

The Terebrantia (Insecta: Thysanoptera) of the Maltese Islands

1
Institute of Earth Systems, Rural Sciences and Food Systems, University of Malta, MSD 2080 Msida, Malta
2
Biological Sciences Institute, Carreiros Campus, Federal University of Rio Grande, Rio Grande 96203-900, Brazil
3
National Museum of Natural Sciences-CSIC, Department of Biodiversity and Evolutionary Biology National Reference Laboratory for Arthropods, C/Serrano Duplicado, CP 28006 Madrid, Spain
*
Author to whom correspondence should be addressed.
Diversity 2023, 15(4), 514; https://doi.org/10.3390/d15040514
Submission received: 29 January 2023
/
Revised: 8 March 2023
/
Accepted: 14 March 2023
/
Published: 3 April 2023
(This article belongs to the Special Issue Diversity, Taxonomy and Evolution of Insects)
Abstract
:Thirty-nine species of terebrantian Thysanoptera belonging to four families and 22 genera are here recorded from the Maltese Islands. Of these, 33 represent new records to this archipelago. Thrips were collected from 65 different locations over a seven-year period, covering the main habitat types found across the Maltese Islands, namely steppe, garigue, maquis and woodland, but also sand dunes and saltmarshes as well as roadsides, private and public gardens, greenhouses and cultivated fields. An illustrated dichotomous key to identify the Terebrantia of the Maltese Islands is presented. Chorological data for the species researched in the current study shows that the majority of these insects are of a European Mediterranean origin, though the geographical distribution of some of them extends to Africa and the Middle East. Seven species associated with agricultural commodities were found to be of alien origin; however they were locally found in small numbers and do not pose a threat to horticulture.
1. Introduction
Thrips belong to the insect order Thysanoptera which comprises more than 6400 described species worldwide. These insects are found on plants, leaf litter or dead twigs where they feed on pollen, plant sap and fungi. The order is divided into two suborders: Terebrantia and Tubulifera, which are physiologically and behaviorally different. Around 100 thrips species are reported to impact crop productivity around the world, almost all belonging to Terebrantia [1].
Generally, thrips are associated with plants, but these plants are only considered as host-plants if the insect completes its life cycle on the mentioned plant [2]. Thus, the establishment of whether a plant is a true host of a particular thrips relies on the presence of immature stages. Although a small number of thrips species do not stray from their host-plant, the majority, as adults, tend to use other plants as a source of food or shelter, but these should never be regarded as host plants.
The Maltese archipelago consists of the habitable islands of Malta and Gozo and a number of much smaller islands including Comino, Selmunett and Filfla. The archipelago is situated in the center of the Mediterranean basin and covers an area of 316 km2 [3]. The climate of the archipelago alternates between a mild rainy cold season and a dry hot summer [4] lasting from early June to the end of September, with temperatures averaging about 30 °C, though sometimes peaking at 40 °C. Water availability is rather scarce, with typical average rainfall for the period 1900–2000 being 550 mm [5]. Abstraction of groundwater, necessary to sustain the relatively limited perennial surface water ecosystems, exceeds aquifer recharge rate [5].
The Maltese Islands lack many habitats such as mountains, rivers, large forests, alpine environments and others and thus insects associated with these habitat types are lacking. Nonetheless a range of natural terrestrial habitats still occur, which can be categorized into four main types: the steppe, the garigue, the maquis and the woodland [6]. A number of valleys with temporary fresh water courses as well as a few degraded sand dunes and saltmarshes can also be found. All of these habitats can potentially host thrips on the vegetation therein. Despite the limited variety of habitats, the biodiversity of the flora of the Maltese Islands is rather rich, with some 1100 indigenous plants [7] and a relatively large number of cultivated plants, some of which have become established escapees.
The literature on the Thysanoptera of the Maltese Islands is very fragmentary. Seven species of Terebrantia have been recorded. These include Thrips tabaci a widely known agricultural pest that affects onions, garlic and cauliflowers [8,9,10], Tenothrips discolor [11], Frankliniella occidentalis, Heliothrips haemorrhoidalis [10,12], Aeolothrips tenuicornis, Melanthrips fuscus [13] and Melanthrips lybicus [14,15].
The Maltese Islands are becoming urbanized at a rapid rate due to the high popula-tion density, scoring at 1649 per km2 [16]. This problem is compounded by the fact that the economy of the Maltese Islands heavily depends on tourism, with c. 1.5 million tourists visiting the islands every year. These factors leave a considerable negative impact on the Maltese natural environment. Extensive use of plant protection products is also causing the decline of a number of indigenous insect species, many of which serve as ecosystem providers. A knowledge of insect biodiversity is a prerequisite to enable proper conservation programs, thus avoiding loss of species diversity due to climate change, among other issues.
This present study was thus undertaken to document the biodiversity of the terebrantians that inhabit the Maltese Islands. It also provides an illustrated dichotomous key.
2. Materials and Methods
During the current study, thrips specimens were collected from 65 different locations from Malta and Gozo (as shown in Figure 1) between Spring 2015 and Spring 2022. The habitats and flora therein are described in Table 1. Thrips were collected from the main habitats found in the Maltese Islands as mentioned earlier, but also from private and public gardens, town and country roadsides, public green areas, private farms and cultivated open fields.
Thrips were collected from 398 different species of indigenous and cultivated plants. The literature on some 700 thrips species recorded mostly from the Palaearctic region was consulted, e.g., zur Strassen (2003) [14] in order to find out which plant species these thrips were collected from. If these plant species were present in the Maltese Islands, they were chosen for sampling purposes. Moreover, other plants which prevailed in the different habitats of the Maltese archipelago were also sampled. Plants were beaten using a plastic rod onto a white plastic tray [17]. All specimens collected in the tray were transferred into collecting Eppendorf tubes using a fine brush (e.g., size 00) dipped in AGA fluid mixture consisting of 10 parts of 60% ethyl alcohol, one part of glycerin and one part of glacial acetic acid. Additional specimens of thrips were collected from Malaise traps present in two locations in Malta (Fawwara and Buskett).
Four hundred and sixty of the collected specimens were mounted on microscope slides for examination under compound microscopy. Prior to mounting, thrips specimens were macerated by immersing in 5% sodium hydroxide. Dark specimens were immersed in this agent for about 12 h while lighter specimens were left for less time depending on the colour intensity of the specimen. The specimens were subsequently dehydrated by placing the specimens in increasing concentrations of ethanol (75, 80, 95, and 100%) [18]. The specimens were finally cleared by immersing in clove oil prior to mounting in Canada balsam [18], after which they were mounted by placing them on the cover slip and then placing the glass slide on top [18].
Mounted specimens were viewed using a Leica DVM6 microscope and identified with the use of a number of printed [14,17,19,20,21,22,23,24] and on-line sources [25,26,27,28] as well as published works having identification keys and species descriptions. Different morphological features observed in the species identified were used to construct the dichotomous key.
The majority of the specimens examined for the current study form part of the private collection of Godwin Degabriele (GD), though some 15 specimens come from the private collection of David Mifsud (DM), which were collected during the late 1990s to the mid 2000s. A number of species from the collection of GD were donated to Arturo Goldarazena (AG) who will deposit them in the collection of the the Museo Nacional de Ciencias Naturales, Madrid.
Abbreviations used in the following sections include the following: Godwin Degabriele (GD); Charles Farrugia (CF); Sylvan Farrugia (SF); David Mifsud (DM); E.R. Speyer (ERS); I.A. Speyer (IAS); Gillian Watson (GW); J.W. Ismay (JWI); Niki Young (NY); slide mounted specimens (sm); specimens conserved in AGA (aga); specimens housed in the British Museum of Natural History (BMNH); thrips which represent new record for the Maltese Islands (†). Representative species from the private collection of GD will eventually be donated to the BMNH.
3. Results
Thirty-nine species of Terebrantia belonging to four families and 22 genera were identified, with 33 of these being new records to the Maltese Islands. These species are discriminated by the illustrated dichotomous key provided below.
3.1. Key to the Terebrantia of the Maltese Islands
- 1
- Abdominal segment X conical in females (Figure 2a) and rounded posteriorly in males; fore wings with one or two longitudinal veins (Figure 2c); females with external ovipositor (Figure 3e,f) ………………………………………… Terebrantia .. 2
Figure 2.
Abdominal segment X (a,b): (a) conical; (b) tubular; fore wing (c,d): (c) with longitudinal veins; (d) with no veins.
Figure 2.
Abdominal segment X (a,b): (a) conical; (b) tubular; fore wing (c,d): (c) with longitudinal veins; (d) with no veins.

- 2
- Antennal segments III and IV with sensoria consisting of a ridge-like structure set in parallel (Figure 3a) or perpendicular (or oblique) to each segment (Figure 3b), never produced as trichomes; fore wing broad, being five to ten times as long as broad (Figure 3c); females with ovipositor curving upwards towards abdominal segments (Figure 3e) ………………………………………………………………………………….. 3Antennal segments III and IV with emergent sensoria, consisting of an acorn-shaped structure (Figure 3g), or a simple (Figure 3h) or forked trichome (Figure 3i); fore wing narrow, being 11–18 times as long as broad and often tapering apically (Figure 3d); females with ovipositor curving downwards (Figure 3f) ……………………………. 15
Figure 3.
Sensoria on antennal segments III and IV (a,b): (a) parallel to length of segment; (b) perpendicular to length of segment; fore wing tip (c,d): (c) broad; (d) narrows apically; ovipositor (e,f): (e) curving upwards towards body: (f) curving downwards; antennal sensoria on segments III and IV (g,i): (g) cone-shaped; (h) simple or hair-like; (i) forked.
Figure 3.
Sensoria on antennal segments III and IV (a,b): (a) parallel to length of segment; (b) perpendicular to length of segment; fore wing tip (c,d): (c) broad; (d) narrows apically; ovipositor (e,f): (e) curving upwards towards body: (f) curving downwards; antennal sensoria on segments III and IV (g,i): (g) cone-shaped; (h) simple or hair-like; (i) forked.

- 3
- Antennal segments III and IV with sensoria set in parallel to length of segment (Figure 3a); antennal segments VI–IX broadly joined basally (Figure 4a) Aeolothripidae .. 4
Figure 4.
Antennal segments VI–IX: (a) broadly joined basally; (b) distinct from each other.

- 4
- Antennal segment III about 15 times as long as wide (Figure 5a); antennal segments III and IV with sensoria consisting of a multitude of dot-like structures lined up parallel to length of segment (Figure 5b); fore wings with dark band sub-apically (Figure 5c); abdominal segments I–III significantly narrower than other segments, giving the appearance of a wasp-like waist (Figure 5e) ………………………………………………………………………………………….. Franklinothrips megalops TrybomAntennal segments III 3.3–5 times as long as wide; antennal segments III and IV with sensoria consisting of one or a small number of continuous ridge shapes lined up parallel to length of segment (Figure 5b); fore wings with light coloured sub-apical region (Figure 5d); abdominal segments I–III only slightly narrower than other segments (Figure 5f) ……………………………………………………………………….. 5
Figure 5.
(a) antennal segments III–IX; (b) sensorium on antennal segment III; fore wing tip (c,d): (c) with dark band sub-apically; (d) with light-coloured sub-apical region; upper abdominal segments (e,f): (e) being considerably narrower than other segments; (f) only slightly narrower than other segments.
Figure 5.
(a) antennal segments III–IX; (b) sensorium on antennal segment III; fore wing tip (c,d): (c) with dark band sub-apically; (d) with light-coloured sub-apical region; upper abdominal segments (e,f): (e) being considerably narrower than other segments; (f) only slightly narrower than other segments.

- 5
- Antennal segments III and IV with sensoria consisting of one continuous ridge-like structure lined up parallel to length of segment (Figure 6a); fore wings broad, being less than six times as long as broad and with two dark bands (Figure 6c); pronotum postero-marginal setae as long as discal setae ……………….. Aeolothrips Haliday .. 6Antennal segments III and IV with sensoria consisting of lens-shaped structures lined up parallel to length of segment (Figure 6b); fore wings narrow, being more than six times as long as broad and of a uniformly pale brown hue (Figure 6d); pronotum bearing a pair of postero-marginal setae that are longer than discal setae ……………… ……………………………………………………………………… Rhipidothrips Uzel .. 9
Figure 6.
Antennal sensoria on segment III (a,b): (a) continuous and ridge-shaped; (b) lens-shaped; fore wing (c,d): (c) broad and banded; (d) narrow and monochrome.
Figure 6.
Antennal sensoria on segment III (a,b): (a) continuous and ridge-shaped; (b) lens-shaped; fore wing (c,d): (c) broad and banded; (d) narrow and monochrome.

- 6
- Fore wing margin at tip as light as wing membrane (Figure 7a) …………………….. 7Fore wing margin at tip darker than wing membrane (Figure 7b) …………………... 8
Figure 7.
Fore wing: (a) pale coloured margin at tip; (b) dark coloured margin at tip.

- 7
- Body bicoloured, with head and thorax yellow with brown areas medially (Figure 8a); antennal segment III bicoloured, with the basal half of segment pale yellow, while the apical half dark brown (Figure 8c) …………………….. Aeolothrips gloriosus Bagnall
Figure 8.
Body (a,b): (a) bicoloured; (b) uniformly brown; antennal segment III (c,d): (c) distal half brown; (d) only distal fifth brown.
Figure 8.
Body (a,b): (a) bicoloured; (b) uniformly brown; antennal segment III (c,d): (c) distal half brown; (d) only distal fifth brown.

- 8
- Antennal segment III with sensorium being one third the length of the segment (Figure 9a); antennal segment IV with sensorium two thirds the length of the segment (Figure 9a); pronotum pale, often yellow (Figure 9c) …….. Aeolothrips melisi PriesnerAntennal segment III and IV with sensoria about half the length of the segment (Figure 9b); pronotum dark, usually brown (Figure 9d) ……………………………………………………………………………………………… Aeolothrips tenuicornis Bagnall
Figure 9.
Antennal segments III and IV (a,b): (a) sensoria about one third the length of segment in III, two thirds the length of segment in IV; (b) sensoria about half the length of segment in both III and IV; pronotum (c,d): (c) pale; (d) dark.
Figure 9.
Antennal segments III and IV (a,b): (a) sensoria about one third the length of segment in III, two thirds the length of segment in IV; (b) sensoria about half the length of segment in both III and IV; pronotum (c,d): (c) pale; (d) dark.

- 9
- Antennal segment II yellow (Figure 10c); pronotum yellow to light brown and with three to four long pairs of postero-marginal setae (Figure 10a); females always macropterous (Figure 10f); abdominal tergites with little or no sculpture or microtrichia (Figure 10g) ………………………………… Rhipidothrips gratiosus Uzel.
Figure 10.
Antennal segment II (a,b): (a) yellow; (b) brown; pronotum (c,d): (c) with four pairs of long postero-marginal setae; (d) with three pairs of postero-marginal setae; fore wings (e,f): (e) micropterous; (f) macropterous; abdominal tergites (g,h): (g) with no sculpture; (h) with sculpture.
Figure 10.
Antennal segment II (a,b): (a) yellow; (b) brown; pronotum (c,d): (c) with four pairs of long postero-marginal setae; (d) with three pairs of postero-marginal setae; fore wings (e,f): (e) micropterous; (f) macropterous; abdominal tergites (g,h): (g) with no sculpture; (h) with sculpture.

- 10
- Internal body colour orange brown (Figure 11a); females macropterous. On Stipa capensis …………………………………………… Rhipidothrips unicolor zur Strassen.
Figure 11.
Abdominal segments II and III showing internal body color: (a) brown; (b) red.

- 11
- Abdominal tergites with microtrichia on sculpture lines (Figure 12a); females usually macropterous ………………………………….. Rhipidothrips niveipennis O.M. ReuterAbdominal tergites with no microtrichia on sculpture (Figure 12b); females usually micropterous ………………………………………… Rhipidothrips brunneus Williams
Figure 12.
Section of abdominal tergites: (a) with microtrichia on sculpture; (b) with no microtrichia on sculpture.
Figure 12.
Section of abdominal tergites: (a) with microtrichia on sculpture; (b) with no microtrichia on sculpture.

- 12
- Fore wing banded (Figure 13a) …………………………….. Melanthrips ficalbii BuffaFore wing evenly brown, not banded (Figure 13b) and with a small pale sub-basal area ………………………………………………………………………………………… 13
Figure 13.
Fore wing: (a) banded; (b) not banded.

- 13
- Anterior margin of fore wing between transverse veins with one row of setae (Figure 14a) ……………………………………………………….. Melanthrips knechteli PriesnerAnterior margin of fore wing between transverse veins with two rows of setae (Figure 14b) ……………………………………………………………………………………….. 14
Figure 14.
Fore wing: (a) with anterior margin between transverse veins with one row of setae; (b) with anterior margin between transverse veins with two rows of setae.
Figure 14.
Fore wing: (a) with anterior margin between transverse veins with one row of setae; (b) with anterior margin between transverse veins with two rows of setae.

- 14
- Hind tibia with one long seta (Figure 15a) ……………… Melanthrips fuscus (Sulzer)Hind tibia with two long setae (Figure 15b) …………… Melanthrips lybicus Priesner
Figure 15.
Hind tibia: (a) with one long seta; (b) with two long setae.

- 15
- Antennae nine-segmented (Figure 16a); antennal segments III and IV with cone-shaped sensoria (Figure 3g) ………………………….Stenurothripidae ……………………………………………………..Holarthrothrips tenuicornis Priesner
Figure 16.
Antenna (a–d): (a) nine-segmented; (b) eight-segmented; (c) seven-segmented; (d) six-segmented.
Figure 16.
Antenna (a–d): (a) nine-segmented; (b) eight-segmented; (c) seven-segmented; (d) six-segmented.

- 16
- Metathoracic endofurca lyre-shaped, with arms extending well beyond length of segment (Figure 17a); median tergal setae on abdomen arising close to each other (Figure 17c) ………………………………………………………….. Dendrothripinae …………………………………………………………………. Dendrothrips saltator Uzel
Figure 17.
Pterothorax (a,b): (a) with lyre-shaped metathoracic endofurca; (b) with metathoracic endofurca not lyre-shaped; tergites (c,d): (c) median setae close to each other; (d) median setae far away from each other.
Figure 17.
Pterothorax (a,b): (a) with lyre-shaped metathoracic endofurca; (b) with metathoracic endofurca not lyre-shaped; tergites (c,d): (c) median setae close to each other; (d) median setae far away from each other.

- 17
- Fore wing with one row of postero-marginal cilia (Figure 18a); antennal segments VI–VIII long and slender when compared with basal segments. (Figure 18c) ………………………………… ………………………………… Panchaetothripinae .. 18
Figure 18.
Fore wing (a,b): (a) one row of postero-marginal cilia; (b) two rows of postero-marginal cilia; terminal antennal segments (c,d): (c) ending in a sharp point; (d) not ending in a sharp point.
Figure 18.
Fore wing (a,b): (a) one row of postero-marginal cilia; (b) two rows of postero-marginal cilia; terminal antennal segments (c,d): (c) ending in a sharp point; (d) not ending in a sharp point.

- 18
- Fore tarsi one-segmented (Figure 19a); fore wings uniformly light brown; postero-marginal cilia straight (Figure 19c); fore wing apex rounded (Figure 19c) …………………………………………………… Heliothrips haemorrhoidalis (Bouchè)
Figure 19.
Fore tarsus (a,b): (a) one-segmented; (b) two-segmented; fore wing (c,d): (c) postero-marginal cilia straight and apex rounded; (d) postero-marginal cilia wavy and apex pointed.
Figure 19.
Fore tarsus (a,b): (a) one-segmented; (b) two-segmented; fore wing (c,d): (c) postero-marginal cilia straight and apex rounded; (d) postero-marginal cilia wavy and apex pointed.

- 19
- Pronotum considerably narrower at anterior margin than at posterior margin (Figure 20a) ………………………………………………………………. Chirothrips Haliday .. 20Pronotum having the same width at both margins (Figure 20b) ……………………. 22
Figure 20.
Pronotum: (a) considerably narrower at anterior margin than at posterior margin; (b) being almost the same width at both margins.
Figure 20.
Pronotum: (a) considerably narrower at anterior margin than at posterior margin; (b) being almost the same width at both margins.

- 20
- Antennal segment II symmetrical ……………………….. Chirothrips hamatus TrybomAntennal segment II asymmetrical, projecting laterally (Figure 21a,b) ……………. 21
Figure 21.
Antennal segment II showing outer edge (a,b): (a) nearly straight; (b) at an obtuse angle from the base of the segment; antennal segment IV (c,d): (c) with a forked sense cone; (d) with a simple sense cone.
Figure 21.
Antennal segment II showing outer edge (a,b): (a) nearly straight; (b) at an obtuse angle from the base of the segment; antennal segment IV (c,d): (c) with a forked sense cone; (d) with a simple sense cone.

- 21
- Antennal segment II with the outer edge nearly straight (Figure 21a); antennal segment IV with forked sense cone (Figure 21c); antennal segment VII longer than VIII; males usually macropterous ………………….. Chirothrips meridionalis BagnallAntennal segment II with the outer edge at an obtuse angle from the base of segment, creating a tip with bulge (Figure 21b); antennal segment IV with simple sense cone (Figure 21d); antennal segment VII shorter than VIII; males micropterous …………………………………………………………… Chirothrips manicatus Haliday
- 22
- Abdominal segment X bearing a pair of thorn-shaped setae (Figure 22a) …………………………………………………………………… Limothrips Haliday .. 23Abdominal segment X without thorn-shaped setae (Figure 22b) …………………… 24
Figure 22.
Abdominal segment X: (a) with thorn-shaped setae; (b) lacking thorn-shaped setae.

- 23
- Antennal segment II asymmetrical, with elongated extremity at outer side of segment (Figure 23a); antennal segments III and IV with forked sense cones (Figure 23c) …………………………………………………….. Limothrips angulicornis Jablonowski
Figure 23.
Antennal segment II (a,b): (a) asymmetrical; (b) symmetrical; antennal segments III and IV (c,d): (c) with forked sense cones; (d) with simple sense cones.
Figure 23.
Antennal segment II (a,b): (a) asymmetrical; (b) symmetrical; antennal segments III and IV (c,d): (c) with forked sense cones; (d) with simple sense cones.

- 24
- Antennal segments III and IV with simple sense cones (Figure 3h) …………..…… 25Antennal segments III and IV with forked sense cones (Figure 3i) ………………… 28
- 25
- Antennae six-segmented (Figure 16d); head and body having a distinct bright yellow colour (Figure 24a) …………………………………………. Aptinothrips rufus Haliday
Figure 24.
Male habitus: (a) yellow body colour; (b) body bicoloured.

- 26
- Fore wing with setae on vein always long, dark and capitate (blunt tipped) (Figure 25b); second vein with no setae (Figure 25a); antennal segment VIII ending in a sharp point (Figure 18c); males macropterous ……………. Echinothrips americanus Morgan
Figure 25.
(a) Fore wing with second vein with no setae; (b) setae on vein of fore wing with capitate tips; (c) fore wing with second vein with setae.
Figure 25.
(a) Fore wing with second vein with no setae; (b) setae on vein of fore wing with capitate tips; (c) fore wing with second vein with setae.

- 27
- Females micropterous; head with reticulate (network-like) sculpture; ridge present between antennal sockets (Figure 26a) ……………… Prosopothrips nigriceps BagnallFemales macropterous; head with no distinct sculpture; ridge lacking between antennal sockets (Figure 26b) ………………… Bregmatothrips dimorphus (Priesner)
Figure 26.
Head: (a) with reticulate sculpture and with a ridge between antennal sockets; (b) with no distinct sculpture and with no ridge between antennal sockets.
Figure 26.
Head: (a) with reticulate sculpture and with a ridge between antennal sockets; (b) with no distinct sculpture and with no ridge between antennal sockets.

- 28
- Pronotum with no long postero-angular setae (Figure 27a); fore wing with dark band at proximal region (Figure 27c) …………………….. Anaphothrips sudanensis Trybom
Figure 27.
Pronotum (a,b): (a) with no long postero-angular setae; (b) with long postero-angular setae present; fore wing (c,d): (c) with a dark band at proximal region; (d) with no dark band at proximal region.
Figure 27.
Pronotum (a,b): (a) with no long postero-angular setae; (b) with long postero-angular setae present; fore wing (c,d): (c) with a dark band at proximal region; (d) with no dark band at proximal region.

- 29
- Both sexes micropterous; antennal segment V yellow with very little brown on distal margin (Figure 28a) ……………………………… Asphodelothrips croceicollis (Karny)
Figure 28.
Antennal segment V: (a) pale yellow; (b) evenly brown; (c) bicoloured.

- 30
- Pronotum with five (rarely 4) pairs of prominent postero-marginal setae, with submedian pair more than twice as long as the discal setae (Figure 29a); male antennal segment VI as long as segments IV and V together (Figure 29c); male with a row of around 12 pore plates on each of abdominal sternites II–VII (Figure 29e) …………………………………………………………….. Pezothrips kellyanus (Bagnall)Pronotum with variable number of prominent postero-marginal setae, usually fewer than five and submedian pair never twice as long as discal setae (Figure 29b); male antennal segment VI shorter than segments IV and V together (Figure 29d); male with only one or no pore plate on each of abdominal sternites II–VII (Figure 29f) ……… 31
Figure 29.
Pronotum (a,b): (a) with five pairs of prominent postero-marginal setae; (b) with four pairs of prominent postero-marginal setae; antennal segments V–VIII (c,d): (c) antennal segment VI as long as IV + V; (d) antennal segment VI shorter than IV + V; male abdominal sternite (e,f): (e) with around 12 small pore plates per sternite; (f) with one large pore plate per sternite.
Figure 29.
Pronotum (a,b): (a) with five pairs of prominent postero-marginal setae; (b) with four pairs of prominent postero-marginal setae; antennal segments V–VIII (c,d): (c) antennal segment VI as long as IV + V; (d) antennal segment VI shorter than IV + V; male abdominal sternite (e,f): (e) with around 12 small pore plates per sternite; (f) with one large pore plate per sternite.

- 31
- Antennal segment VI with a sensorium of which the length of the base (insertion) of the sense cone is longer than the width of segment VII (Figure 30a) …………………………………………………………….. Odontothrips meliloti PriesnerAntennal segment VI with sensorium of which base is shorter than the width of segment VII (Figure 30b) …………………………………………………………………. 32
Figure 30.
Sensorium on antennal segment VI: (a) with base of sensorium longer than width of segment VII; (b) with base of sensorium shorter than width of segment VII.
Figure 30.
Sensorium on antennal segment VI: (a) with base of sensorium longer than width of segment VII; (b) with base of sensorium shorter than width of segment VII.

- 32
- Abdominal tergite VIII without paired lateral ctenidia (Figure 31a) often replaced by rows of microthrichia …………………………………………………………………….. 33Abdominal tergite VIII with paired lateral ctenidia (Figure 31b) sometimes with only few and sparse (not arranged in a single row) ………………………………………… 35
Figure 31.
Left side of abdominal segment VII: (a) lacking ctenidia; (b) with ctenidia.

- 33
- Fore wing clavus with five marginal setae (Figure 32a); second vein of fore wing with 8–10 setae ……………………………………………………. Tenothrips discolor (Karny)Fore wing clavus with six marginal setae (Figure 32b); second vein of fore wing with 11 or more setae …………………………………………………………………………… 34
Figure 32.
Clavus (a,b): (a) with six marginal setae; (b) with five marginal setae.

- 34
- Abdominal sternites bearing discal setae; ocellar setae iii arising just antero-lateral to ocellar triangle (Figure 33a); pronotum with one pair of postero-angular setae longer than discal setae (Figure 33c) …………………………………… Oxythrips ajugae Uzel
Figure 33.
Head (a,b): (a) showing ocellar setae iii just anterolateral to ocellar triangle; (b) showing ocellar setae iii arising outside ocellar triangle; pronotum (c,d): (c) with one pair of postero-angular setae longer than discal setae; (d) with two pairs of postero-angular setae longer than discal setae.
Figure 33.
Head (a,b): (a) showing ocellar setae iii just anterolateral to ocellar triangle; (b) showing ocellar setae iii arising outside ocellar triangle; pronotum (c,d): (c) with one pair of postero-angular setae longer than discal setae; (d) with two pairs of postero-angular setae longer than discal setae.

- 35
- Ctenidia on abdominal segment VIII situated antero-laterally to spiracle (Figure 34a); anterior margin of pronotum bearing at least one pair of long setae …………………………………………………………………….. Frankliniella Karny .. 36Ctenidia on abdominal segment VIII situated postero-mesad to spiracles (Figure 34b). anterior margin of pronotum with no long setae ………………….. Thrips Linnaeus 37
Figure 34.
Position of ctenidia on abdominal segment VIII: (a) antero-laterally to spiracle; (b) postero-mesad to spiracles.
Figure 34.
Position of ctenidia on abdominal segment VIII: (a) antero-laterally to spiracle; (b) postero-mesad to spiracles.

- 36
- Ocellar setae iii arising on anterior margins of ocellar triangle (Figure 35a); meta-thoracic campaniform sensilla present (Figure 35e); microtrichial comb at the posterior margin of tergite VIII fully developed (Figure 35d) ………………………………………………………. Frankliniella occidentalis PergandeOcellar setae iii arising close together between hind ocelli (Figure 35b); meta-thoracic campaniform sensilla absent (Figure 35d); microtrichial comb at the posterior margin of tergite VIII not developed or with few short teeth on lateral margins (Figure 35f) …………………………………………………………….. Frankliniella schultzei Trybom
Figure 35.
Head (a,b): (a) with ocellar setae iii arising on anterior margins of ocellar triangle; (b) with ocellar setae iii arising close together between hind ocelli; metanotum (c,d): (c) with campaniform sensilla present; (d) with no campaniform sensilla; abdominal tergite VIII (e,f): (e) with complete microtrichial comb; (f) with no microtrichial comb.
Figure 35.
Head (a,b): (a) with ocellar setae iii arising on anterior margins of ocellar triangle; (b) with ocellar setae iii arising close together between hind ocelli; metanotum (c,d): (c) with campaniform sensilla present; (d) with no campaniform sensilla; abdominal tergite VIII (e,f): (e) with complete microtrichial comb; (f) with no microtrichial comb.

- 37
- Antennae eight-segmented (Figure 16b) ………………………. Thrips simplex MorisonAntennae seven-segmented (Figure 16c) ………………………………………………. 38
- 38
- Abdominal tergite II with four lateral setae (Figure 36a); fore wing clavus with six marginal setae (Figure 32a) ………………………………….. Thrips australis (Bagnall)
Figure 36.
Abdominal tergite II (a,b): (a) with four lateral setae; (b) with three lateral setae.

- 39
- Abdominal tergite VIII with microtrichial comb medially incomplete (Figure 37a) ……………………………………………………………………………. Thrips major UzelAbdominal tergite VIII with microtrichial comb complete (Figure 37b) …………………………………………………………………….. Thrips tabaci Lindeman
Figure 37.
Abdominal tergite VIII: (a) medially incomplete; (b) complete.

3.2. Species Catalogue
- FAMILY AEOLOTHRIPIDAE
- Aeolothrips gloriosus Bagnall, 1914 †
- Material examined: MALTA: Buskett, 19.iv.2016, 2 ♀♀ (sm) on Laurus nobilis, GD; Buskett, 21.ii.2018, 1 ♀ (sm) on Rhamnus alaternus, GD; Xemxija, 3.iii.2018, 2 ♀♀ (sm) on Pistacea lentiscus, GD
- Body length: ♀: 1760–1940 µm; ♂: no records.
- Wing type: Both sexes are macropterous.
Aeolothrips gloriosus occurs on woody, often deciduous plants such as Tilia (Malvaceae), Fraxinus (Oleaceae), Sambucus (Adoxaceae), Prunus (Rosaceae), but also Citrus (Rutaceae) and Olea (Oleaceae) [14,20,26,27]. In the Maltese Islands, A. gloriosus was exclusively found on trees and shrubs including Laurus nobilis (Lauraceae), Pistacea lentiscus (Anacardiaceae) and Rhamnus alaternus (Rhamnaceae) in woodland and maquis habitats. This species was recorded as a facultative predator [29]. This Aegean species has a distribution which spans from Anatolia to Morocco and the Azores [14]. It has also been recorded from Iran [30] and the Near East [13].
- Aeolothrips intermedius Bagnall, 1934 †
- Material examined: MALTA: 03.iv.1959, 1 ♀ and instar larva (sm) on Gladiolus sp., ERS (BMNH); Wied Qirda, 06.iv.2016, 1 ♂ (sm) on Glebionis coronaria, GD; Wied Ħesri, 22.iv.2016, 2 ♀♀ (sm) on Avena sp., GD (BMNH); Wied Qirda, 17.iii.2017, 1 ♀ (sm) on Asphodelus ramosus, GD; Manikata, 29.iv.2017, 1 ♀ (sm) on Pallenis spinosa, GD; Fawwara, 22.v.2017, 1 ♀ (sm) from Malaise trap, DM; Lapsi, 09.x.2018, 1 ♀ (sm) on Hyparrhenia hirta, GD.
- Body length: ♀: 1700–1840 µm; ♂: 1300 µm.
- Wing type: Both sexes are macropterous.
A. intermedius has been recorded on a wide variety of unrelated plant species, often on yellow flowered plants such as those of Asteraceae, Cruciferae, Leguminosae, and Poaceae [15,21,24,26,31,32,33,34]. In the Maltese Islands, a larval instar was found on Gladiolus (Iridaceae) implying that this plant is used as a host by this species. Adults were found on annual herbaceous plants namely Asparagaceae, Asteraceae and Poaceae. This species was recorded as a facultative predator feeding on different insects including other thrips [32]. This species is widespread in Europe and Palaearctic Asia and the Middle East [13,14]. It has also been intercepted in the US from plant material originating from Europe [35].
- Aeolothrips melisi Priesner, 1936 †
- Material examined: MALTA: Wied Ħesri, 22.iv.2016, 1 ♀ (sm) on Capparis orientalis, GD; Popeye Village, 29.iv.2017, 1 ♀ (sm) on Tamarix africana, GD. GOZO: Ramla Bay, 18.iv.2017, 2 ♀♀ and 1 ♂ (sm) on Cakile maritima and 2 ♀♀ (sm) on Medicago marina, 18.iv.2017, GD.
- Body length: ♀: 1960–2550 µm; ♂: 1780 µm
- Wing type: Both sexes are macropterous.
Aeolothrips melisi was recorded on flowers of unrelated plants [14] including Brassicaceae [22,23] and Fabaceae [30] and this was also the case with the material collected from the Maltese Islands. This species was recorded as a facultative predator [29]. A. melisi is widespread throught the Mediterranean basin [13,19].
- Aeolothrips tenuicornis Bagnall, 1926
- Material examined: MALTA: Għammieri, 19.xii.1996, on Galium sp., 2 ♀♀ (sm), DM; M’Scala, 02.ii.1997, on Hedysarum coronarium, 2 ♀♀ (sm), DM; Siġġiewi (private garden), 04.xi.2015, 2 ♀♀ (sm) on Rosa sp., GD; Siġġiewi (private garden), 28.x.2015, 1 ♀ (sm) on Rosa sp., GD; Wied Ħesri, 15.i.2016, 1 ♂ (sm) on Silene colorata, GD; Siġġiewi (private garden), 29.iii.2016, 4 ♀♀ (sm) and 2 ♂♂ (sm) on Ranunculus asiaticus, GD; Siġġiewi (road), 04.iv.2016, 1 ♀ and 1 ♂ (sm) copula pair on Glebionis coronaria, GD; Wied Ħesri, 15.iv.2016, 1 ♀ (sm) on Convolvulus arvensis, GD; Wied Qirda, 06.iv.2016, 2 ♀♀ (sm) on Glebionis coronaria, GD; Wied Ħesri, 22.iv.2016, 1 ♀ (sm) on Capparis orientalis, GD; Kunċizzjoni, 22.iv.2016, 1 ♂ (sm) on Reichardia picroides, GD; Siġġiewi (road), 23.v.2016, 1 ♀ (sm) on Glebionis coronaria, GD; Siġġiewi (road), 21.vi.2016, 3 ♀♀ (sm, aga) on Glebionis coronaria, GD; Msida, Junior College grounds, 08.x.2016, 1 ♀ (sm) on Cynodon dactylon, GD; Lapsi 09.i.2017, 1 ♀ (sm) on Periploca angustifolia, GD; Siġġiewi (road), 21.iv.2017, 1 ♀ (sm) on Argyranthemum frutescens, GD; Mellieħa road l/o Popeye Village, 29.iv.2017, 1 ♂ (sm) on Convolvulus althoides, GD; Wied Ħesri, 30.iv.2017, 1 ♀ (sm) on Gladiolus communis, GD; Pembroke, 05.xi.2018, 1 ♀ (sm) on Reichardia picroides, GD. GOZO: Ramla Bay, 18.iv.2017, 1 ♀ (sm) on Malva arborea, GD.
- Body length: ♀: 1900–2400 µm; ♂: 1200–1680 µm.
- Wing type: Both sexes are macropterous.
Adult thrips of this species are found on flowers of unrelated plant species including Asteraceae, Brassicaceae, Fabaceae, [15,23,24,33,36], Citrus (Rutaceae) [24] and Cratageus (Rosaceae) [28,29], Triticum aestivum (Poaceae) and also on Verbascum (Scrophulariaceae) [24], Vitis (Vitaceae). In the Maltese Islands, this species was also found on flowers of unrelated indigenous plants including Asteraceae, Convolvulaceae, Fabaceae and Ranunculaceae. Aeolothrips tenuicornis is recorded as a facultative predator [32]. It occurs throughout most of Europe, the Middle East, the Mediterranean Region and the Atlantic Islands (Azores, Madeira, Canaries) [13,14,20,22,28,31].
- Franklinothrips megalops (Trybom, 1912) †
- Material examined: MALTA: Msida, University of Malta grounds, 04.xi.2016, 1 ♀ (sm) on Rosmarinus officinalis, DM; Fawwara, 22.v.2017, 1♀ (sm) from Malaise trap, DM.
- Body length: ♀: 2380–2480 µm, ♂: no records.
- Wing type: ♀: macropterous; ♂: no records.
Franklinothrips megalops is a predatory species [37], and has been observed to feed on other small arthropods including juvenile and adult stages of other Thysanoptera such as Heliothrips hemorrhoidalis and Thrips spp [14,23]. This species was described originally from Kenya but is in fact widespread in Africa [36], Spain and Israel, Yemen and southern India [14,23,36].
- Rhipidothrips brunneus Williams, 1913 †
- Material examined: MALTA: Wied Ħesri, 24.ii.2017, 2 ♀♀ (micropterous) (sm) on Bromus diandrus, GD.
- Body length: ♀: 1880–2060 µm; ♂: no records.
- Wing type: ♀: micropterous; ♂: no records.
This phytophagous species feeds on flowers of Poaceae [14,23,24,27,28,34] but was also recorded as having predatory tendencies [27]. R. brunneus is distributed throughout the western Palaearctic and the Near East, and has been accidentally introduced to North America and Australia [13,14,23] where it is an established species [28].
- Rhipidothrips gratiosus Uzel, 1895 †
- Material examined: MALTA: Wied Ħesri, 03.iv.2016, 1 ♀ (sm) on Avena sterilis, GD; Wied Ħesri, 22.iv.2016, 2 ♀♀ (sm) on Avena sterilis, GD; Fiddien, 14.iv.2016, 1 ♀ (sm) on Avena sp., GD; Wied Għollieqa 02.xii.2016, 1 ♂ (micropterous–sm) on Cynodon dactylon, GD; Wied Ħesri, 03.iv.2017, 4 ♀♀ (sm, aga) on Gladiolus communis, GD; Fiddien, 24.iv.2017, 1 ♀ (sm) on Avena sp., 1 ♀ (sm) on Medicago sp., GD; Wied Qirda, 17.iii.2017, 1 ♀ (sm) on Medicago sp., GD.
- Body size: ♀: 1900–2120 µm; ♂: 1240 µm.
- Wing type: ♀: macropterous; ♂: micropterous.
Males of this species have been described as being macropterous just like females [14,26], however, the male specimen examined from the Maltese Islands was found to be micropterous and marginally smaller than other males described from continental Europe [14]. Rhipidothrips gratiosus is phytophagous, typically found on the flowers of Poaceae [26,27,34,38,39] but also on other plants such as Erysimum cheiranthoides (Brassicaceae) [39] and Vicia faba (Fabaceae) [40]. In the Maltese Islands, this species has been largely collected from Poaceae. R. gratiosus was also recorded as having predatory habits [26]. The distribution of this species extends across the Western Palaearctic, the Near East and North Africa and has been introduced [13] and has become established in North America [14,24].
- Rhipidothrips niveipennis Reuter, 1899 †
- Material examined: MALTA: Wied Ħesri, 04.iv.2016, 1 ♀ (sm) on Avena sterilis, GD; Wied Ħesri, 22.iv.2016, 1 ♀ (sm) on Avena sterilis, GD; Gudja, l/o Malta International Airport, 26.ix.2016, 1 ♀ (sm) on Ficus microcarpa, GD.
- Body size: ♀: 2000–2014 µm; ♂: no records
- Wing type: ♀: macropterous; ♂: no records
Although material from the Maltese Islands consisted of only macropterous specimens, micropterous individuals have been described from continental Europe [14]. This species is phytophagous on different Poaceae [14] as was also largely the case for the specimens collected from the Maltese Islands. R. niveipennis is recorded from Finland, Sweden, the French Alps, [14] and Norway [41].
- Rhipidothrips unicolor zur Strassen, 1965 †
- Material examined: MALTA: Għajn Tuffieħa, 29.iv.2017, 7 ♀♀ (sm, aga) and instar larva (sm) on Stipa capensis, GD.
- Body size: ♀: 1760–2250 µm; ♂: n/a.
- Wing type: Both sexes are macropterous.
Rhipidothrips unicolor has been recorded, often in large numbers, on Stipa capensis (Poaceae) which has also been described as the host plant for this species [14,42]. In the Maltese Islands, both adults and larvae were also collected from the mentioned host plant. The geographical distribution for R. unicolor includes Greece, Italy, Spain, Canary Islands [14,42,43], Morocco [13,14] and Iran [31].
- FAMILY MELANTHRIPIDAE
- Melanthrips ficalbii Buffa, 1907 †
- Material examined: GOZO: Victoria, (road), 20.iv.2019, 2 ♀♀ (sm) on Galium aparine, GD.
- Body length: ♀: 1840–1960 µm; ♂: n/a.
- Wing type: Both sexes are macropterous.
Melantrhips ficalbii is a phytophagous species with larval stages being found on Rubiaceae [33] indicating that these plants are used as hosts by this species. Adults have also been recorded on Galium (Rubiaceae), Euphorbia cyparissias (Euphorbiaceae), Reseda (Resedaceae) as well as on different Poaceae [14]. In the current study, M. ficalbii has been recorded from Galium aparine (Rubiaceae). M. ficalbii is widespread in western Europe and the western Mediterranean Region [13,14,25,35].
- Melanthrips fuscus Sulzer, 1776
- Material examined: MALTA: Siġġiewi (private garden), 04.xi.2015, 3 ♂♂ (sm) on Rosa sp., GD; Siġġiewi (private garden), 12.i.2016, 3 ♀♀ and 3 ♂♂ (sm) on Rosa sp., GD; Siġġiewi (private garden), 15.i.2016, 2 ♀♀ (sm) and 2 ♂♂ (sm) on Mercurialis annua, GD; Wied Ħesri, 15.i.2016, 1 ♀ (sm) on Silene colorata, GD; Wied Qirda, 29.i.2016, 2 instar larvae (aga) and 1 ♂ (sm) on Brassica rapa, GD; Siġġiewi (private garden), 29.iii.2016, 1 ♀ (sm) and 1 ♂ (sm) on Ranunculus asiaticus, GD; Siġġiewi (private garden), 16.x.2016, 1 ♀ (sm) on Lobularia maritima, GD; Wied Qirda, 17.iii.2017, 1 ♂ (sm) on Asphodelus ramosus, GD; Wied Ħesri, 03.iv.2017, 2 ♀♀ (sm) and 1 ♂ (sm) on Acacia saligna, GD; Dingli Cliffs, 27.iv.2017, 1 ♂ (sm) on Brassica rapa, GD; Lapsi, 09.x.2017, 1 ♂ (sm) on Potentilla reptans, GD; Wied Baqqiegħa, 22.i.2018, 1 ♀ (sm) on Cerinthe major, GD.
- Body length: ♀: 1540–2140 µm; ♂: 1360–1560 µm.
- Wing type: Both sexes are macropterous.
This species is phytophagous with overwhelming records from plants in the Brassicaceae [14,23,24,26,32,34,36]. It has also been recorded from annual plants [24,32,34,36,44], grasses and trees [45]. In The Maltese Islands, larvae were found on Brassica rapa (Brassicaceae). Melantrips fuscus adults were collected together with M. lybicus on flowers of many unrelated indigenous and cultivated plants including Brassicaceae, Rosaceae, Caryophyllaceae, Xanthorrhoeaceae and Fabaceae. M. fuscus is found across Europe, the East Palaearctic, the Nearctic and North Africa [22,23]. It has also been intercepted in US from plant material imported from Europe, the Mediterranean and Africa [35].
- Melanthrips knechteli Priesner, 1936 †
- Material examined: MALTA: Siġġiewi (private garden), 15.i.2016, 1 ♀ (sm) on Mercurialis annua, GD; Siġġiewi (private garden), 29.iii.2016, 1 ♂ (sm) on Ranunculus asiaticus, GD; Wied Xkora, 30.x.2017, 1 ♂ (sm) on Mercurialis annua, GD; Wied Baqqiegħa, 22.i.2018, 1 ♀ (sm) on Cerinthe major, GD.
- Body length: ♀: 1580 µm; ♂: 1160–1240 µm.
- Wing type: Both sexes are macropterous.
Melanthrips knechteli has been recorded on Cerinthe major (Boraginaceae) and Thymus (Lamiaceae) [14,23]. In the Maltese Islands, this species was found on Cerinthe major but also on Ranunculus asiaticus (Ranunculaceae) and Mercurialis annua (Euphorbiacceae). The geographical distribution for this species includes Turkey, Bulgaria, Romania, Albania, Czech Republic (Moravia), Spain and Iran [14,26,46,47].
- Melanthrips libycus Priesner, 1936
- Material examined: MALTA: Siġġiewi (private garden), 12.i.2016, 1 ♀ (sm) and 1 ♂ (sm) on Rosa sp., GD; Wied Ħesri, 15.i.2016, 1 ♀ (sm) and 2 ♂♂ (sm) on Silene colorata, GD; Wied Qirda, 29.i.2016, 1 ♀ (sm) on Trifolium nigrescens, 1 ♀ (sm) on Brassica sp., GD; Qormi (private farm), 07.ii.2016, 1 ♀ (sm) on Brassica oleracea var. botrytis, GD; Wied Qirda, 24.ii.2016, 1 ♀ (sm) on Bromus diandrus, GD; Ta’ Qali, 27.ii.2016, 5 instar larvae (aga) and 2 ♀♀ (sm) on Diplotaxis tenuifolia, SF; Siġġiewi (private garden), 29.ii.2016, 1 ♀ (sm) on Ranunculus asiaticus, GD; Wied Qirda, 17.iii.2017, 1 ♀ (sm) on Medicago sp., GD; Wied Ħesri, 03.iv.2017, 1 ♀ (sm) on Acacia saligna, GD; Dingli Cliffs, 29.x.2017, 1 ♀ (sm) on Brassica rapa, GD. GOZO: Ramla Bay, 18.iv.2017, 1 ♂ (sm) on Cakile maritima, GD.
- Body length: ♀: 1600–2300 µm; ♂: 1400–1725 µm.
- Wing type: Both sexes are macropterous.
The long setae present on the head of this species were described to be as long as three pairs of post-ocular setae [14,48] but in specimens collected from the Maltese Islands these setae were found to be shorter. Melanthrips lybicus has been recorded on Poaceae [24], Brassicaceae [15,23,44,48] and also from Citrus x paradisi (Rutaceae) [15], though the literature does not specify which species are used as host plants. In the current study, larvae of this species were found on Diplotaxis tenuifolia (Brassicaceae). M. lybicus adults together with M. fuscus has been recorded on a variety of unrelated plants such as Brassicaceae, Fabaceae and Poaceae. M. lybicus is widely distributed across southern Europe and the Mediterranean basin [14,15,23,49].
- FAMILY STENUROTHRIPIDAE
- Holarthrothrips tenuicornis Bagnall, 1927 †
- Material examined: MALTA: Qormi (roundabout), 14.ix.2017, 13 ♀♀ (sm, aga) on male Phoenix dactylifera, GD; Qormi (roundabout), 03.x.2017, 13 ♀♀ (sm, aga) on male flowers of Phoenix dactylifera, GD; Msida, Junior College grounds, 10.x.2017, 1 ♀ on male flowers of Phoenix dactylifera, GD; Siġġiewi (private garden), 30.vii.2018, 2 ♀♀ (sm), 10 ♀♀ (aga) and 1 ♂ (sm), 1 ♂ (aga) on male flowers of Phoenix dactylifera, GD.
- Body length: ♀: 1550–1820 µm; ♂: 1260 µm.
- Wing type: Both sexes are macropterous.
Thrips of the genus Holarthrothrips are mostly associated with male flowers of Phoenix trees (Arecaceae) [15,50] where both larvae and adults occur. Adults of this species have also been recorded on other plants such as Vitis (Vitaceae), Citrus (Rutaceae), and Rubia (Rubiaceae) [14,24,29] which, however, should not be considered as host-plants. In the Maltese Islands larvae and adults were always found on male flowers of Phoenix dactylifera. H. tenuicornis has been recorded from across the Mediterranean basin [13,14,15,23,29,43].
- FAMILY THRIPIDAE
- Subfamily Dendrothripinae
- Dendrothrips saltator Uzel, 1895 †
- Material examined: MALTA: Wied Babu, 15.xii.1996, 7 ♀♀ (sm) on Ferula melitensis, DM; Wied Qirda, 06.iv.2016, 13 instar larvae (aga) on Foeniculum vulgare, GD; Maqluba, l/o Qrendi, 08.iv.2016, 9 ♀♀ (sm, aga) on Foeniculum vulgare, GD; Qormi (road), 19.iv,2016, 1 instar larva (aga) on Ferula melitensis, GD; 09.vii.2018, 6 ♀♀ (sm, aga) and 1 ♂ (sm) on Foeniculum vulgare, GD.
- Body length: ♀: 1260–1440 µm; ♂: 860 µm.
- Wing type: Both sexes are macropterous.
Dendrothrips saltator has been reported from leaves of various deciduous trees and a variety of unrelated plants [14,26], particularly on Apiaceae, Asteraceae [14,23,26], Fabaceae, as well as plants such as Triticum aestivum (Poaceae) and Olea europaea (Oleaceae) [34]. In the Maltese Islands, both adults and larvae of D. saltator were found on Foeniculum vulgare and Ferula melitensis (both in Apiaceae). Thus, most likely, the host plants for Dendrothrips saltator are only plants within the family Apiaceae. In fact, in the UK, this species is only recorded on Paucedanum officinale (Apiaceae) [27]. D. saltator is widespread in Europe, the Oriental Region and the Near East [13,14]. It was also intercepted in the US on plant material originating from Europe [35,51].
- Subfamily Panchaetothripinae
- Heliothrips haemorrhoidalis (Bouchè, 1883)
- Material examined: MALTA: Żabbar, 06.v.1996, 1 ♀ (sm) on Viburnum sp., CF; Msida, University of Malta grounds, 28.iv.2017, 1 ♀ (sm) on Apium graveolens, GD.
- Body length: ♀: 1660–1900 µm; ♂: no records.
- Wing type: ♀: macropterous; ♂: no records.
Heliothrips haemorrhoidalis is a polyphagous species recorded on different unrelated host-plants [23,24,26,32]. It is an agricultural pest and can potentially inflict damage on a number of crop plants in both greenhouses [23] and open fields [26]. In the Maltese Islands H. haemorrhoidalis was collected on Viburnum (Adoxaceae), Apium graveolens (Apiaceae) as well as from plants with low levels of nitrogen [12]. This species is preyed upon by Franklinothrips megalops [26]. H. haemorrhoidalis is believed to have originated in South America, probably Peru [51], but has become widespread around the world in tropical and subtropical areas [24], being introduced with exotic ornamental plants. In northern Europe, H. haemorrhoidalis thrives in greenhouses and indoor places [14].
- Hercinothrips femoralis (Reuter, 1881) †
- Material examined: MALTA: Msida, University of Malta grounds, 04.v.2016, 1 ♀ (sm) on Origanum majorana, GD; Siġġiewi (private garden), 05.xi.2016, 14 ♀♀ (sm, aga) and 1 instar larva (aga) on Hippeastrum sp., GD; Birguma, 20.viii.2020, 13 ♀♀ (sm, aga) on Ocimum basilicum, NY; Siġġiewi private garden, 04.xi.2020, 8 ♀♀ (sm, aga) on Calendula officinalis, GD.
- Body length: ♀: 1320–1580 µm: ♂: no records.
- Wing type: ♀: macropterous; ♂: no records.
Hercinothrips femoralis is a polyphagous species occurring on herbs and cultivated plants, mostly in greenhouses [23,24] and often on Amaryllidaceae [14] but also on other crops [26]. In the Maltese Islands, H. femoralis was found on different unrelated plants but only the Hippeastrum sp. (Amaryllidaceae) can be regarded as a host-plant for the mentioned thrips. Although this species was recorded in Europe from greenhouses, it was found in plants grown outside in the Maltese Islands, probably due to the warmer climate of the archipelago. H. femoralis has been recorded as a pest, inflicting damage in greenhouse crops [27]. H. femoralis is native to Central Africa, but is now subcosmopolitan in distribution with many records in tropical and subtropical countries [14,27,35].
- Subfamily Thripinae
- Anaphothrips sudanensis Trybom, 1911 †
- Material examined: MALTA: Wied Ħesri, 04.xi.2016, 1 ♀ (sm) on Cynodon dactylon, GD.
- Body length: ♀: 1340 µm; ♂: no records.
- Wing type: the locally collected female specimen is macropterous.
Anaphothrips sudanensis is a phytophagous species associated with Poaceae [23]. It is commonly found in the leaf axils and tend to develop streaks on leaves as these expand and mature [28]. This species has been recorded on different Poaceae, particularly on Panicum maximum, Pennisetum, Sorghum halepense, Zea mays and Saccharum officinarum [14,26,28], as well as on Allium cepa (Amaryllidaceae) [24] and Tagetes minuta (Asteraceae) [26]. In the Maltese Islands, the single female specimen of this species was collected on Poaceae (Cynodon dactylon). A. sudanensis is recorded from the East Palaearctic, Afrotropical, Australian, Neotropical and Oriental Regions, the Near East and North Africa [13].
- Aptinothrips rufus (Haliday, 1836) †
- Material examined. MALTA: Wied Ħesri, 04.iv.2016, 1 ♀ (sm) and 1 instar larva (aga) on Hyparrhenia hirta and 4 ♀♀ (sm) and 1 instar larva (aga) on Avena sterilis, GD; Wied Qirda, 06.iv.2016, 1 ♀ (sm) on Plantago major, GD; Maqluba l/o Qrendi, 15.iv.2016, 8 ♀♀ (sm) on Triticum aestivum, GD; Wied Ħesri, 15.iv.2016, 1 ♂ (sm) on Convolvulus arvensis, GD; Msida, Junior College grounds, 27.iv.2016, 1 ♀ (sm) and 2 instar larvae (aga) on Hordeum leporinum, GD; Kunċizzjoni, 22.i.2017, 1 ♀ (sm) on Hordeum leporinum, GD; Msida, Junior College grounds, 03.iv.2017, 2 ♀♀ (sm) on Hordeum leporinum, GD; Għajn Tuffieħa, 29.iv.2017, 1 ♀ (sm) on Stipa capensis, GD. GOZO: Ramla Bay, 18.iv.2017, 1 ♀ (sm) on Medicago marina, GD.
- Body length: ♀: 1460–1700 µm; ♂: 900 µm.
- Wing type: Both sexes are apterous.
Aptinothrips rufus is associated with Poaceae [15,22,26,44,52] but adults were recorded on unrelated plants belonging to the following families: Fabaceae, Lamiaceae, Plumbaginaceae [44], Polygonaceae [34], and Asteraceae [26]. In the Maltese Islands, both adults and larvae of A. rufus were only found on Poaceae (Avena sp., Hordeum leporinum and Hyparrhenia hirta), though adults were also found on other plants as well as grasses. A rufus is a species that originated from Europe, but has become widespread in temperate regions around the world [14,32,52]. In fact, it is recorded from the East Palaearctic, Afrotropical, Australian, Nearctic, Neotropical and Oriental Regions, the Near East and North Africa [13]. It has been intercepted in the US from plants originating from Europe, the Mediterranean and Africa [34,53].
- Asphodelothrips croceicollis (Karny, 1914) †
- Material examined: MALTA: Dingli Cliffs, 27.i.2016, 4 ♂♂ (sm) on Asphodelus ramosus, GD; Buskett, 19.ii.2016, 2 instar larvae (aga) on Asphodelus ramosus, GD; Dingli Cliffs, 24.i.2017, 5 ♀♀ (sm) on Asphodelus ramosus, GD; Kunċizzjoni, 28.i.2017, 2 ♀♀ (sm) and 1 ♂ (sm) on Asphodelus ramosus (ag) and 1 ♀ (sm) on Erica multiflora, GD; Wied Ħesri, 24.ii.2017, 1 ♀ (sm) and 1 ♂ (sm) and on Asphodelus ramosus, GD; Buskett, 23.ii.2018, 2 ♀♀ (sm) on Asphodelus ramosus, GD; Kunċizzjoni, 29.i.2018, 1 ♀ (sm) on Glebionis coronaria, GD.
- Body length: ♀: 1760–2160 µm; ♂: 1000–1580 µm.
- Wing type: Both sexes of locally recorded specimens are micropterous.
Both micropterous as well as macropterous forms for this species exist, with the macropterous females being dark brown in colour and the micropterous forms being light brown [14]. All specimens collected from Malta were found to be micropterous, with females being of a dark brown colour. Asphodelothrips croceicollis is associated with Asphodelus spp. (Asphodelaceae) [14,29] and in the Maltese Islands it was almost exclusively collected on flowers of Asphodelus ramosus. A. croceicollis is found throughout the Ponto-Mediterranean area and the Canary Islands, occurring up to 1500 m altitude [14], as well in the Near East [13].
- Bregmatothrips dimorphus (Priesner, 1919) †
- Material examined: MALTA: Wied Ħesri, 15.i.2016, 5 ♀♀ (aga) and 4 ♂♂ (sm, aga) on Pipatherum miliaceum, GD; Buskett, 03.ii.2016, 1 ♀ (sm) on Cynodon dactylon, 3 ♀♀ (sm, aga) and 2 ♂♂ (sm, aga) on Pipatherum miliaceum, GD.
- Body length: ♀: 1440–1460 µm; ♂: 1140–1240 µm.
- Wing type: ♀: macropterous; ♂: micropterous.
The female specimens collected in the Maltese Islands were all macropterous, but micropterous forms are also described from the European continent [32]. Bregmatothtips dimorphus is associated with Poaceae [34] and this was also the case for the material collected from the Maltese Islands which was found on the grass species Cynodon dactylon and Pipatherum miliaceum. This species is widespread in Europe but is also recorded from Yemen, Sudan, southern Africa [14,23,26], the Near East and North Africa [13].
- Ceratothrips ericae (Haliday, 1836) †
- Material examined: MALTA: Lapsi, 9.x.2017, 1 ♀ (sm) on Limbarda crithmoides, GD. GOZO: Qbajjar, 31.iii.2018, 1 ♀ (sm) on Helychrysum melitense, GD.
- Body length: ♀: 1200–1460 µm; ♂: no records.
- Wing type: ♀: macropterous; ♂: no records.
Different authors have recorded this species from unrelated plants [14,27,29,35,54,55]. In the Maltese Islands, Ceratothrips ericae was found on Avena sterilis (Poaceae) and Helychrysum melitense (Asteraceae). C. ericae is an important pollinator, especially in areas where larger pollinators do not occur [56]. This species is of Euro-Siberian origin, but has been accidentally introduced to other parts of the world [14], including the Near East and the Australian Region [13]. It has also been intercepted in the US from plants originating from Europe [35].
- Chirothrips hamatus Trybom, 1895 †
- Material examined: GOZO: Xlendi, 21.vii.1956, 1 ♀ (sm) on Lithospermum arvense (JIS & ERS), (BMNH).
- Body length: ♀: 1453 µm; ♂: no records.
- Wing type: ♀: macropterous; ♂: no records.
Males of this species are described as being micropterous [14,24,28]. Chirothrips hamatus is mostly associated with Poaceae, particularly Alopecurus pratensis [14,24,27] and thus, Lithospermum arvense (Boraginaceae), on which the single female specimen was collected during the present study, cannot be considered as a host plant. C. hamatus is recorded from western Siberia, the Near East and the Nearctic Region; it is also widespread in Europe being generally less common in southern areas [13,14,27].
- Chirothrips manicatus (Haliday, 1836) †
- Material examined: MALTA: Siġġiewi (private farm), 03.v.2016, 1 ♀ and 1 ♂ (sm) on Koeleria cristata, GD; Wied Ħesri, 02.xi.2016, 1 ♂ (sm) on Phragmites australis, GD; Wied Ħesri, 04.xi.2016, 4 ♀♀ (sm, aga) and 5 ♂♂ (sm, aga) on Arundo donax, GD; Naxxar (road), 29.iv.2017 1 ♂ (sm) on Triticum aestivum, GD; Wied Qirda, 31.v.2018, 1 ♂ (sm) on Hyparrhenia hirta, GD; Fiddien 16.vi.2021, 1♀ (sm) on Avena sterilis, GD.
- Body length: ♀: 1400–1700 µm; ♂: 960–1440 µm.
- Wing type: ♀: macropterous; ♂: micropterous.
Chirothrips manicatus is a species which is mostly associated with Poaceae and sometimes Cyperaceae [23]. It has been also recorded on other unrelated plants such as Malva alcea (Malvaceae) and Malus sp. (Rosaceae) [54,57,58,59]. In the Maltese Islands, this species was only collected from Poaceae. C. manicatus may transmit phytopathogenic bacteria and fungi [25]. Chirothrips manicatus probably originated in the Palaearctic but has a Holarctic distribution and is nowadays considered as subcosmopolitan in distribution, with numerous records in temperate parts of the world [14].
- Chirothrips meridionalis Bagnall, 1927 †
- Material examined: MALTA: Wied Ħesri, 04.iv.2016, 2 ♂♂ (sm) on Hyparrhenia hirta, GD. Wied Qirda, 04.xi.2016, 2 ♀♀ (sm) on Arundo donax, GD; Wied Qirda, 31.v.2018, 3 ♀♀ (sm) on Hyparrhenia hirta, GD (1 ♀ ag).
- Body length: ♀: 1600–1680 µm; ♂: 1240 µm.
- Wing type: Both sexes are macropterous.
Similar to other species of the genus Chirothrips, C. meridionalis is also associated with Poaceae [14,22,23,26]. In the Maltese Islands this thrips was also collected on plants belonging to the mentioned plant family. C meridionalis has been recorded from South Europe [22] and is widespread across the Mediterranean basin, Madeira, Canary Islands, India, Pakistan, Iran, Yemen, Nigeria and South Africa [14]. It has also been intercepted in the US from plants originating from Europe, the Mediterranean and Africa [35].
- Echinothrips americanus Morgan, 1913 †
- Material examined: MALTA: Siġġiewi (private farmhouse), 21.v.2017, 1 ♀ (sm) on Salvia officinalis, GD; Siġġiewi (private garden), 14.vii.2017, 13 ♀♀ (sm, aga) on Azalea indica, Siġġiewi (private garden), 16.vii.2017, 8 ♀♀, 3 instar larvae (aga) on Azalea indica, GD.
- Body length: ♀: 1100–1340 µm; ♂: no records.
- Wing type: ♀: macropterous; ♂: no records.
This phytophagous species has been recorded from unrelated plants including many vegetables [50], Impatiens (Balsaminaceae), Dendranthema (Asteraceae) and Euphorbia (Euphorbiaceae) [32]. It is believed to have been introduced to many countries around the world with trade of ornamental plants. This species has been recorded as a pest on cultivated plants such as Philodendron (Araceae) and Syngonium (Araceae), Impatiens, and Euphorbia [32,48]. Echinothrips americanus is native to eastern North America and has been imported with ornamental plants to Europe [14,32], Neotropical and Oriental Regions [13].
- Frankliniella occidentalis (Pergande, 1895)
- Material examined: MALTA: Żabbar, 11.iii.1994, 2 ♀♀ (sm) (BMNH) on Gerbera sp., DM; St. Paul’s Bay, 14.iii.1994, 2 ♀♀ (sm) (BMNH) on Solanum melanogena, GW; St. Paul’s Bay, 14.iv.1994, 1 ♀ (sm) (BMNH) on Dianthus caryophyllus, JWI; St. Paul’s Bay, 14.iv.1994, 2 ♀♀ (sm) (BMNH) on Chrysanthemum sp., JWI; St. Paul’s Bay (glasshouse), 14.iii.1994, 2 ♀♀ (sm) (BMNH) on Fragraria x ananassa, GW; Żabbar, 13.i.1997, 1 ♀ (sm) (dark morph) on Dianthus caryophyllus, DM; Siġġiewi (road), 14.x.2015, 1 ♀ (sm) (yellow morph) on Diplotaxis tenuifolia, GD; Siġġewi (private garden), 29.i.2016, 1 ♀ (sm) (yellow morph) on Narcissus tazzetta, GD; Wied Qirda, 06.iv.2016, 1 ♂ (sm) on Matricaria chamomilla, GD; Wied Ħesri, 04.iv.2016, 1 ♂ (sm) on Avena sterilis, GD; Siġġiewi (private garden), 13.iv.2016, 1♀ (sm) (yellow morph) on Foeniculum vulgare, GD; Manikata, 21.iv.2017, 1 ♂ (sm) on Pallenis spinosa, GD; Mellieħa (road l/o Popeye Village), 21.iv.2017, 1 ♀ (sm) (yellow morph), 1 ♀ (sm) (dark form) on Tamarix africana, GD; Il-Ballut, l/o M’Xlokk, 05.v.2017, 1 ♀ (sm) (pale morph), 1 ♀ (sm) (dark morph) on Malva arborea, GD; road, l/o Wied Ħesri, 07.viii. 2017, 1 ♀ (sm) (pale morph), 3 ♀♀ (aga) on Ipomoea carnosa, GD; Qormi (road), 18.viii.2017, 1 ♀ (sm) (pale morph), 3 ♀♀ (aga) (pale morph) on Yucca gloriosa, GD; San Ġwann (private farm), 29.x.2018, 2 ♀♀ (sm) (yellow morph), 3 ♀♀ (aga) (pale form) and 1 ♂ on Lactuca sativa, GD; Wied Ħesri, 12.iii.2018, 2 ♀♀ (sm) (dark morph) on Trifolium nigrescens, GD; Fiddien, 16.v.2021, 1 ♀ (sm) on Avena sterilis, GD.
- Body length: ♀: 1440–1800 µm; ♂: 1080–1300 µm.
- Wing type: Both sexes are macropterous.
The extent of the brown color pattern on antennae and overall color intensity of this species is likely dependent on temperature, with darker specimens developing in cooler temperatures [27]. Both dark and yellow morphs were collected from the Maltese Islands. Frankliniella occidentalis is polyphagous, with adults and larvae commonly found on both flowers and leaves of unrelated plants [29] including numerous crops [23] and indigenous plants [27,35,40,54,55,59]. In the Maltese Islands this species was recorded on seed crops, cut flowers such as Dahlia (Asteraceae), Dianthus (Caryophyllaceae) and Gerbera (Asteraceae), nursery stock, peaches, plums, nectarines, strawberries, sweet peppers, grapes, cotton and other crops including tomatoes [12]. This species is particularly harmful to crops, fruits and cultivated flowers and is a vector of a number of orthotospoviruses such as Tomato Spotted Wilt Virus (TSWV). Nonetheless, F. occidentalis has also been described to feed on the eggs and adults of mites species which can be harmful to agricultural crops [60]. F. occidentalis is native to Pacific North America, but has been imported with cultivated plants to virtually all countries in the world during the early 1980s, thriving at first in greenhouses but subsequently spreading out in the open especially in areas with warmer climates such as the Mediterranean basin [14,32].
- Frankliniella schultzei (Trybom, 1910) †
- Material examined: MALTA: Siġġiewi (private garden), 04.xi.2015, 1 ♀ (sm) (yellow morph) form) on Rosa sp.
- Body length: ♀: 1400 µm; ♂: no records.
- Wing type: ♀: mactopterous; ♂: no records.
The specimen collected from the Maltese Islands was yellow but dark morphs of this species also exist as in the case of Frankliniella occidentalis. F. schultzei was recorded on flowers and leaves of a wide variety of different unrelated plants [14,25,26,30,33,61], crops [25,48] and stored bulbs [28]. It is known to cause damage to different crops and is an orthotospovirus vector [14,25]. F. schultzei is generally considered as native to South America, but it could also be of Africotropical origin [26]. It is widespread in circum-tropical and subtropical regions, where it is recorded as a common pest [14].
- Limothrips angulicornis Jablonowski, 1894 †
- Material examined: MALTA: Fiddien, 24.iv.2017, 1 ♀ (sm) on Avena sp., GD.
- Body length: ♀: 2080 µm; ♂: no records.
- Wing type: ♀: macropterous; ♂: no records.
Limothrips angulicornis is phytophagous and associated with leaves and leaf axils of grasses [14,44]. Larvae have been found on Poaceae [62] but adults were also recorded from Populus sp. (Salicaceae) and Pinus sp. (Pinaceae) [34]. In the Maltese Islands, this thrips was collected from Avena (Poaceae). L. angulicornis originates from the West Palaearctic, but has been imported to other parts of the world such as Western Asia [22], California, Chile and southern Australia where it has become established [14,26,27].
- Limothrips cerealium (Haliday, 1836) †
- Material examined: MALTA: Wied Qirda, 10.iv.2016, 2 ♀♀ (sm) on Bromus diandrus, GD; Wied Ħesri, 22.iv.2016, 1 ♀ (sm) on Hyparrhenia hirta, GD; Wied Speranza, 26.iv.2016, 1 ♀ (sm) on Mentha pulegium, GD; Junior College grounds, 27.iv.2016, 1 ♀ (sm) on Hordeum leporinum, GD; Dingli Cliffs, 24.v.2016, 1 ♀ (sm) on Brassica rapa, GD; Dingli Cliffs, 27.v.2016, 1 ♀ (sm) on Glebionis coronaria, GD; Fiddien, 24.iv.2017, 2 ♀♀ (sm) on Avena sp., GD; Naxxar, road, 29.iv.2017, 1 ♀ (sm) on Avena sterilis and 1 ♂ (sm) on Triticum aestivum, GD; Wied Ħesri, 12.iii.2018, 1 ♀ (sm) on Trifolium nigrescens, GD.
- Body length: ♀: 1520–2200 µm; ♂: 1260 µm.
- Wing type: ♀: macropterous; ♂: micropterous.
Limothrips cerealium is phytophagous and generally found on Poaceae, namely cereals [25] and grasses [22,34,35,55,63] where larvae were also recorded [27]. In the Maltese Islands, this species was mainly found on Poaceae, but also on unrelated herbaceous plants such as Brassica rapa (Brassicaceae) and Glebionis coronaria (Asteraceae). This species can cause damage to grains, causing “silvering” on leaves [23,26,48] and is possibly a mechanical vector of phytopathogenic bacteria and fungi [23]. L. cerealium probably originated in Atlantic western Europe [14], but has been accidentally introduced on cereals and grains in many parts of the world and is now considered subcosmopolitan in distribution [14].
- Odontothrips meliloti Priesner, 1951 †
- Material examined: MALTA: Wied Qirda, 17.iii.2017, 1 ♀ (sm) on Hedysarum coronarium, GD; Fiddien, 24.iv.2017, 1 ♀ (sm) on Medicago sp., GD; Il-Ballut l/o M’Xlokk, 05.v.2017, 1 ♀ (sm) on Medicago arborea, GD; Siggiewi (private residence), 18.iii.2018, 1 ♀ on glass window pane, GD. GOZO: Xlendi, 31.iii.2016, 2 ♀♀ (sm) on Lotus ornithopodioides, GD; Ramla Bay, 17.iii.2017, 1 ♂ (sm) on Medicago marina, GD.
- Body length: ♀: 1900–2460 µm; ♂: 1500 µm.
- Wing type: Both sexes are macropterous.
Odontothrips meliloti is recorded from flowers of Fabaceae, specifically on those of the genus Melilotus (M. albus and M. officinalis) [23,33]. In the Maltese Islands, it was also collected from flowers of Fabaceae (Hedysarum coronarium, Lotus ornithopodioides, Medicago sp.). This species has a Euro-Siberian distribution and is widespread in central and southern Europe [14].
- Oxythrips ajugae Uzel, 1895 †
- Material examined: MALTA: Wied Għollieqa, 10.iii.2017, 10 ♀♀ (aga, sm) on Pinus halepensis, GD; Wied Qirda, 17.iii.2017, 6 ♀♀ (aga, sm) on Pinus halepensis, GD; Kunċizzjoni, 29.i.2018, 5 ♀♀ (sm) and 3 ♂♂ (sm) on Pinus halepensis, GD; Xemxija, 03.iii.2018, 5 ♀♀ (sm) on Pinus halepensis, GD.
- Body length: ♀: 1500–1700 µm; ♂: 920 µm.
- Wing type: Both sexes are macropterous.
Oxythrips ajugae is a spring occurring [14] dendrophilous foliivore [64]. It has been recorded on Pinaceae such as Pinus and Cupressus [14,29], but also on a number of hardwoods and herbaceous plants [34]. In the Maltese Islands, adults and larvae of this species were found in spring on Pinus halepensis (Pinaceae), indicating that this plant is used as a host by the mentioned thrips. O. ajugae is widespread in Europe [14] and Asia [32], the Nearctic Region, the Near East and North Africa [13].
- Pezothrips kellyanus (Bagnall, 1916) †
- Material examined: MALTA: Siġġiewi (private garden), 04.xi.2015, 3 ♂♂ (sm) on Rosa sp.
- Body length: ♀: no records; ♂: 1460–1660 µm.
- Wing type: ♀: no records; ♂: macropterous.
Pezothrips kellyanus is found on highly scented white flowers [28,65] especially those of Citrus (Rutaceae) [48,65,66], but also on Gardenia (Rubiaceae) [65], Jasminum (Oleaceae) [32,65,66,67], Lonicera japonica (Caprifoliaceae) [67], Olea europea (Oleaceae) [34] and Pittosporum tobira (Pittosporaceae) [27,67]. Immature stages of this species were only found on lemon, grapefruit, Jasminum and Gardenia flowers [65], however, the flowers of Camellia (Theaceae), Chrysanthemum (Asteraceae), Rosa (Rosaceae), Prunus (Rosaceae), Jasminum (Oleaceae), and Passiflora (Passifloraceae) can sustain the breeding of this species in times when citrus plants are not in bloom and at times when the climate becomes unfavourable, to then reinfest citrus trees when the climate temperature becomes warmer and the citrus trees bloom again [68]. In the Maltese Islands this species was collected from flowers of Rosa sp. (Rosaceae) found in close proximity to Citrus trees (Rutaceae). P. kellyanus is often recorded as an agricultural pest, causing scarring on fruits, particularly those of citrus trees [48,66]. This species may have originated in Australia [27] but was accidentally introduced in many territories, particularly in Europe [27].
- Prosopothrips nigriceps Bagnall, 1927 †
- Material examined: MALTA: Pembroke, 05.xi.2018, 1 ♀ (sm) on Reichardia picroides, GD; Xrobb l-Għaġin, 14.iii.2018, 2 ♀♀ (aga) on Hedysarum glomeratum, GD. GOZO: Ramla Bay, 18.iv.2017, 6 ♀♀ (aga, sm) on Silene colorata, GD.
- Body length: ♀: 1340–1500 µm; ♂: no records.
- Wing type: ♀: apterous; ♂: no records.
Prosopothrips nigriceps is mainly recorded from coastal regions on dune grasses such as Lygeum spartum and Poa annua, as well as on low-growing vegetation [14]. In the Maltese Islands, this species was mostly collected from coastal garigue areas on different unrelated plants such as Reichardia picroides (Asteraceae), Hedysarum glomeratum (Fabaceae) and Silene colorata (Caryophyllaceae). P. nigriceps is distributed throughout the Mediterranean basin [14]. It has also been intercepted in the US from plants originating from southern Europe and the Mediterranean [35].
- Tenothrips discolor (Karny, 1907)
- Material examined: MALTA: St. Paul’s Bay, vii.1956, 1 ♀ (sm) (BMNH) on Limbarda crithmoides, ERS.
- Body length: ♀: 1200 µm; ♂: no records.
- Wing type: ♀: macropterous; ♂: no records.
Although mostly associated with Asteraceae, Tenothrips discolor is regarded as polyphagous [14] and recorded on several unrelated plants [29,34,44,55,57]. In the Maltese Islands, this species is recorded from a specimen in the collection of the BMNH [11] collected on Limbarda crithmoides (Asteraceae). This species is known from the Ponto-Mediterranean area extending its distribution range to Kyrgyzstan and the Canary Islands [14].
- Thrips australis (Bagnall, 1915) †
- Material examined: MALTA: Wied Ħesri, 10.x.2015, 6 ♀♀ (sm) and 2 ♂♂ (sm) on Sambucus nigra, GD; Wied Ħesri, 16.iv.2016, 1 ♀ (sm), 5 ♀♀ (aga) and 1 ♂ (sm) on Schinus terebinthifolius, GD; Dingli Cliffs, 27.i.2017, 1 ♂ (sm) on Asphodelus ramosus, GD; Wied Musa l/o Siġġiewi, 30.x.2017, 1 ♀ (sm) on Smilax aspera, GD; Rabat road 2, 23.xi.2017, 2 ♀♀ (sm) on Ceratonia siliqua, GD; Luqa (by-pass), 14.i,2018, 1 ♀ (sm) on Solandra maxima, GD; Siġġiewi (private farm), 29.i.2018, 3 ♀♀ (sm) and 5 ♀♀ (aga) on Eriobotrya japonica, GD.
- Body length: ♀: 1330–1540 µm; ♂: 800–1260 µm.
- Wing type: Both sexes are macropterous.
Thrips australis is mostly, though not exclusively, found on Eucalyptus, and other Myrtaceae, but may disperse to other white flowering plants (e.g., Melaleuca sp.) once Eucalyptus flowers are no longer available [52]. This species has been also recorded on a number of agricultural crops [26], Juncus (Poaceae) [57], Lantana (Verbenaceae), Pentanisia ouranogyne (Rubiaceae) [26] and Acacia (Fabaceae) [26,50], but their presence on these species is probably accidental. In the Maltese Islands, this species was never collected on Eucalyptus flowers, but was always found on white flowers of unrelated trees and shrubs. An Australia species in origin, this species is now present wherever Eucalyptus plantations were introduced [53].
- Thrips major Uzel, 1895 †
- Material examined: MALTA: Wied Babu, 15.xii.1996, 2 ♀♀ (sm), on Erica multiflora, DM; Siġġiewi (private garden), 04.xi.2015, 4 ♀♀ (sm) on Rosa sp., GD; Siġġiewi (private garden), 12.i.2016, 1 ♀ (sm) on Rosa sp., GD; Siġġiewi (private garden), 01, 29.i.2016, 4 ♀♀ (sm) on Narcissus tazzetta, GD; Siġġiewi (private garden), 21.iii.2016, 2 ♀♀ (sm) on Pittosporum tobira, GD; Buskett, 19.iv.2016, 1 ♀ (sm) on Laurus nobilis, GD; Żabbar (private garden), 05.iv.2016, 1 ♀ (sm) on Stephanotis floribunda, GD; Wied Qirda, 15.iv.2016, 1 ♀ (sm) on Bromus madritensis, GD; Siġġiewi (road), 20.iv.2016, 3 ♀♀ (sm) on Olea europaea, GD; Wied Speranza, 26.iv.2016, 1 ♀ (sm) on Tropaeolum majus, GD; Siġġiewi (private garden), 10.v.2016, 2 ♀♀ (sm) on Lonicera caprifolium, GD; Siġġiewi, 21.viii.2016, 3 ♀♀ (sm) on Glebionis coronaria, GD; Msida, University of Malta grounds, 11.x.2016, 3 ♀♀ (sm) on Quercus robur, GD; Siġġiewi (private garden), 21.x.2016, 4 ♀♀ (sm) on Rosa sp., GD; Kunċizzjoni, 19.i.2017, 1 ♀ (sm) on Erica multiflora, GD; Dingli Cliffs, 27.i.2017, 2 ♀♀ (sm) on Asphodelus ramosus, GD; Msida, University of Malta grounds, 28.iv.2017, 2 ♀♀ (sm) on Olea europaea, GD; Luqa (by-pass), 14.i.2018, 1 ♀ (sm) on Solandra maxima, GD; Siġġiewi (private garden), 14.vii.2017, 1 ♀ (sm) on Azalea indica, GD; Siġġiewi (private farm), 29.i.2018, 4 ♀♀ (sm) on Eriobotrya japonica, GD; Buskett, 20.ii.2018, 1 ♀ (sm) on Rhamnus alaternus, GD; Wied Għollieqa, 11.iv.2018, 2 ♀♀ (sm) on Cercis siliquastrum, GD; Wied Għollieqa, 14.iv.2018, 1 ♀ (sm) on Cercis siliquastrum, GD; Għaxqet l-Għajn, l/o Naxxar, 28.iv.18, 2 ♀ (sm) on Pistacea lentiscus, GD; San Ġwann (private farm), 05.vii. 2019, 2 ♀♀ (sm) on Cucurbita sp., GD; Siġġiewi, (private garden), 22.iv.2020, 1 ♀ (sm) on Citrus limon, GD; Mġiebaħ l/o Xemxija, 2.iii.2018, 2 ♀♀ (sm) on Tamarix africana, GD; Siġġiewi(private garden), 22.iv.2020, 4 ♀♀ (sm) on Prunus persica, GD.
- Body length: ♀: 1300–1580 µm; ♂: no records.
- Wing type: ♀: macropterous; ♂: no records.
Thrips major is a polyphagous species, found on flowers and occasionally on leaf buds of unrelated plants [14,24,26,29,34,35,44,54,55,63]. In the Maltese Islands it was also collected on flowers and leaves of different crops, indigenous shrubs and trees belonging to the families Rosaceae, Amaryllidaceae, Pittosporaceae, Fabaceae, Tamaricaceae, Rhamnaceae, Poaceae and Lauraceae amongst others. T. major is distributed across the Palaearctic [14] and has been intercepted in the US from Europe, the Mediterranean basin and Africa [69].
- Thrips simplex (Morison, 1930) †
- Material examined: MALTA: 3.vi.1959, 1♀ (sm–BMNH) on Gladiolus sp., ERS; St. Paul’s Bay, 14.iv.1994, 1♀, Gladiolus sp., JWI;
- Body length: ♀: 1697 µm; ♂: no records.
- Wing type: ♀: macropterous; ♂: no records.
This species is associated with Gladiolus spp. (Iridaceae) [14,27,55,58], though it has also been recorded on Calla (Araceae), Pancratium (Amaryllidaceae), Protea (Proteaceae), Vitis (Vitaceae) [14] and Dianthus (Caryophyllaceae) [58]. In the Maltese Islands, Thrips simplex was only found on Gladiolus (Iridaceae). This species originated from southern Africa [27], but has become cosmopolitan in distribution and occurs wherever Gladiolus is cultivated [14].
- Thrips tabaci Lindeman, 1889
- Material examined: MALTA: St. Paul’s Bay, vii.1956, 1 ♀ (sm—BMNH) on Limbarda crithmoides (JIS & ERS); Żabbar, 11.iii.1994, 2 ♀♀ (sm—BMNH) on Matthiola bicornis, GW; Armier, 14.vi.1994, 1 ♀ (sm—NMNH) on Fragraria x ananassa, GW; Lapsi, 26.i.1997, 1 ♀ (dark morph) on Asphodelus ramosus, DM; Siġġiewi (private garden), 21.iii.2016, 1 ♀ (dark morph) on Pittosporum tobira, GD; Żabbar (private garden), 05.iv.2016, 1 ♀ (dark morph) on Stephanotis floribunda, GD; Siġġiewi (private garden), 13.iv.2016, 2 ♀♀ (dark morph—sm) on Foeniculum vulgare, GD; Wied Ħesri, 16.iv.2016, 1 ♀ (dark morph) on Schinus terebinthifolius, GD; Dingli, Buskett, 19.iv.2016, 1 ♀ (pale form—sm) on Laurus nobilis, GD; Wied Speranza, 26.iv.2016, 2 ♀♀ (dark morph—sm) on Tropaeolum majus, and 3 ♀♀ (dark morph—sm) on Parietaria judaica, GD; Siġġiewi (private garden), 10.v.2016, 1 ♀ (dark morph—sm) on Prunus persica, GD; Wied Ħesri, 15.v.2016, 2 ♀♀ (pale morph–sm) on Hypericum sinuatum, GD; Wied Qirda, 30.v.2016, 1 ♀ on Hyparrhenia hirta, GD; Msida, University of Malta grounds, 11.x.2016, 1 ♀ (dark morph—sm) on Quercus robur, GD; Dingli Cliffs, 27.i.2017, 1 ♀ (dark morph—sm) on Asphodelus ramosus, GD; Fiddien, 24.iv.2017, 2 ♀♀ (dark morph—sm) on Medicago sp., GD; Popeye Village, 29.vii.2017, 1 ♀ (pale morph–sm) on Tamarix africana, GD; Siġġiewi, (road l/o Wied Ħesri), 07.viii.2017, 3 ♀♀ (pale morph–sm) on Ipomoea carnosa, GD; Siġġiewi (private farm), 13.xi.2017, 1 ♀ (pale form–sm) on Eriobotrya japonica, GD; Xemxija, 03.iii.2018, 7 ♀♀ (dark morph—sm) on Anacamptis urvilleana, GD; Qormi (private farm), 7.vii.2018, 1 ♀ (pale morph–sm), 3 ♀♀ (dark morph—sm) on Foeniculum vulgare, GD; Pembroke, 05.xi.2018, 1 ♀ (dark morph—sm) on Reichardia picroides, GD. GOZO: Ramla Bay, 02.vii.2018, 2 ♀♀ (pale form–sm) on Pancratium maritimum, GD.
- Body length: ♀: 1060–1360 µm; ♂: no records.
- Wing type: ♀: macropterous; ♂: no records.
Both the pale and the dark morphs of this species were recorded in the Maltese Islands, with the pale morphs being more common in the warmer months. Thrips tabaci is a polyphagous species, being recorded on numerous unrelated plants, particularly Asteraceae [24], but also on a wide variety of crops, cultivars and indigenous plants [27,34,35,36,54,55,57,58,63,70]. In the Maltese Islands, this species has been originally reported on Allium (Amaryllidaceae) [7], though in the current study, it has been found on a number of indigenous and cultivated herbaceous annuals and trees including species of Brassicaceae, Rosaceae, Tropaeolaceae, Poaceae, Asteraceae, Apiaceae, Orchidaceae, Amaryllidaceae, Convolvulaceae and Fabaceae amongst others. T. tabaci tends to perform thelytokous reproduction, therefore most individuals are female [71] while males are always very rare. This accounts for the fact that in the current study, no male specimens were found despite the large number of specimens collected. T. tabaci is a widely distributed pest, causing both direct damage to agricultural crops as well as acting as a vector of orthotospoviruses [14,26] but it is also recorded as an important pollinator [56]. T. tabaci is probably of Eastern Mediterranean origin [14] but is now cosmopolitan [14,27] and present wherever onion and garlic are grown [27].
4. Discussion
The insect fauna of the Maltese Islands has been well studied, with works covering practically all orders [72,73,74]. The order Thysanoptera is however an exception, owing to the fact that insects belonging to this order are small, difficult to identify and few people are interested to study them. As stated in the results section, the current study has recorded 39 species of Terebrantia from the Maltese Islands. This species richness compares well to other Mediterranean islands such as Sicily with 58 [13], Sardinia with 29 [13] and the Canary Islands in the Atlantic ocean with 75 species [75], despite the fact that all of these islands are considerably larger than the Maltese Islands and feature a larger variety of habitats.
Thirty eight of the identified species are phytophagous, with the four species of the genus Aeolothrips, as well as Frankliniella occidentalis and Thrips tabaci being described as facultative predators. The only terebrantian recorded in the Maltese Islands which is an obligate predator is Franklinothrips megalops.
An analysis of the zoogeographical composition of the thrips fauna based on distributional types (chorotypes) as defined by Vigna Taglianti et al. [76] (Table 2, Table A1 in Appendix A) reveals that the majority of the species (26 species–66.7%) come from Europe and the Mediterranean Region, with 12 (30.7%) of these species extending their geographical distribution towards the Middle East and the Asian continent and four species (10.2%) reaching the African continent.
These findings reflect the geographical locations of the Maltese Islands, which are located in the center of the Mediterranean Sea, around 250 km north of the Libyan coast and 90 km south of Sicily. Previous local studies on insects with the approximate size and mobility of thrips [77] indicate that most of the local fauna is of European origin whereas African representation is very limited. This also seems to be the case for the terebrantian thrips. The limited amount of habitats found on the Maltese Islands may also have contributed to the fact that no endemic species of Terebrantia were recorded in this study.
Twelve (30.7%) of the species occurring in the Maltese Islands have a cosmopolitan or subcosmopolitan distribution. All of these species, with the exception of Limothrips angulicornis, are indicated as agriculturally important [26,32] and are capable of potentially damaging crops and cultivated plants by either feeding directly on plant tissues or by transmitting disease agents, including orthotospoviruses. It is highly likely that there is a link between the chorotypes of these species and the fact that these species are widely indicated as polyphagous (since larval forms were recorded on a large variety of unrelated plants). Being able to use a wider range of plant species as hosts increases the chances of settling into new environments.
Seven (17.9%) of the species with a cosmopolitan or subcosmopolitan distribution were found to be of alien origin. These species include Heliothrips haemorrhoidalis, Hercinothrips femoralis, Echinothrips americanus, Frankliniella occidentalis, F. schultzei, Thrips australis and T. simplex. The geographical distribution of these seven species either extends from distant world regions into the Mediterranean, or else is found in different unconnected world regions since they were imported from their countries of origin to other areas together with crops or cultivated plants. During the current study, the above-listed species, with the exception of Thrips australis, were collected from cultivated fields, greenhouses or gardens, on crops or cultivated plants and were absent in indigenous habitats situated close to the locations where these species were collected from. It is interesting to note that these species were generally not collected in numbers large enough to suggest that these can act as pests in the Maltese Islands, though some of the species such as Echinothrips americanus were observed to have the potential of quickly establishing sizeable populations that can quickly decimate cultivated plants given the right environmental conditions. Species such as Thrips australis may have occurred in the Maltese Islands for quite some time. T. australis is closely related to Eucalyptus trees and may therefore have been present in the Maltese Islands as far back as the 1950s, when fast growing trees, including Eucalyptus gomphocephala, were imported from different parts of the world in an attempt to increase the local tree populations in urban environments.
In the present study, a number of specimens were also collected which could only be identified to genus level. These specimens belonged to the genera Chirothrips (1 species), Odontothrips (2 species), Tenothrips (1 species) and Thrips (2 species) and additional material will be required to properly identify them in future.
In due course, the thrips fauna of the Maltese Islands may change due to a number of factors, namely building over natural habitats due to urbanization requirements and the extensive use of plant protection products in agricultural fields. Another important contributing factor is climate change, with the weather of the Maltese Islands becoming hotter and drier. This can affect the distribution of certain indigenous plants that serve as hosts or even as food or shelter plants and consequently impact the thrips diversity. Importation of cultivated plants will continue to introduce additional alien species to the Maltese Islands. Without proper quarantine monitoring of imported plant material and the devising of eradication policies, these species can definitely become established in the Maltese Islands in the very near future.
Author Contributions
Conceptualization, D.M. and G.D.; methodology, D.M. and G.D.; validation, A.C., A.G. and D.M.; formal analysis, G.D.; investigation, G.D.; resources, G.D.; data curation, G.D.; writing—original draft preparation, G.D.; writing—review and editing, D.M., A.C. and A.G.; visualization, D.M.; supervision, D.M., A.C. and A.G.; project administration, D.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to acknowledge Laurence A. Mound from CSIRO for the identification of A. gloriosus and R. niveipennis and G. (Bert) Vierbergen from the Netherlands Institute for Vectors, Invasive plants and Plant health, for providing information about Tenothrips discolor. Thanks also go to Paul Brown from the BMNH who kindly lent us specimens of Chirothrips hamatus and Thrips simplex, as well as to Edwin Lanfranco for the help in identifying some of the plants on which the thrips specimens were collected.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A

{kind=link}
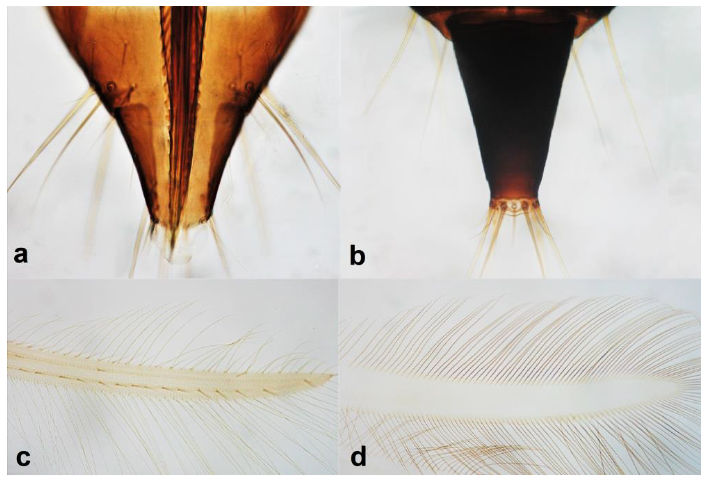{kind=link}
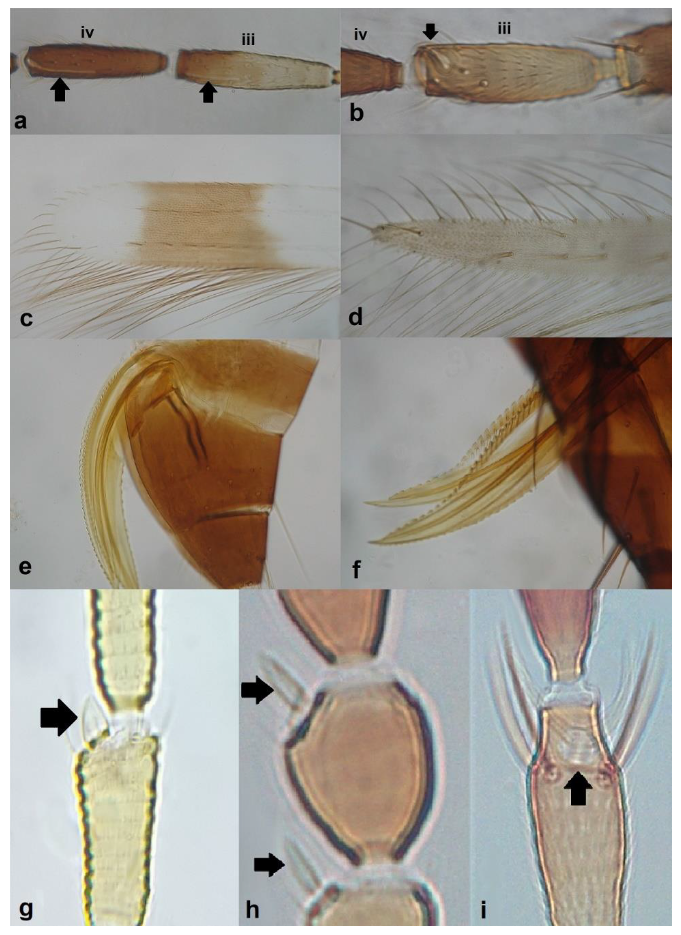{kind=link}
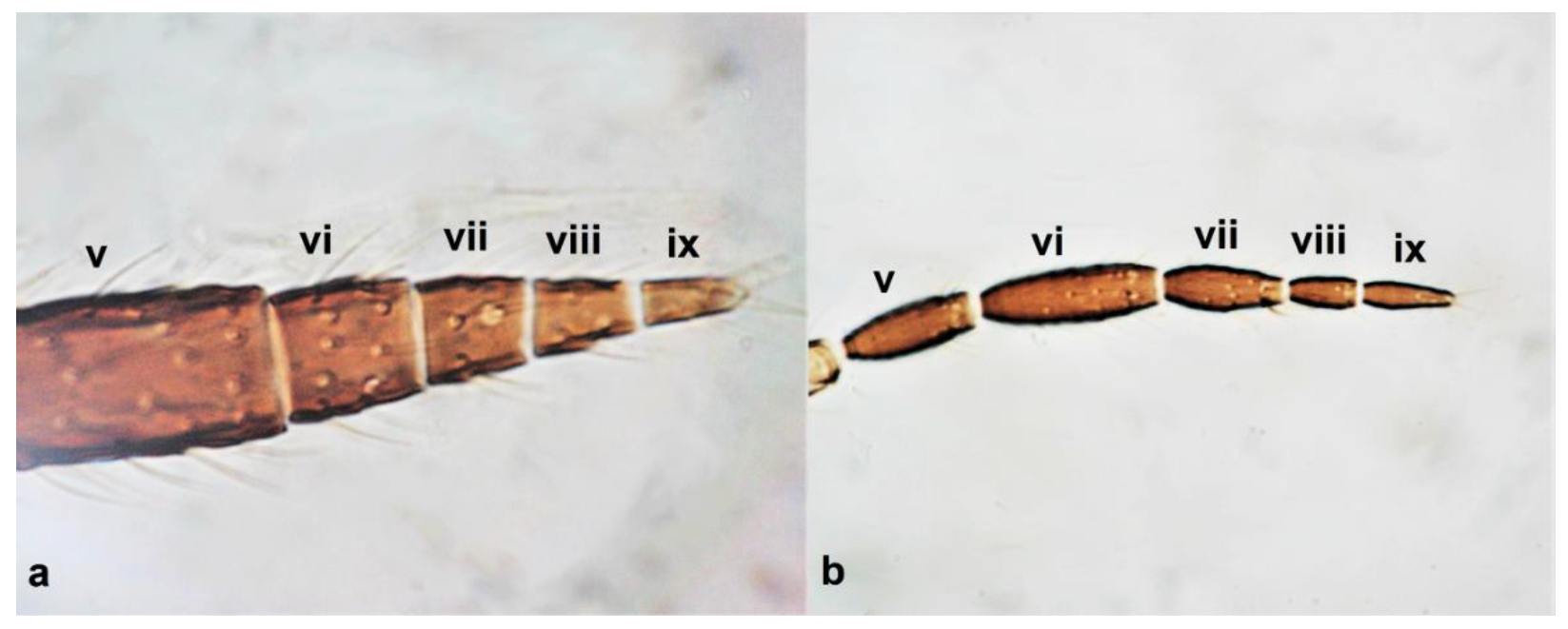{kind=link}
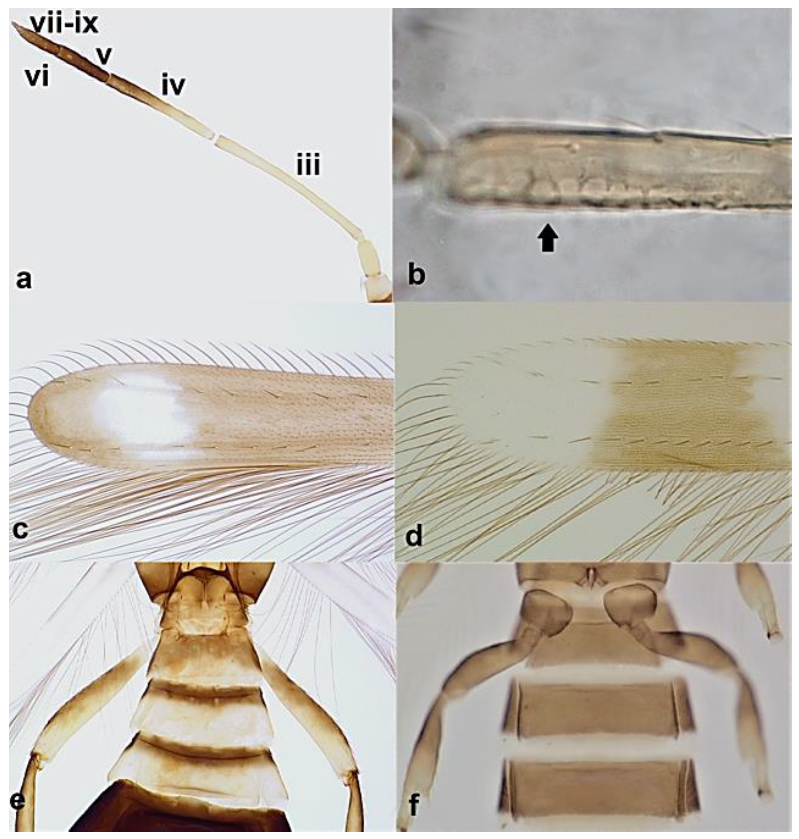{kind=link}
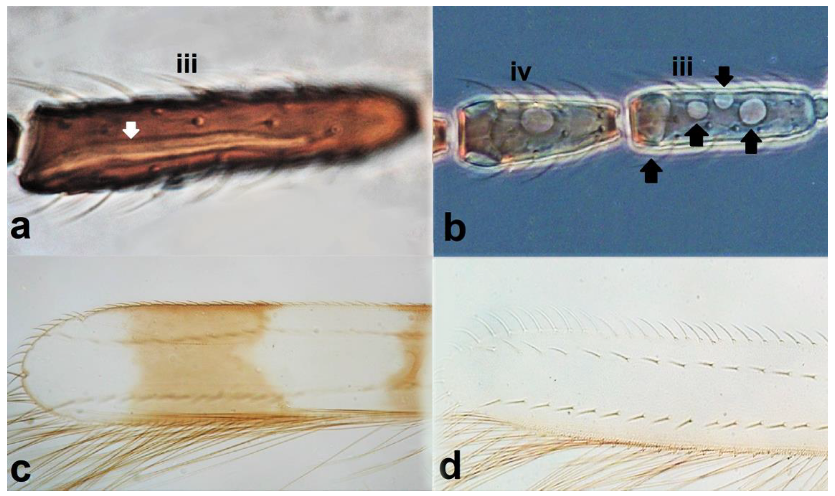{kind=link}
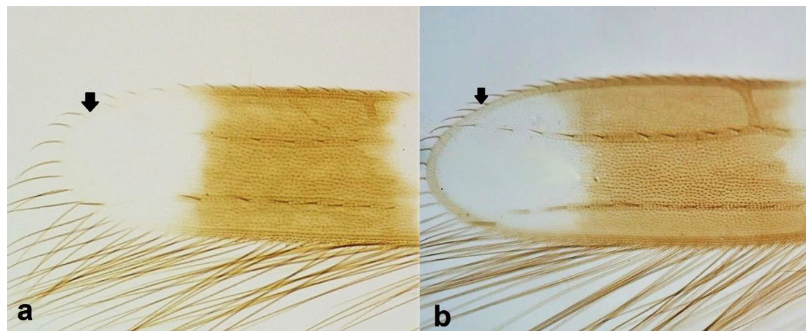{kind=link}
{kind=link}
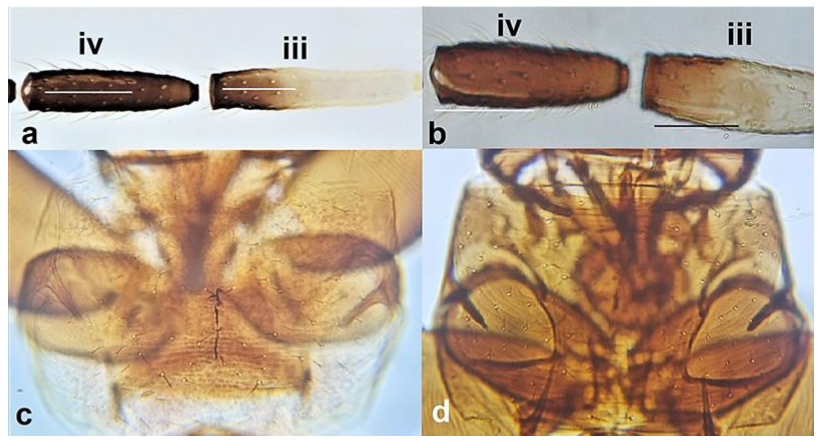{kind=link}
{kind=link}
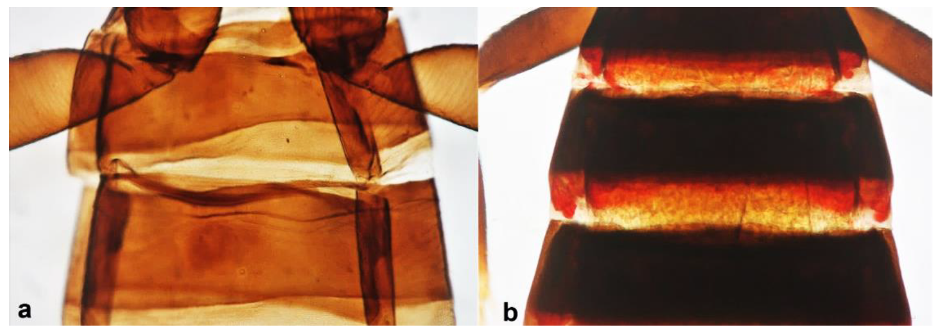{kind=link}
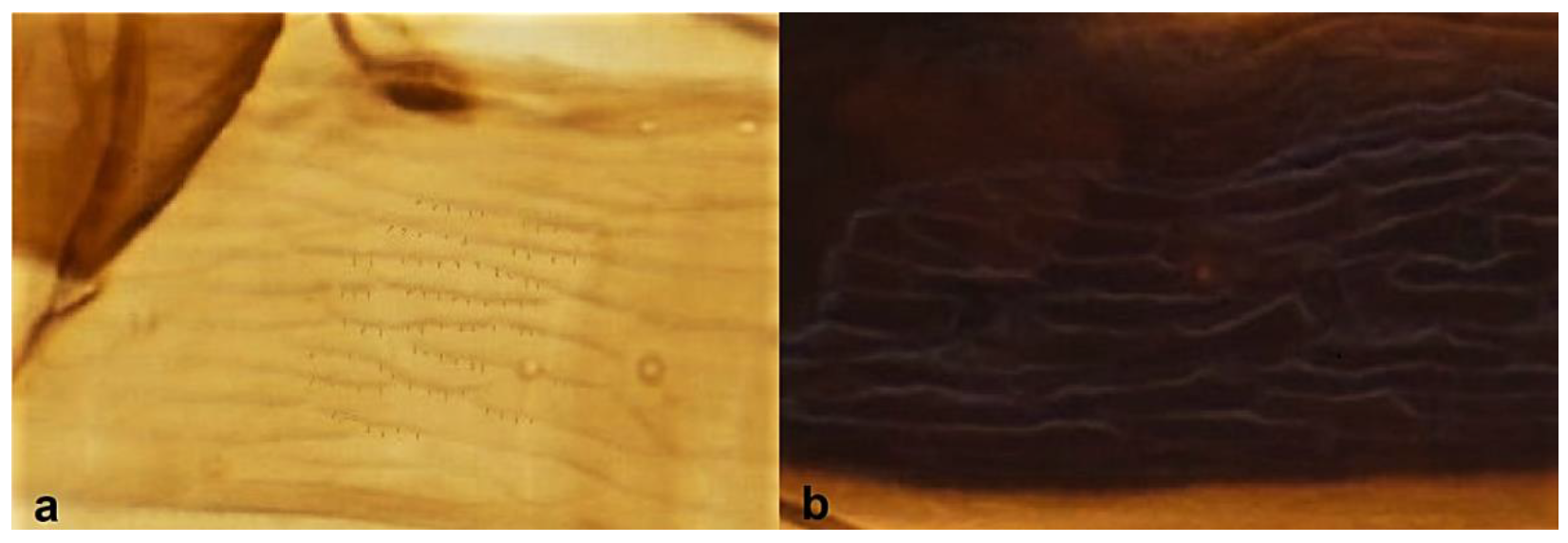{kind=link}
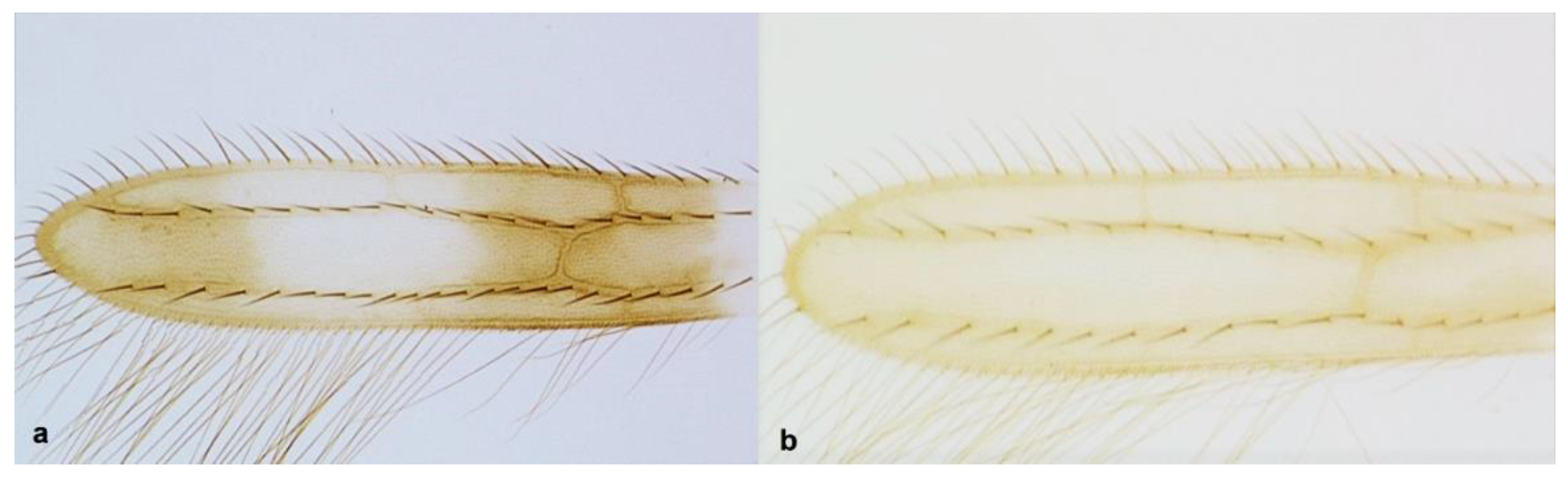{kind=link}
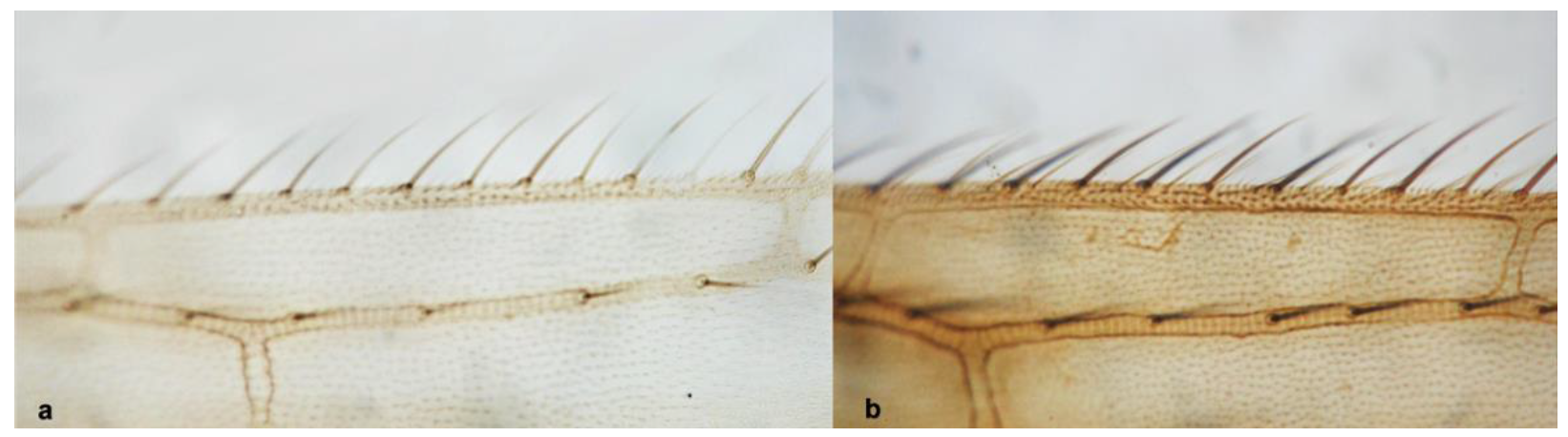{kind=link}
{kind=link}
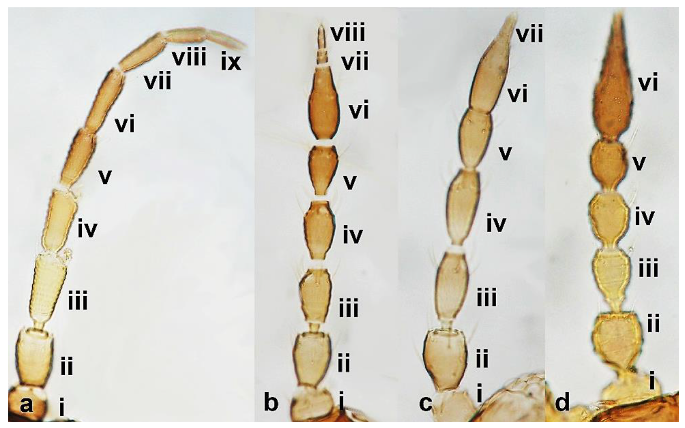{kind=link}
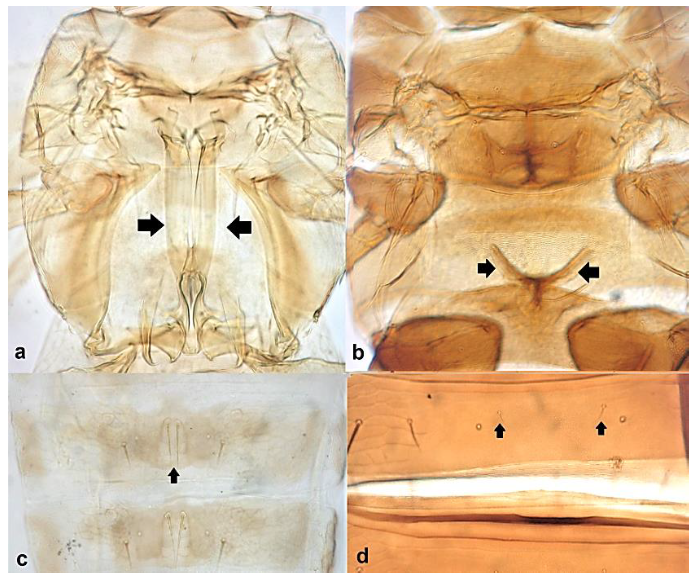{kind=link}
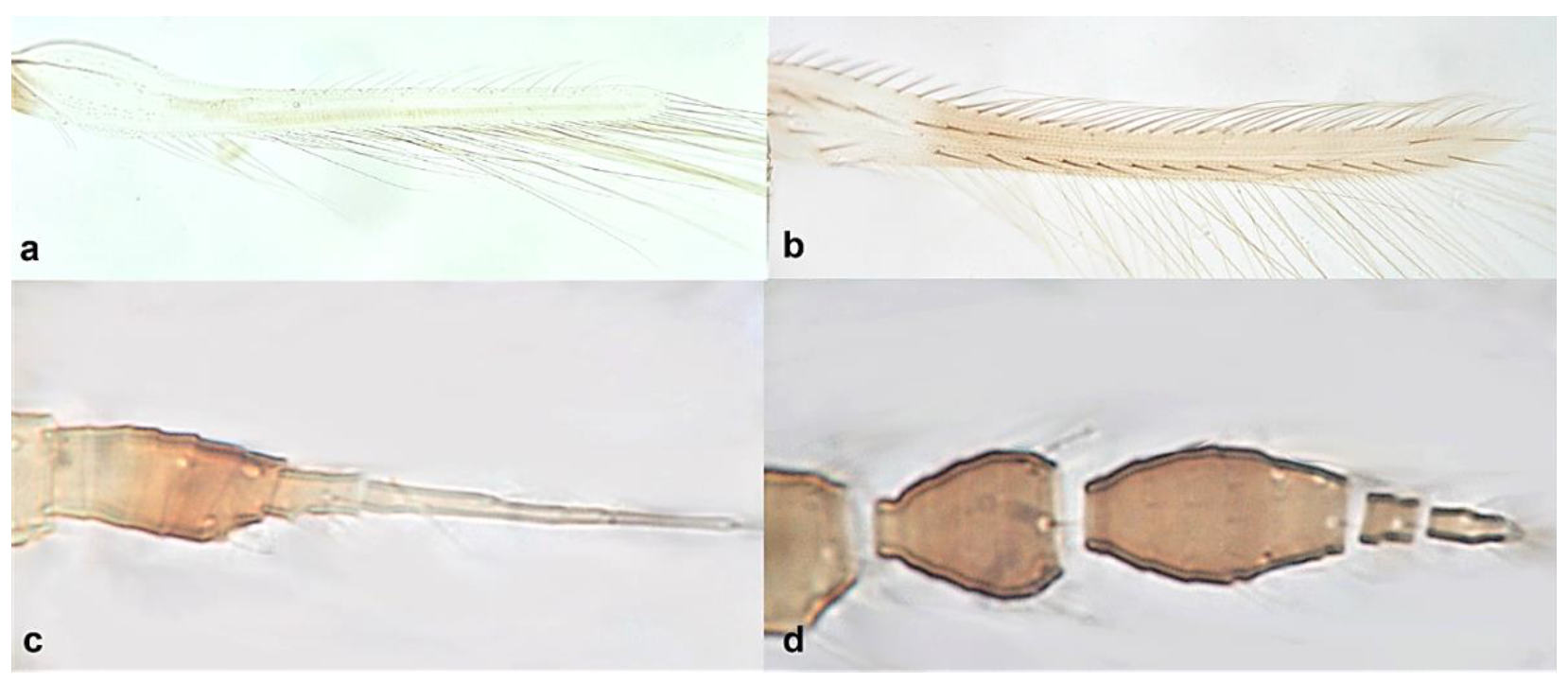{kind=link}
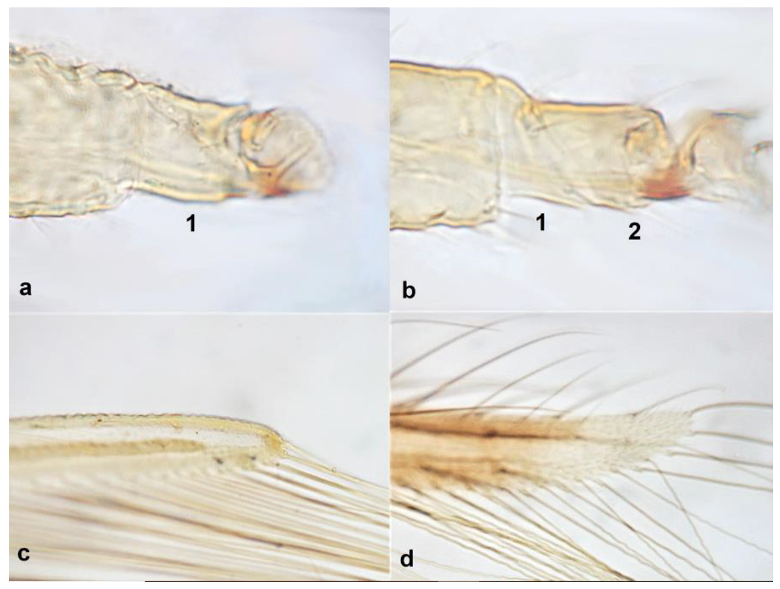{kind=link}
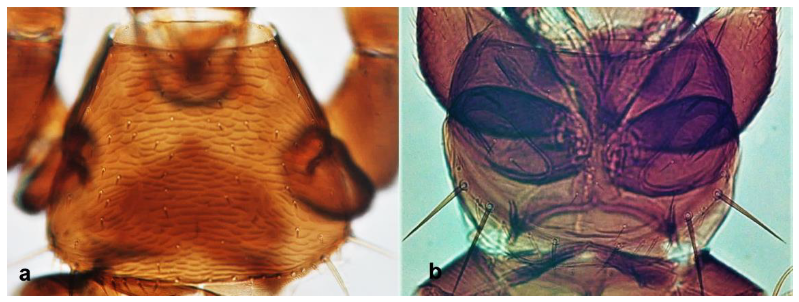{kind=link}
{kind=link}
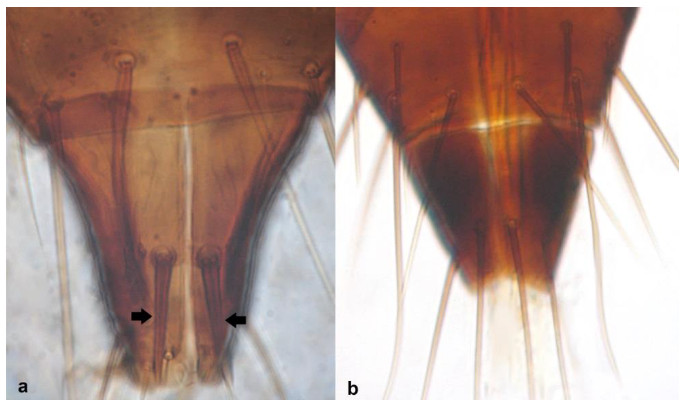{kind=link}
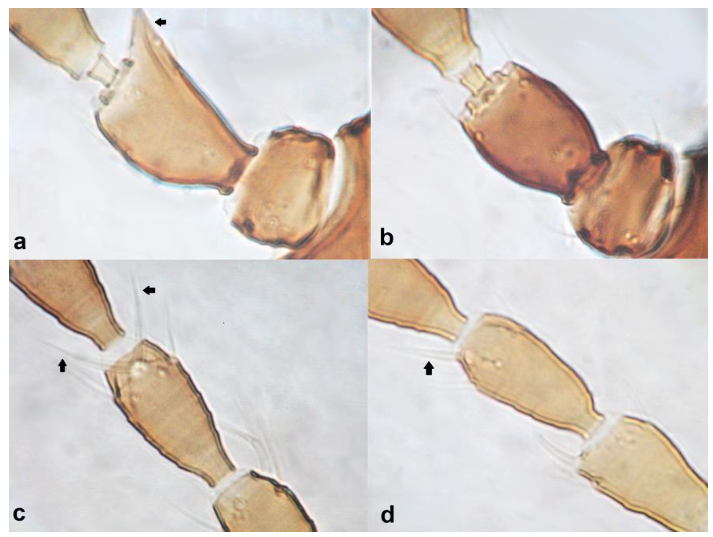{kind=link}
{kind=link}
{kind=link}
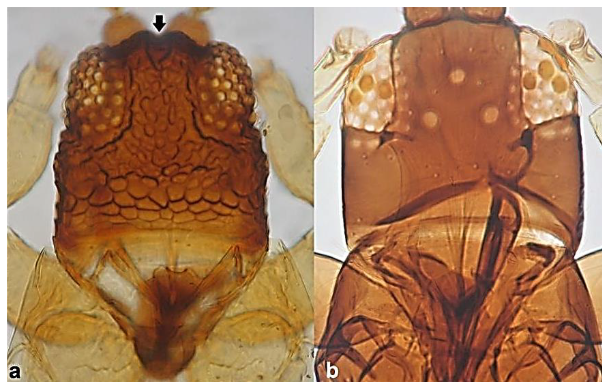{kind=link}
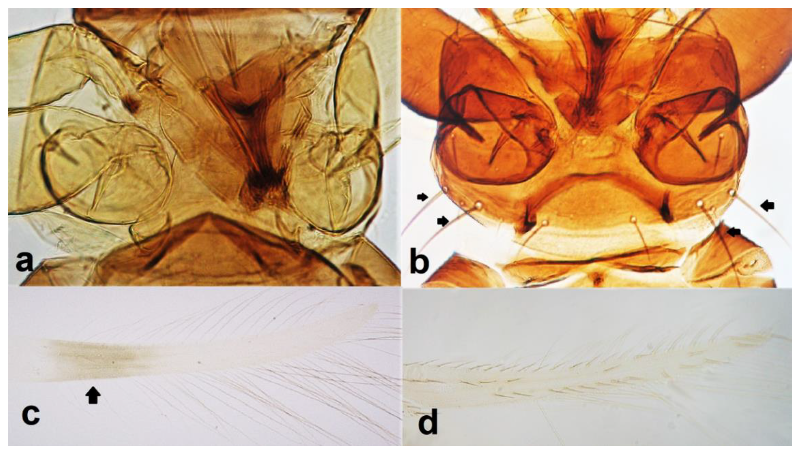{kind=link}
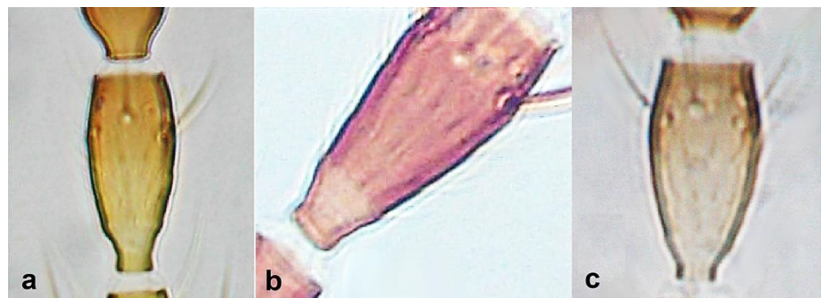{kind=link}
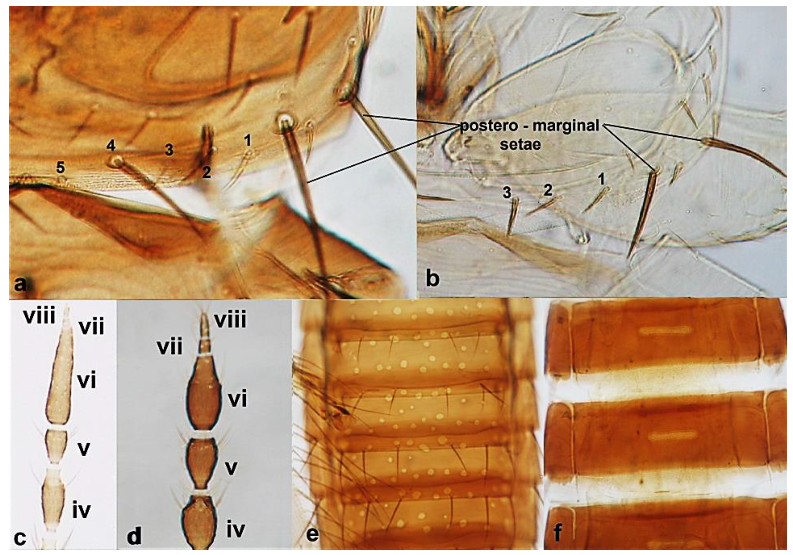{kind=link}
{kind=link}
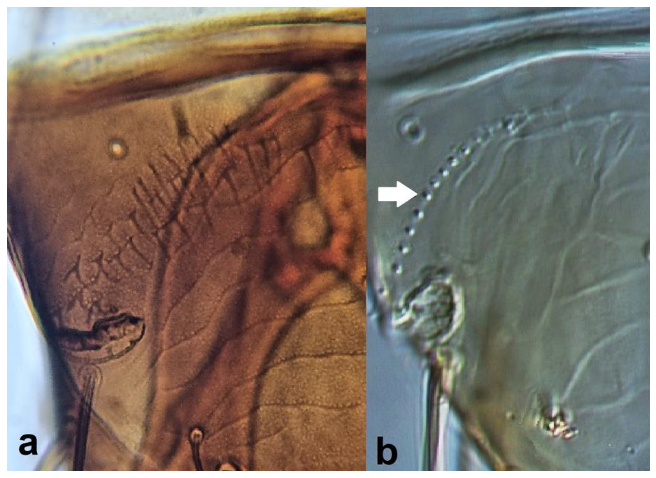{kind=link}
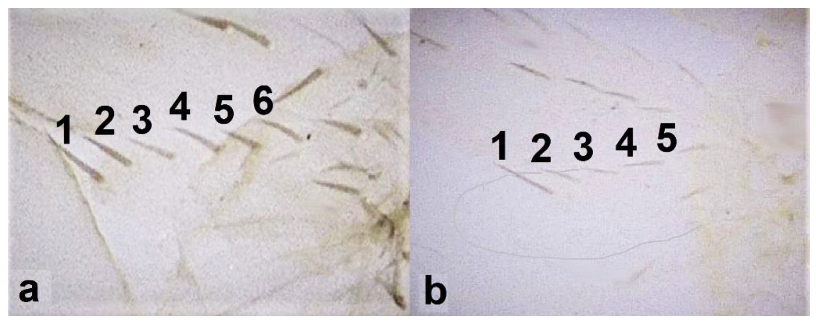{kind=link}
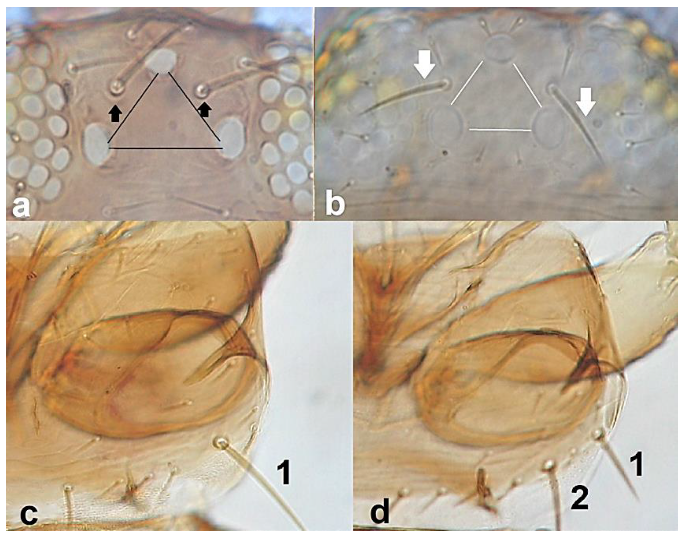{kind=link}
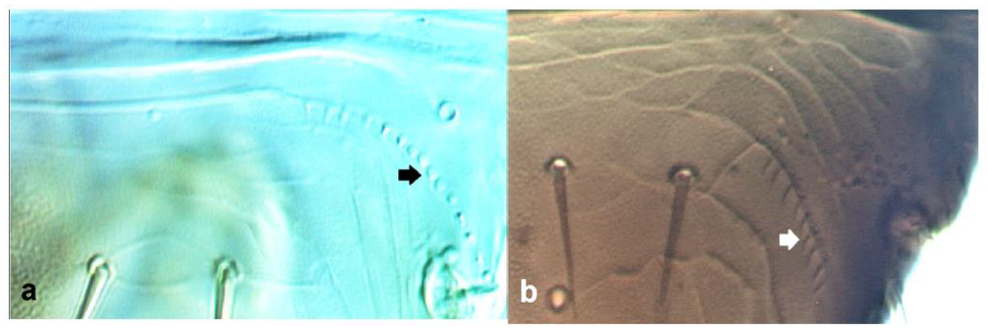{kind=link}
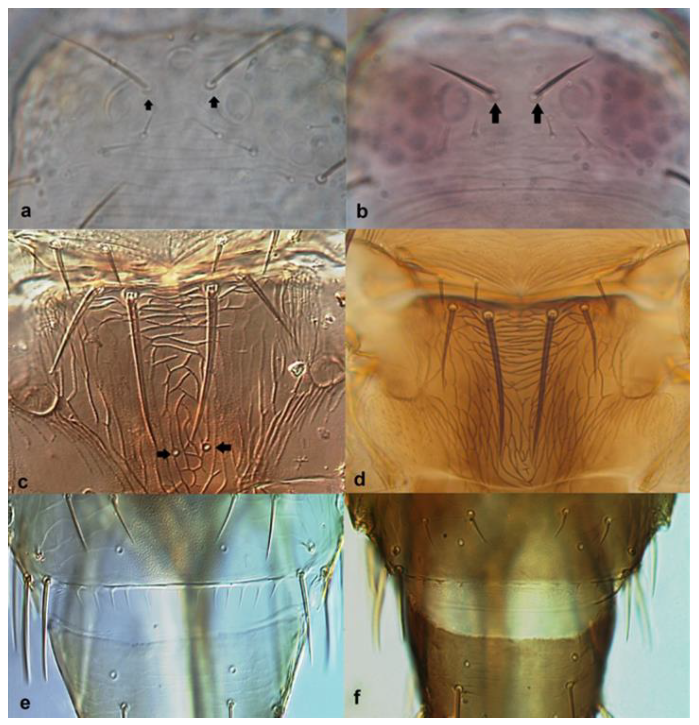{kind=link}
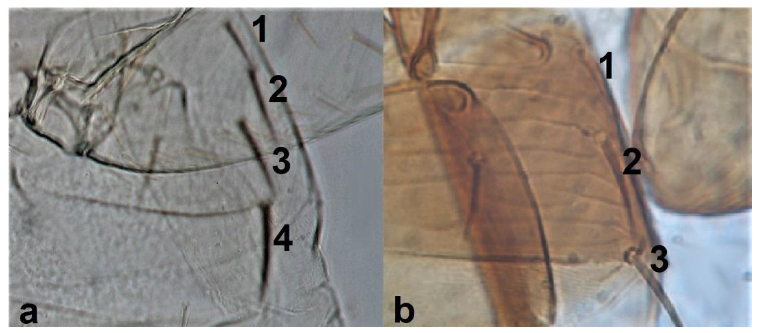{kind=link}
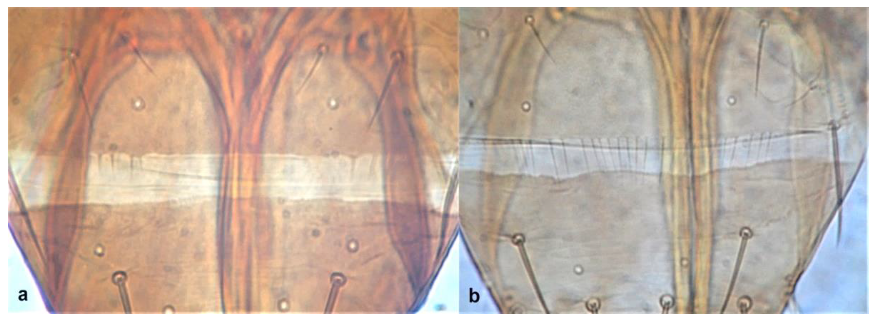{kind=link}
Table A1.
Chorotypes for the Thysanoptera of the Maltese Islands (based on data from previously published material).
Table A1.
Chorotypes for the Thysanoptera of the Maltese Islands (based on data from previously published material).
| Species | Chorotype | Notes |
|---|---|---|
| Aeolothrips gloriosus | West Palaearctic | |
| Aeolothrips imtermedius | Asiatic European | |
| Aeolothrips melisi | Mediterranean | |
| Aeolothrips tenuicornis | Turano Mediterranean | |
| Franklinothrips megalops | Afrotropico Mediterranean | |
| Rhipidothrips brunneus | Palaearctic | also Australian region |
| Rhipidothrips gratiosus | Palaearctic | |
| Rhipidothrips niveipennis | European | |
| Rhipidothrips unicolor | Turano Mediterranean | |
| Melanthrips ficalbii | Palaearctic | |
| Melanthrips fuscus | Palaearctic | |
| Melanthrips knechteli | Turano European | |
| Melanthrips lybicus | Mediterranean | |
| Holarthropthips tenuicornis | Mediterranean | |
| Dendrothrips saltator | Asiatic European | also Oriental region |
| Heliothrips haemorrhoidalis | Cosmopolitan | |
| Hercinothrips femoralis | Cosmoplolitan | |
| Anaphothrips sudanensis | Subcosmopolitan | |
| Aptinothrips rufus | Cosmopolitan | |
| Asphodelothrips croceicollis | Turano Europo Mediterranean | |
| Bregmatothrips dimorphus | Afrotropico Mediterranean | also East Palaearctic |
| Ceratothrips ericae | Asiatic European | also Australian region |
| Chirothrips hamatus | Sibero European | Also Great Britain. Does not extend beyond Turkey |
| Chirothrips manicatus | Subcosmopolitan | |
| Chirothrips meridionalis | Afrotropico Indo Mediterranean | |
| Echinothrips americanus | Subcosmopolitan | |
| Franklinella occidentalis | Cosmopolitan | |
| Frankliniella schultzei | Subcosmopolitan | |
| Limothrips angulicornis | Subcosmopolitan | |
| Limothrips cerealium | Subcosmopolitan | |
| Odontothrips meliloti | Turano European | |
| Oxythrips ajugae | Holarctic | |
| Pezothrips kellyanus | Turano Mediterranean | also the Netherlands and the Australian region |
| Prosopothrips nigriceps | Turano Mediterranean | |
| Tenothrips discolor | Turano Europeo Mediterranean | |
| Thrips australis | Afrotropico Mediterranean | also Australian region |
| Thrips major | Palaearctic | |
| Thrips simplex | Subcosmopolitan | |
| Thrips tabaci | Cosmopolitan |
References
- Mound, L.A.; Wang, Z.; Lima, É.F.B.; Marullo, R. Problems with the Concept of “Pest” among the Diversity of Pestiferous Thrips. Insects 2022, 13, 61. [Google Scholar] [CrossRef] [PubMed]
- Burckhardt, D.; Ouvrard, D.; Queiroz, D.; Percy, D. Psyllid Host-Plants (Hemiptera: Psylloidea): Resolving a Semantic Problem. Fla. Entomol. 2014, 97, 242–246. [Google Scholar] [CrossRef]
- Pedley, M.; Hughes Clarke, M.; Galea, P. Limestone Isles in a Crystal Sea; PEG: San Gwann, Malta, 2002; p. 109. [Google Scholar]
- Chetcuti, D.; Buhagiar, A.; Schembri, P.J.; Ventura, F. The Climate of the Maltese Islands; Malta University Press: Msida, Malta, 1992; p. 108. [Google Scholar]
- FAO. Malta Water Resources Review; FAO: Rome, Italy, 2006. [Google Scholar]
- MEPA Terrestral Habitats. Available online: http://www.mepa.org.mt/biodiversity-habitats-terrestrial##8 (accessed on 25 February 2015).
- Mifsud, S. Malta Wild Plants. Available online: www.maltawildplants.com (accessed on 14 July 2018).
- Saliba, L.J. Insect Pests of Crop Plants in the Maltese Islands; Department of Information: Msida, Malta, 1963. [Google Scholar]
- Farrugia, C. Insect Pests on Cauliflower (Brassica oleracea var. botrytis) in Gozo (Maltese Islands, Central Mediterranean); University of Malta: Msida, Malta, 1997. [Google Scholar]
- Mifsud, D. Biological control in the Maltese Islands—Past initiatives and future programmes. Bull. OEPP 1997, 27, 77–84. [Google Scholar] [CrossRef]
- Mound, L.A.; Palmer, J.M. Notes on Thysanoptera from Israel. Entomol. Mon. Mag. 1973, 109, 102–106. [Google Scholar]
- Mifsud, D.; Watson, G.W. Introduced Sap-Feeding Insect Pests of Crop Plants in the Maltese Islands; University of Malta: Msida, Malta, 1999. [Google Scholar]
- Vierbergen, G. Fauna Europaea—All European animal species on the web: Thysanoptera Section. Biodivers. Data J. 2014, 2, e4034. [Google Scholar]
- Zur Strassen, R. Die Terebrantien Thysanopteren Europas; Die Tierwelt Deutschlands; Goecke & Evers: Keltern, Germany, 2003; p. 277. [Google Scholar]
- Zur Strassen, R.; Kuslitzky, W. An annotated checklist of the thrips of Israel (Thysanoptera). Isr. J. Entomol. 2012, 41, 53–66. [Google Scholar]
- National Statistics Office. Census of Population and Housing 2021; Preliminary Report; National Statistics Office: Valletta, Malta, 2021; p. 100. [Google Scholar]
- Mound, L.A.; Kibby, G. Thysanoptera; CAB International: Wallingford, UK, 1998; p. 70. [Google Scholar]
- Palmer, J.M.; Du Heaume, G.J.; Betts, C. 1959 CIE Guides to Insects of Importance to Man. 2, Thysanoptera. Betts, C.R., Ed.; CAB International: Wallingford, UK, 1989; p. 73. [Google Scholar]
- Marullo, R. I Tisanotteri dell’ Italia meridionale. II Contributo. Le specie italiane del genere Aeolothrips Haliday. Boll. Lab. Entomol. Agrar. Filippo Silvestri 1993, 50, 121–140. [Google Scholar]
- Marullo, R. I Tisanotteri dell’Italia meridionale. III Contributo. La Collezione del Museo Civico di Storia Naturale “G. Doria” di Genova. Boll. Lab. Entomol. Agrar. Filippo Silvestri 1996, 51, 57–65. [Google Scholar]
- Marullo, R. Thysanoptera of southern Italy. V Contribution. Morphological remarks and biological notes on some southern Mediterranean species. Boll. Lab. Entomol. Agrar. Filippo Silvestri Portici 2004, 59, 49–57. [Google Scholar]
- Priesner, H. A Monograph of the Thysanoptera of the Egyptian Deserts; Desert Institute: El Mataria, Egypt, 1960; p. 582. [Google Scholar]
- Priesner, H. Ordnung Thysanoptera (Fransenfluger Thripse); Academie-Verlag: Berlin, Germany, 1964; p. 242. [Google Scholar]
- Mound, L.A.; Morrison, G.D.; Pitkin, B.R.; Palmer, J.M. Thysanoptera. Handbooks for the Identification of British Insects; Royal Entomological Society of London: London, UK, 1976; Volume 1, p. 79. [Google Scholar]
- Zhang, S.M.; Mound, L.A.; Hastings, A. Thysanoptera Chinensis—Thripidae Genera from China. Available online: https://keys.lucidcentral.org/keys/v3/thysanoptera_chinensis/ (accessed on 9 May 2018).
- Moritz, G.; Brandt, S.; Sseruwagi, P.; Waiganjo, M.; Subramanian, S. Pest thrips of Eastern Africa—Identification and Information based on LucID 3.5. Mitt. Dtsch. Ges. Allg. Angew. Entomol. 2012, 18, 533–539. [Google Scholar]
- Mound, L.A.; Collins, D.W.; Hastings, A. Thysanoptera Britannica et Hibernica—Thrips of the British Isles. Available online: https://keys.lucidcentral.org/keys/v3/british_thrips/.html (accessed on 9 May 2018).
- Mound, L.A.; Hoddle, M.S.; Hastings, A. Thysanoptera Californica—Thrips of California. Available online: https://keys.lucidcentral.org/keys/v3/thrips_of_california_2019/index.html (accessed on 26 April 2019).
- Marullo, R.; De Grazia, A. Territorial distribution, classification and relationships amongst Italian Thysanoptera. Bull. Insectology 2013, 66, 127–134. [Google Scholar]
- Bhatti, J.S.; Alavi, J.; zur Strassen, R. Thysanoptera in Iran 1938–2007: An Overview. Thrips 2009, 7–8, 373. [Google Scholar]
- Canale, A.; Conti, B.; Petacchi, R.; Rizzi, I. Thysanoptera collected in an olive growing area of the northern Tuscany (Italy). Entomol. Probl. 2003, 33, 105–110. [Google Scholar]
- Moritz, G. Thripse; Meiling Druck: Haldensleben, Germany, 2006; p. 384. [Google Scholar]
- Kirk, W.D.J. Thrips. Naturalist’s Handbook No 25; Richmond Publishing Co., Ltd.: Richmond, UK, 1996; p. 70. [Google Scholar]
- Fallahzedah, M.; Elaheh, A.; Nazila, S.; Hassan, A.; Jalil, A. Faunistic survey of Thysanoptera in Fars province, Iran. Munis Entomol. Zool. 2011, 6, 251–261. [Google Scholar]
- Nickle, D. A Checklist of Commonly Intercepted Thrips (Thysanoptera) from Europe, the Mediterranean, and Africa at U.S. Ports-of Entry (1983–1999). Part 1. Key to Genera. In Proceedings of the Entomological Society of Washington, Washington, DC, USA, 2 May 2003; Volume 105, pp. 80–99. [Google Scholar]
- Tunç, İ.; Bahşi, Ş.Ü.; Göçmen, H. Thysanoptera fauna of the Aegean region, Turkey, in the spring. Turk. J. Zool. 2012, 36, 592–606. [Google Scholar] [CrossRef]
- Mound, L.A.; Reynaud, P. Franklinothrips; a pantropical Thysanoptera genus of ant-mimicking obligate predators (Aeolothripidae). Zootaxa 2005, 864, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Alavi, J.; zur Strassen, R.; Bagherani, N. Thrips (Thysanoptera) species associated with wheat and barley in Golestan province, Iran. J. Entomol. Soc. Iran 2007, 27, 1–28. [Google Scholar]
- Mirab-balou, M.; Chen, X.; Tong, X. Tenothrips bhatti, a newly recorded genus of Thripinae (Thysanoptera: Thripidae) from China. Entomotaxonomia 2012, 34, 162–166. [Google Scholar]
- Razi, S.; Laamari, M. Thysanoptera survey on Vicia faba (broad bean) in the arid Biskra region of Algeria. Agric. Biol. J. North Am. 2013, 4, 268–274. [Google Scholar] [CrossRef]
- Gertsson, C. An annotated checklist of Thysanoptera (thrips) from the Nordic countries. [Provinsförteckning över Nordens tripsar.]. Entomol. Tidskr. 2015, 134, 185–198. [Google Scholar]
- Marullo, R. Un Profilo della Tisanotterofauna italiana: Le Interazioni tra Specie e Piante Ospiti e gli Effetti delle Colture sulla Diversita’ delle Popolazioni Naturali; Congresso Nazionale Italiano di Entomologia: Rome, Italy, 2002; pp. 179–184. [Google Scholar]
- Stoch, F. Fauna Italica. Available online: http://www.faunaitalia.it/ (accessed on 20 February 2015).
- Zur Strassen, R. Thysanoptera on islands of the northern Sporades in the Aegean (Greece) (Insecta: Thysanoptera). Senckenberg. Biol. 1986, 67, 85–129. [Google Scholar]
- Tunc, I. Studies on the Thysanoptera of Antalya I. Aeolothripidae Uzel. Türk. Entomoloji Derg. 1991, 15, 129–141. [Google Scholar]
- Sierka, W.; Fedor, P.; Vasiliu-Oromulu, L.; Jenser, G.; Bărbuceanu, D. The state of knowledge of thrips (Insecta: Thysanoptera) of the Carpathian Mountains. Acta Phytopathol. Entomol. Hung. 2008, 43, 355–366. [Google Scholar] [CrossRef]
- Karadjova, O.; Krumov, V. Thysanoptera of Bulgaria. ZooKeys 2015, 504, 93–131. [Google Scholar]
- Marullo, R. Conoscere i Tisanotteri; Edizioni Agricole de Il Sole 24 ore Edagricole S.r.l. via Goito 13—40126; Università degli Studi Mediterranea di Reggio Calabria: Reggio Calabria, Italy, 2003; p. 75. [Google Scholar]
- ThripsWiki Contributors zur Strassen Distribution Lists. Available online: http://thrips.info/w/index.php?title=Zur_Strassen_distribution_lists&oldid=41628 (accessed on 6 December 2016).
- ThripsWiki. ThripsWiki—Providing Information on the World’s Thrips in the Catalogue of Life; ThripsWiki: Leiden, The Netherlands, 2018. [Google Scholar]
- Nakahara, S.; O’donnell, C.A.; Mound, L.A. Heliothrips haemorrhoidalis and its relatives, with one new species and one new genus (Thysanoptera: Thripidae). Zootaxa 2015, 4021, 578–584. [Google Scholar] [CrossRef] [Green Version]
- Mound, L.A.; Nielsen, M.; Hastings, A. Thysanoptera Aaotearoa—Thrips of New Zealand. Available online: https://keys.lucidcentral.org/keys/v3/nz_thrips/ (accessed on 3 April 2019).
- Nickle, D.A. Commonly Intercepted Thrips at U.S. Ports-of-Entry from Africa, Europe, and the Mediterranean. IV. Miscellaneous Thripine Genera Excluding Frankliniella, Iridothrips, and Thrips (Thysanoptera: Thripidae). In Proceedings of the Entomological Society of Washington, Washington, DC, USA, 1 January 2009; Volume 111, pp. 215–238. [Google Scholar]
- Raspudić, E.; Ivezić, M.; Brmež, M.; Trdan, S. Distribution of Thysanoptera species and their host plants in Croatia. Acta Agric. Slov. 2009, 93, 275–283. [Google Scholar] [CrossRef] [Green Version]
- Trdan, S. Thrips in Slovenia: Thrips and tospoviruses. In Proceedings of the 7th International Symposium on Thysanoptera, Reggio Calabria, Italy, 2–7 July 2001; pp. 351–356. [Google Scholar]
- Garcia-Fayos, P.; Goldarazena, A. Role of Thrips in Pollination of Arctostaphyllos uva-ursi. Int. J. Plant Sci. 2008, 169, 776–781. [Google Scholar] [CrossRef] [Green Version]
- Jenser, G. Data to the Thysanoptera fauna of Tunisia. Folia Entomol. Hung. 1982, 43, 55–57. [Google Scholar]
- Trdan, S.; Andjus, L.; Raspudić, E.; Kač, M. Distribution of Aeolothrips intermedius Bagnall (Thysanoptera: Aeolothripidae) and its potential prey Thysanoptera species on different cultivated host plants. J. Pest Sci. 2005, 78, 217–226. [Google Scholar] [CrossRef]
- Badieritakis, E.G.; Thanopoulos, R.C.; Fantinou, A.A.; Emmanouel, N.G. Emmanouel qualitative and quantitative study of thrips (Thysanoptera) on alfalfa and records of thrips species on cultivated and wild Medicago species of Greece. Biologia 2015, 70, 504–515. [Google Scholar] [CrossRef]
- Mound, L.A.; Teulon, D.A.J. Thysanoptera as Phytophagous Opportunists; Thrips Biology and Management; Springer: Boston, MA, USA, 1995; pp. 3–19. [Google Scholar]
- Tyagi, K.; Kumar, V. Thrips of Economic importance in India—An Identification guide. Zool. Surv. India 2020, 2020, 96. [Google Scholar]
- Goldarazena, A. Orden Thysanoptera. Rev. SEA 2015, 52, 1–20. [Google Scholar]
- Jenser, G.; Tzanakakis, M.E. Records of Thysanoptera from Northern Greece. Entomol. Hell. 2017, 3, 59. [Google Scholar] [CrossRef] [Green Version]
- Kucharczyk, H.; Zawirska, I. The occurrence of Thysanoptera in Poland; Thrips and Tospoviruses. In Proceedings of the 7th International Symposium on Thysanoptera, Reggio Calabria, Italy, 2–7 July 2001; pp. 341–344. [Google Scholar]
- Vassiliou, V.A. Ecology and Behavior of Pezothrips kellyanus (Thysanoptera: Thripidae) on Citrus. J. Econ. Entomol. 2010, 103, 47–53. [Google Scholar] [CrossRef]
- Conti, F.; Tumminelli, R.; Amico, C.; Fisicaro, R.; Frittitta, C.; Perrotta, G.; Marullo, R. Monitoring Pezothrips kellyanus on citrus in eastern Sicily. Thrips and Tospoviruses. In Proceedings of the 7th International Symposium on Thysanoptera, Reggio Calabria, Italy, 2–7 July 2001; pp. 207–210. [Google Scholar]
- Navarro Campos, C. Pezothrips Kellyanus (Thysanoptera: Thripidae), Nueva Plaga en Cítricos; Comportamiento de sus Poblaciones, Muestreo y Enemigos Naturales. Unpublished. Ph.D. Thesis, Universitat Politècnica de València, Valencia, Spain, 2013; p. 166. [Google Scholar]
- Varikou, K.; Tsitsipis, I.; Alexandrakis, V.; Hoddle, M. Effect of Temperature on the Development and Longevity of Pezothrips kellyanus (Thysanoptera: Thripidae). Ann. Entomol. Soc. Am. 2009, 102, 835–841. [Google Scholar] [CrossRef]
- Nickle, D.A. Commonly Intercepted Thrips at U.S. Ports-of-Entry from Africa, Europe, and the Mediterranean. III. The Genus Thrips Linnaeus, 1758 (Thysanoptera: Thripidae). Proc. Entomol. Soc. Wash. 2008, 110, 165–185. [Google Scholar] [CrossRef]
- Kucharczyk, H.; Kucharczyk, M. Characteristic and diagnostic features of the most frequently occurring species of the Thripidae family (Insecta, Thysanoptera) in crown canopies of Central European forests. Leśne Pr. Badaw. 2013, 74, 5–11. [Google Scholar]
- Khan, F.; Roy, M.C.; Kim, Y. Thelytokous Reproduction of Onion Thrips, Thrips tabaci Lindeman 1889, Infesting Welsh Onion and Genetic Variation among Their Subpopulations. Insects 2022, 13, 78. [Google Scholar] [CrossRef]
- Mifsud, D. Present knowledge of the Entomofauna of the Maltese Islands. Entomol. Basiliensia 2000, 22, 75–86. [Google Scholar]
- Schembri, P.J. Current state of knowledge of the Maltese non-marine fauna. In Malta Environment and Planning Authority Annual report and accounts. Malta Environ. Plan. Auth. 2003, 2003, 33–65. [Google Scholar]
- Cassar, T.; Mifsud, D. Insects of the Maltese Islands. Entomol. Soc. Malta 2023, 12, 711. [Google Scholar]
- Berzosa, J. Thysanoptera checklist (Insecta, Thysanoptera) of the Canary Islands. Geographical distribution, host·plants and bibliography. Boll. Real Soc. Esp. Hist. Nat. Sec. Biol. 2000, 96, 93–112. [Google Scholar]
- Vigna Taglianti, A.; Audisio, P.A.; Biondi, M.; Bologna, M.A.; Carpaneto, G.M.; De Biase, A.; Fattorini, S.; Piattella, E.; Sindaco, R.; Venchi, A.; et al. A proposal for a chorotype classification of the Near East fauna, in the framework of the Western Palearctic region. Biogeogr. J. Integr. Biogeogr. 1999, 20, 1. [Google Scholar] [CrossRef] [Green Version]
- Carapezza, A.; Mifsud, D. New records of true bugs (Hemiptera, Heteroptera) from the Maltese Islands 2015. Bull. Entomol. Soc. Malta 2015, 7, 27–50. [Google Scholar]
Figure 1.
Map of the Maltese Islands (Gozo and Malta) showing the locations where data were collected between 2015–2022 (with names, habitat description and predominant vegetation of locations indicated in Table 1).
Figure 1.
Map of the Maltese Islands (Gozo and Malta) showing the locations where data were collected between 2015–2022 (with names, habitat description and predominant vegetation of locations indicated in Table 1).

Table 1.
Locations where data were collected for this study: habitat type and brief description.
| Site No. | Location | Habitat Type and Description |
|---|---|---|
| 1 | Ta’ Sabbara, Żabbar | Steppe, with typical vegetation including Foeniculum vulgare, Vicia sativa and, especially when in a degraded state, Asphodelus ramosus, Galactites tomentosa and Stipa capensis, and. Most plants in this habitat dry up during the dry summer period [6]. |
| 2 | Wied Anġlu, Birguma | |
| 3 | Dingli Cliffs, Dingli | Garigue, characterised by low lying, often aromatic shrubs adapted to resist drought and strong winds, e.g., Thymbra capitata. Commonly found species include Drimia (=Urginea) maritima, Erica multiflora, and Euphorbia melitensis [6]. |
| 4 | Country road, l/o Rabat | |
| 5 | Manikata | |
| 6 | Ċirkewwa | |
| 7 | Lapsi, Siġġiewi | |
| 8 | Xrobb l-Għaġin | |
| 9 | Għaxqet l-Għajn, Għargħur | |
| 10 | Mġiebaħ, Xemxija | |
| 11 | Lippija Tower, Mġarr | |
| 12 | Pembroke | |
| 13 | l/o Majjistral Park, Għajn Tuffieħa | |
| 14 | Xlendi Tower, Xlendi, Gozo | |
| 15 | Ta Ċenċ, Gozo | |
| 16 | Il-Maqluba, Qrendi | Maquis, with plants consisting largely of evergreen small trees such as Ceratonia siliqua and Olea europaea, large shrubs such as Pistacea lentiscus and climbers such as Hedera helix and Smilax aspera, as well as herbaceous shade loving species such as Acanthus mollis [6]. |
| 17 | Il-Fawwara, Siġġiewi | |
| 18 | Mgarr ix-Xini, Gozo | |
| 19 | Ħas-Saptan, Gudja | Woodland, comprising of Mediterranean sclerophyll forest with trees including Quercus ilex and Pinus halepensis and an undergrowth of smaller shrubs. Tamarix spp. prevails on coastal wood remnants, while Populus alba occurs in riparian woodland remnants [6]. |
| 20 | Buskett, Dingli | |
| 21 | Wied Għollieqa, San Ġwann | Valleys, with slopes having plants typically found in maquis environments and the valley itself. Plants occurring in these habitats include Hyparrhenia hirta, Rubus ulmifolius and Potentilla reptans and, where water courses are present also some hydrophytes such as Ranunculus saniculifolius. |
| 22 | Wied Ħesri, Siġġiewi | |
| 23 | Wied Musa, Siġġiewi | |
| 24 | Wied Xkora, Siġġiewi | |
| 25 | Wied Qirda, Żebbuġ | |
| 26 | Wied il-Baqqiegħa, Żebbuġ | |
| 27 | Wied Speranza, Mosta | |
| 28 | Wied Qlejgħa, Rabat | |
| 29 | Wied tal-Fiddien, Rabat | |
| 30 | Wied Babu, Żurrieq | |
| 31 | Il-Qattara, Dwejra, Gozo | |
| 32 | Ramla l-Ħamra, Gozo. | Sand dune, with vegetation that needs to be suited to harsh conditions including high temperatures, dryness, occasional inundation by seawater and accumulation of sand. These plants include Cakile maritima, Elytrigia juncea, Eryngium maritimum, Lotus cytisoides and Pancratium maritimum. |
| 33 | Il-Ballut, Marsaxlokk | Saltmarshes, with plants that tolerate high salinity such as Athrocnemum macrostachyum, Salsola soda and Salicornia ramosissima. |
| 34 | Siġġiewi (roads) | Town roads. Plants growing along these roads include a variety of both indigenous species (e.g., Diplotaxis tenuifolia) as well as non-indigenous species. These sites yield plants which are cultivated, non-indigenous species. |
| 35 | Naxxar (road) | |
| 36 | Rabat (roads) | |
| 37 | Żabbar (road) | |
| 38 | Attard (road) | |
| 39 | Victoria, Gozo (roads) | |
| 40 | Popeye Village, Mellieħa road | Country roads. These host a number of indigenous plants including Glebionis coronaria and Antirrhinum siculum. |
| 41 | Għajn Tuffieħa road | |
| 42 | Ġebel San Pietru, Madliena | |
| 43 | Il-Kunċizzjoni, l/o Rabat | |
| 44 | Żebbuġ (roads) | By-passes. A number of central strips and roundabouts on the proximity of these roads include cultivated species such as Solandra maxima, Albizia julibrissin and Nerium oleander. |
| 45 | Luqa road | |
| 46 | Mosta road | |
| 47 | Mellieha by-pass | |
| 49 | Zabbar (private gardens) | Private gardens. These include a variety of cultivated herbaceous plants and fruit trees such as Citrus. |
| 50 | Mosta garden | |
| 51 | Attard garden | |
| 52 | Junior College Ringroad, Msida | |
| 53 | University of Malta Grounds, Msida | |
| 54 | Siggiewi plant nursery | |
| 55 | St. Thomas Bay, Marsaskala garden | Public gardens. These feature a combination of cultivated plants, e.g., Yucca gloriosa and Salvia coccinea and indigenous species including Tamarix africana and Atriplex halimus. |
| 56 | Sta. Lucia garden | |
| 57 | Golden Bay garden | |
| 58 | Qbajjar, Gozo garden | |
| 59 | Floriana Car Park | Public cultivated green areas with plants as described in the “Town roads” section. |
| 60 | Qormi (roundabout) | |
| 61 | San Ġwann farm | Private farms, which grow a variety of crop plants. |
| 62 | Qormi farm | |
| 63 | Siġġiewi farm | |
| 64 | Ta’ Qali l/o Attard field | Private open cultivated fields with a variety of plants as described in private farms. |
| 65 | Tal-Ferħa, Għargħur field |
Table 2.
Proportion of chorotypes for the Terebrantia of the Maltese Islands.
| Chorotype | Number of Species | % |
|---|---|---|
| Subcosmopolitan | 7 | 17.9 |
| Cosmopolitan | 5 | 12.8 |
| Palaearctic | 5 | 12.8 |
| Turano Mediterranean | 4 | 10.3 |
| Asiatic European | 3 | 7.7 |
| Afrotropico Mediterranean | 3 | 7.7 |
| Mediterranean | 3 | 7.7 |
| Turano European | 2 | 5.1 |
| Afrotropico-Indo-Mediterranean | 1 | 2.6 |
| European | 1 | 2.6 |
| Europeo Mediterranean | 1 | 2.6 |
| Holarctic | 1 | 2.6 |
| Sibero European | 1 | 2.6 |
| Turano-Europeo-Mediterranean | 1 | 2.6 |
| West Palaearctic | 1 | 2.6 |
| Total: | 39 | 100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Degabriele, G.; Cavalleri, A.; Goldarazena, A.; Mifsud, D. The Terebrantia (Insecta: Thysanoptera) of the Maltese Islands. Diversity 2023, 15, 514. https://doi.org/10.3390/d15040514
AMA Style
Degabriele G, Cavalleri A, Goldarazena A, Mifsud D. The Terebrantia (Insecta: Thysanoptera) of the Maltese Islands. Diversity. 2023; 15(4):514. https://doi.org/10.3390/d15040514
Chicago/Turabian StyleDegabriele, Godwin, Adriano Cavalleri, Arturo Goldarazena, and David Mifsud. 2023. "The Terebrantia (Insecta: Thysanoptera) of the Maltese Islands" Diversity 15, no. 4: 514. https://doi.org/10.3390/d15040514
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.