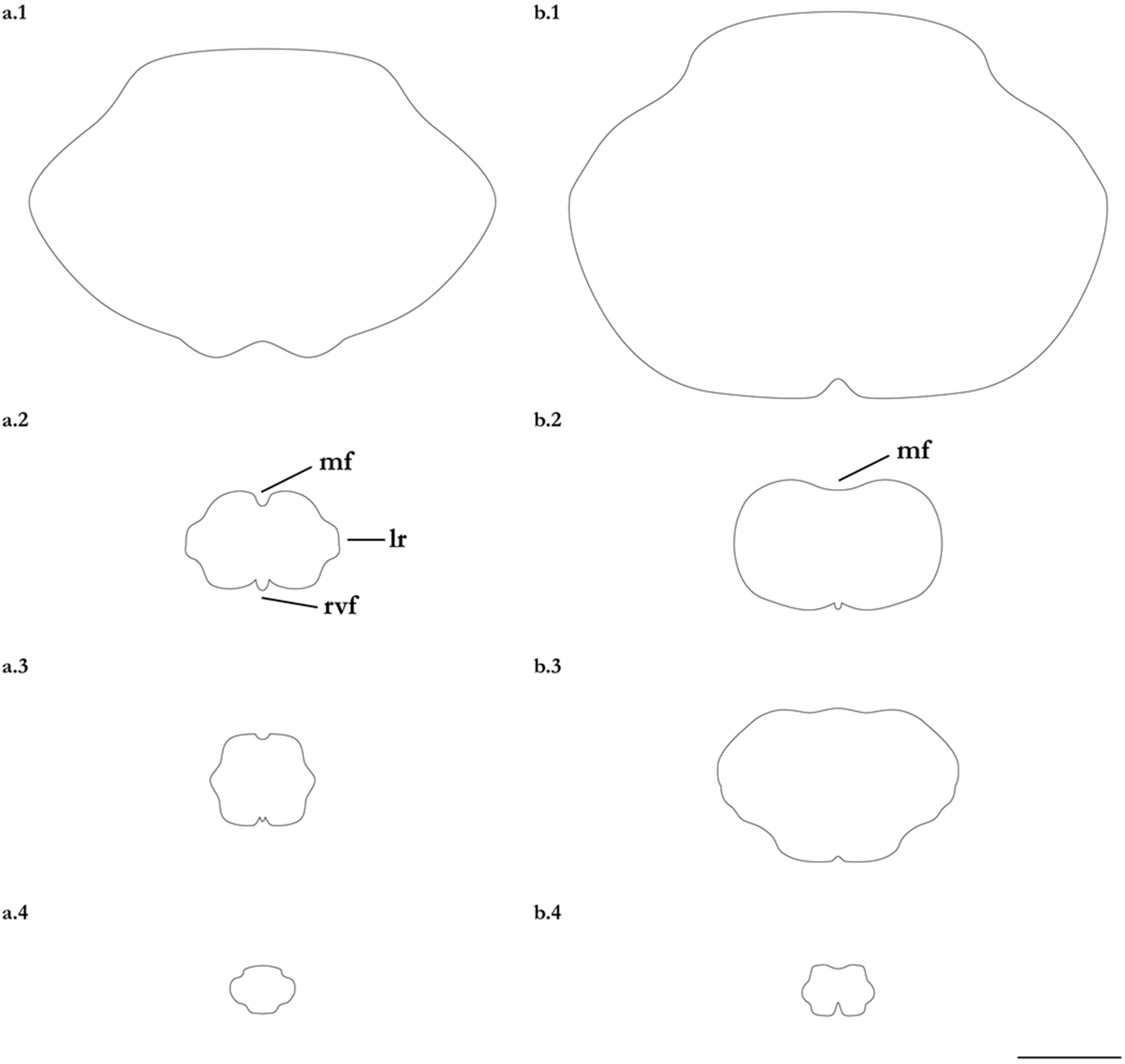
Figure 9.
Schematic cross-sections of the post-sting tail of Brevitrygon imbricata for (a) non-type BPBM 27597, adult male 205 mm DW, Kerala, India; and (b) non-type NTM S 13160-009, female 186 mm DW, Sri Lanka. Sections taken at: 1. sting origin; 2. axis 25% from origin; 3. through midlength; and 4. axis 75% from origin. Abbreviations: df = dorsal cutaneous fold, fr = low fleshy ridge, lr = lateral ridge, vf = ventral cutaneous fold. Scale bar: 2 mm.
Figure 9.
Schematic cross-sections of the post-sting tail of Brevitrygon imbricata for (a) non-type BPBM 27597, adult male 205 mm DW, Kerala, India; and (b) non-type NTM S 13160-009, female 186 mm DW, Sri Lanka. Sections taken at: 1. sting origin; 2. axis 25% from origin; 3. through midlength; and 4. axis 75% from origin. Abbreviations: df = dorsal cutaneous fold, fr = low fleshy ridge, lr = lateral ridge, vf = ventral cutaneous fold. Scale bar: 2 mm.
Figure 10.
Brevitrygon imbricata, non-type BPBM 27597 (2 of 2), adult male 205 mm DW, Kerala, India (preserved): (a) dorsal surface; (b) ventral surface. Scale bar: 2 cm.
Figure 10.
Brevitrygon imbricata, non-type BPBM 27597 (2 of 2), adult male 205 mm DW, Kerala, India (preserved): (a) dorsal surface; (b) ventral surface. Scale bar: 2 cm.
Figure 11.
Brevitrygon imbricata, non-type BPBM 27597 (1 of 2), female 202 mm DW, Kerala, India (preserved): (a) dorsal surface; (b) ventral surface. Scale bar: 2 cm.
Figure 11.
Brevitrygon imbricata, non-type BPBM 27597 (1 of 2), female 202 mm DW, Kerala, India (preserved): (a) dorsal surface; (b) ventral surface. Scale bar: 2 cm.
Figure 12.
Dorsal disc and tail of early juveniles of (a) B. imbricata (USNM 222564, juvenile female 88 mm DW, Sri Lanka); (b) B. walga (CSIRO H 8666-02, late embryo male 82 mm DW, Myanmar); (c) B. heterura (BPBM 26574, juvenile female 104 mm DW, Indonesia); (d) B. javaensis (AMS I 46280-001, juvenile male 120 mm DW, Indonesia); (e) B. manjajiae sp. nov. (BPBM 33199 (1 of 2), juvenile male 103 mm DW, Kuwait). Scale bars: 2 cm.
Figure 12.
Dorsal disc and tail of early juveniles of (a) B. imbricata (USNM 222564, juvenile female 88 mm DW, Sri Lanka); (b) B. walga (CSIRO H 8666-02, late embryo male 82 mm DW, Myanmar); (c) B. heterura (BPBM 26574, juvenile female 104 mm DW, Indonesia); (d) B. javaensis (AMS I 46280-001, juvenile male 120 mm DW, Indonesia); (e) B. manjajiae sp. nov. (BPBM 33199 (1 of 2), juvenile male 103 mm DW, Kuwait). Scale bars: 2 cm.
Figure 13.
Map of the Indo-West Pacific showing the localities of the primary types (symbols) and confirmed ranges (polygons) of Brevitrygon imbricata (black circle, green polygons; occurrences off Myanmar, Singapore, and Taiwan questionable, indicated by green question marks), B. walga (white circle, red polygon), B. heterura (black star, yellow polygon), B. javaensis (white star, pink polygon; yellow stripes indicate overlapping range with B. heterura), and B. manjajiae sp. nov. (black triangle, orange polygon; occurrence in the Red Sea questionable, indicated by orange question mark).
Figure 13.
Map of the Indo-West Pacific showing the localities of the primary types (symbols) and confirmed ranges (polygons) of Brevitrygon imbricata (black circle, green polygons; occurrences off Myanmar, Singapore, and Taiwan questionable, indicated by green question marks), B. walga (white circle, red polygon), B. heterura (black star, yellow polygon), B. javaensis (white star, pink polygon; yellow stripes indicate overlapping range with B. heterura), and B. manjajiae sp. nov. (black triangle, orange polygon; occurrence in the Red Sea questionable, indicated by orange question mark).
Figure 14.
Illustrations of Brevitrygon from Bleeker’s (1874) ichthyological atlas: (a) Leiobatis (Himantura) heterurus (=B. heterura, female, Pl. 35, 1a,b); (b) Leiobatis (Himantura) walga (=B. heterura, male, Pl. 41, 1a,b); (c) Leiobatis (Himantura) dadong (=B. imbricata, Pl. 36, 1a).
Figure 14.
Illustrations of Brevitrygon from Bleeker’s (1874) ichthyological atlas: (a) Leiobatis (Himantura) heterurus (=B. heterura, female, Pl. 35, 1a,b); (b) Leiobatis (Himantura) walga (=B. heterura, male, Pl. 41, 1a,b); (c) Leiobatis (Himantura) dadong (=B. imbricata, Pl. 36, 1a).
Figure 15.
Brevitrygon cf. imbricata, non-type SAM—LJVC 517, adult male 185 mm DW, Taiwan: (a) dorsal surface, fresh; (b) ventral surface, preserved. Scale bar: 2 cm.
Figure 15.
Brevitrygon cf. imbricata, non-type SAM—LJVC 517, adult male 185 mm DW, Taiwan: (a) dorsal surface, fresh; (b) ventral surface, preserved. Scale bar: 2 cm.
Figure 16.
Trygon walga Müller & Henle, lectotype MNHN 0000-2431, female 170 mm DW, Ganges Delta, India, preserved: (a) dorsal surface, (b) ventral surface.
Figure 16.
Trygon walga Müller & Henle, lectotype MNHN 0000-2431, female 170 mm DW, Ganges Delta, India, preserved: (a) dorsal surface, (b) ventral surface.
Figure 17.
Trygon walga Müller & Henle, putative syntype BMNH 1889.2.1.4196, female 223 mm DW, Chennai, India, preserved: dorsal surface.
Figure 17.
Trygon walga Müller & Henle, putative syntype BMNH 1889.2.1.4196, female 223 mm DW, Chennai, India, preserved: dorsal surface.
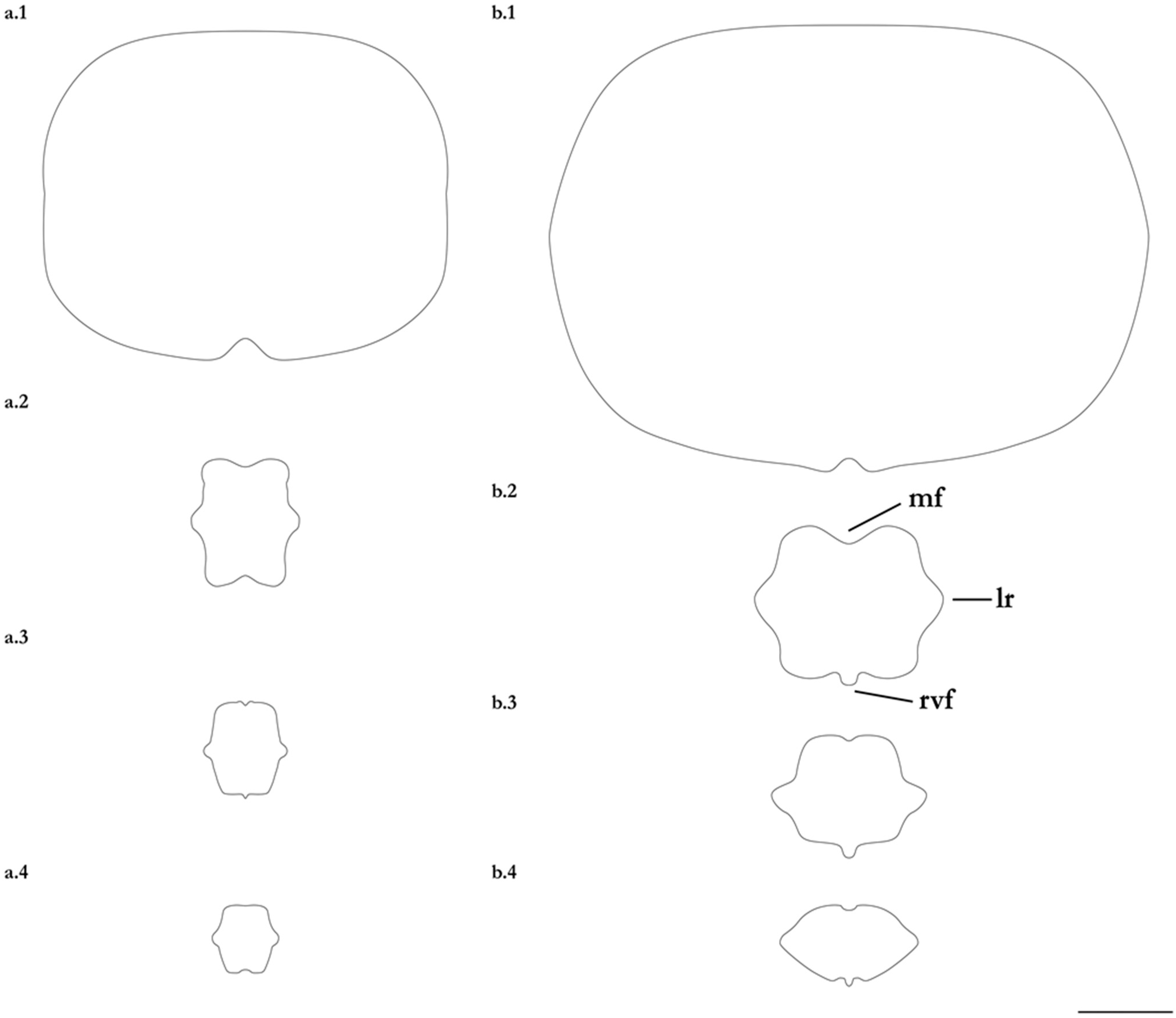
Figure 18.
Schematic cross-sections of the post-sting tail of Brevitrygon walga for: (a) non-type CSIRO H 8667-01, adult male 193 mm DW, Myanmar; and (b) non-type CSIRO H 8669-01, female 197 mm DW, Myanmar. Sections taken at: 1. sting origin; 2. axis 25% distance from origin; 3. through midlength; and 4. axis 75% distance from origin. Abbreviations: mf = median furrow or groove, lr = lateral ridge, rvf = rudimentary ventral fold. Scale bar: 2 mm.
Figure 18.
Schematic cross-sections of the post-sting tail of Brevitrygon walga for: (a) non-type CSIRO H 8667-01, adult male 193 mm DW, Myanmar; and (b) non-type CSIRO H 8669-01, female 197 mm DW, Myanmar. Sections taken at: 1. sting origin; 2. axis 25% distance from origin; 3. through midlength; and 4. axis 75% distance from origin. Abbreviations: mf = median furrow or groove, lr = lateral ridge, rvf = rudimentary ventral fold. Scale bar: 2 mm.
Figure 19.
Brevitrygon walga, non-type CSIRO H 8667-01, adult male 193 mm DW, Myanmar, fresh: (a) dorsal surface, (b) ventral surface (photos P. Psomadakis). Scale bar: 2 cm.
Figure 19.
Brevitrygon walga, non-type CSIRO H 8667-01, adult male 193 mm DW, Myanmar, fresh: (a) dorsal surface, (b) ventral surface (photos P. Psomadakis). Scale bar: 2 cm.
Figure 20.
Brevitrygon walga, not retained, female 218 mm DW, Myanmar, fresh: (a) dorsal surface, (b) ventral surface (photos P. Psomadakis). Scale bar: 2 cm.
Figure 20.
Brevitrygon walga, not retained, female 218 mm DW, Myanmar, fresh: (a) dorsal surface, (b) ventral surface (photos P. Psomadakis). Scale bar: 2 cm.
Figure 21.
(a) Trygon walga Müller & Henle 1841, Systematische Beschreibung der Plagiostomen, Pl. 51 from manuscript, ~105 mm DW. (b) Trygon nuda Günther 1870, syntype BMNH 1845.3.7.19, 105 mm DW.
Figure 21.
(a) Trygon walga Müller & Henle 1841, Systematische Beschreibung der Plagiostomen, Pl. 51 from manuscript, ~105 mm DW. (b) Trygon nuda Günther 1870, syntype BMNH 1845.3.7.19, 105 mm DW.
Figure 22.
Brevitrygon walga, non-type MTUF 29999, female 184 mm DW, Hooghly River, India: dorsal surface, preserved. Scale bar: 2 cm.
Figure 22.
Brevitrygon walga, non-type MTUF 29999, female 184 mm DW, Hooghly River, India: dorsal surface, preserved. Scale bar: 2 cm.
Figure 23.
Trygon heterurus Bleeker, holotype BMNH 1867.11.28.158, female 162 mm DW, Jakarta, Indonesia, preserved): (a) dorsal surface; (b) ventral surface.
Figure 23.
Trygon heterurus Bleeker, holotype BMNH 1867.11.28.158, female 162 mm DW, Jakarta, Indonesia, preserved): (a) dorsal surface; (b) ventral surface.
Figure 24.
Trygon walga not Müller & Henle (=B. heterura), syntype RMNH.PISC.2453, female 167 mm DW, Java, Indonesia: dorsal surface, dry mounted.
Figure 24.
Trygon walga not Müller & Henle (=B. heterura), syntype RMNH.PISC.2453, female 167 mm DW, Java, Indonesia: dorsal surface, dry mounted.
Figure 25.
Brevitrygon heterura, non-type KA 351, adult male 162 mm DW, Kalimantan, Indonesia: (a) dorsal surface, fresh; (b) ventral surface, fresh (photos K. Jensen). Scale bar: 2 cm.
Figure 25.
Brevitrygon heterura, non-type KA 351, adult male 162 mm DW, Kalimantan, Indonesia: (a) dorsal surface, fresh; (b) ventral surface, fresh (photos K. Jensen). Scale bar: 2 cm.
Figure 26.
Brevitrygon heterura, non-type Phuket fishing port, female 162 mm DW, Phuket, Thailand: (a) dorsal surface, fresh; (b) ventral surface, fresh.
Figure 26.
Brevitrygon heterura, non-type Phuket fishing port, female 162 mm DW, Phuket, Thailand: (a) dorsal surface, fresh; (b) ventral surface, fresh.
Figure 27.
Schematic cross-sections of the post-sting tail of Brevitrygon heterura for (a) non-type CSIRO H 4924-02, adult male, 175 mm DW, Gulf of Thailand; and (b) non-type CSIRO H 4924-03, female 175 mm DW, Gulf of Thailand. Sections taken: 1. sting origin; 2. axis 25% distance from origin; 3. through midlength; and 4. axis 75% distance from origin. Abbreviations: mf = median furrow or groove, lr = lateral ridge, rvf = rudimentary ventral fold. Scale bar: 2 mm.
Figure 27.
Schematic cross-sections of the post-sting tail of Brevitrygon heterura for (a) non-type CSIRO H 4924-02, adult male, 175 mm DW, Gulf of Thailand; and (b) non-type CSIRO H 4924-03, female 175 mm DW, Gulf of Thailand. Sections taken: 1. sting origin; 2. axis 25% distance from origin; 3. through midlength; and 4. axis 75% distance from origin. Abbreviations: mf = median furrow or groove, lr = lateral ridge, rvf = rudimentary ventral fold. Scale bar: 2 mm.
Figure 28.
Brevitrygon heterura, non-type ZMH, adult female 185 mm DW, Prachuap Khiri Khan, Thailand: (a) dorsal surface, preserved; (b) ventral surface, preserved. Scale bar: 2 cm.
Figure 28.
Brevitrygon heterura, non-type ZMH, adult female 185 mm DW, Prachuap Khiri Khan, Thailand: (a) dorsal surface, preserved; (b) ventral surface, preserved. Scale bar: 2 cm.
Figure 29.
Dasybatis uylenburgi Giltay (=Brevitrygon heterura), holotype IRSNB 38, adult male 180 mm DW, East Indies (Indonesia): (a) dorsal surface, preserved; (b) ventral surface, preserved.
Figure 29.
Dasybatis uylenburgi Giltay (=Brevitrygon heterura), holotype IRSNB 38, adult male 180 mm DW, East Indies (Indonesia): (a) dorsal surface, preserved; (b) ventral surface, preserved.
Figure 30.
Schematic cross-sections of the post-sting tail of Brevitrygon javaensis for (a) paratype CSIRO H 6129-01, adolescent male 173 mm DW, Cilacap, Indonesia; and (b) paratype CSIRO H 5860-08, female 197 mm DW, Cilacap, Indonesia. Sections taken at: 1. sting origin; 2. axis 25% distance from origin; 3. through midlength; and 4. axis 75% distance from origin. Abbreviations: mf = median furrow or groove, lr = lateral ridge. Scale bar: 2 mm.
Figure 30.
Schematic cross-sections of the post-sting tail of Brevitrygon javaensis for (a) paratype CSIRO H 6129-01, adolescent male 173 mm DW, Cilacap, Indonesia; and (b) paratype CSIRO H 5860-08, female 197 mm DW, Cilacap, Indonesia. Sections taken at: 1. sting origin; 2. axis 25% distance from origin; 3. through midlength; and 4. axis 75% distance from origin. Abbreviations: mf = median furrow or groove, lr = lateral ridge. Scale bar: 2 mm.
Figure 31.
Brevitrygon javaensis, holotype MZB 21461, female 164 mm DW, Cilacap, Indonesia: (a) dorsal surface (fresh); (b) ventral surface (preserved). Scale bar: 2 cm.
Figure 31.
Brevitrygon javaensis, holotype MZB 21461, female 164 mm DW, Cilacap, Indonesia: (a) dorsal surface (fresh); (b) ventral surface (preserved). Scale bar: 2 cm.
Figure 32.
Brevitrygon javaensis, paratype CSIRO H 6129-01, adult male 173 mm DW, Cilacap, Indonesia: dorsal surface (preserved).
Figure 32.
Brevitrygon javaensis, paratype CSIRO H 6129-01, adult male 173 mm DW, Cilacap, Indonesia: dorsal surface (preserved).
Figure 33.
Brevitrygon manjajiae sp. nov., holotype USNM 222555, adult male 231 mm DW, Miani Hor, Pakistan, Arabian Sea (preserved): (a) dorsal surface; (b) ventral surface. Scale bar: 2 cm.
Figure 33.
Brevitrygon manjajiae sp. nov., holotype USNM 222555, adult male 231 mm DW, Miani Hor, Pakistan, Arabian Sea (preserved): (a) dorsal surface; (b) ventral surface. Scale bar: 2 cm.
Figure 34.
Schematic cross-sections of the post-sting tail of Brevitrygon manjajiae sp. nov. for (a) paratype LACM 38314-24, adult male 246 mm DW, Pakistan; and (b) paratype LACM 38134-37 (1 of 2), female 207 mm DW, Pakistan. Sections taken at: 1. sting origin; 2. axis 25% distance from origin; 3. through midlength; and 4. axis 75% distance from origin. Abbreviations: mf = median furrow or groove, lr = lateral ridge, rvf = rudimentary ventral fold. Scale bar: 2 mm.
Figure 34.
Schematic cross-sections of the post-sting tail of Brevitrygon manjajiae sp. nov. for (a) paratype LACM 38314-24, adult male 246 mm DW, Pakistan; and (b) paratype LACM 38134-37 (1 of 2), female 207 mm DW, Pakistan. Sections taken at: 1. sting origin; 2. axis 25% distance from origin; 3. through midlength; and 4. axis 75% distance from origin. Abbreviations: mf = median furrow or groove, lr = lateral ridge, rvf = rudimentary ventral fold. Scale bar: 2 mm.
Figure 35.
Brevitrygon manjajiae sp. nov., paratype LACM 38134-37 (1 of 2), female 207 mm DW, Pakistan (preserved): (a) dorsal surface; (b) ventral surface. Scale bar: 2 cm.
Figure 35.
Brevitrygon manjajiae sp. nov., paratype LACM 38134-37 (1 of 2), female 207 mm DW, Pakistan (preserved): (a) dorsal surface; (b) ventral surface. Scale bar: 2 cm.
Figure 36.
Brevitrygon manjajiae sp. nov., upper jaws (a,d), oral papillae (b,e), and lower jaws (c,f) of (a–c) paratype LACM 38314-24, adult male 246 mm DW, Sonmiani Bay, Baluchistan, Pakistan and (d–f) paratype LACM 38134-37, female 207 mm DW, off mouth of Hojambro Creek, Sindh, Pakistan. Scale bars: 5 mm.
Figure 36.
Brevitrygon manjajiae sp. nov., upper jaws (a,d), oral papillae (b,e), and lower jaws (c,f) of (a–c) paratype LACM 38314-24, adult male 246 mm DW, Sonmiani Bay, Baluchistan, Pakistan and (d–f) paratype LACM 38134-37, female 207 mm DW, off mouth of Hojambro Creek, Sindh, Pakistan. Scale bars: 5 mm.
Figure 37.
Trygon walga non Müller & Henle, syntype MNHN 0000-2337, adult male 198 mm DW, Red Sea: dorsal surface. Scale bar: 2 cm.
Figure 37.
Trygon walga non Müller & Henle, syntype MNHN 0000-2337, adult male 198 mm DW, Red Sea: dorsal surface. Scale bar: 2 cm.
Figure 38.
Brevitrygon manjajiae sp. nov., non-type BPBM 33199 (2 of 2), female 187 mm DW, Kuwait: dorsal surface, preserved. Scale bar: 2 cm.
Figure 38.
Brevitrygon manjajiae sp. nov., non-type BPBM 33199 (2 of 2), female 187 mm DW, Kuwait: dorsal surface, preserved. Scale bar: 2 cm.
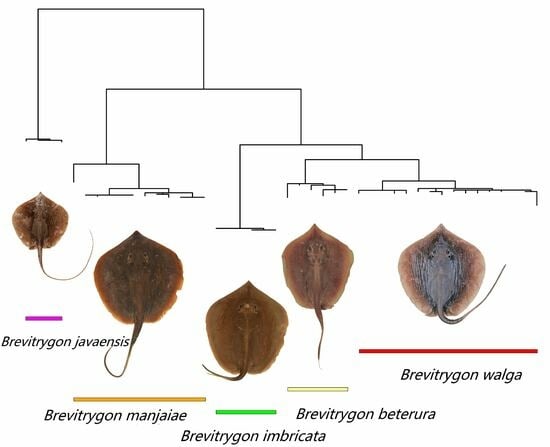
Figure 39.
Maximum likelihood (ML) tree based on General Time Reversible (GTR) model accommodating observed proportion of Invariant sites (I) and Gamma distributed distribution of Variable sites Γ (i.e., the GTR + I + Γ model).
Figure 39.
Maximum likelihood (ML) tree based on General Time Reversible (GTR) model accommodating observed proportion of Invariant sites (I) and Gamma distributed distribution of Variable sites Γ (i.e., the GTR + I + Γ model).
Table 1.
Morphometric data for Brevitrygon imbricata: putative syntype of Trygon walga Müller & Henle (MNHN 2438), non-types from India and Sri Lanka, and 3 specimens from Taiwan. Measurements expressed as percentages of disc width (mm).
Table 1.
Morphometric data for Brevitrygon imbricata: putative syntype of Trygon walga Müller & Henle (MNHN 2438), non-types from India and Sri Lanka, and 3 specimens from Taiwan. Measurements expressed as percentages of disc width (mm).
| | Pondicherry, India | Southern India and Sri Lanka | Taiwan |
|---|
| | MNHN 2438 | All Data (n = 11) | Males (n = 5) | Females (n = 6) | Males (n = 3) |
|---|
| | | Min | Max | Min | Max | Min | Max | Min | Max |
|---|
| Disc, width (mm) | 159 | 138 | 205 | 171 | 205 | 138 | 204 | 162 | 182 |
| Total length | 197.6 | 169.8 | 207.9 | 187.2 | 198.5 | 169.8 | 207.9 | 191.9 | 202.0 |
| Disc, length (direct) | 106.9 | 106.1 | 113.3 | 107.1 | 113.1 | 106.1 | 113.3 | 105.7 | 109.4 |
| Disc, thickness | 10.6 | 7.9 | 13.3 | 10.5 | 12.8 | 7.9 | 13.3 | 12.3 | 12.9 |
| Snout to origin of cloaca | 91.2 | 89.0 | 94.7 | 89.0 | 94.7 | 91.2 | 93.4 | 88.8 | 91.9 |
| Cloaca origin to tail tip | 106.4 | 76.6 | 116.5 | 94.4 | 105.4 | 76.6 | 116.5 | 100.9 | 110.7 |
| Snout to pectoral insertion | 98.6 | 95.0 | 103.8 | 95.7 | 103.8 | 95.0 | 103.2 | 95.4 | 98.9 |
| Snout to maximum width | 48.2 | 42.9 | 52.3 | 42.9 | 49.2 | 46.0 | 52.3 | 42.7 | 52.6 |
| End of orbit to pectoral insertion | 62.6 | 58.1 | 66.1 | 59.1 | 65.7 | 58.1 | 66.1 | 61.6 | 63.6 |
| Snout, preorbital (direct) | 30.5 | 29.0 | 31.9 | 29.7 | 31.1 | 29.0 | 31.9 | 28.1 | 31.8 |
| Snout, preorbital (horizontal) | 29.8 | 26.1 | 30.7 | 27.6 | 30.7 | 26.1 | 30.0 | 25.0 | 28.4 |
| Orbit diameter | 6.7 | 5.8 | 7.5 | 6.1 | 6.9 | 5.8 | 7.5 | 7.0 | 7.6 |
| Eye diameter | 4.7 | 3.8 | 5.0 | 3.8 | 5.0 | 4.0 | 4.5 | 4.2 | 4.4 |
| Spiracle length | 6.2 | 5.7 | 6.7 | 5.9 | 6.5 | 5.7 | 6.7 | 5.5 | 6.7 |
| Orbit and spiracle length | 10.0 | 9.7 | 11.1 | 10.4 | 11.1 | 9.7 | 10.8 | 10.1 | 11.3 |
| Interorbital width | 11.6 | 10.0 | 12.1 | 11.0 | 12.1 | 10.0 | 11.5 | 11.1 | 11.3 |
| Intereye width | 17.3 | 14.6 | 17.1 | 15.7 | 17.1 | 14.6 | 16.6 | 15.8 | 17.5 |
| Distance between spiracles | 15.5 | 15.5 | 16.6 | 15.5 | 16.6 | 15.6 | 16.4 | 16.8 | 17.9 |
| Head length (direct) | 57.0 | 55.2 | 58.6 | 56.5 | 58.6 | 55.2 | 57.8 | 55.7 | 59.1 |
| Snout, prenasal (direct) | 24.2 | 24.0 | 26.9 | 24.0 | 26.0 | 24.0 | 26.9 | 22.4 | 26.3 |
| Nostril length | 3.9 | 3.5 | 4.3 | 3.7 | 4.3 | 3.5 | 4.3 | 3.7 | 4.6 |
| Nasal curtain, length | 5.4 | 4.9 | 6.6 | 5.7 | 6.6 | 4.9 | 6.2 | 6.3 | 7.0 |
| Nasal curtain, width | 11.4 | 10.2 | 12.5 | 11.6 | 12.5 | 10.2 | 11.7 | 12.2 | 12.4 |
| Distance between nostrils | 11.8 | 10.1 | 12.1 | 10.7 | 12.1 | 10.1 | 10.9 | 12.3 | 13.1 |
| Snout, preoral (direct) | 30.6 | 29.8 | 32.4 | 29.8 | 31.7 | 30.0 | 32.4 | 27.3 | 32.0 |
| Mouth width | 9.8 | 9.3 | 10.9 | 9.6 | 10.9 | 9.3 | 10.5 | 8.4 | 9.4 |
| Width, 1st gill slit | 2.3 | 2.8 | 3.7 | 2.8 | 3.2 | 2.9 | 3.7 | 3.2 | 3.7 |
| Width, 3rd gill slit | 2.5 | 3.0 | 4.0 | 3.0 | 4.0 | 3.2 | 4.0 | 3.9 | 4.2 |
| Width, 5th gill slit | 1.7 | 2.3 | 2.7 | 2.3 | 2.7 | 2.4 | 2.7 | 2.5 | 2.6 |
| Distance between 1st gill slits | 23.2 | 22.0 | 24.4 | 23.1 | 24.4 | 22.0 | 24.3 | 23.9 | 24.7 |
| Distance between 5th gill slits | 14.9 | 13.5 | 15.3 | 13.6 | 15.3 | 13.5 | 15.3 | 14.6 | 15.4 |
| Length pelvic fin | 21.1 | 18.1 | 22.6 | 18.3 | 20.9 | 18.1 | 22.6 | 20.4 | 21.2 |
| Width across pelvic fin base | 14.7 | 15.5 | 20.3 | 15.5 | 18.2 | 17.1 | 20.3 | 16.4 | 17.1 |
| Greatest width across pelvic fins | 33.6 | 26.6 | 42.9 | 26.6 | 38.2 | 35.6 | 42.9 | 34.9 | 38.0 |
| Tail width, axil of pelvics | 8.1 | 7.3 | 9.8 | 8.2 | 9.8 | 7.3 | 9.1 | 7.8 | 9.7 |
| Tail height, axil of pelvics | 5.4 | 5.0 | 5.7 | 5.0 | 5.6 | 5.1 | 5.7 | 5.9 | 6.1 |
| Tail width, base of sting | 5.2 | 4.1 | 5.2 | 4.2 | 5.2 | 4.1 | 4.9 | 3.7 | 5.5 |
| Tail height, base of sting | 3.9 | 3.2 | 4.2 | 3.2 | 3.7 | 3.2 | 4.2 | 3.4 | 4.2 |
| Cloaca length | 5.4 | 4.8 | 8.9 | 4.8 | 8.9 | 5.9 | 7.0 | 5.5 | 6.6 |
| Clasper, postcloaca length | - | 18.4 | 20.0 | 18.4 | 20.0 | 0.0 | 0.0 | 17.6 | 19.5 |
| Clasper, length from pelvic axil | - | 7.9 | 10.1 | 7.9 | 10.1 | 0.0 | 0.0 | 8.6 | 9.8 |
| Pect. insertion to sting origin | 26.3 | 25.4 | 29.5 | 25.4 | 28.7 | 25.9 | 29.5 | 22.9 | 26.0 |
| Cloaca origin to sting | 28.8 | 29.2 | 36.1 | 29.6 | 36.1 | 29.2 | 35.7 | 29.1 | 32.4 |
| Caudal sting 1 length | - | 17.4 | 31.9 | 17.4 | 25.8 | 21.0 | 31.9 | 21.2 | 22.0 |
| Caudal sting 2 length | - | 22.9 | 34.5 | 24.6 | 30.4 | 22.9 | 34.5 | 31.7 | 32.7 |
Table 2.
Morphometric data for Brevitrygon walga: newly designated lectotype (MNHN 2431), putative syntype of Trygon walga Müller & Henle (BMNH 1889.2.1.4196), and other material from the Bay of Bengal. Measurements expressed as a percentage of disc width (mm).
Table 2.
Morphometric data for Brevitrygon walga: newly designated lectotype (MNHN 2431), putative syntype of Trygon walga Müller & Henle (BMNH 1889.2.1.4196), and other material from the Bay of Bengal. Measurements expressed as a percentage of disc width (mm).
| | MNHN 2431 | BMNH 1889.2.1.4196 | Other Material (n = 6) |
|---|
| | Lectotype | Syntype T. walga? | Bay of Bengal |
|---|
| | Female | Female | Min | Max |
|---|
| Disc, width (mm) | 170 | 223 | 141 | 233 |
| Total length | 231.2 | 215.8 | 211.4 | 270.8 |
| Disc, length (direct) | 111.5 | 106.0 | 100.6 | 111.7 |
| Disc, thickness | 8.9 | 11.2 | 9.4 | 13.3 |
| Snout to origin of cloaca | 94.2 | 90.5 | 89.5 | 99.7 |
| Cloaca origin to tail tip | 137.1 | 125.3 | 118.1 | 181.3 |
| Snout to pectoral insertion | 101.0 | 97.2 | 95.1 | 103.1 |
| Snout to maximum width | 52.0 | 45.9 | 44.2 | 48.8 |
| End of orbit to pectoral insertion | 62.1 | 61.1 | 56.3 | 108.4 |
| Snout, preorbital (direct) | 33.7 | 31.1 | 31.0 | 33.5 |
| Snout, preorbital (horizontal) | 33.2 | 27.3 | 30.3 | 32.9 |
| Orbit diameter | 6.1 | 5.3 | 5.3 | 5.9 |
| Eye diameter | 4.2 | 3.5 | 3.5 | 3.9 |
| Spiracle length | 7.2 | 6.4 | 5.5 | 6.5 |
| Orbit and spiracle length | 10.9 | 9.8 | 9.2 | 10.4 |
| Interorbital width | 11.4 | 12.6 | 11.4 | 14.0 |
| Intereye width | 16.5 | 15.5 | 15.6 | 16.4 |
| Distance between spiracles | 16.6 | 16.9 | 15.8 | 17.7 |
| Head length (direct) | 61.0 | 57.2 | 57.1 | 59.2 |
| Snout, prenasal (direct) | 30.2 | 25.8 | 26.0 | 28.6 |
| Nostril length | 3.9 | 3.7 | 4.1 | 5.1 |
| Nasal curtain, length | 6.6 | 5.3 | 5.8 | 7.5 |
| Nasal curtain, width | - | 11.1 | 11.7 | 13.1 |
| Distance between nostrils | - | 11.1 | 11.4 | 12.3 |
| Snout, preoral (direct) | - | 31.4 | 31.9 | 34.5 |
| Mouth width | - | 9.4 | 9.3 | 11.0 |
| Width, 1st gill slit | 2.5 | 3.3 | 2.5 | 3.2 |
| Width, 3rd gill slit | 2.4 | 3.5 | 2.4 | 3.3 |
| Width, 5th gill slit | 1.3 | 2.5 | 2.0 | 2.6 |
| Distance between 1st gill slits | 25.4 | 24.9 | 22.4 | 24.4 |
| Distance between 5th gill slits | 14.5 | 15.6 | 13.0 | 15.9 |
| Length pelvic fin | 21.7 | 20.3 | 17.9 | 21.9 |
| Width across pelvic fin base | 17.6 | 17.0 | 14.4 | 21.5 |
| Greatest width across pelvic fins | 39.1 | 13.2 | 32.3 | 40.8 |
| Tail width, axil of pelvics | 10.0 | 8.6 | 6.1 | 10.4 |
| Tail height, axil of pelvics | 7.0 | 5.9 | 5.6 | 7.2 |
| Tail width, base of sting | 5.3 | 4.1 | 4.2 | 6.1 |
| Tail height, base of sting | 3.9 | 3.2 | 3.4 | 4.0 |
| Cloaca length | 5.1 | 6.0 | 4.8 | 7.7 |
| Clasper, postcloaca length | - | - | 12.1 | 16.8 |
| Clasper, length from pelvic axil | - | - | 4.3 | 8.0 |
| Pect. insertion to sting origin | 31.9 | 34.6 | 25.2 | 35.2 |
| Cloaca origin to sting | 36.1 | 37.6 | 32.9 | 39.1 |
| Caudal sting 1 length | - | - | 21.0 | 28.0 |
| Caudal sting 2 length | - | 8.2 | 16.0 | 37.9 |
Table 3.
Morphometric data for
Brevitrygon heterura: holotype (BMNH 1867.11.28.158), and non-types from Indonesia, Malaysia, Thailand, and Vietnam. Measurements expressed as percentages of disc width (mm). Data for non-types based on new and selective remeasurements of material from Manjaji [
3].
Table 3.
Morphometric data for
Brevitrygon heterura: holotype (BMNH 1867.11.28.158), and non-types from Indonesia, Malaysia, Thailand, and Vietnam. Measurements expressed as percentages of disc width (mm). Data for non-types based on new and selective remeasurements of material from Manjaji [
3].
| | Holotype | Non-Types |
|---|
| | BMNH 1867.11.28.158 | Indonesia (n = 4) | Malaysia (n = 21) | Thailand (n = 16) | Vietnam (n = 1) |
|---|
| | Female, Java, Indonesia | Min | Max | Min | Max | Min | Max | |
|---|
| Disc, width (mm) | 199 | 163 | 244 | 101 | 210 | 161 | 210 | 176 |
| Total length | 213.6 | 195.6 | 237.6 | 184.9 | 255.1 | 156.3 | 231.6 | - |
| Disc, length (direct) | 110.1 | 105.5 | 112.7 | 103.3 | 109.8 | 103.4 | 110.6 | 107.4 |
| Disc, thickness | 10.2 | 11.0 | 13.6 | 10.6 | 13.4 | 10.6 | 13.5 | 12.3 |
| Snout to origin of cloaca | 91.5 | 88.7 | 97.9 | 85.0 | 92.9 | 85.6 | 95.6 | 91.4 |
| Cloaca origin to tail tip | 122.1 | 100.4 | 142.3 | 93.1 | 168.3 | 64.3 | 142.7 | - |
| Snout to pectoral insertion | 98.9 | 95.2 | 104.7 | 92.7 | 100.8 | 93.5 | 102.1 | 97.4 |
| Snout to maximum width | 50.3 | 47.1 | 52.0 | 39.8 | 53.4 | 43.5 | 49.3 | 46.5 |
| End of orbit to pectoral insertion | 63.2 | 61.7 | 67.2 | 56.9 | 65.7 | 59.5 | 65.0 | 62.9 |
| Snout, preorbital (direct) | 31.7 | 29.6 | 32.4 | 28.3 | 32.1 | 27.6 | 31.1 | 30.0 |
| Snout, preorbital (horizontal) | 30.6 | 28.6 | 31.5 | 27.0 | 30.7 | 25.6 | 29.5 | 27.0 |
| Orbit diameter | 6.6 | 5.3 | 6.7 | 5.8 | 7.7 | 5.8 | 7.9 | 6.5 |
| Eye diameter | 4.4 | 3.3 | 4.8 | 3.7 | 5.2 | 3.8 | 4.6 | 4.1 |
| Spiracle length | 5.5 | 5.2 | 7.0 | 5.2 | 6.9 | 4.9 | 6.2 | 5.7 |
| Orbit and spiracle length | 10.1 | 9.7 | 10.6 | 9.4 | 11.8 | 9.2 | 11.0 | 10.7 |
| Interorbital width | 12.4 | 10.5 | 13.4 | 10.3 | 13.0 | 10.0 | 12.3 | 12.5 |
| Intereye width | 15.3 | 15.8 | 16.9 | 16.1 | 18.6 | 15.7 | 17.2 | 16.9 |
| Distance between spiracles | 16.7 | 16.6 | 18.0 | 15.4 | 18.0 | 15.5 | 17.4 | 18.8 |
| Head length (direct) | 57.6 | 55.9 | 59.3 | 52.3 | 59.3 | 54.6 | 58.8 | 58.2 |
| Snout, prenasal (direct) | 26.4 | 23.9 | 27.2 | 23.5 | 26.7 | 22.7 | 26.6 | 22.7 |
| Nostril length | 4.8 | 4.2 | 5.3 | 3.6 | 5.2 | 3.8 | 5.2 | 3.6 |
| Nasal curtain, length | 5.5 | 5.6 | 7.6 | 5.1 | 7.1 | 6.3 | 7.8 | 6.9 |
| Nasal curtain, width | 11.6 | 11.7 | 13.8 | 10.3 | 17.9 | 11.0 | 14.5 | 13.4 |
| Distance between nostrils | 11.7 | 11.1 | 13.2 | 10.4 | 12.9 | 10.8 | 13.7 | 12.6 |
| Snout, preoral (direct) | 33.0 | 29.5 | 33.4 | 28.5 | 33.1 | 28.2 | 32.8 | 29.7 |
| Mouth width | 9.8 | 9.3 | 10.4 | 7.9 | 9.9 | 8.3 | 10.2 | 10.6 |
| Width, 1st gill slit | 3.1 | 2.7 | 3.7 | 2.6 | 3.4 | 2.6 | 3.7 | 3.0 |
| Width, 3rd gill slit | 3.5 | 2.7 | 4.1 | 2.9 | 3.8 | 3.1 | 3.9 | 3.5 |
| Width, 5th gill slit | 1.9 | 2.0 | 2.6 | 1.4 | 2.5 | 1.9 | 2.5 | 2.0 |
| Distance between 1st gill slits | 23.7 | 22.7 | 25.1 | 21.0 | 25.1 | 22.3 | 24.4 | 25.2 |
| Distance between 5th gill slits | 15.3 | 14.9 | 15.7 | 14.1 | 16.5 | 14.6 | 16.6 | 16.4 |
| Length pelvic fin | 26.8 | 21.2 | 25.1 | 20.0 | 24.1 | 18.6 | 24.5 | 23.7 |
| Width across pelvic fin base | 17.1 | 16.1 | 16.7 | 12.9 | 18.7 | 13.8 | 17.8 | 15.2 |
| Greatest width across pelvic fins | - | 35.6 | 40.4 | 32.1 | 45.8 | 33.5 | 42.5 | 40.3 |
| Tail width, axil of pelvics | 7.8 | 8.4 | 10.0 | 8.1 | 11.2 | 7.4 | 11.2 | 9.7 |
| Tail height, axil of pelvics | 6.6 | 5.4 | 5.9 | 5.0 | 6.9 | 5.3 | 6.6 | 6.0 |
| Tail width, base of sting | 5.0 | 5.7 | 6.2 | 3.7 | 6.4 | 4.2 | 5.9 | - |
| Tail height, base of sting | 4.1 | 4.0 | 4.1 | 2.9 | 4.0 | 3.3 | 4.2 | - |
| Cloaca length | 5.7 | 5.0 | 8.3 | 4.0 | 7.3 | 4.5 | 7.2 | 5.9 |
| Clasper, postcloaca length | - | 18.7 | 20.4 | 12.0 | 22.6 | 20.6 | 22.8 | 19.2 |
| Clasper, length from pelvic axil | - | 9.2 | 13.3 | 7.5 | 20.3 | 9.3 | 17.1 | 13.6 |
| Pect. insertion to sting origin | 28.7 | 29.1 | 29.6 | 23.7 | 32.1 | 20.2 | 27.0 | - |
| Cloaca origin to sting | 34.0 | 32.5 | 33.6 | 29.4 | 37.1 | 26.8 | 31.3 | - |
| Caudal sting 1 length | - | - | - | - | - | - | - | - |
| Caudal sting 2 length | - | - | - | - | - | - | - | - |
Table 4.
Morphometric data for Brevitrygon heterura: males and females from Malaysia and Thailand, respectively. Measurements expressed as percentages of disc width (mm) and given as ranges with means.
Table 4.
Morphometric data for Brevitrygon heterura: males and females from Malaysia and Thailand, respectively. Measurements expressed as percentages of disc width (mm) and given as ranges with means.
| | Malaysia (n = 11) | Malaysia (n = 10) | Thailand (n = 7) | Thailand (n = 10) |
|---|
| | Adult Males | Females | Adult Males | Females |
|---|
| | Min | Max | Mean | Min | Max | Mean | Min | Max | Mean | Min | Max | Mean |
|---|
| Disc, width (mm) | 107 | 200 | 173 | 136 | 210 | 184 | 170 | 207 | 183 | 161 | 199 | 180 |
| Total length | 227.9 | 255.1 | 239.0 | 185.5 | 215.6 | 202.6 | 215.3 | 231.6 | 224.1 | 156.3 | 205.3 | 182.7 |
| Disc, length (direct) | 103.7 | 108.0 | 105.3 | 104.3 | 109.8 | 107.0 | 103.4 | 107.6 | 105.0 | 105.3 | 110.6 | 108.3 |
| Disc, thickness | 11.2 | 13.4 | 12.6 | 10.6 | 13.2 | 12.1 | 11.8 | 13.5 | 12.7 | 10.9 | 13.1 | 12.4 |
| Snout to origin of cloaca | 86.8 | 92.3 | 88.8 | 87.8 | 92.9 | 90.2 | 85.6 | 93.2 | 88.8 | 88.9 | 95.6 | 92.2 |
| Cloaca origin to tail tip | 139.5 | 168.3 | 150.6 | 93.1 | 127.8 | 112.7 | 122.4 | 142.7 | 134.2 | 64.3 | 116.1 | 90.4 |
| Snout to pectoral insertion | 92.9 | 98.3 | 94.8 | 93.4 | 100.8 | 97.0 | 93.5 | 98.2 | 95.4 | 95.0 | 102.1 | 98.8 |
| Snout to maximum width | 45.4 | 53.4 | 48.7 | 43.0 | 50.6 | 46.5 | 43.5 | 49.1 | 46.3 | 46.5 | 49.3 | 48.0 |
| End of orbit to pectoral insertion | 57.0 | 62.0 | 59.4 | 57.3 | 65.7 | 61.6 | 59.5 | 61.4 | 60.7 | 60.0 | 65.0 | 62.8 |
| Snout, preorbital (direct) | 29.5 | 32.0 | 30.8 | 28.3 | 32.1 | 30.5 | 27.6 | 30.1 | 29.0 | 28.8 | 31.1 | 29.7 |
| Snout, preorbital (horizontal) | 28.2 | 30.6 | 29.4 | 27.0 | 30.7 | 29.4 | 26.9 | 29.5 | 28.1 | 25.6 | 29.0 | 27.8 |
| Orbit diameter | 5.9 | 7.7 | 6.4 | 5.8 | 6.6 | 6.1 | 6.2 | 7.3 | 6.7 | 6.3 | 7.9 | 7.1 |
| Eye diameter | 3.9 | 5.2 | 4.2 | 3.8 | 4.5 | 4.2 | 4.1 | 4.5 | 4.1 | 3.8 | 4.6 | 4.3 |
| Spiracle length | 5.5 | 6.9 | 6.3 | 5.2 | 5.8 | 5.5 | 4.9 | 6.1 | 5.6 | 5.5 | 6.2 | 5.9 |
| Orbit and spiracle length | 10.0 | 11.8 | 10.5 | 9.4 | 10.2 | 9.8 | 10.0 | 11.0 | 10.5 | 9.2 | 10.9 | 10.3 |
| Interorbital width | 11.9 | 13.0 | 12.5 | 10.9 | 11.6 | 11.3 | 10.3 | 12.3 | 11.1 | 10.0 | 11.6 | 10.9 |
| Intereye width | 16.3 | 18.6 | 17.5 | 16.1 | 17.5 | 16.6 | 16.2 | 17.2 | 16.7 | 15.7 | 16.8 | 16.4 |
| Distance between spiracles | 16.7 | 18.0 | 17.1 | 15.4 | 16.8 | 16.1 | 16.7 | 17.4 | 17.0 | 15.5 | 16.4 | 16.0 |
| Head length (direct) | 56.4 | 59.3 | 57.6 | 52.3 | 57.6 | 56.0 | 54.6 | 58.8 | 56.6 | 55.1 | 57.5 | 56.3 |
| Snout, prenasal (direct) | 23.5 | 26.6 | 25.0 | 24.2 | 26.6 | 25.4 | 22.7 | 25.0 | 23.5 | 23.4 | 26.6 | 24.9 |
| Nostril length | 3.6 | 5.2 | 4.3 | 4.2 | 4.9 | 4.5 | 3.8 | 4.9 | 4.5 | 4.4 | 5.1 | 4.7 |
| Nasal curtain, length | 6.4 | 7.1 | 6.8 | 5.4 | 6.9 | 6.2 | 6.7 | 7.8 | 7.4 | 6.3 | 7.1 | 6.8 |
| Nasal curtain, width | 11.2 | 14.1 | 12.7 | 10.9 | 11.7 | 11.3 | 12.9 | 14.5 | 13.2 | 11.0 | 12.6 | 11.7 |
| Distance between nostrils | 11.3 | 12.9 | 12.0 | 10.5 | 11.1 | 10.8 | 12.2 | 13.7 | 12.6 | 10.8 | 11.4 | 11.0 |
| Snout, preoral (direct) | 29.4 | 32.4 | 31.1 | 29.1 | 33.0 | 31.5 | 28.2 | 31.6 | 29.7 | 29.8 | 32.8 | 31.0 |
| Mouth width | 8.8 | 9.9 | 9.5 | 8.3 | 8.8 | 8.5 | 9.0 | 10.2 | 9.7 | 8.3 | 10.2 | 9.0 |
| Width, 1st gill slit | 2.7 | 3.3 | 2.9 | 2.6 | 3.4 | 3.0 | 2.7 | 3.7 | 3.0 | 2.6 | 3.2 | 2.9 |
| Width, 3rd gill slit | 2.9 | 3.7 | 3.4 | 2.9 | 3.6 | 3.2 | 3.1 | 3.9 | 3.4 | 3.1 | 3.7 | 3.3 |
| Width, 5th gill slit | 1.7 | 2.3 | 1.9 | 1.6 | 2.5 | 2.1 | 1.9 | 2.2 | 2.1 | 1.9 | 2.5 | 2.1 |
| Distance between 1st gill slits | 22.3 | 25.1 | 23.2 | 21.3 | 22.4 | 21.9 | 23.0 | 24.4 | 23.8 | 22.3 | 24.4 | 23.3 |
| Distance between 5th gill slits | 15.1 | 16.5 | 15.7 | 14.1 | 15.3 | 14.8 | 14.6 | 16.5 | 15.6 | 14.6 | 16.6 | 15.4 |
| Length pelvic fin | 20.7 | 23.4 | 22.3 | 20.0 | 23.3 | 21.2 | 22.0 | 24.5 | 23.3 | 18.6 | 24.0 | 22.4 |
| Width across pelvic fin base | 12.9 | 15.2 | 13.8 | 13.7 | 18.7 | 16.6 | 13.8 | 15.5 | 14.8 | 15.4 | 17.8 | 16.9 |
| Greatest width across pelvic fins | - | - | - | - | - | - | 40.4 | 42.5 | 41.6 | 34.3 | 42.1 | 37.8 |
| Tail width, axil of pelvics | 8.1 | 10.4 | 9.6 | 10.0 | 11.2 | 10.7 | 7.4 | 9.1 | 8.8 | 9.2 | 11.2 | 10.0 |
| Tail height, axil of pelvics | 5.4 | 6.5 | 5.8 | 5.2 | 6.5 | 5.9 | 5.3 | 6.1 | 5.6 | 5.4 | 6.6 | 5.9 |
| Tail width, base of sting | 3.7 | 6.4 | 5.2 | 4.1 | 5.4 | 4.8 | 4.2 | 5.6 | 5.3 | 5.1 | 5.9 | 5.5 |
| Tail height, base of sting | 3.0 | 4.0 | 3.5 | 2.9 | 3.6 | 3.3 | 3.3 | 3.9 | 3.8 | 3.5 | 4.2 | 3.8 |
| Cloaca length | 4.0 | 5.6 | 4.9 | 4.5 | 7.3 | 5.8 | 4.5 | 5.3 | 4.9 | 4.9 | 7.2 | 5.9 |
| Clasper, postcloaca length | 13.0 | 22.6 | 20.2 | - | - | - | 20.6 | 22.8 | 21.3 | - | - | - |
| Clasper, length from pelvic axil | 7.5 | 20.3 | 15.1 | - | - | - | 9.3 | 17.1 | 15.7 | - | - | - |
| Pect. insertion to sting origin | 25.1 | 32.1 | 28.1 | 24.8 | 30.9 | 27.8 | 20.2 | 26.7 | 23.4 | 20.7 | 27.0 | 23.7 |
| Cloaca origin to sting | 29.4 | 37.1 | 32.2 | 29.7 | 35.6 | 33.4 | 26.8 | 31.3 | 28.7 | 28.8 | 31.3 | 30.1 |
| Caudal sting 1 length | - | - | - | - | - | - | - | - | - | - | - | - |
| Caudal sting 2 length | - | - | - | - | - | - | - | - | - | - | - | - |
Table 5.
Morphometric data for all types of
Brevitrygon javaensis based on Last & White [
17]. Measurements expressed as percentages of disc width (mm).
Table 5.
Morphometric data for all types of
Brevitrygon javaensis based on Last & White [
17]. Measurements expressed as percentages of disc width (mm).
| | Min | Max |
|---|
| Disc, width (mm) | 164 | 234 |
| Total length | 296.3 | 309.2 |
| Disc, length (direct) | 99.8 | 105.5 |
| Disc, thickness | 11.0 | 13.5 |
| Snout to origin of cloaca | 83.8 | 89.5 |
| Cloaca origin to tail tip | 209.8 | 220.7 |
| Snout to pectoral insertion | 90.9 | 96.6 |
| Snout to maximum width | 46.8 | 49.0 |
| End of orbit to pectoral insertion | 57.2 | 61.5 |
| Snout, preorbital (direct) | 28.3 | 30.2 |
| Snout, preorbital (horizontal) | 26.2 | 29.1 |
| Orbit diameter | 5.1 | 6.0 |
| Eye diameter | 3.2 | 3.8 |
| Spiracle length | 5.6 | 6.6 |
| Orbit and spiracle length | 9.3 | 9.9 |
| Interorbital width | 11.4 | 12.9 |
| Intereye width | 15.1 | 16.7 |
| Distance between spiracles | 17.0 | 18.4 |
| Head length (direct) | 52.1 | 55.5 |
| Snout, prenasal (direct) | 24.1 | 26.0 |
| Nostril length | 3.3 | 3.9 |
| Nasal curtain, length | 10.0 | 11.9 |
| Nasal curtain, width | 4.8 | 5.7 |
| Distance between nostrils | 9.0 | 10.3 |
| Snout, preoral (direct) | 28.6 | 30.7 |
| Mouth width | 7.5 | 9.1 |
| Width, 1st gill slit | 2.8 | 3.4 |
| Width, 3rd gill slit | 2.8 | 3.3 |
| Width, 5th gill slit | 2.0 | 2.8 |
| Distance between 1st gill slits | 20.5 | 22.3 |
| Distance between 5th gill slits | 13.2 | 14.3 |
| Length pelvic fin | 17.3 | 19.7 |
| Width across pelvic fin base | 15.1 | 17.9 |
| Greatest width across pelvic fins | 33.7 | 42.2 |
| Tail width, axil of pelvics | 8.3 | 10.0 |
| Tail height, axil of pelvics | 5.2 | 6.7 |
| Tail width, base of sting | 3.8 | 4.6 |
| Tail height, base of sting | 2.9 | 3.2 |
| Cloaca length | 5.5 | 7.3 |
| Clasper, postcloaca length | 18.4 | 18.9 |
| Clasper, length from pelvic axil | 8.1 | 8.7 |
| Pect. insertion to sting origin | 33.9 | 39.1 |
| Cloaca origin to sting | 38.1 | 42.1 |
| Caudal sting 1 length | - | - |
| Caudal sting 2 length | - | - |
Table 6.
Morphometric data for holotype and 5 paratypes of Brevitrygon manjajiae sp. nov., and two other non-type specimens from Kuwait. Measurements expressed as percentages of disc width (mm).
Table 6.
Morphometric data for holotype and 5 paratypes of Brevitrygon manjajiae sp. nov., and two other non-type specimens from Kuwait. Measurements expressed as percentages of disc width (mm).
| | Pakistan | Kuwait |
|---|
| | Holotype | Paratypes | Non-Types |
|---|
| | USNM 222555 | Min | Max | BPBM 33199 | MCZ 59269 (1 of 5) |
|---|
| Disc, width (mm) | 231 | 174 | 226 | 187 | 167 |
| Total length | 224.2 | 220.2 | 239.0 | 191.8 | 238.7 |
| Disc, length (direct) | 103.2 | 101.3 | 102.9 | 109.1 | 107.5 |
| Disc, thickness | 12.6 | 10.6 | 13.2 | 10.9 | 10.1 |
| Snout to origin of cloaca | 87.6 | 84.5 | 87.2 | 92.3 | 93.2 |
| Cloaca origin to tail tip | 136.6 | 134.1 | 152.2 | 98.6 | 145.4 |
| Snout to pectoral insertion | 94.0 | 88.7 | 93.7 | 100.2 | 100.1 |
| Snout to maximum width | 47.0 | 42.9 | 47.4 | 46.2 | 50.7 |
| End of orbit to pectoral insertion | 60.5 | 56.3 | 59.6 | 62.6 | 60.1 |
| Snout, preorbital (direct) | 28.5 | 28.0 | 30.0 | 31.1 | 34.2 |
| Snout, preorbital (horizontal) | 27.5 | 27.2 | 28.4 | 29.2 | 33.8 |
| Orbit diameter | 6.2 | 5.3 | 6.3 | 6.5 | 6.8 |
| Eye diameter | 4.4 | 3.8 | 4.5 | 4.4 | 4.4 |
| Spiracle length | 5.3 | 5.0 | 6.7 | 5.8 | 6.4 |
| Orbit and spiracle length | 10.0 | 9.1 | 10.5 | 11.4 | 11.6 |
| Interorbital width | 13.0 | 12.0 | 13.8 | 12.5 | - |
| Intereye width | 16.9 | 15.1 | 16.8 | 16.6 | - |
| Distance between spiracles | 16.4 | 16.2 | 17.7 | 16.6 | - |
| Head length (direct) | 55.8 | 53.9 | 56.1 | 57.0 | 59.7 |
| Snout, prenasal (direct) | 22.5 | 21.0 | 24.6 | 26.3 | 28.0 |
| Nostril length | 4.6 | 3.6 | 4.6 | 4.7 | 5.0 |
| Nasal curtain, length | 7.3 | 5.6 | 6.7 | 7.1 | 7.0 |
| Nasal curtain, width | 12.5 | 10.8 | 13.3 | 11.6 | 11.6 |
| Distance between nostrils | 11.7 | 11.1 | 12.4 | 10.6 | 11.5 |
| Snout, preoral (direct) | 28.2 | 26.5 | 30.0 | 32.1 | 34.2 |
| Mouth width | 10.0 | 8.8 | 10.4 | 9.8 | - |
| Width, 1st gill slit | 3.1 | 2.7 | 3.6 | 3.7 | 3.3 |
| Width, 3rd gill slit | 3.4 | 3.3 | 3.9 | 3.5 | 3.1 |
| Width, 5th gill slit | 2.6 | 2.1 | 2.8 | 2.6 | 2.5 |
| Distance between 1st gill slits | 24.1 | 21.5 | 23.1 | 21.9 | 22.1 |
| Distance between 5th gill slits | 16.0 | 13.6 | 15.6 | 14.9 | 15.0 |
| Length pelvic fin | 19.3 | 17.1 | 19.3 | 20.2 | 18.1 |
| Width across pelvic fin base | 16.3 | 15.9 | 18.3 | 18.0 | 14.9 |
| Greatest width across pelvic fins | 36.5 | 29.8 | 38.7 | 35.1 | 36.6 |
| Tail width, axil of pelvics | 8.3 | 8.1 | 10.4 | 9.4 | 9.8 |
| Tail height, axil of pelvics | 5.6 | 5.4 | 6.1 | 6.4 | 5.8 |
| Tail width, base of sting | 4.0 | 4.2 | 4.6 | 4.3 | 4.8 |
| Tail height, base of sting | 3.2 | 3.4 | 3.7 | 3.2 | 3.8 |
| Cloaca length | 5.7 | 4.6 | 7.0 | 7.2 | 5.9 |
| Clasper, postcloaca length | 18.5 | 11.7 | 17.1 | - | 17.9 |
| Clasper, length from pelvic axil | 7.2 | 5.5 | 7.4 | - | 8.2 |
| Pect. insertion to sting origin | 30.7 | 28.7 | 35.9 | 31.2 | 30.1 |
| Cloaca origin to sting | 35.8 | 31.5 | 40.2 | 37.4 | 36.0 |
| Caudal sting 1 length | 17.4 | 6.9 | 21.0 | 18.2 | 21.6 |
| Caudal sting 2 length | 24.6 | 23.0 | 23.5 | - | 29.6 |











































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}