
Revisiting Biodiversity and Ecosystem Functioning through the Lens of Complex Adaptive Systems

1
Department of Animal Biology, Faculty of Sciences, University of Lisbon, Campo Grande, 1749-016 Lisbon, Portugal
2
MARE-Marine and Environmental Sciences Centre, Faculty of Sciences, University of Lisbon, Campo Grande, 1749-016 Lisbon, Portugal
3
Global Health and Tropical Medicine, GHTM, Associate Laboratory in Translation and Innovation towards Global Health, LA-REAL, Instituto de Higiene e Medicina Tropical, IHMT, Universidade NOVA de Lisboa, UNL, Rua da Junqueira 100, 1349-008 Lisboa, Portugal
4
Ce3C-Centre for Ecology, Evolution and Environmental Changes, Faculty of Sciences, University of Lisbon, Campo Grande, 1749-016 Lisbon, Portugal
*
Author to whom correspondence should be addressed.
Diversity 2023, 15(8), 895; https://doi.org/10.3390/d15080895
Submission received: 19 June 2023
/
Revised: 21 July 2023
/
Accepted: 25 July 2023
/
Published: 28 July 2023
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Understanding the relationship between biodiversity and ecosystem functioning (BEF) is essential to comprehend the impacts of biodiversity changes on ecosystem functioning. This knowledge helps to detect and anticipate significant trends in global biodiversity loss and the homogenization of biota worldwide to prevent them. Species act together with climate, resource availability, and disturbance regimes to modulate ecological processes defining ecosystems’ complexity and their dynamic adaptation to variability. In this article, we revisit the BEF paradigm by addressing current knowledge of how biodiversity connects to ecosystem functioning across scales in the context of complex adaptive systems (CAS). We focus on ecosystem processes that lead to the emergence of the BEF relationship, considering ecosystem functioning as a macroscopic emergent property. Specifically, this work integrates the knowledge of the processes that connect biodiversity to ecosystem functioning. It addresses how biodiversity supports ecosystem multifunctionality across scales, resulting in the persistence of CAS in a rapidly changing world. We present a framework for ecological management considering the BEF relationship within the scope of CAS. The CAS standpoint brings new insights into the BEF field and its relevance for future ecological conservation of the Earth’s life support.
1. Introduction
Global climate change, overexploitation of natural resources, destruction and fragmentation of natural habitats, and biological invasions are now having measurable effects on ecosystem functioning and are expected to cause unprecedented effects on biodiversity by increasing the rates of population and species extinctions in the near future [1,2]. Even though the loss of species is a natural process on the evolutionary scale [3,4], the current speed of non-random extinctions in natural systems due to the direct or indirect influence of human activity [5] is alarming. The effects of extinctions may have considerable time-lags that are challenging to identify [5,6], complicating long-term predictions of global-changing biodiversity effects. The relationship between biodiversity and ecosystem functioning (BEF) has thus led to significant interest in ecology [6,7,8,9,10,11].
Research on BEF is fundamental to understand the influence of species decline and the homogenization of biota on general ecosystem properties. It also has particular societal relevance when studying the consequences for ecosystems on which humans depend [5,12,13,14]. BEF relationships are highly complex, i.e., with diverse, interacting, and adapting elements [2,11,15]. Thus, addressing BEF within the complex adaptive systems (CAS) framework can improve our understanding of the links between biodiversity and ecosystem functioning. This requires relating the adaptive interactions that characterize ecosystems with the dynamics of ecological processes operating on BEF, along different levels of ecological organization, environmental heterogeneity, and spatiotemporal scales [12,15,16,17,18,19,20,21]. This knowledge is crucial to guide an effective management of resources to preserve ecosystem services critical for the continuity of present and future generations.
Biodiversity encompasses the number, distribution, and functional traits of individuals and species, intra- and interspecific genetic diversity, levels of organization (populations, communities, ecosystems, biomes), and ecological interactions (trophic, behavioral, competitive, mutualistic, and parasitic) at different spaciotemporal scales, defining the basis of ecosystems as complex adaptive systems [4,11,15,22,23,24]. Ecosystem functioning refers to the state or trajectory of ecosystems in terms of innate pathways and fluxes of energy, matter, and information occurring through essential ecosystem processes, such as productivity, nutrient and biogeochemical cycling, and ecological network dynamics, from which is derived the stability that supports ecosystem complexity at a larger scale [15,16,22,25,26]. Complex adaptive systems are characterized by interconnected components, non-linear dynamics, and the ability to self-organize and adapt in response to environmental changes [16,27]. Within this framework, biodiversity and ecosystem functioning are not static entities but dynamic processes that emerge from local interactions within a larger system [4,16,17]. The importance of examining BEF within the scope of CAS lies in the recognition that ecosystem functioning may be perceived as an aggregate (or macroscopic) property since it displays the emergent nature of ecosystems that self-organize from local evolved components, as well as feed back loops [22]. CAS map our understanding of local biological interactions and ecosystem processes that lead to emerging patterns, such as uncertainty, biodiversity, collective behavior, common pool resources, robustness, redundancy, heterogeneity and connectivity, that underlay ecosystem functioning [4,16,28]. These emergent properties may, in turn, feed back to the local biotic and abiotic interactions generating regular changes, critical phenomena, and flips among alternative stable states [4,16,17,27,29,30]. These feed backs result from evolutionary processes at lower levels of the ecological organization. Biodiversity acts as a buffer and homeostasis agent contributing to the coevolution of ecosystems and the biosphere [16,31]. Thus, comprehending the processes linking biodiversity to ecosystem functioning, and the overall contribution of biodiversity to improve ecosystem multifunctionality by considering this complexity and adaptability, is essential to develop conservation actions that will help to prevent future biodiversity loss and ecosystem oversimplification.
This article revisits the biodiversity and ecosystem functioning paradigm by focusing on the links between biodiversity and ecosystem functioning across scales in the context of CAS. We present an integrative approach to examining why biodiversity is important for ecosystem functioning by relating the processes that connect biodiversity to ecosystem functioning. Then, we focus on how biodiversity ensures ecosystem multifunctionality across distinct scales, highlighting its CAS nature, which enables adaptation to continuous and abrupt changes. We ask four main questions: (1) How does biodiversity relate to ecosystem functioning within the framework of complex adaptive systems? (2) How does biodiversity interconnect with ecosystem processes, such as productivity, nutrient and biogeochemical cycling, ecological networks dynamics, and stability, to modulate BEF? (3) How does biodiversity link to ecosystem multifunctionality, particularly its role in sustaining multiple simultaneous processes to generate adaptability? (4) What are the implications of understanding BEF under the lens of CAS for ecological management? From the CAS perspective, these questions focus on the ascending effects that lead to BEF relationships considering that ecosystem functioning is a macroscopic property shaped by emerging patterns. The rationale for this work encompassed the review of the empirical and theoretical literature on BEF and CAS across systems at different scales (temporal, spatial, evolutionary, ecological). We discuss major findings, from seminal to current research, that have provided increasingly rigorous answers to the above questions. Then, using a BEF approach and adopting CAS thinking, we present an exploratory framework for ecological management. This approach can be used as a guideline to develop plans aiming at sustaining complex adaptive ecosystems, which are the foundation of all life on Earth. We finalize by bringing forward contemporary challenges and prospects. By connecting key data-driven and theoretical findings from a long body of research, this review contributes with a comprehensive background knowledge coupling BEF with CAS.
2. The Biodiversity–Ecosystem Functioning Paradigm
The study of the relationship between biodiversity and ecosystem functioning (BEF) has been a dynamic and thriving research area in the field of ecology for the last three decades. Until the beginning of the 1990s, biodiversity was viewed as the result of the interaction between abiotic and biotic factors, with little or no consequences on ecosystem functioning [32,33]. It was only after the 1992 Bayreuth conference that the connection between biodiversity and ecosystem functioning (BEF) emerged as a new research field [34,35,36,37,38]. Within the 1990s, due to the increasing rate of species loss, research started to reveal some effects of species diversity on the environment and the functioning of ecosystems [32,36,38,39]. These effects manifested in habitats through changes in ecosystem structure, and fluxes of nutrients, productivity, and stability [38], leading to a paradigm shift in ecology [32]. Seminal work in this area ([14,36,38,39] can be seen for further details) focused primarily on fieldwork and experimentation to test the hypothesis that species diversity affects ecosystem functioning e.g., [40,41,42,43,44,45,46,47]. In addition, theoretical studies integrating empirical results emerged in an attempt to determine if, and how, biodiversity predictably affected essential ecological processes [48,49,50,51,52,53,54]. The growing interest in this area led to the publication of contradictory results that instigated a strong degree of dispute around the validity of BEF experiments (design, mechanisms involved, applicability to natural systems) [10,34,35,36,39,47,55,56,57,58,59,60,61,62,63,64,65]. However, after three decades of research on BEF, several papers have been published, underlining a general trend on the positive link between different metrics of biodiversity and ecosystem functioning throughout terrestrial, freshwater, and marine ecosystems [2,34,35,38,39,66,67,68,69,70,71,72].
Several biodiversity attributes have been used to test the BEF hypothesis at different scales (α-diversity: local; β-diversity: regional; and γ-diversity: global). These include taxonomic approaches (species presence or absence); richness (number of species) and evenness (equity of species abundances); functional and phylogenetic diversity (functional groups within the ecosystem, the distribution of species traits within a community, or the degree of species similarities and differences); and genetic diversity (genotypic variation and genotype identity). However, the best measure to capture the BEF relationship is still a subject of debate [10,14,34,67,68,73,74,75,76,77]. Richness and evenness generally show positive or neutral effects on ecosystem productivity and related properties, while functional, phylogenetic, and genetic diversity seem to exert an impact on mixed ecosystem processes, with a tendency for genetic diversity to better predict community productivity, robustness, invasibility, and vulnerability to disease [14,31,34,68,73,74,75]. Adopting multidimensional biodiversity attributes allows a better understanding of the intricate relationship between biodiversity and ecosystem functioning, and avoids underestimating the repercussions that result from biodiversity loss [67]. In principle, the use of particular biodiversity metrics will depend on the question, the processes, and the scale under analysis.
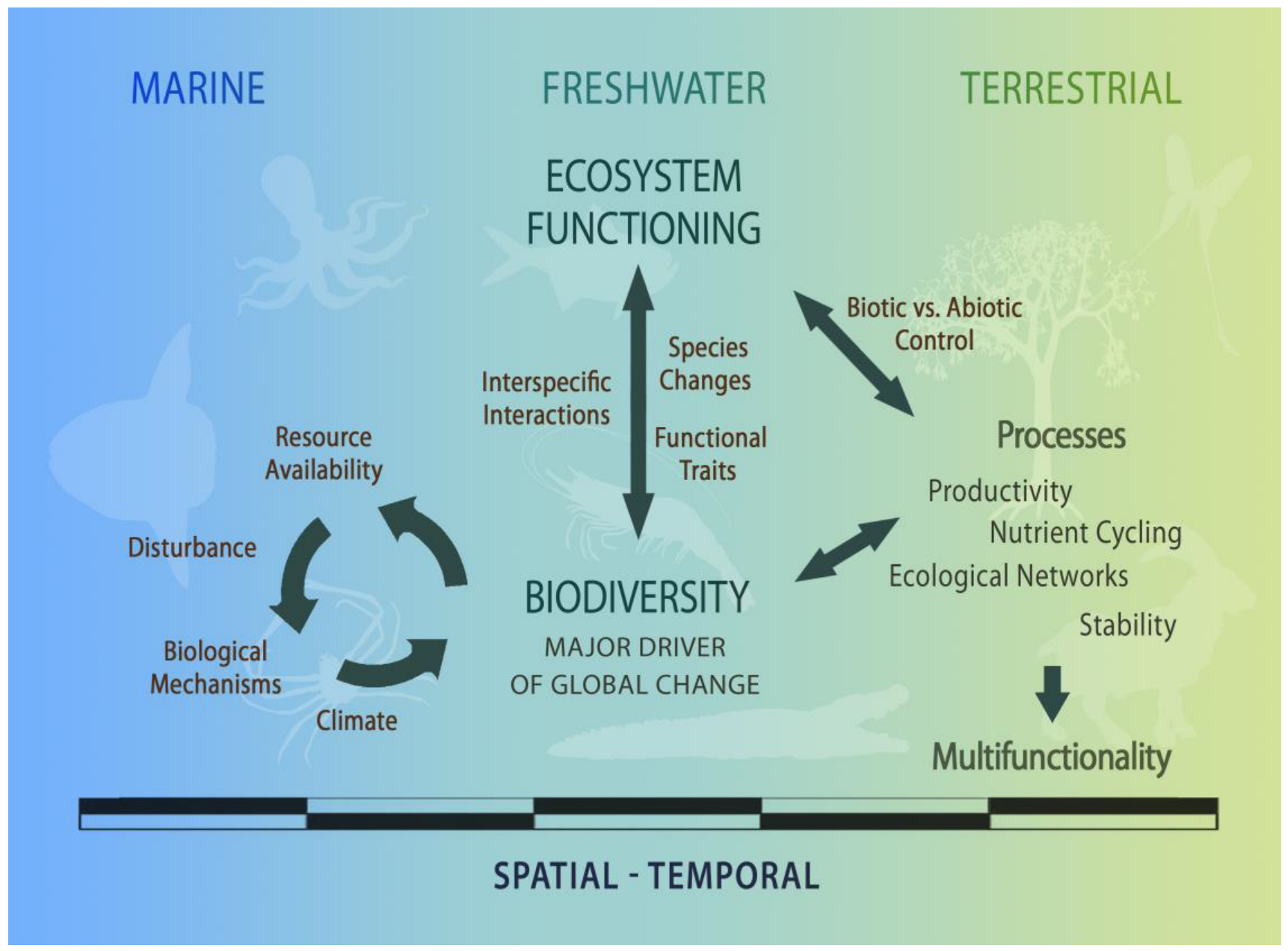
Despite this intricacy, the still-expanding scope of answers, and the emerging new questions on BEF, various studies have identified common patterns regarding the links between biodiversity and ecosystem functioning [5,10,14,26,34,38,67,76,77,78,79], which are put into perspective in Figure 1. In accordance, biodiversity, either at the level of genes, species, communities, or functional traits, interacts with climate, resource availability, and disturbance regimes at different scales to shape ecosystem functioning. By regulating distinct ecosystem processes through a common set of biological mechanisms, biodiversity fosters ecosystem multifunctionality and sustains dynamic complex adaptive ecosystems [67]. The magnitude of biodiversity effects over ecosystem functioning depends on the functional traits of component species, the strength of interspecific interactions, the sequence of species changes across space and time, and the relative biotic versus abiotic control over ecosystem process rates [5,10,56,67,76,77].
The integration of concepts and empirical studies on BEF has emphasized the crucial role of biodiversity in the functioning of ecosystems and increased our understanding of the consequences of diversity loss for ecosystems and humanity [5,12,26,38,66]. In fact, recent advances on BEF have indicated that minimal biodiversity changes may have profound effects on ecosystem functioning. Consequently, the impact of species loss may be comparable to, or even greater than, that of other environmental factors held responsible for global change [2,13,14,32,71,76]. These studies represented a new shift in the biodiversity ecosystem functioning paradigm by acknowledging that biodiversity, in conjunction with environmental heterogeneity, is a pivotal player in the regulation of ecosystem functioning.
3. Biodiversity and Ecosystem Functioning within the Scope of CAS
The intricate relationships and interactions among diverse species and their environment create complex interdependencies that contribute to ecosystem functioning. Environmental conditions act on biodiversity, indirectly influencing ecosystem functioning [10,76]. Furthermore, environmental factors may directly affect ecosystem functioning, especially within the context of global environmental changes [67]. This implies a balance between species adaptation to existing settings and plasticity to endure environmental changes. Biodiversity encompasses different species with unique adaptations and traits that enable them to occupy specific niches and perform distinct ecological functions affecting different ecosystem processes [5,7,10,14,26,34,38,76,77,79,80]. These processes are nonlinear [12,77] because the properties of ecosystems accumulate, deplete, and feed back to shape future ecological interactions, leading to uncertainty [3,16,17,24,28]. Ecosystem functioning emerges from the interplay between different levels of biological organization, ecosystem processes, and environmental heterogeneity at small scales. Its macroscopic dynamic patterns feed back to shape the development of local interactions into subsequent aggregate properties [22,24,31]. Consequently, biodiversity sustains the ecosystem’s adaptive capacity to adjust to biological and environmental changes across different ecological and evolutionary scales [3,16,17,21,24,28]. Individual interactions and ecosystem processes result in collective behaviors and the production of common pool resources that extend to the whole ecosystem, which, in turn, slowly influence individual behaviors and the production of natural resources through expanded time scales [3,28,31]. Thus, individual biological components induce evolutionary changes that percolate throughout the ecosystem and evolve from the interchange of ecological processes at different scales [17]. These processes produce macroscopic properties, e.g., ecosystem functioning, emerging from trade-offs during the course of evolution and the capacity of biological components to adapt to rapid ecological shifts [16,22,24,31]. Ecosystem functioning is also adaptive because biological interactions tend to respond to interference. For instance, the presence of diverse species increases the likelihood of some degree of redundancy, i.e., some species have traits that enable them to tolerate or adapt to changing conditions [81]. The adaptive capacity of ecosystems confers robustness, i.e., a balance between adaptation to existing settings and plasticity to endure environmental changes. Biodiversity and ecological heterogeneity sustain permanent local environmental and biological changes which balance the turnover of biotic components through continuous abiotic modifications [28]. Heterogeneity results from the interaction among ecological processes that occur in different spatial and temporal scales. This encompasses, for example, the counterbalance between positive local short-term feed back and negative broad long-term feed back, which create and stabilize heterogeneity, respectively [3].
The BEF relationship reflects the multifunctionality and continuous adaptation of biotic and environmental interactions at different spatial and temporal scales [72,75,82,83,84] from short-term behavioral responses, and medium-term physiological and developmental phenotypic plasticity, to long-term evolutionary changes [17,75,82,85,86,87]. Some studies have detected a connection between biodiversity and ecosystem functioning at local but not at regional or landscape levels [34]. However, several studies have indicated that the effects of biodiversity increase through time and at larger spatial scales [10,38,76,77,85,88,89]. Large spatial scales provide environmental heterogeneity and connectivity, allowing the coexistence and dispersal of species, and are key to evolution and the maintenance of biodiversity [3,21,90]. Additionally, the prevalence of multiple ecosystem processes at larger scales entails higher biodiversity than at smaller scales [10,38]. Multiple scales unite features of space, time, and ecological complexity that sustain biodiversity. Broad scales allow the patterns of biological and environmental variability to expand, generating more patches and resources through the evolutionary divergence of species’ ecological niches and life traits [3]. This drives speciation through the short-term advantage of new traits capable of exploring new niches and permeating throughout biological assemblies over ecological and evolutionary scales [3,17].
From the CAS point of view, integrating different biodiversity descriptors, and multiple ecosystem processes across temporal and spatial scales, will more likely provide a better understanding of BEF.
4. Linking Biodiversity to Ecosystem Processes
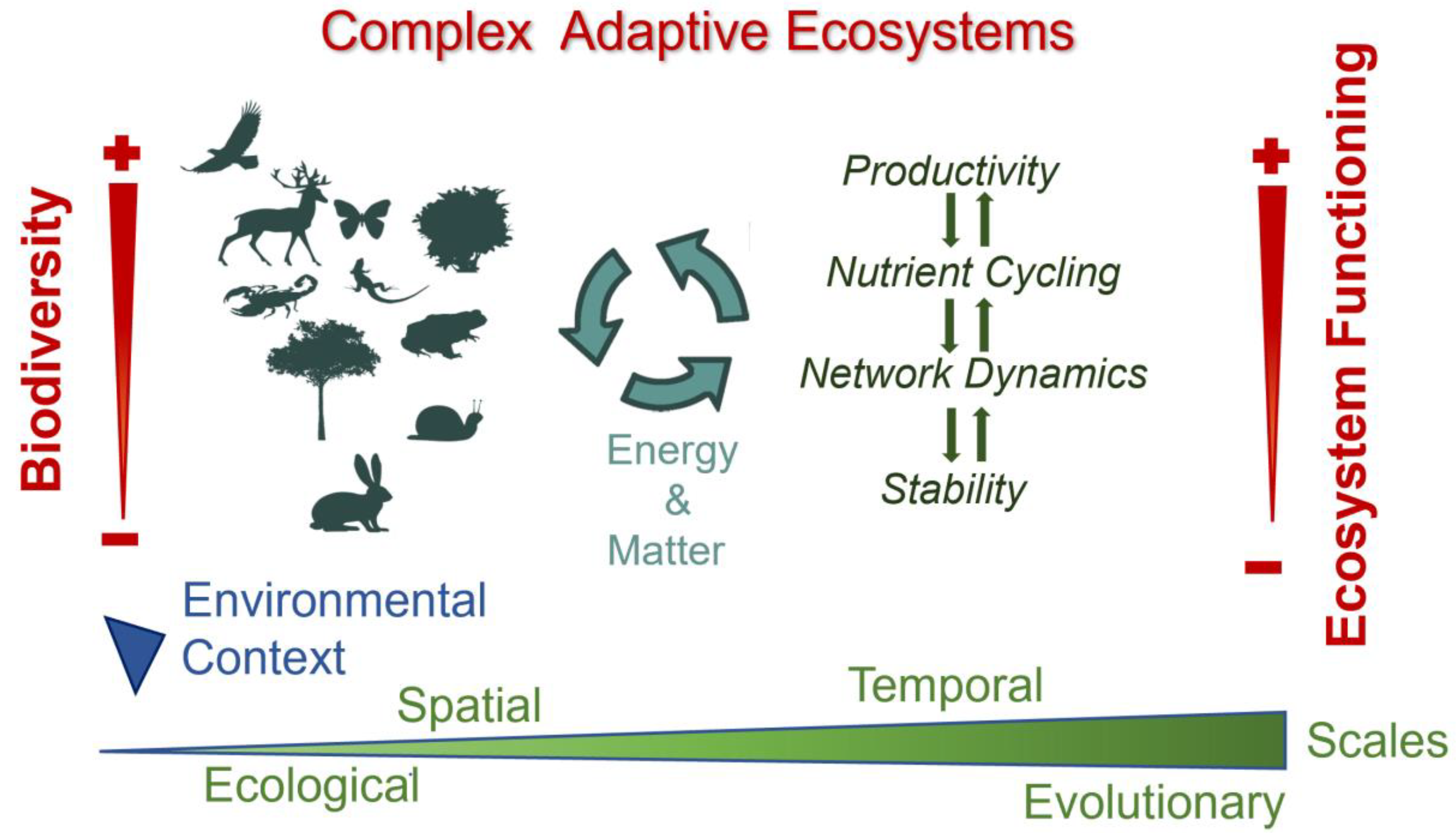
Moving our understanding of BEF forward through the lens of CAS requires looking at basic nonlinear ecosystem processes (such as productivity, nutrient and biogeochemical cycling, ecological network dynamics, and stability), which foster fluxes of matter, energy, and information [15,16,22,25,26], through the local interplay between biodiversity and abiotic features that translate into macroscopic properties, such as ecosystem functioning (Figure 2); we address this in the sub-sections below. In accordance, biodiversity simultaneously interacts with several dynamic processes in different environmental contexts, leading to ecosystem functioning, across different spatial and temporal scales, as well as ecological and evolutionary scales. In this context, highly diverse communities contribute to enhanced ecosystem functioning in heterogeneous environmental conditions, promoting complexity and adaptation (i.e., complex adaptive ecosystems). However, this is one of the most difficult challenges for the field of BEF because spatiotemporal interconnectedness, adaptability, variability, and uncertainty characterize the reciprocal interactions among biotic/biotic and biotic/abiotic components that regulate ecosystem functioning and maintain complex adaptive ecosystems [5,15,38]. Further, the simultaneous effect of environmental factors and biodiversity on multiple ecosystem processes, and thus on ecosystem functioning, remains unclear [72].
4.1. Productivity
Productivity, i.e., the rate of biomass production in ecosystems, is an essential aspect of BEF due to transformations of energy and matter flow through interspecific interactions that determine the productivity of the entire ecosystem [7,66,91]. Decades of theoretical and empirical work have shown that biodiversity has congruent outcomes in primary and secondary productivity—herein termed productivity—and vice versa [7,14,25,61,92,93]. Overall, highly diverse communities will maximize resource use efficiency, being more productive than systems characterized by reduced biodiversity, and this disparity increases with time [77]. These differences in productivity related to biodiversity are triggered by distinct causes, namely, interspecific niche partitioning, more effective use of limiting resources (e.g., nutrients, water, prey, space, and light), energy that is more efficiently converted into biomass or decomposed, reduced herbivory and disease, and nutrient-cycling feed backs that increase elements such as nutrient stocks and allocation rates over time [10,14,38,75,76,94,95]. More diverse assemblages may contain key species with a substantial impact on productivity, as well as species with diverse functional traits that sustain an increase in total resource uptake [38]. Findings from [73] support the idea that the correlation between biodiversity and productivity, due to resource partitioning, is related to evolutionary and genetic divergence. In situations of heterogeneous limiting resources, biomass production increases with biodiversity as more species will be able to capture the few available nutrient supplies [11,66]. Therefore, reduced biodiversity will lead to less efficient use of resources that will translate into decreased productivity [26,35,59,68,73,96], and thus accessible common pool resources. Furthermore, in cases of extreme climate events, biodiversity is also a pivotal factor in balancing and upholding ecosystem productivity [97].
Biological mechanisms, such as complementarity [10,63,66,72,80,83,95,98,99,100], facilitation [10,26,66,76,95,98,101], and selection [26,50,59,63,66,72,83,95,98,99,102] effects, primarily control the relationship between biodiversity and productivity [72,103,104]. This is because dominant species contribute to increase productivity, which is further increased by the occurrence of multiple complementary and facilitating species [63,72,99,103]. However, in extreme unstable conditions or during habitat transformation, there may be a significant shift in species composition, resulting in lower biodiversity characterized by dominant species with high adaptive capacity [105]. Complementarity and facilitation are the main drivers behind the effects of biodiversity on the productivity of aquatic ecosystems, while, in terrestrial ecosystems, these effects may be similarly triggered by complementarity and selection [66,72,101].
Most studies have indicated that the relationship between biodiversity and productivity varies through time, space, ecosystems, and ecological levels as species interactions and community structure adjust to successional, seasonal, and environmental gradients and changes [10,51,63,87,94,99,103,106,107,108,109].
4.2. Nutrient and Biogeochemical Cycling
Ecosystem functioning relies on nutrient and biogeochemical capacity, i.e., the aptitude of an ecosystem to store and recycle nutrients across its biotic/abiotic components [110]. It depends on the availability of nutrients and their interconnected dynamics, which uphold metabolic processes and biodiversity feed backs [111]. In fact, natural ecosystems are characterized by complex pathways that capture, store, and transfer nutrients as energy flows through different trophic levels and microbial processing [111,112]. Nutrient and biogeochemical cycling is an important ecosystem process supported by biodiversity and the physical complexity of the habitat. The biota regulates nutrient and biogeochemical processes by transferring materials among the hydro-, litho-, and atmospheres, sustaining and shaping ecosystems over ecological and evolutionary scales [113]. Heterogeneous habitats encompass high levels of biodiversity and resources, fostering nutrient and biogeochemical cycling [11,111,112]. The diversity of autotrophs, consumers (omnivores, predators, and herbivores), and decomposers (sensu lato) regulates nutrient availability and cycling throughout the whole ecosystem [114,115]. Biodiversity, either directly or indirectly, conveys multifold routes for nutrient processing, such as absorption by primary producers, transfer to and by consumers and pathogens, and decomposition, sustaining ecosystem functioning and reducing nutrient losses [66,115]. Increased biodiversity maximizes nutrient uptake and energy flows through ecological networks as more diverse communities include species with different traits. These assemblies provide additional pathways to nutrient intercept, assimilation, and conversion into biomass or dead organic matter, which comprise supplemental sources of nutrients and energy that are essential for ecosystem functioning [26,63,96,111,112].
The connection between biodiversity and nutrient and biogeochemical cycling seems to be primarily mediated by complementarity and facilitation [10,26,94,116]. Although some studies have pointed out the importance of biodiversity on nutrient and biogeochemical dynamics [26,60,66,94,110,111,114,117,118,119,120,121,122], we need a better understanding of the influence of biodiversity on this process.
4.3. Ecological Network Dynamics
Ecosystem functioning is the outcome of intricate linkages between biotic and abiotic factors [37] mediated by trophic and non-trophic interactions [123,124]. The set of direct and indirect trophic interactions and non-trophic links have a key role in tuning the connection between biodiversity and ecosystem functioning [100,125]. Complex direct (e.g., predation, competition, and parasitism) and indirect (e.g., behavior, habitat shift, morphology, and life history) trophic interactions (within and among trophic levels and trophic webs), together with non-trophic interactions (e.g., mutualism and commensalism) shape ecological networks, which promotes adaptive changes and influence ecosystem functioning through space and time [9,12,18,75,91,123,125,126,127,128,129,130]. Thus, understanding ecological networks as a dynamic process is essential to unveil the effects of biodiversity on ecosystem functioning underlying CAS. Ecological network dynamics are highly complex, displaying non-random and persistent spatiotemporal features, which convey consistent patterns linked to ecosystem functioning [18,123,124,130,131]. In an expansive approach, ecological network dynamics may be perceived through ecological interactions assessed by the trophic and non-trophic links and the degree of connection among species [125,132].
In the trophic web context, biodiversity links to ecosystem functioning through horizontal (within trophic levels) and vertical (among trophic levels) diversity [12]. So, biodiversity has the potential to simultaneously affect several trophic links in such a way that changes in contiguous levels may produce contrasting results on specific ecosystem processes [12,59,91,128,133,134].The existence of multiple trophic levels in natural trophic webs generates complex BEF relationships [9,128]. As multitrophic diversity increases, average ecosystem functioning may increase or be maintained, displaying complex nonlinear configurations [12,20,126,134]. Therefore, the trophic web dynamics (e.g., the flow of energy and matter through trophic levels) depends on the nature of the trophic interactions and the biodiversity within and across trophic levels [91,135], affecting productivity and nutrient cycling [9,112,127,128,129,130,136,137]. Further, it confers ecosystem adaptability by providing alternative routes for slow and fast fluxes of energy and matter and by reducing the strength of consumer–resource interactions that have potentially destabilizing effects on ecosystems [9,12,124,129,131]. Biodiversity also influences the strength of ecological interactions and the dynamics of ecological networks by balancing the top-down and bottom-up control of trophic webs [129,136]. Robust top-down and bottom-up forces have extensive indirect effects on ecological network dynamics and ecosystem functioning [118,126,134].Top-down control of biodiversity relates to the impact consumers have on lower trophic levels, i.e., the rate at which consumers convert resources into matter. Bottom-up effects of biodiversity refer to the impact of producers on higher trophic levels, i.e., the rate at which energy flows and resources are captured and converted into consumer matter [135,136,138]. Top-down control by predator diversity varies across ecosystems depending on whether the predator is a specialist or a generalist, on migration, dispersal, and habitat complexity [12,90,128,129,133,136,138]. Omnivores and intra-guild predators may have either positive or negative results on predator, consumer, and producer diversity [100], and are the main factors determining the effects of biodiversity at higher trophic levels [12]. Strict predators (carnivores) generally control their prey by either decreasing or maintaining their biodiversity, which in turn increases or maintains producer biodiversity [129]. High herbivore diversity controls producers by promoting a diversified resource use and buffering top-down effects of top predators [91]. This results from the trade-offs between herbivores’ competitive ability and anti-predator behaviours, e.g., asynchronous foraging activity or habitat shifts, which also dampen the impact of herbivory [91,100]. The diversity of consumers (omnivores, predators, and herbivores) may also exert top-down or bottom-up control on decomposition rates and vice versa [34,35,112,135]. Consumer diversity is essential for supporting balanced ecosystem functioning, dampening the risk of decoupling trophic cascade effects [131,133]. Producer diversity, on the other hand, upholds bottom-up control through competitive advantage and resistance to consumption [90,126,130,136,138], meaning that biodiversity may lead to the dominance of producers with relevant effects on ecosystem functioning [90,126]. Further, high consumer and producer biodiversity enhance mutualistic interactions that may elicit positive feed backs on the biodiversity of symbiotic interactors and on several ecosystem processes [14,70,87] and reduce the incidence of pathogens [14,70,114]. The balance between top-down vs. bottom-up control by biodiversity strongly determines the outcome of ecosystem functioning, the adjustment of ecosystems to biodiversity and environmental changes, and the decoupling of trophic cascade effects [27,127,128].
Complementarity, facilitation, and selection effects are the most common mechanisms underlying ecological network processes [18,26,91,126,128,129,137]. However, these mechanisms differentially mediate the effects of consumer vs. producer biodiversity on ecosystem functioning [12,26,126,129,137]. In an ecological network setting, complementarity and facilitation are not only the result of resource partitioning among competitors or positive interspecific interactions, but also of top-down control by upper trophic levels [18,100,129,137]. Likewise, selection effects may emerge from both the prevalence of species displaying anti-predator behavior and competitive dominance [129,137]. Consumer biodiversity can drive ecosystem functioning through different mechanisms, such as adaptation (i.e., adaptive protection by prey and adaptive food search by predators), non-additive interactions (i.e., the outcome of multi-consumer interactions is different from the sum of each component), and indirect effects (i.e., one species has an indirect effect on a third species), which are tiers of complementarity, facilitation, and selection [98,137,139]. These mechanisms may be related to consumer–consumer synergistic interactions or interference competition and to consumer–prey antagonistic interactions [18,98]. Adaptation, non-additive, and indirect mechanisms may decrease prey extinction as consumer diversity increases [100,137].
The set of direct and indirect trophic interactions and non-trophic links have a key role in tuning the connection between biodiversity and ecosystem functioning [100,125]. Combining distinct interactions and mechanisms at different levels of ecological organization, environmental heterogeneity, and scale conveys an in-depth insight into the connection between biodiversity and ecosystem functioning [12,15,16,17,18,19,20].
4.4. Stability
Ecosystem stability is a fundamental characteristic of CAS that may be perceived as the capacity ecosystems have to achieve and sustain a stable state by re-adjusting their functioning to return to equilibrium after natural or anthropogenic stressful events (e.g., decline in population abundances, introduction of invasive species, species extinction, introduction of pollutants, land use and habitat fragmentation, and climate change) [125,140,141]. Thus, an ecosystem’s equilibrium state is not static. Rather, it is the result of a dynamic time-dependent state (such as a limit cycle in population abundances or a chaotic attractor), which provides adaptability benefits in the face of perturbation. Ecosystems can, thus, display multiple states at specific periods of time, and are considered dynamically stable [142]. Local stability occurs when (all considered properties of) the ecosystem return (in the considered time scale) to their initial state after small disturbances or the new state presents reduced deviations, but alternative (attractor) states exist; and global stability occurs when an equilibrium state is maintained after wide-ranging perturbations [140]. At broad spatial and temporal scales, shifts between the alternative (attractor) states may result in landscape transformation or in sustained mosaics of states. Either way, local short-term instability may drive biodiversity at larger scales [3], paradoxically resulting in higher stability at either higher spatial or temporal scales. The notion of stability needs further refinement, and it may encompass five core properties [31,37,140,143,144,145]: variability—the long-term variability of ecosystem properties in relation to environmental fluctuations; persistence or constancy—capacity to maintain constant characteristics through time; resistance—the ability to preserve primary features under disturbance; resilience—the ability to absorb changes of state variables under disturbance; and robustness—endurance of ecosystem integrity combining resistance with resilience to recover from displacement. Robustness is considered by some authors as more appropriate to define stability in terms of ecosystem functioning [31]. Clearly, all these concepts need to be connected to a specific spatial and temporal scale, which would clarify the still-active debate (see below). The multiple components of stability are related, having divergent connections with biodiversity, and acting at different levels of the biological hierarchy (population, community, and ecosystem) and spatiotemporal scales [21,37,140,144].
The biodiversity–stability relationship has been the subject of a long debate [25,37,41,79,80,81,125,140,141,144,145,146,147,148,149,150,151,152]. Although the biodiversity–stability relationship is not yet fully resolved, experimental and theoretical advances have indicated that biodiversity may promote the stability of aggregate ecosystem functioning, and concomitantly destabilize species abundances, which increases robustness [5,25,31,42,79,81,151,152,153,154]. Fluctuations at the population level result in asynchronous responses of individual species to environmental variation, the rate at which species react to distress, and the reduction in competitive interactions [81,152]. Accordingly, increased biodiversity incorporates species at the community level that will perform a wide range of functions (i.e., the role and functional attributes of each species within the ecosystem) and will respond differently to changes enhancing overall ecosystem robustness [31] and, thus, stability [37,38,80]. This multiplicity contributes to a variable adjustment (variability) of aggregate ecosystem functioning while maintaining a dynamic equilibrium state through time and space (persistence/constancy) in the face of environmental fluctuations, and enhances robustness, i.e., resistance and resilience to disturbance (e.g., invasion, drought, fire, acute climate events, nutrient overloads, disease, etc.) [31,37,155,156,157].
The relationship between stability and diversity is intrinsically related to the ecosystem processes described previously and may be modulated by a variety of mechanisms that drive biological insurance [5,25,37,50,76,83,129,144,146,155,156,158,159,160] on ecosystem functioning. For instance, productivity increases over time through biological insurance, i.e., increased biodiversity protects ecosystem aggregate properties against environmental oscillations through differential responses of each species in the community [63,94,97,103,106,108,109]. This is primarily explained by ecosystem feed backs such as nutrient cycling over space and time [65,94], which contribute to the build-up of common pool resources. Interspecific complementarity and facilitation [63,73,95,98,101,161,162] result in different and unsynchronized responses by diverse component species to nutrient and ecological network fluctuations. These generate compensatory dynamics through negative covariance, adjusting ecosystem functioning to variability and fostering persistence [5,81,163,164]. This asynchrony improves robustness to environmental and biological disturbances by balancing bottom-up and top-down effects [76,81,131,143,165], which may also be enhanced by the presence of more stable and resistant species in diverse communities [26,76,81]. By favoring individual adaptation, cooperative interactions, collective behaviors, and complexity through strong and weak links, biodiverse ecological networks lead to aggregate ecosystem robustness [15,24,163]. Thus, biodiversity ensures manifold biotic interactions and distinct functional attributes that interact with abiotic factors to elicit multiple dynamic feed backs. These prompt complex adaptive responses to environmental heterogeneity and ecological variation that minimize the propagation of stressful events, encompassing ecosystem functioning stability across scales [15,21,24,143,157,163]. Redundancy is another important factor of stability related to biological insurance, as functionally identical species will ensure that a particular ecosystem process will overcome perturbance [16,28,31,83,94,151,159,163]. However, interspecific complementarity may increase through space and time due to both evolutionary and ecological adaptations, decreasing redundancy in highly diverse assemblages as species will perform more unique functions [86,91,94,146].
To assess the relationship between biodiversity and stability, it is crucial to adopt a CAS lens and compartmentalize the notions of stability into responses at different levels of functional and spatiotemporal aggregations (e.g., the discussion presented above between stability in species abundance vs. stability of ecosystem functioning). Moreover, since ecosystems are adaptive, it is critical to develop notions of stability at a set scale and period, not only in terms of the magnitude of the perturbations but, more importantly, in terms of the frequency of these perturbations and the rate of their magnitude change.
5. Biodiversity and Ecosystem Multifunctionality
Ecosystem functioning combines various biological and environmental interactions, sustaining multiple simultaneous ecological processes—multifunctionality—that will translate into services essential for humanity [4,72,84,88,166]. Since biodiversity regulates different ecosystem dimensions, i.e., ecosystem structure and related processes [4], it is fundamental to uncover how it strengthens multifunctionality [84,85,88,167]. This will enhance our ability to predict the persistence and robustness of complex adaptive ecosystems in a fast-changing world.
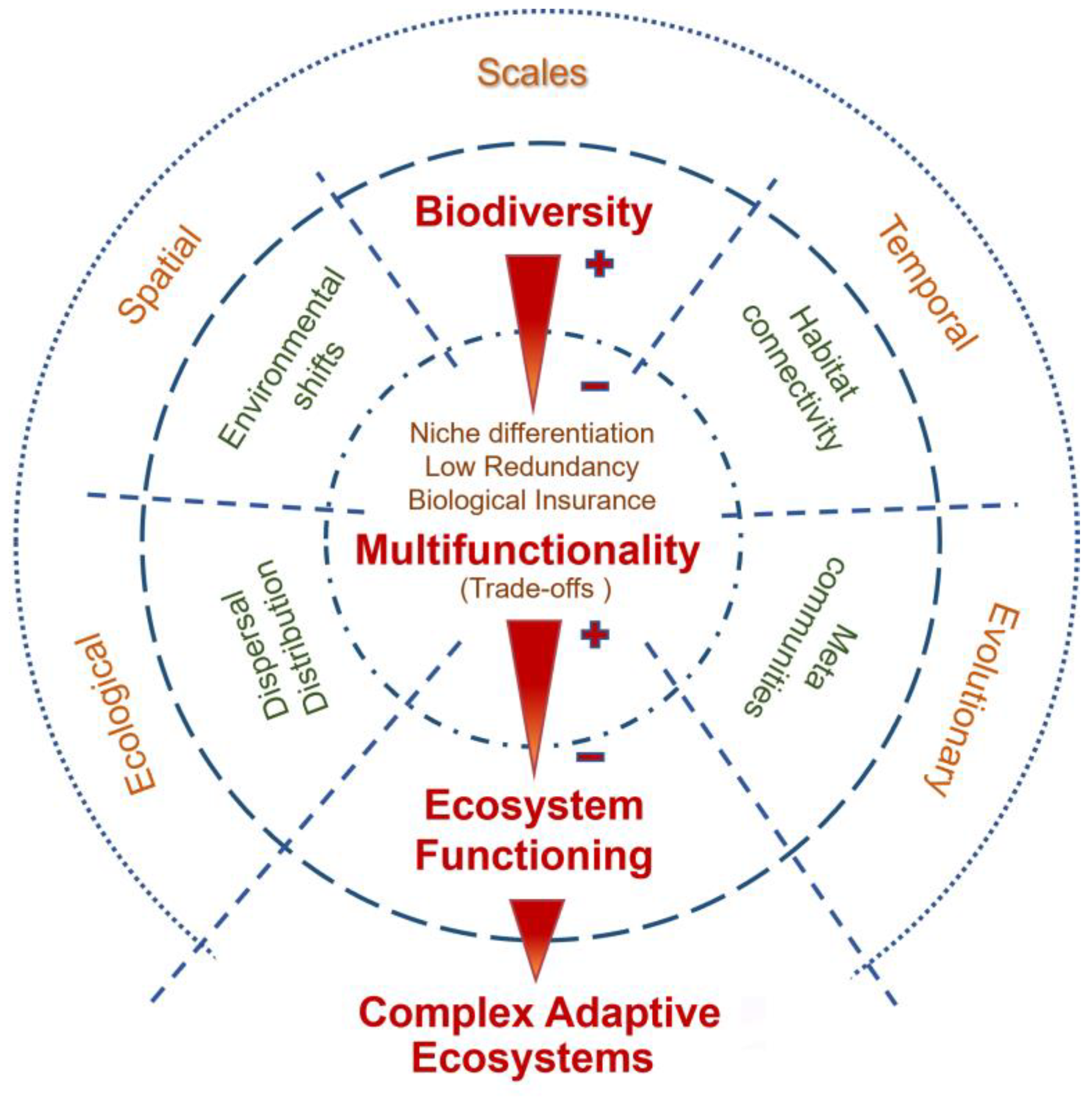
Initial studies on BEF typically focused on unidimensional effects of biodiversity on single processes and ecological levels, providing conflicting results. However, a framework of recent research using multifunctional BEF methodologies [5,72,78,82,84,85,86,88,166,167,168,169,170] has systematically shown a positive relationship between biodiversity and the overall ecosystem functioning. In fact, growing evidence suggests that comprehending how biodiversity mediates the functioning of natural ecosystems entails the integration (Figure 3) of multiple ecological processes, different levels of biodiversity, various environmental contexts, habitat connectivity and dispersal rates, diverse spatiotemporal and ecological–evolutionary scales, and the interaction among all these factors [18,20,66,90,166].
Biodiversity is essential to support dynamic adaptive ecosystem multifunctionality at different temporal and spatial scales within variable environmental contexts. Increased biodiversity sustains cumulative multifunctionality across local habitats and landscapes through niche partitioning, low redundancy, and biological insurance over multiple seasonal and time-based periods [79,82,83,85,86,88,166,167,168,169,170], especially when environmental conditions are highly variable [38,88,166,170]. Ecosystem multifunctionality is fostered by biodiversity since different species frequently affect distinct processes; one species may contribute to several ecological processes at the same time; and these processes may be influenced asymmetrically by various species [85,86,88,170]. In a multifunctional ecosystem, increased biodiversity contributes to an array of different processes because each species becomes unique, i.e., with reduced functional overlap, strengthening ecosystem multifunctionality [82,84,85,86]. Reduced redundancy supports increasing ecosystem complexity through niche differentiation among species with different functional traits [82,85,86,166,169,171]. Indeed, the degree to which ecosystem functioning changes after biodiversity shifts strongly depends on the functional traits that are gained by or made extinct in the ecological system, i.e., the functional role of the new or extinct species [5,38,172]. Different functional traits, either individually or interactively, have the capacity to directly and/or indirectly convey alternative pathways to fully achieve ecosystem multifunctionality [72,82,86,167]. Dispersal is also a key process in temporal and spatial multifunctionality [170]. By tracking local environmental shifts, species reshape their distribution and link metacommunities through connected habitats, upholding high biodiversity levels and overall ecosystem functioning [20,22,90,146,170].
The strength of the positive effects of biodiversity on ecosystem multifunctionality depends on the trade-offs between negative and additive/synergistic intra/interspecific contributions that simultaneously affect multiple processes [78,82,88,166,170,173]. Since different species may enhance different processes, biodiversity levels promoting multifunctionality will increase ecosystem complexity and adaptability through, for example, variability, persistence, resistance to invasions and/or disease, resilience to perturbation and, thus, ecosystem robustness and stability [31,82,86,88,166].
6. Ecological Management—A BEF Approach Adopting CAS Thinking
A paramount achievement of the BEF research is that it provides a comprehensive background to assess the effects of biodiversity loss and homogenization of biota triggered by humans and a supporting framework for ecological management [14,37,62,174]. Management strategies for the sustainable use of ecosystems are difficult to implement because the structural and functional complexities of ecosystems call for multiple and interdisciplinary methodologies to study and model them. In this sense, embracing CAS thinking for the BEF approach provides a tool for explicit management, focusing on sustainability, conservation, and restoration problems. Doing so requires detailed knowledge of specific ecosystem features to solve key management challenges [28].
An integrated understanding of the properties that convey CAS is essential to implement effective ecosystem management and informed policy decisions. The challenge is to develop management strategies that simultaneous acknowledge trade-offs relating to fundamental attributes of both BEF and CAS to avoid unintended consequences, and accepting that interventions are prone to providing new knowledge that needs to be integrated into ecological management. Taking this into account will ensure the ability of ecosystems to adapt to changing environmental conditions while performing well under existing conditions and providing sustainable ecosystem services [28].
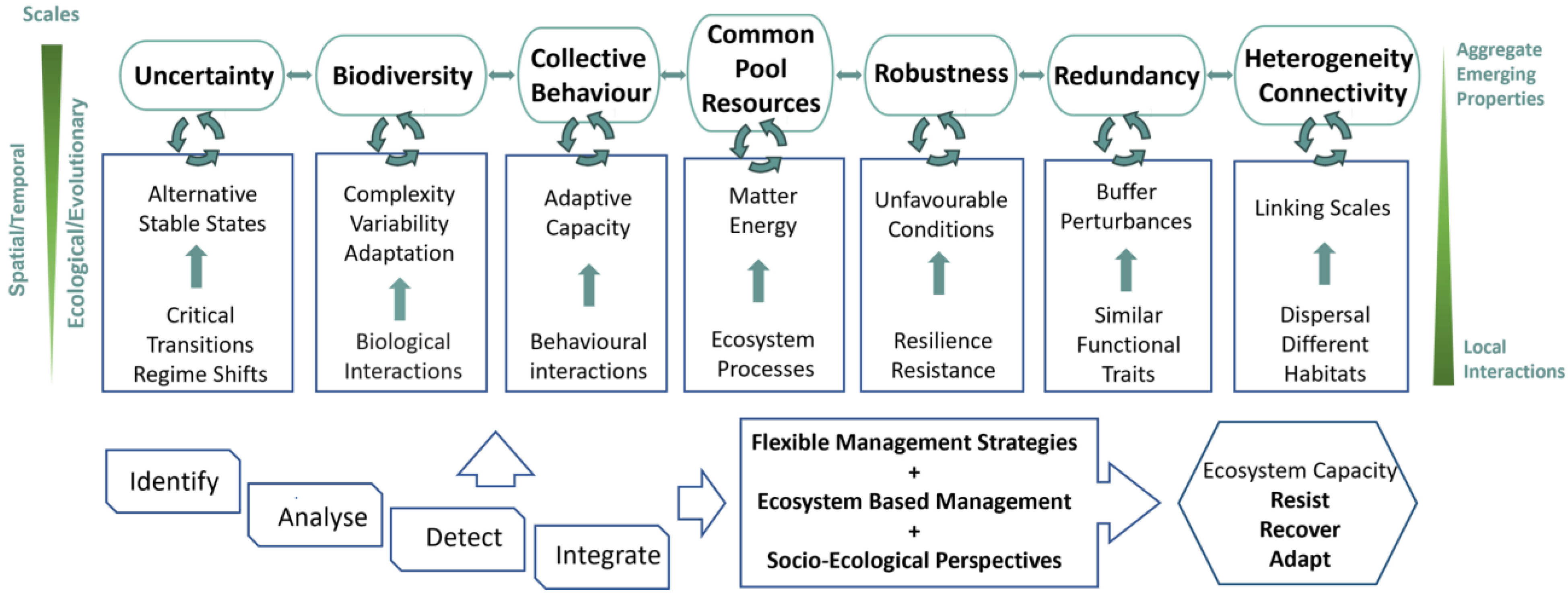
We propose an exploratory framework for ecological management that integrates BEF attributes taking into account CAS properties (Figure 4). Ecological management encompasses the identification and analysis of local interactions and processes. These include critical transitions and regime shifts, biological interactions, behavioral interactions, ecosystem processes, resistance and resilience, species losses, dispersal, and different habitats. The goal is to detect emergent patterns of key interconnected ecosystem properties common in CAS (uncertainty, biodiversity, collective behavior, common pool resources, robustness, redundancy, heterogeneity, and connectivity) across spatiotemporal scales and different levels of biological and abiotic complexity. Critical transitions and regime shifts between alternative stable states reveal the level of uncertainty. Biological components and their interactions convey variability, complexity, and adaptation to sustain biodiversity. Behavioral interactions within and among species enhance adaptation, resulting in collective behavior. Ecosystem processes ensure the flow of matter and energy, producing common pool resources. Resistance and resilience to unfavorable conditions confer robustness. Similar functional species may buffer species losses, contributing to redundancy. Species dispersal and different habitats enable individuals and species to interact and track favorable conditions, conveying environmental heterogeneity and connectivity among different scales. Accounting for fast ecological and slow evolutionary processes by analyzing ecological and co-evolutionary features is also important. Spatial scales and different levels of organization need to be considered due to spatial heterogeneity and feed backs between local and emergent ecosystem patterns. The role of local interactions and aggregate emerging properties, such as ecosystem functioning, must be acknowledged. Management strategies need to be flexible and ecosystem-based, and to integrate socio-ecological perspectives that leverage and foster the capacity of ecosystems to resist, recover, and adapt.
This framework can be used to develop sustainable management plans, conservation prioritization, ecosystem restoration, climate change adaptation, and effective policies. For example, recognizing the influence of biodiversity on ecosystem functioning highlights the importance of maintaining biodiversity for the sustainable provision of ecosystem services such as food production, water purification, climate regulation, and cultural activities. Sustainable management practices can integrate biodiversity through the lens of CAS into decision making to ensure the long-term sustainability of natural resources and the well-being of human communities that rely on these resources. Conservation strategies can focus on protecting heterogenous areas with high biodiversity, supporting essential ecosystem processes that result in collective behavior and the production of common pool resources, which provide essential ecosystem services. Moreover, considering the functional traits of species and their interactions within complex adaptive systems allows for more targeted conservation actions, ensuring robustness, redundancy, and connectivity in the preservation of key ecological functions. By considering diversity, restoration projects can aim to reestablish functions and ecosystem processes that have been lost or disrupted. Restoring biodiverse assemblages can help rebuild trophic interactions, nutrient cycling, and other essential ecosystem processes, enhancing robustness and the functioning of degraded ecosystems. Understanding the connection between biodiversity and ecosystem functioning can inform climate change adaptation strategies. Sustainable management, conservation, and restoration practices can focus on maintaining and enhancing biodiversity to support the adaptive capacity of ecosystems. Diverse ecosystems are more likely to include species with a range of adaptive traits, allowing them to respond to changing environmental conditions and reducing their vulnerability and the services they provide. Finally, understanding the connection between biodiversity and ecosystem functioning within complex adaptive systems provides a scientific basis for policy development. Policies can be informed by the need to protect and restore biodiversity to maintain ecosystem resilience and functioning. Additionally, incorporating the complexity and dynamics of ecosystems into policy frameworks can help address emerging challenges and promote adaptive management approaches that consider the interplay between biodiversity, ecosystem functioning, and human activities.
This framework is designed to combine with multi-factor empirical studies that collect pre-and post-intervention information, as well as with analytical and computational models of ecological dynamics and driving forces of change over ecological and evolutionary scales [15,175] considered in the intervention. The models need then to be confronted with the new post intervention data and updated, either in terms of their recalibration and extension, or simply by clarifying their scope. Using complementary approaches will help to keep a diverse portfolio of increasingly valuable tools. Knowledge graphs [176] present a promising tool to organize and connect such diverse information on data, models, and interventions. These will lead to the identification of various types of feed back among natural ecological systems, anthropogenic pressures, and the evolutionary trends of different scenarios. However, advancing fully cohesive information and delivering science-based tools to back up management plans interconnected with biodiversity and ecosystem sustainability, conservation, and restoration is still a challenge as it requires multidisciplinary and multi-method teams.
7. Biodiversity Loss, Hindered Ecosystem Functioning, and CAS
Research undertaken in recent decades has highlighted the pivotal role of biodiversity in the functioning of ecosystems (see sections above), and the consequences of biodiversity loss for ecosystems and humanity. These studies have indicated that net biodiversity loss threatens multiple ecosystem processes. Therefore, we know that impaired ecosystem functioning will exacerbate with biodiversity loss [5]. Nonetheless, and in spite of various international commitments (e.g., CBD, Aichi Targets, IPBES, SDGs), biodiversity is still experiencing increasing rates of decline and ecosystems are facing ongoing oversimplification worldwide due to human-driven environmental pressures. Using natural resources is vital for human survival and development, but paradoxically the increasing exploitation of ecosystems is severely reducing global biodiversity with detrimental effects on the same resources that humans depend on to survive.
Current knowledge stresses the importance of BEF to comprehend the potential impacts of biodiversity changes on ecosystem functioning, especially to detect the adverse effects on ecosystems of immediate interest to humans and the major trends in global biodiversity loss and the homogenization of biota worldwide. The BEF connection secures complex adaptive ecosystems, as ecosystems emerge through biodiverse interactive communities within changing environmental contexts that simultaneously mediate multiple ecological processes across scales. Increasing biodiversity loss and ecosystem degradation will compromise the fluxes of energy, matter, and information, and thus the robustness and adaptive capacity of ecosystems to endure changes and support the Earth’s system that is so vital to humanity.
8. Conclusions
Understanding how biodiversity relates to ecosystem functioning is still the main focus of the BEF research [4,18,38]. BEF is one of the most critical challenges for ecological science because complex interactions and adaptation play a crucial role in ecosystem functioning [5,38]. Field, experimental, and theoretical studies on BEF have consistently indicated a general trend on the positive link between biodiversity and ecosystem functioning throughout terrestrial, freshwater, and marine ecosystems. The BEF relationship simultaneously regulates multiple dynamic processes, leading to emergent properties across different spatial and temporal scales, as well as ecological and evolutionary scales. Ecosystems are CASs that rely on biodiverse interactive local assemblages within variable environmental conditions, habitat connectivity, and dispersal rates. These components interact and evolve to foster fluxes of matter, energy, and information through nonlinear, basic processes. Biodiversity interacts with climate, resource availability, and disturbance regimes at different scales to configure ecosystem functioning. By regulating distinct ecosystem processes, biodiversity fosters multifunctionality and sustains complex adaptive ecosystems that are capable of adjusting to a fast-changing planet and providing ecosystem services essential to the survival of humanity.
Ecosystem functioning may be perceived as a macroscopic emerging property, which, from the CAS perspective, must affect smaller scales. Using the CAS perspective, we identify a knowledge gap: the feed backs between macrolevel properties, such as ecosystem functioning, and the adaptive capacity at smaller scales, which renders the adaptive capacity of the macroscopic properties themselves. Disentangling this complexity is a long-term conundrum in the BEF relationship. It requires unraveling the role of biodiversity in the dynamics of aggregate ecological patterns, which result from the interplay of all ecosystem compartments at different scales (spatial versus temporal; ecological versus evolutionary; disaggregated properties versus aggregated), and the role of these patterns in those compartments to ensure the persistence of complex adaptive ecosystems.
Recognizing the potential trade-offs and synergies between biodiversity conservation and ecosystem functioning within complex adaptive systems is essential for developing integrated and sustainable management strategies. Balancing these dynamics requires careful consideration of ecological, social, and economic factors, as well as long-term sustainability goals. It emphasizes the importance of adopting an interdisciplinary and systems-based approach to management and conservation practices. Advances in the combined facets of BEF and CAS are critical to a comprehensive assessment of the effects of biodiversity loss triggered by humans.
9. Future Directions
Further progress on BEF entails a forward-looking strategy towards in-depth field research in real-world ecosystems, at landscape and management scales over long periods of time, comprising the integration of data on several biodiversity metrics (namely genetic diversity, species composition, and functional traits); the link between biodiversity and multiple fundamental ecosystem processes; the adaptation of biodiversity and ecosystem functioning to both natural and non-random anthropogenic ecological fluctuations; and the human-induced forces driving species extinction and ecosystem functioning frailty at different scales. Scaling up experiments to combine the complex biotic and environmental interactions that shape BEF and CAS in natural ecosystems will be an important complement to field studies. This experimentation requires the development of new designs that further account for issues of scale, complexity, and applicability to natural systems. As ecosystems operate at multiple spatial and temporal scales, increasing our knowledge still calls for the acquisition of data revealing the scales at which interactions operate and changes occur [4,66].
Integrating currently available meta-datasets and models with previous and novel observational, experimental, and theoretical approaches will identify gaps and produce new knowledge. Such an approach can emerge in parallel with interventionist conservation efforts that, beyond their primary conservation goal, act as theory testers and partners in building a knowledge database that integrates diverse sets of experiments, models, and interventions. The created reference knowledge should be able to produce increasingly accurate scenarios regarding the connection between biodiversity and ecosystem functioning and, consequently, biosphere preservation. Such an integrated approach will be essential to tackle biodiversity loss, and the functionality and sustainability of the Earth’s complex adaptive ecosystems. It will further expand the application of better-informed solutions in conservation policy and decision making addressing the protection, restoration, and sustainable use of biodiversity and ecosystems by humanity.
Author Contributions
A.M.C.—Conceptualization (lead); Data curation (lead); Formal analysis (equal); Investigation (lead); Methodology (lead); Project administration (lead); Visualization (equal); Writing–original draft (lead); Writing—review and editing (equal). L.F.L.—Conceptualization (supporting); Data curation (supporting); Formal analysis (supporting); Funding acquisition (supporting); Investigation (supporting); Methodology (supporting); Project administration (supporting); Visualization (equal) Writing–original draft (supporting); Writing—review and editing (equal). All authors have read and agreed to the published version of the manuscript.
Funding
This review is a result of a collaborative work initiated during a sabbatical leave by AM Correia at Princeton University (PU) supported by the University of Lisbon (ULisboa), Fundação Para a Ciência e a Tecnologia (FCT) and PU. This work also benefited from support by the Princeton Environmental Institute (PEI). LF Lopes acknowledges the financial support of Fundação para a Ciência e a Tecnologia to GHTM-UID/04413/2020 and LA-REAL–LA/P/0117/2020.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We gratefully acknowledge Vítor Vasconcelos, Sólveig Thorsteinsdóttir and Simon A. Levin for their insightful discussions, constructive comments, and important inputs to improve this manuscript. We also thank Steven Carpenter and the Diversity reviewers for their valuable suggestions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ebenman, B.; Jonsson, T. Using Community Viability Analysis to Identify Fragile Systems and Keystone Species. Trends Ecol. Evol. 2005, 20, 568–575. [Google Scholar] [CrossRef] [PubMed]
- Hooper, D.U.; Adair, E.C.; Cardinale, B.J.; Byrnes, J.E.K.; Hungate, B.A.; Matulich, K.L.; Gonzalez, A.; Duffy, J.E.; Gamfeldt, L.; Connor, M.I. A Global Synthesis Reveals Biodiversity Loss as a Major Driver of Ecosystem Change. Nature 2012, 486, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Levin, S.A. Multiple Scales and the Maintenance of Biodiversity. Ecosystems 2000, 3, 498–506. [Google Scholar] [CrossRef]
- Levin, S.A. Evolution at the Ecosystem Level: On the Evolution of Ecosystem Patterns. Contrib. Sci. 2011, 7, 11–16. [Google Scholar] [CrossRef]
- Isbell, F.; Gonzalez, A.; Loreau, M.; Cowles, J.; Díaz, S.; Hector, A.; MacE, G.M.; Wardle, D.A.; O’Connor, M.I.; Duffy, J.E.; et al. Linking the Influence and Dependence of People on Biodiversity across Scales. Nature 2017, 546, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Essl, F.; Dullinger, S.; Rabitsch, W.; Hulme, P.E.; Pyšek, P.; Wilson, J.R.U.; Richardson, D.M. Historical Legacies Accumulate to Shape Future Biodiversity in an Era of Rapid Global Change. Divers. Distrib. 2015, 21, 534–547. [Google Scholar] [CrossRef]
- Loreau, M.; Naeem, S.; Inchausti, P.; Bengtsson, J.; Grime, J.P.; Hector, A.; Hooper, D.U.; Huston, M.A.; Raffaelli, D.; Schmid, B.; et al. Ecology: Biodiversity and Ecosystem Functioning: Current Knowledge and Future Challenges. Science 2001, 294, 804–808. [Google Scholar] [CrossRef] [Green Version]
- Naeem, S.; Loreau, M.; Inchausti, P. Biodiversity and Ecosystem Functioning: The Emergence of a Synthetic Ecological Framework. In Biodiversity and Ecosystem Functioning: Synthesis and Perspectives; Oxford University Press: Oxford, UK, 2002; pp. 3–11. [Google Scholar]
- Worm, B.; Duffy, J.E. Biodiversity, Productivity and Stability in Real Food Webs. Trends Ecol. Evol. 2003, 18, 628–632. [Google Scholar] [CrossRef]
- Hooper, D.U.; Chapin, F.S.; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of Biodiversity on Ecosystem Functioning: A Consensus of Current Knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Tylianakis, J.M.; Rand, T.A.; Kahmen, A.; Klein, A.M.; Buchmann, N.; Perner, J.; Tscharntke, T. Resource Heterogeneity Moderates the Biodiversity-Function Relationship in Real World Ecosystems. PLoS Biol. 2008, 6, 947–956. [Google Scholar] [CrossRef] [Green Version]
- Duffy, J.E.; Cardinale, B.J.; France, K.E.; McIntyre, P.B.; Thébault, E.; Loreau, M. The Functional Role of Biodiversity in Ecosystems: Incorporating Trophic Complexity. Ecol. Lett. 2007, 10, 522–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balvanera, P.; Siddique, I.; Dee, L.; Paquette, A.; Isbell, F.; Gonzalez, A.; Byrnes, J.; O’Connor, M.I.; Hungate, B.A.; Griffin, J.N. Linking Biodiversity and Ecosystem Services: Current Uncertainties and the Necessary next Steps. Bioscience 2014, 64, 49–57. [Google Scholar] [CrossRef]
- Tilman, D.; Isbell, F.; Cowles, J.M. Biodiversity and Ecosystem Functioning. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 471–493. [Google Scholar] [CrossRef]
- Hagstrom, G.I.; Levin, S.A. Marine Ecosystems as Complex Adaptive Systems: Emergent Patterns, Critical Transitions, and Public Goods. Ecosystems 2017, 20, 458–476. [Google Scholar] [CrossRef]
- Levin, S.A. Ecosystems and the Biosphere as Complex Adaptive Systems. Ecosystems 1998, 1, 431–436. [Google Scholar] [CrossRef] [Green Version]
- Levin, S.A. Self-Organization and the Emergence of Complexity in Ecological Systems. Bioscience 2005, 55, 1075. [Google Scholar] [CrossRef] [Green Version]
- Poisot, T.; Mouquet, N.; Gravel, D. Trophic Complementarity Drives the Biodiversity-Ecosystem Functioning Relationship in Food Webs. Ecol. Lett. 2013, 16, 853–861. [Google Scholar] [CrossRef] [PubMed]
- Sentis, A.; Gémard, C.; Jaugeon, B.; Boukal, D.S. Predator Diversity and Environmental Change Modify the Strengths of Trophic and Nontrophic Interactions. Glob. Chang. Biol. 2017, 23, 2629–2640. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.; Germain, R.M.; Srivastava, D.S.; Filotas, E.; Dee, L.E.; Gravel, D.; Thompson, P.L.; Isbell, F.; Wang, S.; Kéfi, S.; et al. Scaling-up Biodiversity-ecosystem Functioning Research. Ecol. Lett. 2020, 23, 757–776. [Google Scholar] [CrossRef] [Green Version]
- Qiao, X.; Hautier, Y.; Geng, Y.; Wang, S.; Wang, J.; Zhang, N.; Zhang, Z.; Zhang, C.; Zhao, X.; von Gadow, K. Biodiversity Contributes to Stabilizing Ecosystem Productivity across Spatial Scales as Much as Environmental Heterogeneity in a Large Temperate Forest Region. For. Ecol. Manag. 2023, 529, 120695. [Google Scholar] [CrossRef]
- Norberg, J. Biodiversity and Ecosystem Functioning: A Complex Adaptive Systems Approach. Limnol. Oceanogr. 2004, 49, 1269–1277. [Google Scholar] [CrossRef]
- Levin, S.A. Learning to Live in a Global Commons: Socioeconomic Challenges for a Sustainable Environment. Ecol. Res. 2006, 21, 328–333. [Google Scholar] [CrossRef]
- Levin, S.A. Fragile Dominion: Complexity and the Commons; Perseus Books: Cambridge, MA, USA, 1999. [Google Scholar]
- Tilman, D. The Ecological Consequences of Changes in Biodiversity: A Search for General Principles. Ecology 1999, 80, 1455–1474. [Google Scholar] [CrossRef]
- Stachowicz, J.J.; Bruno, J.F.; Duffy, J.E. Understanding the Effects of Marine Biodiversity on Communities and Ecosystems. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 739–766. [Google Scholar] [CrossRef] [Green Version]
- Solé, R.; Levin, S. Ecological Complexity and the Biosphere: The next 30 Years. Philos. Trans. R. Soc. B Biol. Sci. 2022, 377. [Google Scholar] [CrossRef] [PubMed]
- Levin, S.; Xepapadeas, T.; Crépin, A.-S.; Norberg, J.; de Zeeuw, A.; Folke, C.; Hughes, T.; Arrow, K.; Barrett, S.; Daily, G.; et al. Social-Ecological Systems as Complex Adaptive Systems: Modeling and Policy Implications. Environ. Dev. Econ. 2013, 18, 111–132. [Google Scholar] [CrossRef] [Green Version]
- Petraitis, P.S.; Dudgeon, S.R. Detection of Alternative Stable States in Marine Communities. J. Exp. Mar. Bio. Ecol. 2004, 300, 343–371. [Google Scholar] [CrossRef]
- Fogarty, M.J.; Gamble, R.; Perretti, C.T. Dynamic Complexity in Exploited Marine Ecosystems. Front. Ecol. Evol. 2016, 4, 68. [Google Scholar] [CrossRef] [Green Version]
- Levin, S.A.; Lubchenco, J. Resilience, Robustness, and Marine Ecosystem-Based Management. Bioscience 2008, 58, 27–32. [Google Scholar] [CrossRef] [Green Version]
- Naeem, S.; Bunker, D.E.; Hector, A.; Loreau, M.; Perrings, C. Biodiversity, Ecosystem Functioning and Human Wellbeing: An Ecological and Economic Perspective; Oxford University Press: Oxford, UK, 2009; ISBN 9780199547951. [Google Scholar]
- Naeem, S. Disentangling the Impacts of Diversity on Ecosystem Functioning in Combinatorial Experiments. Ecology 2002, 83, 2925–2935. [Google Scholar] [CrossRef]
- Balvanera, P.; Pfisterer, A.B.; Buchmann, N.; He, J.S.; Nakashizuka, T.; Raffaelli, D.; Schmid, B. Quantifying the Evidence for Biodiversity Effects on Ecosystem Functioning and Services. Ecol. Lett. 2006, 9, 1146–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loreau, M. Linking Biodiversity and Ecosystems: Towards a Unifying Ecological Theory. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 49–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naeem, S.; Bunker, D.E.; Hector, A.; Loreau, M.; Perrings, C. Introduction: The Ecological and Social Implications of Changing Biodiversity. An Overview of a Decade of Biodiversity and Ecosystem Functioning Research. In Biodiversity, Ecosystem Functioning, and Human Wellbeing; Naeem, S., Bunker, D.E., Hector, A., Loreau, M., Perrings, C., Eds.; Oxford University Press: Oxford, UK, 2009; pp. 3–13. ISBN 9780191720345. [Google Scholar]
- Cardinale, B.J.; Srivastava, D.S.; Duffy, J.E.; Wright, J.P.; Downing, A.L.; Sankaran, M.; Jouseau, C. Effects of Biodiversity on the Functioning of Trophic Groups and Ecosystems. Nature 2006, 443, 989–992. [Google Scholar] [CrossRef] [PubMed]
- Cardinale, B.J.; Duffy, J.E.; Gonzalez, A.; Hooper, D.U.; Perrings, C.; Venail, P.; Narwani, A.; Mace, G.M.; Tilman, D.; Wardle, D.A.; et al. Biodiversity Loss and Its Impact on Humanity. Nature 2012, 486, 59–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- deLaplante, K.; Picasso, V. The Biodiversity-Ecosystem Function Debate in Ecology. Philos. Ecol. 2011, 11, 169–200. [Google Scholar] [CrossRef]
- Naeem, S.; Thompson, L.J.; Lawler, S.P.; Lawton, J.H.; Woodfin, R.M. Declining Biodiversity Can Alter the Performance of Ecosystems. Nature 1994, 368, 734–737. [Google Scholar] [CrossRef]
- Tilman, D.; Downing, J.A. Biodiversity and Stability in Grasslands. Nature 1994, 367, 363–365. [Google Scholar] [CrossRef]
- Tilman, D.; Wedin, D.; Knops, J. Productivity and Sustainability Influenced by Biodiversity in Grassland Ecosystems. Nature 1996, 379, 718–720. [Google Scholar] [CrossRef]
- Tilman, D.; Knops, J.; Wedin, D.; Reich, P.; Ritchie, M.; Siemann, E. The Influence of Functional Diversity and Composition on Ecosystem Processes. Science 1997, 277, 1300–1302. [Google Scholar] [CrossRef] [Green Version]
- Díaz, S.; Cabido, M. Plant Functional Types and Ecosystem Function in Relation to Global Change. J. Veg. Sci. 1997, 8, 463–474. [Google Scholar] [CrossRef]
- Hooper, D.U.; Vitousek, P.M. The Effects of Plant Composition and Diversity on Ecosystem Processes. Science 1997, 277, 1302–1305. [Google Scholar] [CrossRef] [Green Version]
- Hector, A.; Schmid, B.; Beierkuhnlein, C.; Caldeira, M.C.; Diemer, M.; Dimitrakopoulos, P.G.; Finn, J.A.; Freitas, H.; Giller, P.S.; Good, J.; et al. Plant Diversity and Productivity Experiments in European Grasslands. Science 1999, 286, 1123–1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wardle, D.A.; Bonner, K.I.; Nicholson, K.S. Biodiversity and Plant Litter: Experimental Evidence Which Does Not Support the View That Enhanced Species Richness Improves Ecosystem Function. Oikos 1997, 79, 247. [Google Scholar] [CrossRef]
- Loreau, M. Separating Sampling and Other Effects in Biodiversity Experiments. Oikos 1998, 82, 600–602. [Google Scholar] [CrossRef]
- Loreau, M. Biodiversity and Ecosystem Functioning: A Mechanistic Model. Proc. Natl. Acad. Sci. USA 1998, 95, 5632–5636. [Google Scholar] [CrossRef] [PubMed]
- Loreau, M. Biodiversity and Ecosystem Functioning: Recent Theoretical Advances. Oikos 2000, 91, 3–17. [Google Scholar] [CrossRef] [Green Version]
- Cardinale, B.J.; Nelson, K.; Palmer, M.A. Linking Species Diversity to the Functioning of Ecosystems: On the Importance of Environmental Context. Oikos 2000, 91, 175–183. [Google Scholar] [CrossRef] [Green Version]
- Chesson, P. Mechanisms of Maintenance of Species Diversity. Annu. Rev. Ecol. Syst. 2000, 31, 343–366. [Google Scholar] [CrossRef] [Green Version]
- Lehman, C.L.; Tilman, D. Biodiversity, Stability, and Productivity in Competitive Communities. Am. Nat. 2000, 156, 534–552. [Google Scholar] [CrossRef]
- Doak, D.F.; Bigger, D.; Harding, E.K.; Marvier, M.A.; O’Malley, R.E.; Thomson, D. The Statistical Inevitability of Stability-Diversity Relationships in Community Ecology. Am. Nat. 1998, 151, 264–276. [Google Scholar] [CrossRef]
- Duffy, J.E. Biodiversity Loss, Trophic Skew and Ecosystem Functioning. Ecol. Lett. 2003, 6, 680–687. [Google Scholar] [CrossRef] [Green Version]
- Covich, A.P.; Austen, M.C.; Bärlocher, F.; Chauvet, E.; Cardinale, B.J.; Biles, C.L.; Inchausti, P.; Dangles, O.; Solan, M.; Gessner, M.O.; et al. The Role of Biodiversity in the Functioning of Freshwater and Marine Benthic Ecosystems. Bioscience 2004, 54, 767. [Google Scholar] [CrossRef]
- Finke, D.L.; Denno, R.F. Predator Diversity and the Functioning of Ecosystems: The Role of Intraguild Predation in Dampening Trophic Cascades. Ecol. Lett. 2005, 8, 1299–1306. [Google Scholar] [CrossRef]
- Srivastava, D.S.; Vellend, M. Biodiversity-Ecosystem Function Research: Is It Relevant to Conservation? Annu. Rev. Ecol. Evol. Syst. 2005, 36, 267–294. [Google Scholar] [CrossRef] [Green Version]
- Cardinale, B.J.; Wright, J.P.; Cadotte, M.W.; Carroll, I.T.; Hector, A.; Srivastava, D.S.; Loreau, M.; Weis, J.J. Impacts of Plant Diversity on Biomass Production Increase through Time Because of Species Complementarity. Proc. Natl. Acad. Sci. USA 2007, 104, 18123–18128. [Google Scholar] [CrossRef]
- Hector, A.; Joshi, J.; Scherer-Lorenzen, M.; Schmid, B.; Spehn, E.M.; Wacker, L.; Weilenmann, M.; Bazeley-White, E.; Beierkuhnlein, C.; Caldeira, M.C.; et al. Biodiversity and Ecosystem Functioning: Reconciling the Results of Experimental and Observational Studies. Funct. Ecol. 2007, 21, 998–1002. [Google Scholar] [CrossRef] [Green Version]
- Cardinale, B.J.; Bennett, D.M.; Nelson, C.E.; Gross, K. Does Productivity Drive Diversity or Vice Versa? A Test of the Multivariate Productivity-Diversity Hypothesis in Streams. Ecology 2009, 90, 1227–1241. [Google Scholar] [CrossRef]
- Huston, M.A. Hidden Treatments in Ecological Experiments: Re-Evaluating the Ecosystem Function of Biodiversity. Oecologia 1997, 110, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Schläpfer, F.; Schmid, B. Ecosystem Effects of Biodiversity: A Classification of Hypotheses and Exploration of Empirical Results. Ecol. Appl. 1999, 9, 893–912. [Google Scholar] [CrossRef]
- Downing, A.L.; Leibold, M.A. Ecosystem Consequences of Species Richness and Composition in Pond Food Webs. Nature 2002, 416, 837–841. [Google Scholar] [CrossRef] [PubMed]
- Díaz, S.; Symstad, A.J.; Chapin, F.S.; Wardle, D.A.; Huenneke, L.F. Functional Diversity Revealed by Removal Experiments. Trends Ecol. Evol. 2003, 18, 140–146. [Google Scholar] [CrossRef]
- Genung, M.A.; Fox, J.; Winfree, R. Species Loss Drives Ecosystem Function in Experiments, but in Nature the Importance of Species Loss Depends on Dominance. Glob. Ecol. Biogeogr. 2020, 29, 1531–1541. [Google Scholar] [CrossRef]
- Yan, P.; Fernández-Martínez, M.; Van Meerbeek, K.; Yu, G.; Migliavacca, M.; He, N. The Essential Role of Biodiversity in the Key Axes of Ecosystem Function. Glob. Chang. Biol. 2023, 29, 4569–4585. [Google Scholar] [CrossRef]
- Cardinale, B.J.; Matulich, K.L.; Hooper, D.U.; Byrnes, J.E.; Duffy, E.; Gamfeldt, L.; Balvanera, P.; O’Connor, M.I.; Gonzalez, A. The Functional Role of Producer Diversity in Ecosystems. Am. J. Bot. 2011, 98, 572–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isbell, F.; Calcagno, V.; Hector, A.; Connolly, J.; Harpole, W.S.; Reich, P.B.; Scherer-Lorenzen, M.; Schmid, B.; Tilman, D.; Van Ruijven, J.; et al. High Plant Diversity Is Needed to Maintain Ecosystem Services. Nature 2011, 477, 199–202. [Google Scholar] [CrossRef]
- Peh, K.S.H.; Lewis, S.L. Conservation Implications of Recent Advances in Biodiversity-Functioning Research. Biol. Conserv. 2012, 151, 26–31. [Google Scholar] [CrossRef]
- Tilman, D.; Reich, P.B.; Isbell, F. Biodiversity Impacts Ecosystem Productivity as Much as Resources, Disturbance, or Herbivory. Proc. Natl. Acad. Sci. USA 2012, 109, 10394–10397. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Van Der Putten, W.H. Belowground Biodiversity and Ecosystem Functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef]
- Cadotte, M.W.; Cardinale, B.J.; Oakley, T.H. Evolutionary History and the Effect of Biodiversity on Plant Productivity. Proc. Natl. Acad. Sci. USA 2008, 105, 17012–17017. [Google Scholar] [CrossRef]
- Cardinale, B.J.; Venail, P.; Gross, K.; Oakley, T.H.; Narwani, A.; Allan, E.; Flombaum, P.; Joshi, J.; Reich, P.B.; Tilman, D.; et al. Further Re-Analyses Looking for Effects of Phylogenetic Diversity on Community Biomass and Stability. Funct. Ecol. 2015, 29, 1607–1610. [Google Scholar] [CrossRef] [Green Version]
- Mora, C.; Danovaro, R.; Loreau, M. Alternative Hypotheses to Explain Why Biodiversity-Ecosystem Functioning Relationships Are Concave-up in Some Natural Ecosystems but Concave-down in Manipulative Experiments. Sci. Rep. 2014, 4, 5427. [Google Scholar] [CrossRef] [Green Version]
- Isbell, F.; Cowles, J.; Dee, L.E.; Loreau, M.; Reich, P.B.; Gonzalez, A.; Hector, A.; Schmid, B. Quantifying Effects of Biodiversity on Ecosystem Functioning across Times and Places. Ecol. Lett. 2018, 21, 763–778. [Google Scholar] [CrossRef] [Green Version]
- Van Der Plas, F. Biodiversity and Ecosystem Functioning in Naturally Assembled Communities. Biol. Rev. 2019, 94, 1220–1245. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Isbell, F.; Deng, W.; Hong, P.; Dee, L.E.; Thompson, P.; Loreau, M. How Complementarity and Selection Affect the Relationship between Ecosystem Functioning and Stability. Ecology 2021, 102, e03347. [Google Scholar] [CrossRef] [PubMed]
- Byrnes, J.E.K.; Gamfeldt, L.; Isbell, F.; Lefcheck, J.S.; Griffin, J.N.; Hector, A.; Cardinale, B.J.; Hooper, D.U.; Dee, L.E.; Emmett Duffy, J. Investigating the Relationship between Biodiversity and Ecosystem Multifunctionality: Challenges and Solutions. Methods Ecol. Evol. 2014, 5, 111–124. [Google Scholar] [CrossRef] [Green Version]
- Hooper, D.U.; Solan, M.; Symstad, A.; Diaz, S.; Gessner, M.O.; Buchmann, N.; Degrange, V.; Grime, P.; Hulot, F.; Mermillod-Blondin, F.; et al. Species Diversity, Functional Diversity, and Ecossystem Functioning. In Biodiversity and Ecosystem Functioning; Loreau, M., Naeem, S., Inchausti, P., Eds.; Oxford University Press: Oxford, UK, 2002; pp. 195–386, 0198515707. [Google Scholar]
- Loreau, M.; de Mazancourt, C. Biodiversity and Ecosystem Stability: A Synthesis of Underlying Mechanisms. Ecol. Lett. 2013, 16, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Lefcheck, J.S.; Byrnes, J.E.K.; Isbell, F.; Gamfeldt, L.; Griffin, J.N.; Eisenhauer, N.; Hensel, M.J.S.; Hector, A.; Cardinale, B.J.; Duffy, J.E. Biodiversity Enhances Ecosystem Multifunctionality across Trophic Levels and Habitats. Nat. Commun. 2015, 6, 6936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loreau, M.; Barbier, M.; Filotas, E.; Gravel, D.; Isbell, F.; Miller, S.J.; Montoya, J.M.; Wang, S.; Aussenac, R.; Germain, R.; et al. Biodiversity as Insurance: From Concept to Measurement and Application. Biol. Rev. 2021, 96, 2333–2354. [Google Scholar] [CrossRef]
- Manning, P.; van der Plas, F.; Soliveres, S.; Allan, E.; Maestre, F.T.; Mace, G.; Whittingham, M.J.; Fischer, M. Redefining Ecosystem Multifunctionality. Nat. Ecol. Evol. 2018, 2, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Hector, A.; Bagchi, R. Biodiversity and Ecosystem Multifunctionality. Nature 2007, 448, 188–190. [Google Scholar] [CrossRef]
- Gamfeldt, L.; Hillebrand, H.; Jonsson, P.R. Multiple Functions Increase the Importance of Biodiversity for Overall Ecosystem Functioning. Ecology 2008, 89, 1223–1231. [Google Scholar] [CrossRef]
- O’Connor, M.I.; Gonzalez, A.; Byrnes, J.E.K.; Cardinale, B.J.; Duffy, J.E.; Gamfeldt, L.; Griffin, J.N.; Hooper, D.; Hungate, B.A.; Paquette, A.; et al. A General Biodiversity–Function Relationship Is Mediated by Trophic Level. Oikos 2017, 126, 18–31. [Google Scholar] [CrossRef] [Green Version]
- Zavaleta, E.S.; Pasari, J.R.; Hulvey, K.B.; Tilman, G.D. Sustaining Multiple Ecosystem Functions in Grassland Communities Requires Higher Biodiversity. Proc. Natl. Acad. Sci. USA 2010, 107, 1443–1446. [Google Scholar] [CrossRef]
- Scherer-Lorenzen, M.; Gessner, M.O.; Beisner, B.E.; Messier, C.; Paquette, A.; Petermann, J.S.; Soininen, J.; Nock, C.A. Pathways for Cross-Boundary Effects of Biodiversity on Ecosystem Functioning. Trends Ecol. Evol. 2022, 37, 454–467. [Google Scholar] [CrossRef]
- Bauer, S.; Hoye, B.J. Migratory Animals Couple Biodiversity and Ecosystem Functioning Worldwide. Science 2014, 344, 1242552. [Google Scholar] [CrossRef] [PubMed]
- Duffy, J.E.; Richardson, J.P.; France, K.E. Ecosystem Consequences of Diversity Depend on Food Chain Length in Estuarine Vegetation. Ecol. Lett. 2005, 8, 301–309. [Google Scholar] [CrossRef]
- Gross, K. Ecology: Biodiversity and Productivity Entwined. Nature 2016, 529, 293–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Chen, H.Y.H.; Taylor, A.R. Positive Species Diversity and Above-Ground Biomass Relationships Are Ubiquitous across Forest Strata despite Interference from Overstorey Trees. Funct. Ecol. 2017, 31, 419–426. [Google Scholar] [CrossRef]
- Reich, P.B.; Tilman, D.; Isbell, F.; Mueller, K.; Hobbie, S.E.; Flynn, D.F.B.; Eisenhauer, N. Impacts of Biodiversity Loss Escalate through Time as Redundancy Fades. Science 2012, 336, 589–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loreau, M.; Sapijankas, J.; Isbell, F.; Hector, A. Comments: Niche and Fitness Differences Relate the Maintenance of Diversity to Ecosystem Function: Comment. Ecology 2012, 93, 1482–1491. [Google Scholar] [CrossRef] [Green Version]
- Worm, B.; Barbier, E.B.; Beaumont, N.; Duffy, J.E.; Folke, C.; Halpern, B.S.; Jackson, J.B.C.; Lotze, H.K.; Micheli, F.; Palumbi, S.R.; et al. Impacts of Biodiversity Loss on Ocean Ecosystem Services. Science 2006, 314, 787–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cowles, J.M.; Wragg, P.D.; Wright, A.J.; Powers, J.S.; Tilman, D. Shifting Grassland Plant Community Structure Drives Positive Interactive Effects of Warming and Diversity on Aboveground Net Primary Productivity. Glob. Chang. Biol. 2016, 22, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Loreau, M.; Hector, A. Partitioning Selection and Complementarity in Biodiversity Experiments. Nature 2001, 412, 72–76. [Google Scholar] [CrossRef] [PubMed]
- Hooper, D.U.; Dukes, J.S. Overyielding among Plant Functional Groups in a Long-Term Experiment. Ecol. Lett. 2004, 7, 95–105. [Google Scholar] [CrossRef]
- Bruno, J.F.; Cardinale, B.J. Cascading Effects of Predator Richness. Front. Ecol. Environ. 2008, 6, 539–546. [Google Scholar] [CrossRef] [Green Version]
- Cardinale, B.J.; Palmer, M.A.; Collins, S.L. Species Diversity Enhances Ecosystem Functioning through Interspecific Facilitation. Nature 2002, 415, 426–429. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D. Biodiversity and Ecosystem Functioning. In Nature´s Services Societal Dependence on Natural Ecosystems; Daily, G.C., Ed.; Inland Press: Washington, DC, USA, 1997; pp. 93–112. [Google Scholar]
- Cardinale, B.J.; Ives, A.R.; Inchausti, P. Effects of Species Diversity on the Primary Productivity of Ecosystems: Extending Our Spatial and Temporal Scales of Inference. Oikos 2004, 104, 437–450. [Google Scholar] [CrossRef]
- Brooks, E.G.E.; Holland, R.A.; Darwall, W.R.T.; Eigenbrod, F. Global Evidence of Positive Impacts of Freshwater Biodiversity on Fishery Yields. Glob. Ecol. Biogeogr. 2016, 25, 553–562. [Google Scholar] [CrossRef] [Green Version]
- Rozenberg, G.S.; Zinchenko, T.D. The Sustainability of Aquatic Ecosystems: An Overview of the Problem. Arid Ecosyst. 2014, 4, 234–243. [Google Scholar] [CrossRef]
- Tilman, D.; Reich, P.B.; Knops, J.; Wedin, D.; Mielke, T.; Lehman, C. Diversity and Productivity in a Long-Term Grassland Experiment. Science 2001, 294, 843–845. [Google Scholar] [CrossRef] [Green Version]
- Spehn, E.M.; Hector, A.; Joshi, J.; Scherer-Lorenzen, M.; Schmid, B.; Bazeley-White, E.; Beierkuhnlein, C.; Caldeira, M.C.; Diemer, M.; Dimitrakopoulos, P.G.; et al. Ecosystem Effects of Biodiversity Manipulations in European Grassland. Ecol. Monogr. 2005, 75, 37–63. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Chen, H.Y.H. Effects of Species Diversity on Fine Root Productivity in Diverse Ecosystems: A Global Meta-Analysis. Glob. Ecol. Biogeogr. 2016, 25, 1387–1396. [Google Scholar] [CrossRef]
- Ma, Z.; Chen, H.Y.H. Effects of Species Diversity on Fine Root Productivity Increase with Stand Development and Associated Mechanisms in a Boreal Forest. J. Ecol. 2017, 105, 237–245. [Google Scholar] [CrossRef]
- Allgeier, J.E.; Burkepile, D.E.; Layman, C.A. Animal Pee in the Sea: Consumer-Mediated Nutrient Dynamics in the World’s Changing Oceans. Glob. Chang. Biol. 2017, 23, 2166–2178. [Google Scholar] [CrossRef]
- Brookes, J.D.; Aldridge, K.; Wallace, T.; Linden, L.; Ganf, G.G. Multiple Interception Pathways for Resource Utilisation and Increased Ecosystem Resilience. Hydrobiologia 2005, 552, 135–146. [Google Scholar] [CrossRef]
- Zou, K.; Thébault, E.; Lacroix, G.; Barot, S. Interactions between the Green and Brown Food Web Determine Ecosystem Functioning. Funct. Ecol. 2016, 30, 1454–1465. [Google Scholar] [CrossRef] [Green Version]
- Naeem, S. Expanding Scales in Biodiversity-Based Research: Challenges and Solution for Marine Systems. Mar. Ecol. Prog. Ser. 2006, 311, 273–283. [Google Scholar] [CrossRef]
- Borer, E.T.; Lind, E.M.; Ogdahl, E.J.; Seabloom, E.W.; Tilman, D.; Montgomery, R.A.; Kinkel, L.L. Food-Web Composition and Plant Diversity Control Foliar Nutrient Content and Stoichiometry. J. Ecol. 2015, 103, 1432–1441. [Google Scholar] [CrossRef]
- Mischler, J.; Johnson, P.T.J.; Mckenzie, V.J.; Townsend, A.R. Parasite Infection Alters Nitrogen Cycling at the Ecosystem Scale. J. Anim. Ecol. 2016, 85, 817–828. [Google Scholar] [CrossRef]
- Handa, I.T.; Aerts, R.; Berendse, F.; Berg, M.P.; Bruder, A.; Butenschoen, O.; Chauvet, E.; Gessner, M.O.; Jabiol, J.; Makkonen, M.; et al. Consequences of Biodiversity Loss for Litter Decomposition across Biomes. Nature 2014, 509, 218–221. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Pan, Q.; Bai, Y.; Hu, S.; Huang, J.; Wang, Q.; Naeem, S.; Elser, J.J.; Wu, J.; Han, X. Effects of Plant Functional Group Loss on Soil Biota and Net Ecosystem Exchange: A Plant Removal Experiment in the Mongolian Grassland. J. Ecol. 2016, 104, 734–743. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Mommer, L.; van Ruijven, J.; de Kroon, H.; Fischer, C.; Gessler, A.; Hildebrandt, A.; Scherer-Lorenzen, M.; Wirth, C.; Weigelt, A. Plant Species Richness Negatively Affects Root Decomposition in Grasslands. J. Ecol. 2017, 105, 209–218. [Google Scholar] [CrossRef]
- Wei, X.; Reich, P.B.; Hobbie, S.E.; Kazanski, C.E. Disentangling Species and Functional Group Richness Effects on Soil N Cycling in a Grassland Ecosystem. Glob. Chang. Biol. 2017, 23, 4717–4727. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, M.; Malmqvist, B. Mechanisms behind Positive Diversity Effects on Ecosystem Functioning: Testing the Facilitation and Interference Hypotheses. Oecologia 2003, 134, 554–559. [Google Scholar] [CrossRef]
- Allen, D.C.; Vaughn, C.C.; Kelly, J.F.; Cooper, J.T.; Engel, M.H. Bottom-up Biodiversity Effects Increase Resource Subsidy Flux between Ecosystems. Ecology 2012, 93, 2165–2174. [Google Scholar] [CrossRef] [PubMed]
- Boyero, L.; Cardinale, B.J.; Bastian, M.; Pearson, R.G. Biotic vs. Abiotic Control of Decomposition: A Comparison of the Effects of Simulated Extinctions and Changes in Temperature. PLoS ONE 2014, 9, e87426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kéfi, S.; Berlow, E.L.; Wieters, E.A.; Navarrete, S.A.; Petchey, O.L.; Wood, S.A.; Boit, A.; Joppa, L.N.; Lafferty, K.D.; Williams, R.J.; et al. More than a Meal… Integrating Non-Feeding Interactions into Food Webs. Ecol. Lett. 2012, 15, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Yen, J.D.L.; Cabral, R.B.; Cantor, M.; Hatton, I.; Kortsch, S.; Patrício, J.; Yamamichi, M. Linking Structure and Function in Food Webs: Maximization of Different Ecological Functions Generates Distinct Food Web Structures. J. Anim. Ecol. 2016, 85, 537–547. [Google Scholar] [CrossRef] [Green Version]
- Landi, P.; Minoarivelo, H.O.; Brännström, Å.; Hui, C.; Dieckmann, U. Complexity and Stability of Ecological Networks: A Review of the Theory. Popul. Ecol. 2018, 60, 319–345. [Google Scholar] [CrossRef]
- Thébault, E.; Loreau, M. The Relationship between Biodiversity and Ecosystem Functioning in Food Webs. Ecol. Res. 2006, 21, 17–25. [Google Scholar] [CrossRef]
- Montoya, J.M.; Pimm, S.L.; Solé, R.V. Ecological Networks and Their Fragility. Nature 2006, 442, 259–264. [Google Scholar] [CrossRef]
- Duffy, J.E. Biodiversity and Ecosystem Function: The Consumer Connection. Oikos 2002, 99, 201–219. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, O.J.; Krivan, V.; Ovadia, O. Trophic Cascades: The Primacy of Trait-Mediated Indirect Interactions. Ecol. Lett. 2004, 7, 153–163. [Google Scholar] [CrossRef] [Green Version]
- Thébault, E.; Loreau, M. Food-Web Constraints on Biodiversity-Ecosystem Functioning Relationships. Proc. Natl. Acad. Sci. USA 2003, 100, 14949–14954. [Google Scholar] [CrossRef] [PubMed]
- O’Gorman, E.J.; Jacob, U.; Jonsson, T.; Emmerson, M.C. Interaction Strength, Food Web Topology and the Relative Importance of Species in Food Webs. J. Anim. Ecol. 2010, 79, 682–692. [Google Scholar] [CrossRef]
- Trøjelsgaard, K.; Olesen, J.M. Ecological Networks in Motion: Micro- and Macroscopic Variability across Scales. Funct. Ecol. 2016, 30, 1926–1935. [Google Scholar] [CrossRef]
- Finke, D.L.; Denno, R.F. Spatial Refuge from Intraguild Predation: Implications for Prey Suppression and Trophic Cascades. Oecologia 2006, 149, 265–275. [Google Scholar] [CrossRef]
- Wu, D.; Xu, C.; Wang, S.; Zhang, L.; Kortsch, S. Why Are Biodiversity—Ecosystem Functioning Relationships so Elusive? Trophic Interactions May Amplify Ecosystem Function Variability. J. Anim. Ecol. 2023, 92, 367–376. [Google Scholar] [CrossRef]
- Srivastava, D.S.; Cardinale, B.J.; Downing, A.L.; Duffy, J.E.; Jouseau, C.; Sankaran, M.; Wright, J.P. Diversity Has Stronger Top-down than Bottom-up Effects on Decomposition. Ecology 2009, 90, 1073–1083. [Google Scholar] [CrossRef]
- Shurin, J.B.; Borer, E.T.; Seabloom, E.W.; Anderson, K.; Blanchette, C.A.; Broitman, B.; Cooper, S.D.; Halpern, B.S. A Cross-Ecosystem Comparison of the Strength of Trophic Cascades. Ecol. Lett. 2002, 5, 785–791. [Google Scholar] [CrossRef]
- Ives, A.R.; Cardinale, B.J.; Snyder, W.E. A Synthesis of Subdisciplines: Predator-Prey Interactions, and Biodiversity and Ecosystem Functioning. Ecol. Lett. 2005, 8, 102–116. [Google Scholar] [CrossRef]
- Cardinale, B.; Duffy, E.; Srivastava, D.; Loreau, M.; Thomas, M.; Emmerson, M. Towards a Food Web Perspective on Biodiversity and Ecosystem Functioning. In Biodiversity, Ecosystem Functioning, and Human Wellbeing; Naeem, S., Bunker, D.E., Hector, A., Loreau, M., Perrings, C., Eds.; Oxford University Press: Oxford, UK, 2009; pp. 105–120. ISBN 9780199547951. [Google Scholar]
- Griffin, J.N.; O’Gorman, E.J.; Emmerson, M.C.; Jenkins, S.R.; Klein, A.-M.; Loreau, M.; Symstad, A. Biodiversity and the Stability of Ecosystem Functioning. In Biodiversity, Ecosystem Functioning, and Human Wellbeing; Naeem, S., Daniel, E., Bunker, D.E., Hector, A., Loreau, M., Perrings, C., Eds.; Oxford University Press: Oxford, UK, 2009; pp. 78–93. ISBN 9780191720345. [Google Scholar]
- Pimm, S.L. The Complexity and Stability of Ecosystems. Nature 1984, 307, 321–326. [Google Scholar] [CrossRef]
- May, R.M. Will a Large Complex System Be Stable? Nature 1972, 238, 413–414. [Google Scholar] [CrossRef] [PubMed]
- Saint-Béat, B.; Baird, D.; Asmus, H.; Asmus, R.; Bacher, C.; Pacella, S.R.; Johnson, G.A.; David, V.; Vézina, A.F.; Niquil, N. Trophic Networks: How Do Theories Link Ecosystem Structure and Functioning to Stability Properties? A Review. Ecol. Indic. 2015, 52, 458–471. [Google Scholar] [CrossRef]
- Ives, A.R.; Klug, J.L.; Gross, K. Stability and Species Richness in Complex Communities. Ecol. Lett. 2000, 3, 399–411. [Google Scholar] [CrossRef]
- Donohue, I.; Petchey, O.L.; Montoya, J.M.; Jackson, A.L.; McNally, L.; Viana, M.; Healy, K.; Lurgi, M.; O’Connor, N.E.; Emmerson, M.C. On the Dimensionality of Ecological Stability. Ecol. Lett. 2013, 16, 421–429. [Google Scholar] [CrossRef]
- Holling, C.S. Resilience and Stability of Ecological Systems. Annu. Rev. Ecol. Syst. 1973, 4, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Loreau, M.; Mouquet, N.; Gonzalez, A. Biodiversity as Spatial Insurance in Heterogeneous Landscapes. Proc. Natl. Acad. Sci. USA 2003, 100, 12765–12770. [Google Scholar] [CrossRef]
- Ives, A.R.; Carpenter, S.R. Stability and Diversity of Ecosystems. Science 2007, 317, 58–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elton, C.S. The Ecology of Invasions by Animals and Plants; Springer US: Boston, MA, USA, 1958; ISBN 978-0-412-11430-4. [Google Scholar]
- Paine, R.T. Food Web Complexity and Species Diversity. Am. Nat. 1966, 100, 65–75. [Google Scholar] [CrossRef]
- MacArthur, R. Fluctuations of Animal Populations and a Measure of Community Stability. Ecology 1955, 36, 533. [Google Scholar] [CrossRef]
- Peterson, G.; Allen, C.R.; Holling, C.S. Ecological Resilience, Biodiversity, and Scale. Ecosystems 1998, 1, 6–18. [Google Scholar] [CrossRef]
- de Mazancourt, C.; Isbell, F.; Larocque, A.; Berendse, F.; De Luca, E.; Grace, J.B.; Haegeman, B.; Wayne Polley, H.; Roscher, C.; Schmid, B.; et al. Predicting Ecosystem Stability from Community Composition and Biodiversity. Ecol. Lett. 2013, 16, 617–625. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Pu, Z. Different Effects of Species Diversity on Temporal Stability in Single-Trophic and Multitrophic Communities. Am. Nat. 2009, 174, 651–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinha, S. Complexity vs. Stability in Small-World Networks. Phys. A Stat. Mech. Its Appl. 2005, 346, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Thébault, E.; Loreau, M. Trophic Interactions and the Relationship between Species Diversity and Ecosystem Stability. Am. Nat. 2005, 166, E95–E114. [Google Scholar] [CrossRef] [Green Version]
- Leary, D.J.; Petchey, O.L. Testing a Biological Mechanism of the Insurance Hypothesis in Experimental Aquatic Communities. J. Anim. Ecol. 2009, 78, 1143–1151. [Google Scholar] [CrossRef]
- Hautier, Y.; Tilman, D.; Isbell, F.; Seabloom, E.W.; Borer, E.T.; Reich, P.B. Anthropogenic Environmental Changes Affect Ecosystem Stability via Biodiversity. Science 2015, 348, 336–340. [Google Scholar] [CrossRef] [Green Version]
- Ives, A.R.; Gross, K.; Klug, J.L. Stability and Variability in Competitive Communities. Science 1999, 286, 542–544. [Google Scholar] [CrossRef] [Green Version]
- Yachi, S.; Loreau, M. Biodiversity and Ecosystem Productivity in a Fluctuating Environment: The Insurance Hypothesis. Proc. Natl. Acad. Sci. USA 1999, 96, 1463–1468. [Google Scholar] [CrossRef]
- Thibaut, L.M.; Connolly, S.R. Understanding Diversity-Stability Relationships: Towards a Unified Model of Portfolio Effects. Ecol. Lett. 2013, 16, 140–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, A.J.; Barry, K.E.; Lortie, C.J.; Callaway, R.M. Biodiversity and Ecosystem Functioning: Have Our Experiments and Indices Been Underestimating the Role of Facilitation? J. Ecol. 2021, 109, 1962–1968. [Google Scholar] [CrossRef]
- Barry, K.E.; Mommer, L.; van Ruijven, J.; Wirth, C.; Wright, A.J.; Bai, Y.; Connolly, J.; De Deyn, G.B.; de Kroon, H.; Isbell, F.; et al. The Future of Complementarity: Disentangling Causes from Consequences. Trends Ecol. Evol. 2019, 34, 167–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindo, Z. Linking Functional Traits and Network Structure to Concepts of Stability. Community Ecol. 2015, 16, 48–54. [Google Scholar] [CrossRef] [Green Version]
- Ma, F.; Zhang, F.; Quan, Q.; Song, B.; Wang, J.; Zhou, Q.; Niu, S. Common Species Stability and Species Asynchrony Rather than Richness Determine Ecosystem Stability Under Nitrogen Enrichment. Ecosystems 2021, 24, 686–698. [Google Scholar] [CrossRef]
- Jonsson, M.; Wardle, D.A. Structural Equation Modelling Reveals Plant-Community Drivers of Carbon Storage in Boreal Forest Ecosystems. Biol. Lett. 2010, 6, 116–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reiss, J.; Bridle, J.R.; Montoya, J.M.; Woodward, G. Emerging Horizons in Biodiversity and Ecosystem Functioning Research. Trends Ecol. Evol. 2009, 24, 505–514. [Google Scholar] [CrossRef]
- Maestre, F.T.; Quero, J.L.; Gotelli, N.J.; Escudero, A.; Ochoa, V.; Delgado-Baquerizo, M.; García-Gómez, M.; Bowker, M.A.; Soliveres, S.; Escolar, C.; et al. Plant Species Richness and Ecosystem Multifunctionality in Global Drylands. Science 2012, 335, 214–218. [Google Scholar] [CrossRef] [Green Version]
- Mouillot, D.; Villéger, S.; Scherer-Lorenzen, M.; Mason, N.W.H. Functional Structure of Biological Communities Predicts Ecosystem Multifunctionality. PLoS ONE 2011, 6, e17476. [Google Scholar] [CrossRef] [Green Version]
- Soliveres, S.; Van Der Plas, F.; Manning, P.; Prati, D.; Gossner, M.M.; Renner, S.C.; Alt, F.; Arndt, H.; Baumgartner, V.; Binkenstein, J.; et al. Biodiversity at Multiple Trophic Levels Is Needed for Ecosystem Multifunctionality. Nature 2016, 536, 456–459. [Google Scholar] [CrossRef]
- Thompson, P.L.; Gonzalez, A. Ecosystem Multifunctionality in Metacommunities. Ecology 2016, 97, 2867–2879. [Google Scholar] [CrossRef] [PubMed]
- Loreau, M. Does Functional Redundancy Exist? Oikos 2004, 104, 606–611. [Google Scholar] [CrossRef] [Green Version]
- Díaz, S.; Lavorel, S.; De Bello, F.; Quétier, F.; Grigulis, K.; Robson, T.M. Incorporating Plant Functional Diversity Effects in Ecosystem Service Assessments. Proc. Natl. Acad. Sci. USA 2007, 104, 20684–20689. [Google Scholar] [CrossRef] [PubMed]
- Gamfeldt, L.; Roger, F. Revisiting the Biodiversity–Ecosystem Multifunctionality Relationship. Nat. Ecol. Evol. 2017, 1, 0168. [Google Scholar] [CrossRef] [PubMed]
- Kollmann, J.; Meyer, S.T.; Bateman, R.; Conradi, T.; Gossner, M.M.; de Souza Mendonça, M.; Fernandes, G.W.; Hermann, J.M.; Koch, C.; Müller, S.C.; et al. Integrating Ecosystem Functions into Restoration Ecology—Recent Advances and Future Directions. Restor. Ecol. 2016, 24, 722–730. [Google Scholar] [CrossRef]
- Steffen, W.; Richardson, K.; Rockstrom, J.; Cornell, S.E.; Fetzer, I.; Bennett, E.M.; Biggs, R.; Carpenter, S.R.; de Vries, W.; de Wit, C.A.; et al. Planetary Boundaries: Guiding Human Development on a Changing Planet. Science 2015, 347, 1259855. [Google Scholar] [CrossRef] [Green Version]
- Hogan, A.; Blomqvist, E.; Cochez, M.; D’amato, C.; De Melo, G.; Gutierrez, C.; Kirrane, S.; Gayo, J.E.L.; Navigli, R.; Neumaier, S.; et al. Knowledge Graphs. ACM Comput. Surv. 2022, 54, 1–37. [Google Scholar] [CrossRef]
Figure 1.
Common patterns in the relationship between biodiversity and ecosystem functioning (BEF). Biodiversity interacts with climate, resource availability, disturbance regimes, and ecological processes at different scales to configure ecosystem functioning. Biodiversity effects on ecosystem functioning depend on the functional traits of component species, interspecific interactions, the sequence of species changes, and the relative biotic versus abiotic control over process rates.
Figure 1.
Common patterns in the relationship between biodiversity and ecosystem functioning (BEF). Biodiversity interacts with climate, resource availability, disturbance regimes, and ecological processes at different scales to configure ecosystem functioning. Biodiversity effects on ecosystem functioning depend on the functional traits of component species, interspecific interactions, the sequence of species changes, and the relative biotic versus abiotic control over process rates.

Figure 2.
Ecosystem processes linking biodiversity and ecosystem functioning. Ecosystem processes (such as productivity, nutrient and biogeochemical cycling, ecological networks dynamics and stability) link biodiversity to ecosystem functioning in multiple environmental contexts and at distinctive scales. Diverse communities lead to aggregate ecosystem functioning that favors complexity and adaptation, resulting in complex adaptive ecosystems.
Figure 2.
Ecosystem processes linking biodiversity and ecosystem functioning. Ecosystem processes (such as productivity, nutrient and biogeochemical cycling, ecological networks dynamics and stability) link biodiversity to ecosystem functioning in multiple environmental contexts and at distinctive scales. Diverse communities lead to aggregate ecosystem functioning that favors complexity and adaptation, resulting in complex adaptive ecosystems.

Figure 3.
Interplay between biodiversity and ecosystem multifunctionality. Increased biodiversity supports high ecosystem multifunctionality, through niche differentiation, redundancy, and biological insurance, which foster the functioning of the overall ecosystem at different spatial–temporal and ecological–evolutionary scales, sustaining complex adaptive ecosystems. Dispersal affects species distributions in face of environmental shifts and links metacommunities through habitat connectivity.
Figure 3.
Interplay between biodiversity and ecosystem multifunctionality. Increased biodiversity supports high ecosystem multifunctionality, through niche differentiation, redundancy, and biological insurance, which foster the functioning of the overall ecosystem at different spatial–temporal and ecological–evolutionary scales, sustaining complex adaptive ecosystems. Dispersal affects species distributions in face of environmental shifts and links metacommunities through habitat connectivity.

Figure 4.
Framework for ecological management based on a BEF approach adopting CAS thinking. Ecological management should include the identification and analysis of local interactions and processes to detect ecosystem emergent patterns across spatiotemporal scales. Critical transitions and regime shifts between alternative stable states reveal uncertainty. Biological interactions convey variability, complexity, and adaptation to sustain biodiversity. Behavioral interactions enhance adaptative capacity, resulting in collective behavior. Ecosystem processes ensure the flow of matter and energy, producing common pool resources. Resistance and resilience to unfavorable conditions confer robustness. Similar functional species buffer species losses, contributing to redundancy. Species dispersal and different habitats enable species to track favorable conditions, conveying environmental heterogeneity and connectivity among scales. Accounting for fast ecological and slow evolutionary processes is important. The role of local interactions and aggregate properties must be assessed. Management strategies need to be flexible and ecosystem-based, and to integrate socio-ecological perspectives that enhance the capacity of ecosystems to resist, recover, and adapt.
Figure 4.
Framework for ecological management based on a BEF approach adopting CAS thinking. Ecological management should include the identification and analysis of local interactions and processes to detect ecosystem emergent patterns across spatiotemporal scales. Critical transitions and regime shifts between alternative stable states reveal uncertainty. Biological interactions convey variability, complexity, and adaptation to sustain biodiversity. Behavioral interactions enhance adaptative capacity, resulting in collective behavior. Ecosystem processes ensure the flow of matter and energy, producing common pool resources. Resistance and resilience to unfavorable conditions confer robustness. Similar functional species buffer species losses, contributing to redundancy. Species dispersal and different habitats enable species to track favorable conditions, conveying environmental heterogeneity and connectivity among scales. Accounting for fast ecological and slow evolutionary processes is important. The role of local interactions and aggregate properties must be assessed. Management strategies need to be flexible and ecosystem-based, and to integrate socio-ecological perspectives that enhance the capacity of ecosystems to resist, recover, and adapt.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Correia, A.M.; Lopes, L.F. Revisiting Biodiversity and Ecosystem Functioning through the Lens of Complex Adaptive Systems. Diversity 2023, 15, 895. https://doi.org/10.3390/d15080895
AMA Style
Correia AM, Lopes LF. Revisiting Biodiversity and Ecosystem Functioning through the Lens of Complex Adaptive Systems. Diversity. 2023; 15(8):895. https://doi.org/10.3390/d15080895
Chicago/Turabian StyleCorreia, Alexandra M., and Luís F. Lopes. 2023. "Revisiting Biodiversity and Ecosystem Functioning through the Lens of Complex Adaptive Systems" Diversity 15, no. 8: 895. https://doi.org/10.3390/d15080895
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.