
Notes on the Summer Life History Traits of the Non-Native Pumpkinseed (Lepomis gibbosus) (Linnaeus, 1758) in a High-Altitude Artificial Lake

 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area Sampling
2.2. Data Collection
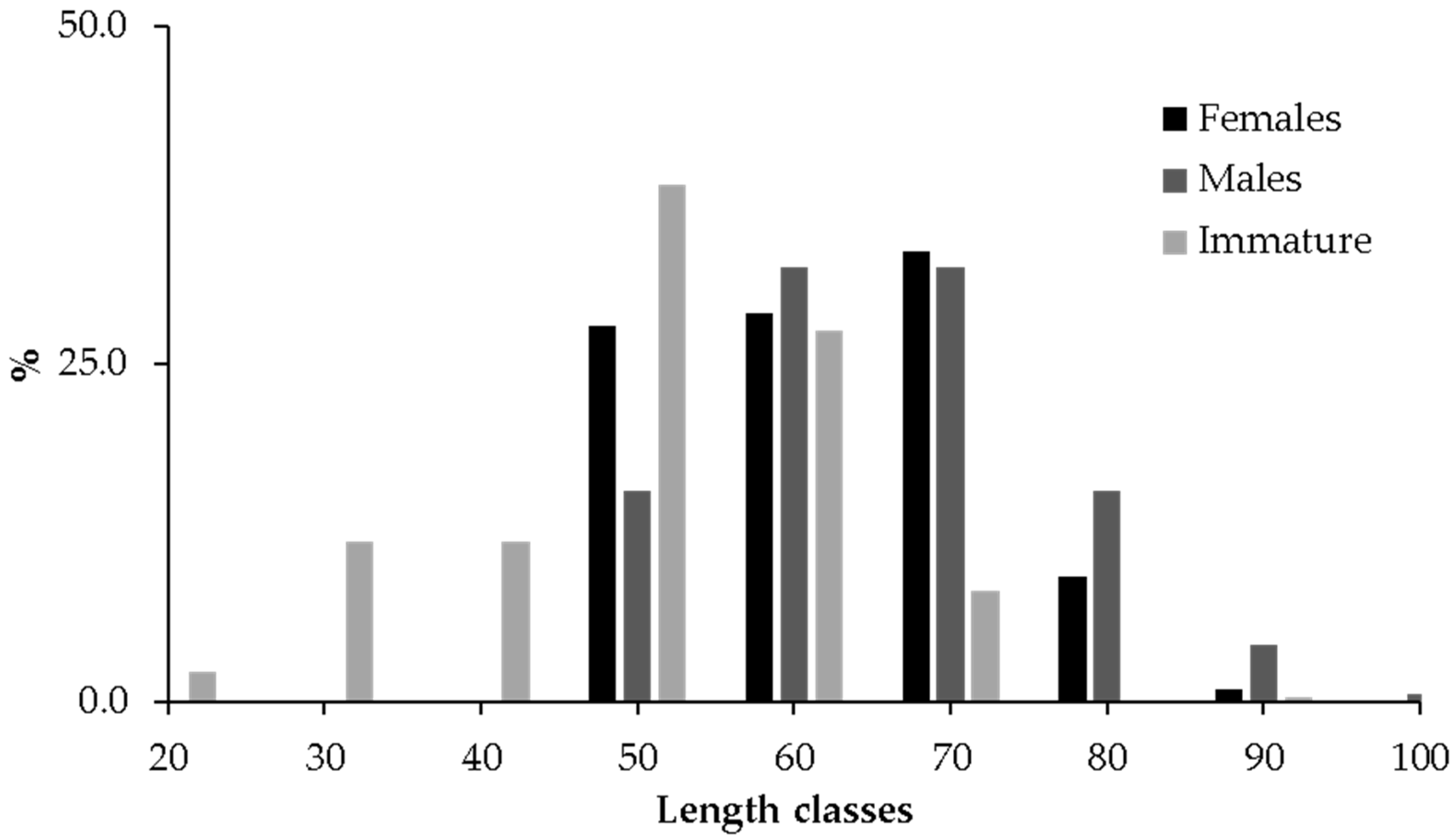
3. Results
3.1. Estimation of Life History Traits
3.2. Metanalytic Approach of Life History Traits
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Country | Length Type | Length Range | a | b | R2 | Year | Reference |
|---|---|---|---|---|---|---|---|---|
| Aoos Springs | Greece | TL | 24–101 | 0.012 | 3.190 | 0.985 | 2021 | Present study |
| River Vit | Bulgaria | SL | 33–92 | 0.024 | 3.187 | 0.963 | 2008 | [65] |
| River Segura | Spain | TL | 48–88 | 0.015 | 2.980 | 0.994 | 2000–2004 | [37] |
| Lake Bara | Croatia | TL | 0.013 | 3.140 | 0.991 | 2015 | [66] | |
| Catchment Basin | Turkey | TL | 27–107 | 0.008 | 3.491 | 0.950 | 2014 | [38] |
| Catchment Basin | Turkey | TL | 39–132 | 0.013 | 3.138 | 0.987 | 2014 | [38] |
| Region | Country | L∞ | k | t0 | tmax | φ′ | r2 | Year |
|---|---|---|---|---|---|---|---|---|
| Europe | ||||||||
| Artificial Lake of Aoos (Present study) | Greece | 119 | 0.36 | −0.31 | 8 | 3.707 | 0.691 | 2021 |
| Tapada Pequena Reservoir | Portugal | 81.3 | 0.41 | −0.151 | 7 | 3.43 | 0.995 | 1996 |
| Tapada Grande Reservoir | Portugal | 112.5 | 0.16 | −1.482 | 17 | 3.32 | 0.897 | 2004 |
| Monte Novo Reservoir | Portugal | 101.1 | 0.50 | −0.108 | 6 | 3.70 | 0.998 | 2004 |
| Divor Reservoir | Portugal | 91.1 | 0.43 | −0.550 | 6 | 3.56 | 0.989 | 1989 |
| Artificial Lake Kerkini | Greece | 163.5 | 0.13 | −2.729 | 20 | 3.53 | 1.000 | 1994 |
| Lake Banyoles | Spain | 131.8 | 0.57 | 0.367 | 6 | 3.99 | 0.998 | – |
| Fumemorte Canal | France | 113.6 | 0.34 | 0.013 | 9 | 3.65 | 0.997 | 2001 |
| Sollac Marsh | France | 133.5 | 0.36 | 0.029 | 8 | 3.80 | 0.997 | 2001 |
| Dabas Pond | Hungary | 94.9 | 0.38 | −0.059 | 8 | 3.54 | 0.996 | 1977 |
| Danube River | Slovakia | 168.3 | 0.23 | −0.158 | 13 | 3.82 | 1.000 | 1973 |
| Cottesmore Pond | England | 94.5 | 0.19 | −0.721 | 15 | 3.23 | 0.996 | 2002 |
| Odra River | Poland | 162.7 | 0.31 | −0.584 | 9 | 3.92 | 0.994 | – |
| North America | ||||||||
| Deep Lake | Michigan, USA | 190.7 | 0.17 | 0.048 | 18 | 3.78 | 0.990 | 1948 |
| Lower Dowsley Pond | Ontario, CAN | 174.2 | 0.24 | 0.195 | 13 | 3.87 | 1.000 | 1990 |
| Opinicon Lake | Ontario, CAN | 184.5 | 0.14 | −0.647 | 21 | 3.67 | 0.994 | 1990 |
| Upper Poole Pond | Ontario, CAN | 145.5 | 0.21 | −0.212 | 14 | 3.65 | 0.996 | 1990 |
| Houghton Lake | Michigan, USA | 256.1 | 0.20 | 0.473 | 15 | 4.11 | 0.990 | 1926 |
| Flora Lake | Wisconsin, USA | 205.5 | 0.16 | −0.327 | 18 | 3.84 | 0.993 | 1958 |
| Plastic Lake | Ontario, CAN | 180.0 | 0.16 | 0.124 | 19 | 3.74 | 0.970 | 1986 |
| Crosson Lake | Ontario, CAN | 168.0 | 0.17 | 0.152 | 18 | 3.68 | 0.986 | 1986 |
| Lake Upper Beverly | Ontario, CAN | 219.0 | 0.13 | −0.026 | 23 | 3.78 | 0.992 | 1987 |
| Lake Lower Beverly | Ontario, CAN | 125.3 | 0.26 | −0.147 | 11 | 3.62 | 0.991 | 1987 |
| Lake Vert | Quebec, CAN | 153.0 | 0.16 | −0.054 | 19 | 3.58 | 0.982 | 1979 |
| Lake Lower Loch Alpine | Michigan, USA | 142.8 | 0.18 | −0.612 | 16 | 3.56 | 0.989 | 1977 |
| A number of 55 lakes | Wisconsin, USA | 195.7 | 0.16 | −0.588 | 18 | 3.79 | 0.995 | 1992 |
| A number of 14 water bodies | Pennsylvania, USA | 160.1 | 0.23 | 0.393 | 13 | 3.78 | 0.982 | 1977 |
| Unnamed Pond | Michigan, USA | 130.4 | 0.24 | −0.802 | 12 | 3.62 | 0.997 | 1938 |
| Unspecified Lake | North Carolina, USA | 280.5 | 0.15 | 0.472 | 20 | 4.07 | 0.981 | 1997 |
| Unspecified Lake | New York, USA | 175.5 | 0.24 | 0.386 | 13 | 3.86 | 0.995 | 1977 |
| A number of 26 ponds | Rhode Island, USA | 172.6 | 0.27 | 0.247 | 11 | 3.91 | 0.999 | 1977 |
| Multiple water bodies | Delaware, USA | 154.0 | 0.63 | 0.287 | 5 | 4.17 | 0.997 | 1997 |
| Multiple streams | North Carolina, USA | 237.0 | 0.17 | 0.312 | 18 | 3.98 | 0.978 | 1977 |
| Multiple water bodies | Illinois, USA | 175.8 | 0.20 | −1.639 | 13 | 3.79 | 0.988 | 1977 |
| Multiple water bodies | Ohio, USA | 189.0 | 0.24 | 0.289 | 13 | 3.93 | 0.999 | 1977 |
| Multiple water bodies | Minnesota, USA | 213.4 | 0.19 | 0.014 | 16 | 3.94 | 0.999 | 1977 |
| Multiple water bodies | Minnesota, USA | 292.5 | 0.17 | 0.211 | 18 | 4.16 | 0.981 | 1977 |
References
- Magliozzi, C.; Tsiamis, K.; Vigiak, O.; Deriu, I.; Gervasini, E.; Cardoso, A.C. Assessing invasive alien species in European catchments: Distribution and impacts. Sci. Total Environ. 2020, 732, 138677. [Google Scholar] [CrossRef]
- Kleitou, P.; Moutopoulos, D.K.; Giovos, I.; Kletou, D.; Savva, I.; Cai, L.L.; Hall-Spencer, J.M.; Charitou, A.; Elia, M.; Katselis, G.; et al. Conflicting interests and growing importance of non-indigenous species in commercial and recreational fisheries of the Mediterranean Sea. Fish. Manag. Ecol. 2022, 2, 169–182. [Google Scholar] [CrossRef]
- Reyjol, Y.; Hugueny, B.; Pont, D.; Bianco, P.G.; Beier, U.; Caiola, N.; Casals, F.; Cowx, I.; Economou, A.; Ferreira, T.; et al. Patterns in species richness and endemism of European freshwater fish. Glob. Ecol. Biogeogr. 2007, 16, 65–75. [Google Scholar] [CrossRef]
- Barbieri, R.; Zogaris, S.; Kalogianni, E.; Stoumboudi, M.Th.; Chatzinikolaou, Y.; Giakoumi, S.; Kapakos, Y.; Kommatas, D.; Koutsikos, N.; Tachos, V.; et al. Freshwater Fishes and Lampreys of Greece: An Annotated Checklist; Monographs on Marine Sciences No. 8; Hellenic Centre for Marine Research: Athens, Greece, 2015; 128p. [Google Scholar]
- Economidis, P.S.; Dimitriou, E.; Pagoni, R.; Michaloudi, E.; Natsis, L. Introduced and translocated fish species in the inland waters of Greece. Fish. Manag. Ecol. 2000, 7, 239–250. [Google Scholar] [CrossRef]
- Hermoso, V.; Clavero, M.; Blanco-Garrido, F.; Prenda, J. Invasive species and habitat degradation in Iberian streams: An analysis of their role in freshwater fish diversity loss. Ecol. Appl. 2011, 21, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Kalogianni, E.; Koutsikos, N.; Vardakas, L.; Giakoumi, S.; Chatzinikolaou, Y.; Oikonomou, A. Impacts of the alien mosquitofish on the abundance and condition of two Mediterranean native fish. Mediterr. Mar. Sci. 2019, 20, 727. [Google Scholar] [CrossRef] [Green Version]
- IUCN. The IUCN Red List of Threatened Species. Version 2022-2 . 2023. Available online: https://www.iucnredlist.org (accessed on 29 July 2023).
- Bobori, D.; Petriki, O.; Aftzi, C. Fish Community Structure in the Mediterranean Temperate Lake Volvi at Two Different Stages of umpkinseed Invasion: Are Natives in Threat? Turk. J. Fish. Aquat. Sci. 2017, 19, 1039–1048. [Google Scholar]
- Arnold, A. Eingebürgerte Fischarten. Neue Brehm-Bücherei Band 1990, 602, 144. [Google Scholar]
- Tandon, K.K. Notes on systematics of the Pumpkin Seed, Lepomis gibbosus (Ostheichthyes, Perciformes, Centrarchidae). Vĕstn. Ceskoslov. Společnosti Zool. 1976, 40, 307–311. [Google Scholar]
- Giapis, A.Ι. Ecological study of Lepomis gibossus (L.) in dam lake Kerkini. Ph.D. Thesis, School of Forestry and Natural Environment, Aristotle University of Thessaloniki, Thessaloniki, Greece, 2003; 240p. [Google Scholar]
- Holčik, J. Fish introductions in Europe with particular reference to its Central and Eastern part. Can. J. Fish. Aquat. Sci. 1991, 48 (Suppl. 1), 13–23. [Google Scholar] [CrossRef]
- Economidis, P.S.; Kattoulas, M.; Stephanidis, E. Fish fauna of the Aliakmon Rriver and the adjacent waters (Macedonia, Greece). Cybium 1981, 5, 89–95. [Google Scholar]
- Moutopoulos, D.K.; Korakis, A.; Katselis, G. Changes of the ichthyofauna in the impoundment of the Aoos Springs, Greece. Acta Zool. Bulg. 2023, 75, 225–233. [Google Scholar]
- Riehl, R.; Baensch, H.A. Aquarien Atlas. Band. 1; Mergus, Verlag für Natur-und Heimtierkunde: Melle, Germany, 1991; 992p. [Google Scholar]
- Blackburn, T.M.; Pyšek, P.; Bacher, S.; Carlton, J.T.; Duncan, R.P.; Jarošík, V.; Wilson, J.R.U.; Richardson, D.M. A proposed unified framework for biological invasions. Trends Ecol. Evol. 2011, 26, 333–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zogaris, S. Information on Measures and Related Costs in Relation to Species Considered for Inclusion on the Union List: Lepomis spp.; Technical Note Prepared by IUCN for the European Commission; European Commission: Brussels, Belgium, 2017. [Google Scholar]
- Przybylski, M.; Zięba, G. NOBANIS—Invasive Alien Species Fact Sheet—Lepomis gibbosus. Available online: https://www.nobanis.org/ (accessed on 29 July 2023).
- Balon, E.K. Nerest Lepomis gibbosus (Linné, 1758), aklimatizovanej v bočných vodach Dunaja a jej vývoj počas embryonalnej periody. Vĕstn. Ceskoslov. Společnosti Zool. 1957, 23, 1–22. [Google Scholar]
- Godinho, F.N.; Ferreira, M.T.; Cortes, R.V. The environmental basis of diet variation in pumpkinseed sunfish, Lepomis gibbosus, and largemouth bass, Micropterus salmoides, along an Iberian river basin. Environ. Biol. Fishes 1997, 50, 105–115. [Google Scholar] [CrossRef]
- Copp, G.H.; Fox, M.G.; Przybylski, M.; Godhinho, F.N.; Vila-Gispert, A. Life-time growth pattern of pumpkinseed Lepomis gibbosus introduced to Europe, relative to native North American populations. Folia Zool. Brno 2004, 56, 237–254. [Google Scholar]
- Vardakas, L.; Koutsikos, N.; Perdikaris, C.; Petriki, O.; Bobori, D.; Zogaris, S.; Giakoumi, S.; Fitoka, E.; Tompoulidou, M.; Tsiaoussi, V.; et al. The fish fauna in lentic ecosystems of Greece. Mediterr. Mar. Sci. 2021, 23, 223–265. [Google Scholar] [CrossRef]
- Ziou, A.; Douligeri, A.S.; Korakis, A.; Kyriazis, N.; Petsis, N.; Moutopoulos, D.K.; Katselis, G. Performance of a pilot application of electrofishing for the management of the invasive species Lepomis gibbosus (Linaeus, 1758) in the Artificial Lake of Aoos springs. In Proceedings of the 18th Panhellenic Congress of Ichthyologists, Messolonghi, Greece, 3–6 November 2022; Volume 18, pp. 275–279. [Google Scholar]
- Zacharias, I.; Doulas, C.; Barbieri, R.; Kousouris, T.; Bertachas, E.; Stoupoudi, M.; Psaras, T.; Giakoumi, S.; Economou, A.N. Comparative study of the physicochemical and biological parameters in the Aoos and Pournari reservoirs. In Proceedings of the 6th Panhellenic Symposium on Oceanography and Fisheries, Chios, Greece, 23–26 May 2000; pp. 224–229. [Google Scholar]
- Economou, A.; Barbieri, R.; Daoulas, C.; Psarras, M.; Stoumboudi, H.; Bertahas, S.; Giakoumi, S.; Patsias, A. Threatened endemic freshwater fish species from western Greece and Peloponnesus. National Centre of Marine Researches, Institute of Inland Waters, Technical Report. Available online: https://www.fishbase.se/References/FBRefSummary.php?ID=56925&database=FB (accessed on 29 July 2023). (In Greek).
- Economou, A.N.; Giakoumi, S.; Vardakas, L.; Barbieri, R.; Stoumboudi, M.Τ.; Zogaris, S. The freshwater ichthyofauna of Greece—An Update based on a hydrographic basin survey. Mediterr. Mar. Sci. 2007, 8, 91–166. [Google Scholar] [CrossRef] [Green Version]
- Moutopoulos, D.K.; Koutsikos, N.; Vardakas, L.; Perdikaris, C. A History of Greek Inland Fishery Development during 1928-2019. Fish Manag. Ecol. 2022, 29, 681–692. [Google Scholar] [CrossRef]
- Moutopoulos, D.K.; Stergiou, K.I. Length-weight and length-length relationships of fish species from the Aegean Sea (Greece). J. Appl. Ichthyol. 2002, 18, 200–203. [Google Scholar] [CrossRef]
- Zar, J.H. Biostatistical Analysis, 5th ed.; Prentice-Hall/Pearson: Upper Saddle River, NJ, USA, 2010; Volume XIII, 944p. [Google Scholar]
- Jakobsen, T.; Fogarty, M.J.; Megrey, B.A.; Moksness, E. Fish Reproductive Biology: Implications for Assessment and Management; Blackwell Publishing: Oxford, UK, 2009. [Google Scholar]
- Pauly, D. Fish Population Dynamics in Tropical Waters: A Manual for Use with Programmable Calculators; ICLARM Studies Review; ICLARM: Manilla, Philippines, 1984; Volume 8, 325p. [Google Scholar]
- Gayanilo, F.C.; Parre, P.S.; Pauly, D. The FAO ICLARM Stock Assessment Tools (FISAT). User’s Guide; FAO Computerized Information Series Fisheries; FAO: Rome, Italy, 1996; 126p. [Google Scholar]
- Pauly, D. Gill size and temperature as governing factors in fish growth: A generalization of von Bertalanffy’s growth formula. Ber. Inst. Meereskd. Univ. Kiel. 1979, 63, xv+156. [Google Scholar]
- Pauly, D.; Munro, J.L. Once more on the comparisons of growth in fish and invertebrates. Fishbyte 1984, 2, 1–21. [Google Scholar]
- Von Bertalanffy, L. A Quantitative Theory of Organic Growth (Inquiries on Growth Laws. II). Hum. Biol. 1938, 10, 181–213. [Google Scholar]
- Andreu-Soler, A.; Oliva-Paterna, F.J.; Torralva, M. A review of length–weight relationships of fish from the Segura River basin (SE Iberian Peninsula). J. Appl. Ichthyol. 2006, 22, 295–296. [Google Scholar] [CrossRef]
- Çiçek, E.; Seçer, B.; Sungur, S.; Eagderi, S.; Bahçeci, H. Length-weight relationships and condition factors of eight exotic fish species from Turkey. Turkish J. Fish. Aquat. Sci. 2022, 6, 260–274. [Google Scholar] [CrossRef]
- Cucherousset, J.; Copp, G.H.; Fox, M.G.; Sterud, E.; van Kleef, H.H.; Verreycken, H.; Záhorská, E. Life-history traits and potential invasiveness of introduced pumpkinseed Lepomis gibbosus populations in northwestern Europe. Biol. Invasions 2009, 11, 2171–2180. [Google Scholar] [CrossRef] [Green Version]
- Villeneuve, F.; Copp, G.H.; Fox, M.G.; Stakėnas, S. Interpopulation variation in growth and life-history traits of the introduced sunfish, pumpkinseed Lepomis gibbosus, in southern England. J. Appl. Ichthyol. 2005, 21, 275–281. [Google Scholar] [CrossRef]
- Maitland, P.S.; Campbell, R.N. Freshwater Fishes; HarperCollins Publishers: London, UK, 1992; p. 369. [Google Scholar]
- Fox, M.G.; Crivelli, A.J. Body size and reproductive allocation in a multiple spawning centrarchid. Can. J. Fish. Aquat. Sci. 1998, 55, 737–748. [Google Scholar] [CrossRef]
- Zieba, G.; Fox, M.G.; Copp, G.H. The effect of elevated temperature on spawning of introduced pumpkinseed Lepomis gibbosus in Europe. J. Fish. Biol. 2010, 77, 1850–1855. [Google Scholar] [CrossRef]
- Burns, J.R. The reproductive cycle and its environmental control in the pumpkinseed, Lepomis gibbosus (Pisces: Centrarchidae). Copeia 1976, 1976, 449–455. [Google Scholar] [CrossRef]
- Wootton, R.J. Ecology of Teleost Fishes; Kluwer Academic Publishers: Amsterdam, The Netherlands, 1999; 386p. [Google Scholar]
- Wootton, R.J.; Evans, G.W.; Mills, L. Annual cycle in female threespined sticklebacks (Gasterosteus aculeatus L.) from an upland and lowland population. J. Fish. Biol. 1978, 12, 183–196. [Google Scholar] [CrossRef]
- Pauly, D. On the interrelationships between natural mortality, growth parameters, and mean environmental temperature in 175 fish stocks. ICES J. Mar. Sci. 1980, 39, 175–192. [Google Scholar] [CrossRef]
- Moutopoulos, D.K.; Stoumboudi, M.Th.; Ramfos, A.; Tsagarakis, K.; Gritzalis, K.C.; Petriki, O.; Patsia, A.; Barbieri, R.; Machias, A.; Stergiou, K.I.; et al. Food web modelling on structure and functioning of a Mediterranean lentic system. Hydrobiologia 2018, 822, 259–283. [Google Scholar] [CrossRef]
- Arranz, I.; Brucet, S.; Bartrons, M.; García-Comas, C.; Benejam, L. Fish size spectra are affected by nutrient concentration and relative abundance of non-native species across streams of the NE Iberian Peninsula. Sci. Total Environ. 2021, 795, 148792. [Google Scholar] [CrossRef] [PubMed]
- Stearns, S.C. The Evolution of Life Histories; Oxford University Press: London, UK, 1992; Volume XII, 249p. [Google Scholar]
- Τomecek, J.V.; Kovac, K.; Stanislav, K. The biological flexibility of the pumpkinseed: A successful colonizer throughout Europe. Chapter 16 in Francesca Gherardi. In Biological Invaders in Inland Waters: Profiles, Distribution, and Threats; Gherardi, F., Ed.; Springer: Dordrecht, The Netherlands, Chapter 16; 2007; pp. 307–336. [Google Scholar]
- Miller, H.C. The Behavior of the Pumpkinseed Sunfish, Lepomis Gibbosus (Lin-neaus), with Notes on the Behavior of Other Species of Lepomis and the Pigmy Sunfish, Elassoma evergladei. Behaviour 1963, 22, 88–150. [Google Scholar] [CrossRef]
- Bhagat, Y.; Fox, M.G.; Ferreira, M.T. Trophic polymorphism in introduced pumpkinseed (Lepomis gibbosus) inhabiting Iberian reservoirs. Environ. Biol. Fish. 2011, 91, 203–217. [Google Scholar] [CrossRef]
- Gkenas, C.; Magalhães, M.F.; Cucherousset, J.; Domingos, I.; Ribeiro, F. Long term patterns in the late summer dietary niche of the invasive pumpkinseed sunfish Lepomis gibbosus. Knowl. Manag. Aquat. Ecosyst. 2016, 417, 19. [Google Scholar] [CrossRef] [Green Version]
- Van Kleef, H.; van der Velde, G.; Leuven, R.; Esselink, H. Pumpkinseed sunfish (Lepomis gibbosus) invasions facilitated by introductions and nature management strongly reduce macroinvertebrate abundance in isolated water bodies. Biol. Invasions 2008, 10, 1481–1490. [Google Scholar] [CrossRef] [Green Version]
- Almeida, D.; Merino-Aguirre, R.; Vilizzi, L.; Copp, G.H. Interspecific aggressive behaviour of invasive pumpkinseed Lepomis gibbosus in Iberian fresh waters. PLoS ONE 2014, 9, e88038. [Google Scholar] [CrossRef] [Green Version]
- Copp, G.H.; Britton, J.R.; Guo, Z.; Edmonds-Brown, V.R.; Pegg, J.; Vilizzi, L.; Davison, P.I. Trophic consequences of non-native pumpkinseed Lepomis gibbosus for native pond fishes. Biol. Invasions 2017, 19, 25–41. [Google Scholar] [CrossRef] [Green Version]
- Miner, J.G.; Stein, R.A. Detection of predators and habitat choice by small bluegills: Effects of turbidity and alternative prey. Trans. of the Americ. Fisheries Soc. 1996, 125, 97–103. [Google Scholar] [CrossRef]
- Holtan, P. Pumpkinseed; Wisconsin Department of Fisheries Management, Bureau of Fisheries Management: Madison, WI, USA, 1991. [Google Scholar]
- Bergmann, C. Über die verhältnisse der wärmeökonomie de thiere zu ihrer grösse. Götting Stud. 1847, 3, 595–708. [Google Scholar]
- Blanchet, S.; Grenouillet, G.; Beauchard, O.; Tedesco, P.A.; Leprieur, F.; Dürr, H.H.; Busson, F.; Oberdorff, T.; Brosse, S. Non-native species disrupt the worldwide patterns of freshwater fish body size: Implications for Bergmann’s rule. Ecol. Lett. 2010, 13, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, D. Pattern and process in the ecological biogeography of European freshwater fish. J. Anim. Ecol. 2006, 75, 734–751. [Google Scholar] [CrossRef] [PubMed]
- Piechocki, A.; Szlauer-Lukaszewska, A. Molluscs of the middle and lower Odra: The role of the river in the expansion of alien species in Poland. Folia Malacol. 2013, 21, 73–86. [Google Scholar] [CrossRef] [Green Version]
- Ohlberger, J. Climate warming and ectotherm body size—From individual physiology to community ecology. Funct. Ecol. 2013, 27, 991–1001. [Google Scholar] [CrossRef]
- Uzunova, E.; Velkov, B.; Studenkov, S.; Georgieva, M.; Nikolova, M.; Pehlivanov, L.; Parvanov, D. Growth, age and size structure of the introduced pumpkinseed (Lepomis gibbosus L.) population from small ponds along the Vit River (Bulgaria). Bulg. J. Agric. Sci. 2008, 14, 227–234. [Google Scholar]
- Khanom, M.; Rouf, M.A.; Ahsan, N.; Siddiqui, N.; Tomljanović, T. Morphological characteristics, growth and age structure of allochthonous fish pumpkinseed, Lepomis gibbosus in Bara Lake, Croatia. Bulg. J. Agric. Sci. 2020, 26, 213–222. [Google Scholar]





| Month/Season | Ν | TL | W = aTLb | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Min | Max | Mean | SD | a-Value | b-Value | R2 | |||
| Month | July | 54 | 42 | 90 | 63 | 11.9 | 0.009 | 3.306 | 0.990 |
| August | 310 | 44 | 101 | 64 | 13 | 0.011 | 3.212 | 0.983 | |
| September | 160 | 30 | 95 | 63 | 15 | 0.013 | 3.156 | 0.982 | |
| October | 57 | 24 | 71 | 44 | 14.8 | 0.012 | 3.174 | 0.994 | |
| Season | Summer | 364 | 42 | 101 | 64 | 13 | 0.011 | 3.233 | 0.984 |
| Autumn | 217 | 24 | 95 | 58 | 15 | 0.012 | 3.194 | 0.986 | |
| TOTAL | 581 | 24 | 101 | 62 | 13 | 0.012 | 3.192 | 0.985 | |
| Length Classes (mm) | 0 | 1 | 2 | 3 | 4 | 5 | 6 | Total | % |
|---|---|---|---|---|---|---|---|---|---|
| Age | |||||||||
| 20–25 | 1 | 1 | 0.2 | ||||||
| 25–30 | 5 | 5 | 0.9 | ||||||
| 30–35 | 17 | 17 | 2.9 | ||||||
| 35–40 | 16 | 16 | 2.8 | ||||||
| 40–45 | 16 | 16 | 2.8 | ||||||
| 45–50 | 17 | 17 | 2.9 | ||||||
| 50–55 | 37 | 15 | 52 | 9.0 | |||||
| 55–60 | 26 | 87 | 2 | 115 | 19.8 | ||||
| 60–65 | 9 | 81 | 11 | 101 | 17.4 | ||||
| 65–70 | 3 | 18 | 45 | 3 | 69 | 11.9 | |||
| 70–75 | 4 | 60 | 21 | 85 | 14.6 | ||||
| 75–80 | 19 | 13 | 4 | 36 | 6.2 | ||||
| 80–85 | 2 | 10 | 12 | 1 | 25 | 4.3 | |||
| 85–90 | 3 | 9 | 3 | 15 | 2.6 | ||||
| 90–95 | 3 | 3 | 6 | 1.0 | |||||
| 95–100 | 4 | 4 | 0.7 | ||||||
| 100–105 | 1 | 1 | 0.2 | ||||||
| Total | 147 | 205 | 139 | 50 | 28 | 11 | 1 | 581 | |
| % | 25.3 | 35.3 | 23.9 | 8.6 | 4.8 | 1.9 | 0.2 | ||
| Region | Country | Month | GSI | ΚR | Year | Reference |
|---|---|---|---|---|---|---|
| Artificial lake of Aoos | Greece | July | 4.67 | 1.08 | 2021 | Present study |
| 19 water bodies | England | June | 7.70 | 1.02 | 2003–04 | [40] |
| Lake Einedammen | Norway | July | 7.22 | 0.91 | 2006 | [39] |
| Lake Schoapedobbe | Netherlands | June | 10.04 | 1.00 | 2006 | [39] |
| Lake Meeuwven | Netherlands | June | 8.78 | 0.91 | 2006 | [39] |
| Lake Cottesmore School | England | June | 9.80 | 0.88 | 2006 | [39] |
| Stream Batts Bridge | England | June | 4.13 | 0.97 | 2006 | [39] |
| Isle of Wight | England | June | 6.71 | 1.07 | 2005 | [39] |
| Lake Webbekomsbroek | Belgium | June | 6.10 | 1.04 | 2006 | [39] |
| Stream Slangebeek | Belgium | June | 3.14 | 1.05 | 2006 | [39] |
| Marsh Brière | France | May | 11.20 | 1.04 | 2006 | [39] |
| Lake Grand-Lieu | France | May | 11.36 | 1.19 | 2006 | [39] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Douligeri, A.S.; Ziou, A.; Korakis, A.; Kiriazis, N.; Petsis, N.; Katselis, G.; Moutopoulos, D.K. Notes on the Summer Life History Traits of the Non-Native Pumpkinseed (Lepomis gibbosus) (Linnaeus, 1758) in a High-Altitude Artificial Lake. Diversity 2023, 15, 910. https://doi.org/10.3390/d15080910
Douligeri AS, Ziou A, Korakis A, Kiriazis N, Petsis N, Katselis G, Moutopoulos DK. Notes on the Summer Life History Traits of the Non-Native Pumpkinseed (Lepomis gibbosus) (Linnaeus, 1758) in a High-Altitude Artificial Lake. Diversity. 2023; 15(8):910. https://doi.org/10.3390/d15080910
Chicago/Turabian StyleDouligeri, Alexandra S., Athina Ziou, Athanasios Korakis, Nikolaos Kiriazis, Nikolaos Petsis, George Katselis, and Dimitrios K. Moutopoulos. 2023. "Notes on the Summer Life History Traits of the Non-Native Pumpkinseed (Lepomis gibbosus) (Linnaeus, 1758) in a High-Altitude Artificial Lake" Diversity 15, no. 8: 910. https://doi.org/10.3390/d15080910