


Active Coral Restoration to Preserve the Biodiversity of a Highly Impacted Reef in the Maldives
by

Irene Pancrazi
1,2,*,
Kayla Feairheller
3,
Hassan Ahmed
2,
Carolina di Napoli
1 and
Monica Montefalcone
1,4
1
Seascape Ecology Laboratory, DiSTAV, Department of Earth, Environment and Life Sciences, University of Genoa, Corso Europa 26, 16132 Genova, Italy
2
Save the Beach Maldives, Boakeyo Goalhi, Villimalé 21022, Maldives
3
Bleu World, Catalina Island, CA 90704, USA
4
National Biodiversity Future Center (NBFC), Piazza Marina, 61, 90133 Palermo, Italy
*
Author to whom correspondence should be addressed.
Diversity 2023, 15(9), 1022; https://doi.org/10.3390/d15091022
Submission received: 15 August 2023
/
Revised: 11 September 2023
/
Accepted: 16 September 2023
/
Published: 19 September 2023
(This article belongs to the Special Issue Marine Ecosystem Restoration: Challenges and Opportunities)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Maldivian coral reefs have been experiencing significant degradation due to a combination of global climate change and local anthropogenic pressures. To enforce the conservation of coral reefs worldwide, coral restoration is becoming a popular tool to restore ecosystems actively. In the Maldives, restoration interventions are performed only around touristic islands, where there are economic resources available to support these projects. Unfortunately, on local islands, coral restoration does not benefit from the same support and is rarely boosted. A challenging coral restoration experiment has been performed, for the first time, on a local island of the Maldives affected by intense human pressures that caused the degradation of its reefs. A total of 242 coral fragments were collected from impacted colonies and transferred to the coral nursery of the island. Survival and growth rates of the fragments were monitored for 12 months. After one year, a survival rate of 70% was recorded. Although this rate might appear lower when compared to other restoration experiences, it is very promising considering the origin of the fragments and the poor quality of the environment where they have been transplanted. Some potential threats to the success of this restoration have also been identified, i.e., water temperature anomaly, diseases, and parasites, the latter being the leading causes of coral mortality. The procedure presented here is less expensive compared to the typical relocation of entire coral colonies from donor-healthy reefs to degraded reefs, thus providing an opportunity and a viable option for local islands to restore their reefs and preserve local biodiversity.
1. Introduction
Coral reefs are considered one of the most biodiverse and productive ecosystems on Earth [1,2]. Referred to as the rainforests of the sea, they host 25% of the total marine biodiversity, even though they cover only 0.2% of the seafloor [3]. Coral reefs provide a wide variety of ecosystem services: they provide nursery and aggregation areas for marine life [4,5], protect coasts, support fishery and tourism [6,7,8], create local identity, and provide cultural connections [6,9]. Despite their incredible value, coral reefs are fragile ecosystems, and in the past decades, they have been declining worldwide due to a combination of global and local anthropogenic pressures [10,11]. Unfortunately, the synergistic effect of global and local impacts is eroding reef resilience [12], making natural recovery slower or even unreachable [13].
The United Nations has recently launched the UN Decade on Ecosystem Restoration, which aims to promote global cooperation for the restoration of degraded ecosystems. From 2021 until 2030, the United Nations are asked to implement projects that combat climate change, preserve and restore biodiversity, and safeguard food and water supplies [14]. Ecological restoration seeks to initiate or accelerate ecosystem recovery following damage [15]. This practice is not a substitute for environmental conservation but aims to assist in the recovery of degraded ecosystems.
Coral reefs are considered priority ecosystems for ecological restoration interventions, which can be broadly categorized into passive or active practices [16]. Both passive and active approaches to coral restoration are crucial components of a conservation strategy that seeks to optimize biodiversity conservation and ecosystem services provision [17]. Passive coral restoration focuses mainly on habitat maintenance, management, and conservation, allowing natural processes to mitigate impacts with minimal to no human interference [18]. Due to the high intensity and frequency of human pressures on coral reefs, passive conservation is no longer enough [16], and severely degraded reefs, especially those missing breeding populations of formerly abundant organisms, might not be able to recover in a foreseeable time frame [18,19]. Active restoration projects have thus been increasing all around the world [15], providing direct human intervention through coral gardening that can be applied either in situ or ex situ within coral nurseries [20]. The ex situ methods involve the sampling and the transfer of either entire colonies or coral fragments on artificial substrates in a nursery area with controlled environmental conditions. When the coral fragments have reached a suitable size, they can be relocated to the degraded reef and then monitored over time [21].
The Republic of Maldives is a nation of coral islands built by the low-lying accumulation of unconsolidated sediment formed from the skeletal remains of carbonate-producing reef organisms, mainly corals [22,23]. Healthy coral reefs are thus vital for the existence and economic sustainability of the Maldives because they improve tourism, provide coastal protection, and sustain the growing population with abundant food [24]. Destruction of coral reefs for coastal development, mass coral mortality due to climate change impacts, and effects of sea level rise might lead to the erosion and the sinking of the islands, thus causing the land where Maldivian people live to disappear.
Coral reefs of the Maldives have been largely affected by climate change and global warming [25]. Rising sea level is causing flooding on many islands [26]. Global warming is exacerbating the ENSO phenomenon across the equatorial Pacific Ocean [11], a natural periodic fluctuation in sea surface temperature (known as El Niño) and in air pressure of the overlying atmosphere (known as Southern Oscillation) [27]. Due to the warming in sea surface temperature (SST), intense ENSO episodes are causing more severe, long-lasting, and frequent coral bleaching events across the country, with consequent mass coral mortalities [28].
In the past 40 years, coral reefs around the Maldives have also experienced increased local anthropogenic pressures, such as coastal development and land reclamations [12]. The capital atoll of North Malé is almost entirely developed, and the surrounding area of the capital city of Malé (i.e., the Greater Malé Area) is the most populated of the Maldives [29]. This intense human development has caused a decline in the water quality [30], challenging even further the surrounding reefs.
Low water quality may increase the spread of coral diseases [31] by reducing coral resistance to microbial infections while increasing pathogen virulence, leaving coral reef ecosystems in a state of disrepair with little hope of recovery [32]. Coral diseases have been identified as one of the most important causes of coral loss, and different environmental factors have been correlated to numerous disease processes [33]. Their occurrence is likely to be driven not by a single factor but rather by a multifactorial process that facilitates the emergence of pathogens in vulnerable ecosystems [34]. Although coral diseases have been largely investigated in some areas of the Caribbean and the Great Barrier Reef, less is known from the Indo-Pacific area [31]. Stony corals also provide microhabitats for thousands of micro and macro-parasitic and commensal organisms, which use the tissue and the skeleton of coral colonies as substrate. Currently, there are more than 165,000 described species of coral-associated invertebrates along a spectrum of symbiosis from mutualism to commensalism and parasitism. Parasites may stress the coral and, to some extent, may cause coral mortality through feeding and boring activities; some corallivores may also be able to spread diseases [35,36].
Coral restoration efforts throughout the Maldives are surprisingly few [21]. While luxury resorts operate small-scale restoration activities for tourism purposes [37], there are no examples of active coral restoration programs at the national scale, making it challenging for local islands and NGOs to find funding for these interventions. Transplanting coral colonies from healthy donor reefs to degraded reefs is considered the best approach to benefit recruitment and accelerate recovery [13,19,21]. However, considering the cost of the equipment needed for the intervention, the cost of the coral relocation (including boat fee, fuel, and dive gear rentals), and the cost of monitoring activities, active restoration projects cannot be afforded by most of the local Maldivian communities because of the lack of funding and local resources. Alternative economically viable initiatives are thus recommended for restoring the degraded reefs of the Maldives.
This study describes the first experiment of small-scale, low-cost, active coral restoration of a degraded reef in the local island of Villimalé (North Malé atoll) subjected to high human pressure. The reefs around Villimalé are dominated by corals of the genus Pocillopora, which have been highly affected by diseases and parasitism in the last years, causing the death of most of the colonies. Instead of transplanting entire healthy coral colonies from a donor reef, fragments of corals were harvested directly from the few colonies still alive in the degraded reef of Villimalé and were transferred to the coral nursery already existing on the site. Their growth was enhanced in this controlled environment in order to produce adult colonies (i.e., sexually mature and reproductive) with the aim of restoring and preserving the local biodiversity in situ. During the 1-year monitoring of the survival and the growth rate of coral fragments, the occurrence of coral diseases and parasites has also been monitored.
2. Materials and Methods
2.1. Study Area and Field Activities
The Maldives is an archipelago of 26 natural atolls with approximately 1200 small, low-lying coral islands stretching from North to South over an area of 860 km2 in the Indian Ocean. Coastal and marine ecosystems dominate the environment in the Maldives, as more than 99% of the territory is covered by the ocean [23].
This study was conducted at Villimalé island (4°10′23.79″ N, 73°29′7.29″ E), in the North Malé atoll (Figure 1), between December 2021 and November 2022. Villimalé is the 5th administrative district of the capital city of Malé, and it is part of the Greater Malé Area along with Hulumalé island [38,39]. Villimalé is the last island of the Greater Malé Area with three natural beaches, and it is the preferred destination for the local community to spend weekends and holidays. With nearly half of the Maldivian population residing in this area [29], the surrounding waters of Villimalé have high concentrations of organic wastes and chemical pollutants [40]. Additionally, Villimalé is near the garbage island of Thilafushi, which has been used as a dumping area since the early 1970s. The waste on the island has been used to fill the lagoon and reclaim the land, causing further pollution of the surrounding waters [41]. A warning alert for coral bleaching was dispatched in this region for the period between the middle of April and the middle of May 2022 due to increased sea surface temperatures because of the severe heat wave consequent of an ENSO episode [42].
For the restoration intervention, six metal frames were deployed on the rocky bottom of the reef between 1.5–2.1 m depth (depending on the tide) on the northwest corner of Villimalé island, where a coral nursery already exists (Figure 1). The metal structures were designed to match the natural height (about 30 cm) of coral rocks on the reef flat and were double-coated with resins and sand to avoid the spread of rust and create a suitable substrate for the corals (Figure 2).
A total of 242 healthy fragments of Pocillopora meandrina (n = 164) and Pocillopora verrucosa (n = 78) were collected from the few living corals remaining on the impacted Villimalé reef by harvesting only healthy branches. The branches were fragmented into 5–6 cm fragments and were transferred to the coral nursery (which is located in the same area of the impacted reef, but it is under controlled conditions) and then attached directly to the frames using cable ties (Figure 2). The first monitoring was carried out two weeks after the transfer of the coral fragments to evaluate their response to mechanical stress, and then the survival and growth rates were monitored monthly for 12 months (from December 2021 to November 2022). To ensure the success of the restoration experiment, scientific divers surveyed the coral nursery three times per week to perform maintenance on the frames. For consistency with the conditions in the donor reef, neither diseased corals nor parasites were removed, and the maintenance focused on cleaning the frames and ensuring the stability of the fragments on the substrate.
2.2. Data Collection and Analysis
During the 1-year monitoring, four key parameters of the coral fragments were analyzed: (i) growth rate, (ii) survival rate, (iii) presence of parasites, and (iv) presence of diseases (including bleaching). The presence of the common corallivore Drupella spp. was also monitored, but due to its absence during the experiment, it was not included among the investigated parameters. During each underwater survey, the 242 fragments were measured by divers to the nearest millimeter with a Vernier caliper along the longest axis of the coral fragment to track their growth. The growth rate of the corals (Gr) was calculated with the formula Gr = T2 − T1, where T1 represents the measurement of the fragment at time 1, and T2 represents the measurement of the same fragment at the subsequent time 2. Monthly average growth rates were calculated with standard errors.
Coral mortality was assessed at each survey from the number of dead fragments with respect to the total number of fragments and then averaged over the months. The survival rate of the corals was computed from the number of living coral fragments found at the end of the 1-year monitoring.
Photographs of each coral fragment were also taken from different angles during each monitoring time to assess the overall health state of the fragments. The presence of parasites and signals of diseases were visually assessed from the analysis of photographs and reported as the % of parasitized (or diseased) fragments found each month with respect to the total number of fragments. For each dead fragment, the putative cause (i.e., parasites, diseases, or bleaching) was also identified.
Inspired by the protocol of Dehnert et al. [43], the health condition of each coral fragment was classified using 4 categories based on the amount of living tissue remaining on the fragment: (1) 100% of living tissue; (2) 100% > living tissue ≥ 50%; (3) <50% of living tissue; and (4) pale or partially bleached fragment. The frequency of each category was measured every month.
To evaluate the effects of thermal stress on the fragments’ state of health, the average monthly growth rate, the average coral mortality, and the average number of new diseased or parasitized corals per month were compared to the average monthly sea surface temperature (SST) of the month before (under the hypothesis that corals need some weeks before reacting to thermal stress). Monthly average local SST data for the period December 2021 and October 2022 were obtained from the Maldives Meteorological Service, which has a station near Hululé Island, approximately 5 km from the restoration site. The average number of new diseased or parasitized corals per month was also compared to the average coral mortality. Correlations between all the investigated variables (growth rate, parasitized corals, diseased corals, local SST, and coral mortality) were performed using the Pearson correlation test using the Past3 software.
3. Results
3.1. Growth Rate and Thermal Anomaly
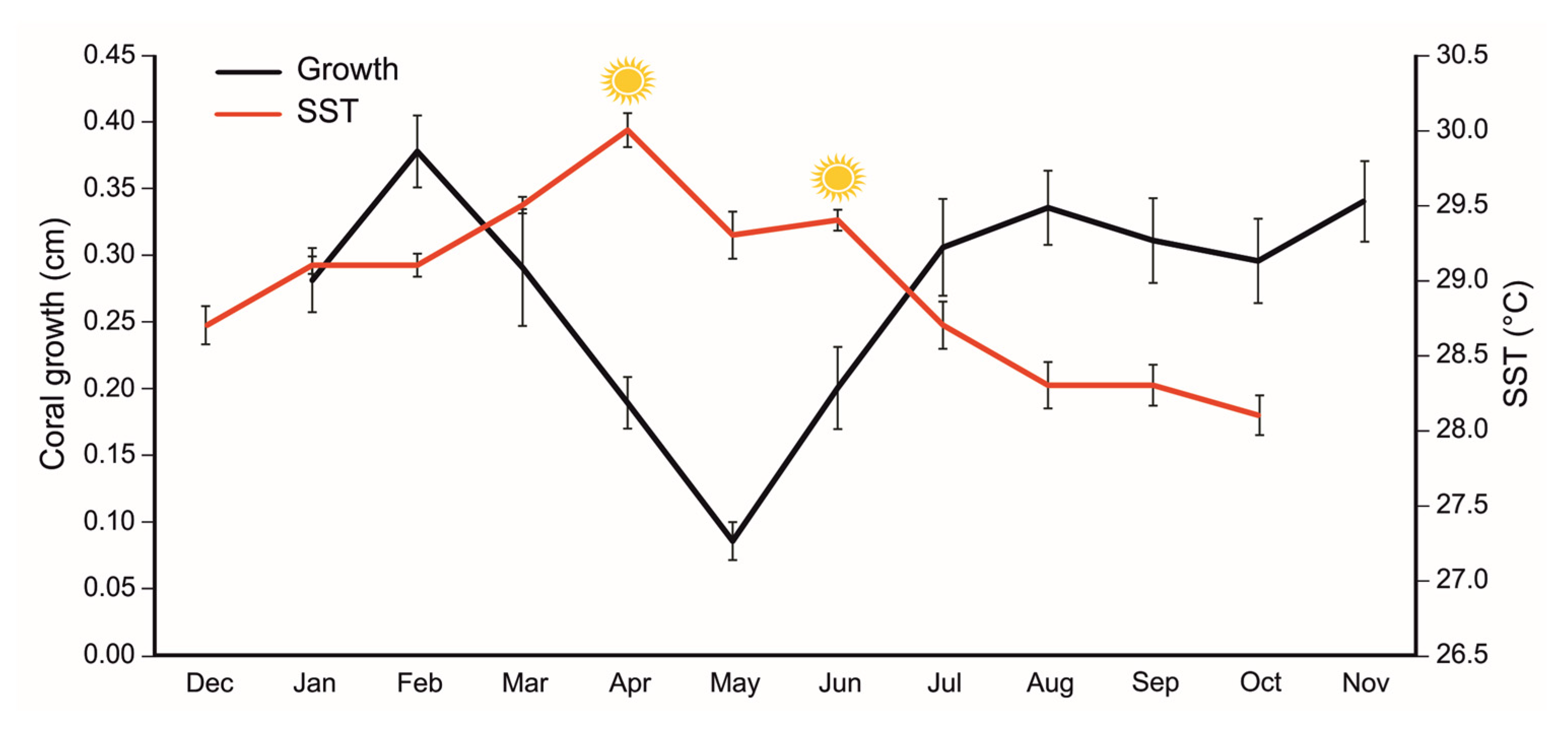
Coral fragments reached the highest growth rate (0.38 ± 0.03 cm) in the month of February 2022, two months after their translocation to the coral nursery (Figure 3, Table S1). Then, the coral growth started to decrease in parallel with the increase in the water temperature, reaching the lowest monthly growth rate in May (0.09 ± 0.01 cm), just one month after the peak in the mean temperature of April (30 ± 0.1 °C) (Figure 3, Table S1). After May, the growth of the corals started to increase again until August (0.34 ± 0.03 cm), in parallel with the decrease of the water temperature. The coral growth remained stable during the last months of the monitoring period.
A significant negative relationship was found between the monthly growth rate of coral fragments and the monthly local SST (r = −0.71, n = 11), being the temporal trend of the two variables specular.
3.2. Survival Rate, Spreading of Diseases and Parasites
After one year of monitoring, the final survival rate of 70% was registered. The highest percentage of coral mortality was reached in February (7% of the fragments), two months after the translocation of fragments to the coral nursery, and in October (8%), whilst in all the other months, the coral mortality ranged between 0% and 5% (Figure 4a, Table S2). A non-significant relationship was found between coral mortality and water temperature (r = −0.09, n = 11).
Diseases were recorded in 53% of the coral fragments by the end of the monitoring period. The highest percentages of diseased corals were recorded in the first months after the translocation of fragments to the coral nursery, i.e., in January (9%), February (8%), March (9%), and May (8%), whilst in all the other months the percentage of newly diseased corals was lower and ranged between 0% and 4% (Figure 4b, Table S2). Two main diseases on the coral fragments were visually identified from the picture analysis and the observation on the field, i.e., the skeletal eroding band (SEB) disease and the rapid tissue necrosis (RTN) (Figure 5a–c).
By the end of the monitoring period, 72% of the coral fragments presented at least one parasite. Parasites spread rapidly in the first four months after the translocation, between December and March, with a maximum number of new parasites recorded in March (14.9%) (Figure 4c, Table S2). By June, 61% of coral fragments were infested with parasites. Two different parasites were predominant in the coral fragments, i.e., the vermetid worm snail Dendropoma maxima and a smaller organism resembling the hermit crab Paguritta spp. (Figure 5d–f). It was impossible to identify the latter parasite with more taxonomical detail as it was not clearly visible from the pictures. Differently from the vermetids, the crabs did not determine the death of the host, but when they reached high abundances in the same colony, they caused a stress response.
The highest rate of coral mortality was experienced by those corals mostly affected by diseases (r = 0.63, n = 12) and by parasites (r = 0.75, n = 12).
3.3. Health Condition of Coral Fragments
The number of fragments with 100% living tissue decreased to about 36% by the end of the year, whilst fragments with more than 50% living tissue became the dominant category from March till the end of the monitoring period (Figure 6, Table S3). The frequency of fragments with less than 50% of living tissue started increasing in July due to the spreading of diseases. The frequency of paled and/or partially bleached fragments showed the highest values in December (27%) when the coral fragments were transferred to the nursery and in April (20%) during the heat wave. From June, the frequencies of bleached corals became negligible or absent, as they successfully recovered when the temperature returned to normal values (Figure 7).
4. Discussion
In this study, an innovative methodology to restore a degraded coral reef in a highly impacted area of the Maldives has been performed. The survival rate of corals in the nursery recorded 1-year later was higher than 70%, indicative of the good resilience of coral fragments notwithstanding the various stressors that undermined their growth. During the 1-year experiment, coral fragments were indeed subject to a number of stressors, namely: (i) mechanical stress due to fragging and translocation to the nursery; (ii) thermal stress due to the heat wave that occurred 5 months after their translocation; (iii) spread of parasites and diseases; and (iv) overall poor water quality of the area. Although our survival rate is lower when compared to other examples of coral restoration performed in the Maldives [43,44], which had about 80–90% survival after the first year, the location of Villimalè Island must be considered in the performance of the experiment, as well as the source of the fragments, which were collected from donor-impacted colonies.
Villimalè proximity to the capital city of Malé and to the garbage island of Thilafushi means an inferior water quality in this area. The high level of human pressure and the huge amount of chemical pollutants and organic waste found in its waters [40,41] make Villimalé a challenging location for corals to grow. The low monthly growth rate registered throughout the year (always <0.4 cm) is likely to be due to the origin of the coral fragments from the impacted colonies of the donor reef, as well as to the local stressful conditions caused by the poor environmental status, as already shown by other studies [43,44,45,46]. It should, therefore, come as no surprise that any restoration program undertaken in these highly impacted locations could be slower and potentially less successful than in other less developed areas. The site-specific characteristics of the coral nursery contributed to the success of the restoration and must, therefore, be considered, especially during the project planning [43]. Moreover, the integrity and maintenance of the three-dimensional structure of the reef must be taken into account to guarantee potential successful restoration interventions since it will provide elevated substrates for the natural recruiting of coral larvae. Even after most of the coral colonies die following a disturbance, the 3D structure of the reef remains long before being eroded and destroyed [28]. In such a situation, an active and prompt restoration intervention becomes necessary to help reefs recover, and it also might reduce the spread of algal overgrowth on dead corals, preventing the risk of irreversible ecosystem shifts.
While dealing with local stressors, transplanted coral fragments also faced a severe heat wave that reached the country between March and April 2022. The increase in the SST also challenged the restored corals, causing a sharp decline in their growth rates for more than three months. Although coral growth has been significantly affected by the water temperature anomaly, it did not cause significant mortality of coral fragments. Fragments appeared pale or partially bleached from March to May, but they successfully recovered when the temperature returned to normal values, showing a high resilience to thermal stress even if the peak of 31.1 °C reached by the SST in April exceeded the severe bleaching threshold [28]. Despite thermal anomalies being one of the most important drivers of coral mortality, results from our restoration experiment are promising and suggest that for planning successful restoration programs, the starting time of the translocation must be carefully selected to avoid those periods of the year more affected by thermal anomalies (i.e., from April to May in the Maldives). Corals are likely to be already acclimatized to the new environmental conditions after some months from the translocation and are thus able to better resist further stress.
Mortality of coral fragments (30% of the translocated fragments after 1 year) was mainly caused by the spread of diseases and parasites. The highest percentage of diseased corals affected by the skeletal eroding band disease [31] and rapid tissue necrosis [47] was recorded in the first months after the translocation (from January to March), likely because the mechanical stress caused by the fragging and the attaching made the fragments more vulnerable to infection. This was also confirmed by the high frequency of pale or partially bleached fragments in the first months of the experiment. In addition, the polluted waters of the North Malé Area favored a high presence of pathogens [31,43]. The extent and the frequency of coral diseases in the Indo-Pacific region have been shown to increase continuously, becoming an imperative environmental issue worldwide that will require further investigation and conservation efforts [48]. This threat is also exacerbated by the instability of climate [49], which negatively affects coral reefs, increasing pathogens’ abundance and virulence [33] and simultaneously decreasing corals’ resistance to pathogens.
Parasites spread rapidly since the fragments were translocated, with more than 80% of parasitized colonies by the end of the year. The spreading of both diseases and parasites was similarly responsible for coral mortality and had the same peaks after 2–4 months from the translocation, confirming the susceptibility of stressed corals to parasites. Infection by diseases is likely to reduce coral growth, making the fragments more susceptible to the establishment of parasites. Parasite abundance has long been suggested as an indicator of ecosystem stress [50], and some species of parasites have been pointed out as bio-indicators of environmental degradation [51].
Fragmentation and transplantation of coral fragments collected from impacted colonies showed effectiveness in restoring a highly degraded reef despite the presence of diseases and parasites. This intervention represents a feasible mitigation measure for biodiversity conservation instead of replanting with corals collected from healthier but more distant donor reefs. Although the reduced growth of corals might require a longer time before the new colonies will reach the suitable size for reproduction and, eventually, will be ready for relocation to other reefs, this challenging approach could revolutionize the coral restoration of highly impacted reefs when the translocation of healthy coral colonies from donor reefs is not an option. Moreover, the restoration methodology experimented with in this study required fewer resources and lower costs compared to the typical restoration approaches, making it a good option for small realities that, otherwise, may not have the means to restore their reefs. Locally tested methodologies and economically viable project designs should be more implemented rather than a single restoration approach scaled up [21]. The Maldives has few examples of coral restoration projects, especially in highly populated islands inhabited by local communities. These findings will be useful to involve the government in planning future restoration programs, mainly on those local islands with limited resources.
Cheap solutions for restoration interventions, along with community-based support, open the opportunity for a broad application of similar studies to restore highly impacted reefs that usually occur in the most populated tropical areas [52,53]. It is vital to act locally and reverse coral reef decline and biodiversity loss through coral restoration initiatives by enhancing coral resilience [18]. The human dimension is particularly relevant to coral restoration since people are involved in all stages of the process, from design to execution and monitoring [37]. The direct involvement of local communities and volunteers can improve local and global management of reef resources [54], increase awareness of threats to coral reefs, and involve every citizen in helping preserve and restore their reefs. After the success of this study, the Villimalé community is still deeply involved in coral restoration activities, and it is currently expanding the restoration site. The good feedback received on the project after one year confirms that the locals are connected to the environment and understand the value of a healthy reef. To upscale similar projects around the Country, it is necessary to increase local knowledge and environmental awareness and directly involve the community in the restoration through restoration training programs.
As a final remark, to ensure successful restoration interventions in highly anthropized and polluted areas, periodic monitoring activities for the maintenance of the nursery are mandatory, especially for cleaning the frames and ensuring the stability of the fragments on the substrate. Without this effort, all the translocated coral colonies would have died before the end of the experiment. We recommend promptly removing the diseased fragments to avoid spreading to the surrounding corals to maximize the survival rate. We also suggest being more cautious with the parasites since both vermetids and crabs grow beneath the coral tissue, and their removal can lead to severe damage to the fragments. In particular, Dendropoma maxima should be removed in its younger stages when the calcareous tube is smaller and more fragile.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/d15091022/s1, Table S1: Average monthly growth rate of corals (in cm) and of SST (in °C) with their standard errors; Table S2: Monthly number of dead, diseased, and parasitized corals; Table S3: Monthly number of coral fragments for each healthy category.
Author Contributions
Conceptualization, I.P. and H.A.; methodology, I.P. and H.A.; software, M.M. and I.P.; validation, M.M., I.P. and H.A.; formal analysis, I.P., K.F., C.d.N. and M.M.; investigation, I.P., K.F., C.d.N. and H.A.; resources, H.A.; data curation, I.P. and K.F.; writing—original draft preparation, I.P. and K.F.; writing—review and editing, M.M., I.P. and K.F.; visualization, I.P. and M.M.; supervision, M.M. and I.P.; project administration, I.P. and H.A.; funding acquisition, H.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data are available on request to the corresponding author due to restrictions and ownership of the NGO Save the Beach Maldives.
Acknowledgments
The Maldivian NGO Save the Beach Maldives supported our research, allowing the study to be consistent for one year. We especially want to thank all the participants in field activities that helped to collect data.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Shaver, E.C.; Silliman, B.R. Time to cash in on positive interactions for coral restoration. PeerJ 2017, 5, e3499. [Google Scholar] [CrossRef] [PubMed]
- Silveira, C.B.; Cavalcant, G.S.; Walter, J.M.; Silva-Lima, A.W.; Dinsdale, E.A.; Bourne David, G.; Thompson, C.C.; Thompson, F.L. Microbial processes driving coral reef organic carbon flow. FEMS Microbiol. Rev. 2017, 41, 575–595. [Google Scholar] [CrossRef] [PubMed]
- Reaka-Kudla, M.L. The global biodiversity of coral reefs: A comparison with rain forests. In Biodiversity II: Understanding and Protecting Our Biological Resources; Reaka-Kudla, M.L., Wilson, D.E., Wilson, E.O., Eds.; Joseph Henry/National Academy Press: Washington, DC, USA, 1997; pp. 83–108. [Google Scholar]
- Heery, E.C.; Hoeksema, B.W.; Browne, N.K.; Reimer, J.D.; Ang, P.O.; Huang, D.; Friess, D.A.; Chou, L.M.; Loke, L.H.L.; Saksena-Taylor, P.; et al. Urban coral reefs: Degradation and resilience of hard coral assemblages in coastal cities of East and Southeast Asia. Mar. Pollut. Bull. 2018, 135, 654–681. [Google Scholar] [CrossRef] [PubMed]
- Olguín-López, N.; Gutiérrez-Chavez, C.; Hérnández-Elizárraga, V.H.; Ibarra-Alvarado, C.; Rojas-Molina, A. Coral reef bleaching: An ecological and biological overview. In Corals in a Changing World; Carmenza, D., Edisson, T.C., Eds.; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef]
- Cinner, J. Coral reef livelihoods. COSUST 2014, 7, 65–71. [Google Scholar] [CrossRef]
- Heron, S.F.; Eakin, C.M.; Douvere, F.; Anderson, K.L.; Day, J.C.; Geiger, E.; Hoegh-Guldberg, O.; Van Hooidonk, R.; Hughes, T.; Marshall, P.; et al. Impacts of Climate Change on World Heritage Coral Reefs: A First Global Scientific Assessment; National Oceanic and Atmospheric Administration: Washington, DC, USA, 2017. Available online: https://repository.library.noaa.gov/view/noaa/16386 (accessed on 12 August 2023).
- Reguero, B.G.; Beck, M.W.; Agostini, V.N.; Kramer, P.; Hancock, B. Coral reefs for coastal protection: A new methodological approach and engineering case study in Grenada. J. Environ. Manag. 2018, 210, 146–161. [Google Scholar] [CrossRef] [PubMed]
- Westoby, R.; McNamara, K.E. Fear, grief, hope and action. Nat. Clim. Chang. 2019, 9, 500–501. [Google Scholar] [CrossRef]
- Morrison, T.H.; Hughes, T.P.; Adger, W.N.; Brown, K.; Barnett, J.; Lemos, M.C. Save reefs to rescue all ecosystems. Nature 2019, 573, 333–336. [Google Scholar] [CrossRef]
- Montefalcone, M.; Morri, C.; Bianchi, C.N. Influence of local pressures on Maldivian coral reef resilience following repeated bleaching events, and recovery perspectives. Front. Mar. Sci. 2020, 7, 587. [Google Scholar] [CrossRef]
- Pancrazi, I.; Ahmed, H.; Cerrano, C.; Montefalcone, M. Synergic effect of global thermal anomalies and local dredging activities on coral reefs of the Maldives. Mar. Pollut. Bull. 2020, 160, 111585. [Google Scholar] [CrossRef]
- Jaap, W.C. Coral reef restoration. Ecol. Eng. 2000, 15, 345–364. [Google Scholar] [CrossRef]
- United Nations Decade in Ecosystem Restoration 2021–2030. Available online: https://www.decadeonrestoration.org/about-un-decade (accessed on 12 August 2023).
- Boström-Einarsson, L.; Babcock, R.C.; Bayraktarov, E.; Ceccarelli, D.; Cook, N.; Ferse Sebastian, C.A.; Hancock, B.; Harrison, P.; Hein, M.; Shaver, E.; et al. Coral restoration-A systematic review of current methods, successes, failures and future directions. PLoS ONE 2020, 15, e0226631. [Google Scholar] [CrossRef] [PubMed]
- Rinkevich, B. Rebuilding coral reefs: Does active reef restoration lead to sustainable reefs? COSUST 2014, 7, 28–36. [Google Scholar] [CrossRef]
- Possingham, H.P.; Bode, M.; Klein, C.J. Optimal conservation outcomes require both restoration and protection. PLoS Boil. 2015, 13, e1002052. [Google Scholar] [CrossRef] [PubMed]
- Westoby, R.; Becken, S.; Laria, A.P. Perspectives on the human dimensions of coral restoration. Reg. Environ. Chang. 2020, 20, 109. [Google Scholar] [CrossRef]
- Bowden-Kerby, A. Coral Transplantation and Restocking to Accelerate the Recovery of Coral Reef Habitats and Fisheries Resources within No-Take Marine Protected Areas: Hands-on Approaches to Support Community-Based Coral Reef Management; ITMEMS, AquaDocs.org: Manila, Philippines, 2003; Available online: https://aquadocs.org/handle/1834/849 (accessed on 12 August 2023).
- Rinkevich, B. Management of coral reefs: We have gone wrong when neglecting active reef restoration. Mar. Pollut. Bull. 2008, 56, 1821–1824. [Google Scholar] [CrossRef]
- Dehnert, I.; Galli, P.; Montano, S. Ecological impacts of coral gardening outplanting in the Maldives. Restor. Ecol. 2022, 31, e13783. [Google Scholar] [CrossRef]
- Morgan, K.M.; Kench, P.S. Reef to island sediment connections on a Maldivian carbonate platform: Using benthic ecology and biosedimentary depositional facies to examine island-building potential. Earth Surf. Process. Landf. 2016, 41, 1815–1825. [Google Scholar] [CrossRef]
- Dhunya, A.; Huang, Q.; Aslam, A. Coastal habitats of Maldives: Status, trends, threats, and potential conservation strategies. Int. J. Sci. Eng. Res. 2017, 8, 47–62. [Google Scholar]
- Giampiccoli, A.; Muhsin, B.A.; Mtapuri, O. Community-based tourism in the case of the Maldives. Geoj. Tour. Geosites 2020, 29, 428–439. [Google Scholar] [CrossRef]
- The World Bank IBRD-IDA. Maldives-Wetland Conservation and Coral Reef Monitoring for Adaptation to Climate Change Project. Available online: https://documents.worldbank.org/en/publication/documents-reports/documentdetail/621291468283182902/maldives-wetlands-conservation-and-coral-reef-monitoring-for-adaptation-to-climate-change-project-restructuring (accessed on 12 August 2023).
- Wadey, M.; Brown, S.; Nicholls, R.J.; Haigh, I. Coastal flooding in the Maldives: An assessment of historic events and their implications. Nat. Hazards 2017, 89, 131–159. [Google Scholar] [CrossRef]
- Dijkstra, H.A. The ENSO phenomenon: Theory and mechanisms. Adv. Geosci. 2006, 6, 3–15. [Google Scholar] [CrossRef]
- Montefalcone, M.; Morri, C.; Bianchi, C.N. Long-term change in bioconstruction potential of Maldivian coral reefs following extreme climate anomalies. Glob. Chang. Biol. 2018, 24, 5629–5641. [Google Scholar] [CrossRef]
- Maldives Bureau of Statistics, Ministry of National Planning Housing & Infrastructure. Available online: https://statisticsmaldives.gov.mv/census-in-2022/ (accessed on 12 August 2023).
- Munavvar, R. Tragedy of the Maldives. Of Fleeting Paradise, Enduring World Power and a Great Sense of Responsibility. The Edition Section “Climate Change”. 2019. Available online: https://edition.mv/comic_of_the_day/12528 (accessed on 12 August 2023).
- Montano, S.; Strona, G.; Seveso, D.; Maggioni, D.; Galli, P. Widespread occurrence of coral diseases in the central Maldives. Mar. Freshw. Res. 2016, 67, 1253–1262. [Google Scholar] [CrossRef]
- Harvell, C.D.; Mitchell, C.E.; Ward, J.R.; Altizer, S.; Dobson, A.P.; Ostfeld, R.S.; Samuel, M.D. Climate warming and disease risks for terrestrial and marine biota. Science 2002, 296, 2158–2162. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.W.; Richardson, L.L. Emerging coral diseases: A temperature-driven process? Mar. Ecol. 2014, 36, 278–291. [Google Scholar] [CrossRef]
- Sokolow, S. Effects of a changing climate on the dynamics of coral infectious disease: A review of the evidence. Dis. Aquat. Org. 2009, 87, 5–18. [Google Scholar] [CrossRef]
- Barton, J.A.; Bourne, D.G.; Humphrey, C.; Hutson, K.S. Parasites and coral-associated invertebrates that impact coral health. Rev. Aquac. 2020, 12, 2284–2303. [Google Scholar] [CrossRef]
- Montano, S. The extraordinary importance of coral-associated fauna. Diversity 2020, 12, 357. [Google Scholar] [CrossRef]
- Hein, M.Y.; Birtles, A.; Willis, B.L.; Gardiner, N.; Beeden, R.; Marshall, N.A. Coral restoration: Socio-ecological perspectives of benefits and limitations. Biol. Conserv. 2019, 229, 14–25. [Google Scholar] [CrossRef]
- Boskalis. Available online: https://boskalis.com/press/press-releases-and-company-news/boskalis-acquires-eur-120-million-contract-for-development-and-climate-adaptive-measures-for-gulhifalhu-in-the-maldives (accessed on 12 August 2023).
- The Maldives Journal Fourth Pillar of Democracy at Work. Available online: https://themaldivesjournal.com/28744 (accessed on 12 August 2023).
- Majeedha, M. State of the Environment 2016. Ministry of Environment, Climate Change and Technology Republic of Maldives. 2017. Available online: https://www.environment.gov.mv/v2/en/download/4270 (accessed on 12 August 2023).
- Peterson, C. Assessment of Solid Waste Management Practices and Its Vulnerability to Climate Risks in Maldives Tourism Sector. Report Submitted to Ministry of Tourism, Arts and Culture, Malé Republic of Maldives. 2013. Available online: Chrome-extension://efaidnbmnnnibpcajpcglclefindmkaj/https://archive.tourism.gov.mv/downloads/tap/2014/Solid_Waste.pdf (accessed on 12 August 2023).
- NOAA Satellite and Informative Service, National Environmental Satellite, Data, and Information Service (NESDIS), Coral Reef Watch. Available online: https://coralreefwatch.noaa.gov/product/vs/gauges/maldives.php (accessed on 12 August 2023).
- Dehnert, I.; Saponari, L.; Isa, V.; Seveso, D.; Galli, P.; Montano, S. Exploring the performance of mid-water lagoon nurseries for coral restoration in the Maldives. Restor. Ecol. 2021, 30, e13600. [Google Scholar] [CrossRef]
- Schopmeyer, S.A.; Lirman, D.; Bartels, E.; Gilliam, D.S.; Goergen, E.A.; Griffin, S.P.; Johnson, M.E.; Lustic, C.; Maxwell, K.; Walter, C.S. Regional Restoration Benchmarks for Acropora cervicornis. Coral Reefs 2017, 36, 1047–1057. [Google Scholar] [CrossRef]
- Cooper, T.F.; Gilmour, J.P.; Fabricius, K.E. Bioindicators of changes in water quality on coral reefs: Review and recommendations for monitoring programmes. Coral Reefs 2009, 28, 589–606. [Google Scholar] [CrossRef]
- Browne, N.K.; Tay, J.K.; Low, J.; Larson, O.; Todd, P.A. Fluctuations in coral health of four common inshore reef corals in response to seasonal and anthropogenic changes in water quality. Mar. Environ. Res. 2015, 105, 39–52. [Google Scholar] [CrossRef] [PubMed]
- Luna, G.M.; Biavasco, F.; Danovaro, R. Bacteria associated with the rapid tissue necrosis of stony corals. Environ. Microbiol. 2007, 9, 1851–1857. [Google Scholar] [CrossRef]
- Sutherland, W.J.; Clout, M.; Depledge, M.; Dicks, L.V.; Dinsdale, J.; Entwistle, A.C.; Fleishman, E.; Gibbons, D.W.; Keim, B.; Lickorish, F.A.; et al. A horizon scan of global conservation issues for 2015. Trends Ecol. Evol. 2015, 30, 17–24. [Google Scholar] [CrossRef]
- Mizuta, R.; Arakawa, O.; Ose, T.; Kusunoki, S.; Endo, H.; Kitoh, A. Classification of CMIP5 future climate responses by the tropical sea surface temperature changes. Sola 2014, 10, 167–171. [Google Scholar] [CrossRef]
- Vidal-Martínez, V.M.; Pech, D.; Sures, B.; Purucker, S.T.; Poulin, R. Can parasites really reveal environmental impact? Trends Parasitol. 2010, 26, 44–51. [Google Scholar] [CrossRef]
- Marcogliese, D.J. Parasites of the superorganism: Are they indicators of ecosystem health? Int. J. Parasitol. 2005, 35, 705–716. [Google Scholar] [CrossRef]
- Sandin, S.A.; Smith, J.E.; DeMartini, E.E.; Dinsdale, E.A.; Donner, S.D.; Friedlander, A.M.; Konotchick, T.; Malay, M.; Maragos, J.E.; Obura, D.; et al. Baselines and degradation of coral reefs in the Northern Line Islands. PLoS ONE 2008, 3, e1548. [Google Scholar] [CrossRef]
- González-Díaz, P.; González-Sansón, G.; Aguilar Betancourt, C.; Álvarez Fernández, S.; Perera Pérez, O.; Hernández Fernández, L.; Rodríguez, V.M.F.; Caballero, Y.C.; Armenteros, M.; de la Guardia Llanso, E. Status of Cuban coral reefs. Bull. Mar. Sci. 2018, 94, 229–247. [Google Scholar] [CrossRef]
- Dean, A.J.; Church, E.K.; Loder, J.; Fielding, K.S.; Wilson, K.A. How do marine and coastal citizen science experiences foster environmental engagement? J. Environ. Manag. 2018, 213, 409–416. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Map of the Maldives with the location of Villimalé Island in the North Malé atoll, close to the capital city of Malé and to the garbage island of Thilafushi. The symbol with the corals shows the location of the impacted reef and the nursery area on Villimalé Island, where the restoration experiment was performed.
Figure 1.
Map of the Maldives with the location of Villimalé Island in the North Malé atoll, close to the capital city of Malé and to the garbage island of Thilafushi. The symbol with the corals shows the location of the impacted reef and the nursery area on Villimalé Island, where the restoration experiment was performed.

Figure 2.
Metal frame double-coated with resin and sand with corals growing on it at the end of the 1-year experiment (left panel); an example of a coral fragment attached to the metal frame at the beginning of the experiment (upper-right panel); and the same fragment one year after (lower-right panel).
Figure 2.
Metal frame double-coated with resin and sand with corals growing on it at the end of the 1-year experiment (left panel); an example of a coral fragment attached to the metal frame at the beginning of the experiment (upper-right panel); and the same fragment one year after (lower-right panel).

Figure 3.
Mean (±se) values of the monthly growth rate of the coral fragments in the nursery (black line) and mean (±se) values of the sea surface temperature (SST, red line). The symbols of suns represent the two peaks registered in the water temperature.
Figure 3.
Mean (±se) values of the monthly growth rate of the coral fragments in the nursery (black line) and mean (±se) values of the sea surface temperature (SST, red line). The symbols of suns represent the two peaks registered in the water temperature.

Figure 4.
Bar plots showing the cumulative monthly fraction of (a) coral mortality, (b) diseased corals, and (c) parasitized corals, expressed as the % of the total coral fragments. The white portions in the bars indicate the % of new dead coral fragments (a), new diseased fragments (b), and new parasitized fragments (c) found every month.
Figure 4.
Bar plots showing the cumulative monthly fraction of (a) coral mortality, (b) diseased corals, and (c) parasitized corals, expressed as the % of the total coral fragments. The white portions in the bars indicate the % of new dead coral fragments (a), new diseased fragments (b), and new parasitized fragments (c) found every month.

Figure 5.
Examples of diseases and parasites: (a) the rapid tissue necrosis (RTN) disease and (b) its progress after 2 months when the algae started covering the dead portions of the coral; (c) the skeletal eroding band (SEB) disease with occurrence of the ciliates Halofolliculina corallasia responsible for the disease (black dots on the white skeleton); (d) juveniles and (e) adults of the vermetids Dendropoma maxima with the mucus net secreted to capture its planktonic prey. Due to this mucus, polyps of the coral are not able to feed anymore, and the colony slowly dies. At the juvenile stage, the tube is fragile and easily removed; (f) the other parasite found in the coral nursery resembling the hermit crab Paguritta spp. The pink color around the parasites indicates a stressful response of the coral.
Figure 5.
Examples of diseases and parasites: (a) the rapid tissue necrosis (RTN) disease and (b) its progress after 2 months when the algae started covering the dead portions of the coral; (c) the skeletal eroding band (SEB) disease with occurrence of the ciliates Halofolliculina corallasia responsible for the disease (black dots on the white skeleton); (d) juveniles and (e) adults of the vermetids Dendropoma maxima with the mucus net secreted to capture its planktonic prey. Due to this mucus, polyps of the coral are not able to feed anymore, and the colony slowly dies. At the juvenile stage, the tube is fragile and easily removed; (f) the other parasite found in the coral nursery resembling the hermit crab Paguritta spp. The pink color around the parasites indicates a stressful response of the coral.

Figure 6.
Frequency of each category of the health condition of coral fragments. The symbols on the top indicate the stressor that mainly affected the coral fragments in the months with the major impacts: hammer and chisel indicate the mechanical stress resulting from the fragging and attaching of the fragments; thermometer indicates peaks in sea surface temperature; pathogen indicates spreading of diseases and parasites.
Figure 6.
Frequency of each category of the health condition of coral fragments. The symbols on the top indicate the stressor that mainly affected the coral fragments in the months with the major impacts: hammer and chisel indicate the mechanical stress resulting from the fragging and attaching of the fragments; thermometer indicates peaks in sea surface temperature; pathogen indicates spreading of diseases and parasites.

Figure 7.
A bleached fragment observed in April 2022 during the peak in SST (left panel), and the same fragment fully recovered after 2 months (in June 2022, right panel).
Figure 7.
A bleached fragment observed in April 2022 during the peak in SST (left panel), and the same fragment fully recovered after 2 months (in June 2022, right panel).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pancrazi, I.; Feairheller, K.; Ahmed, H.; di Napoli, C.; Montefalcone, M. Active Coral Restoration to Preserve the Biodiversity of a Highly Impacted Reef in the Maldives. Diversity 2023, 15, 1022. https://doi.org/10.3390/d15091022
AMA Style
Pancrazi I, Feairheller K, Ahmed H, di Napoli C, Montefalcone M. Active Coral Restoration to Preserve the Biodiversity of a Highly Impacted Reef in the Maldives. Diversity. 2023; 15(9):1022. https://doi.org/10.3390/d15091022
Chicago/Turabian StylePancrazi, Irene, Kayla Feairheller, Hassan Ahmed, Carolina di Napoli, and Monica Montefalcone. 2023. "Active Coral Restoration to Preserve the Biodiversity of a Highly Impacted Reef in the Maldives" Diversity 15, no. 9: 1022. https://doi.org/10.3390/d15091022
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.