
Global Warming Drives Transitions in Suitable Habitats and Ecological Services of Rare Tinospora Miers Species in China

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Collection and Processing
2.2. Suitable Habitat Prediction and Migration Analysis
2.3. Ecological Service Value Assessment
3. Results
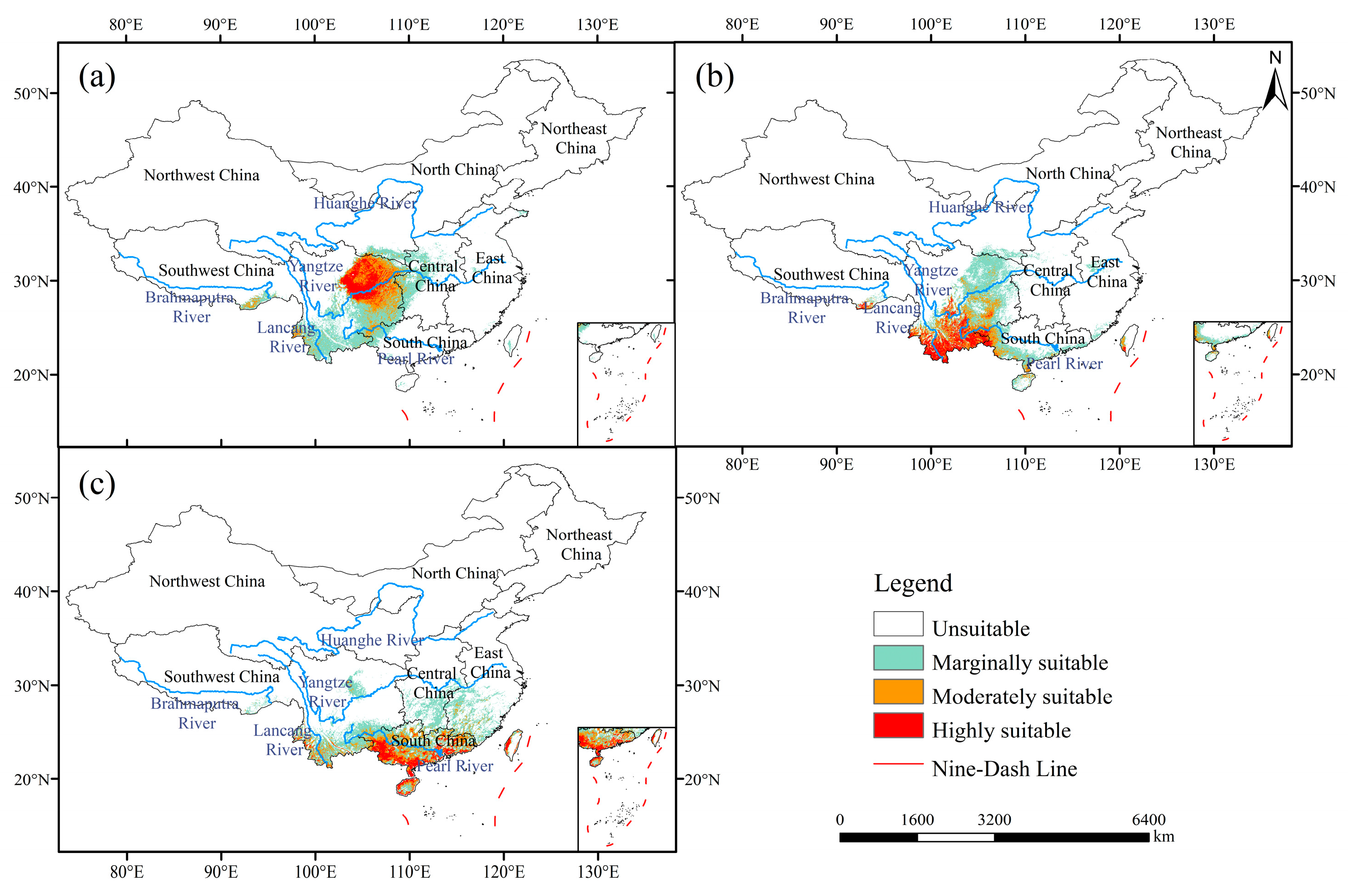
3.1. Contemporary Suitable Habitats
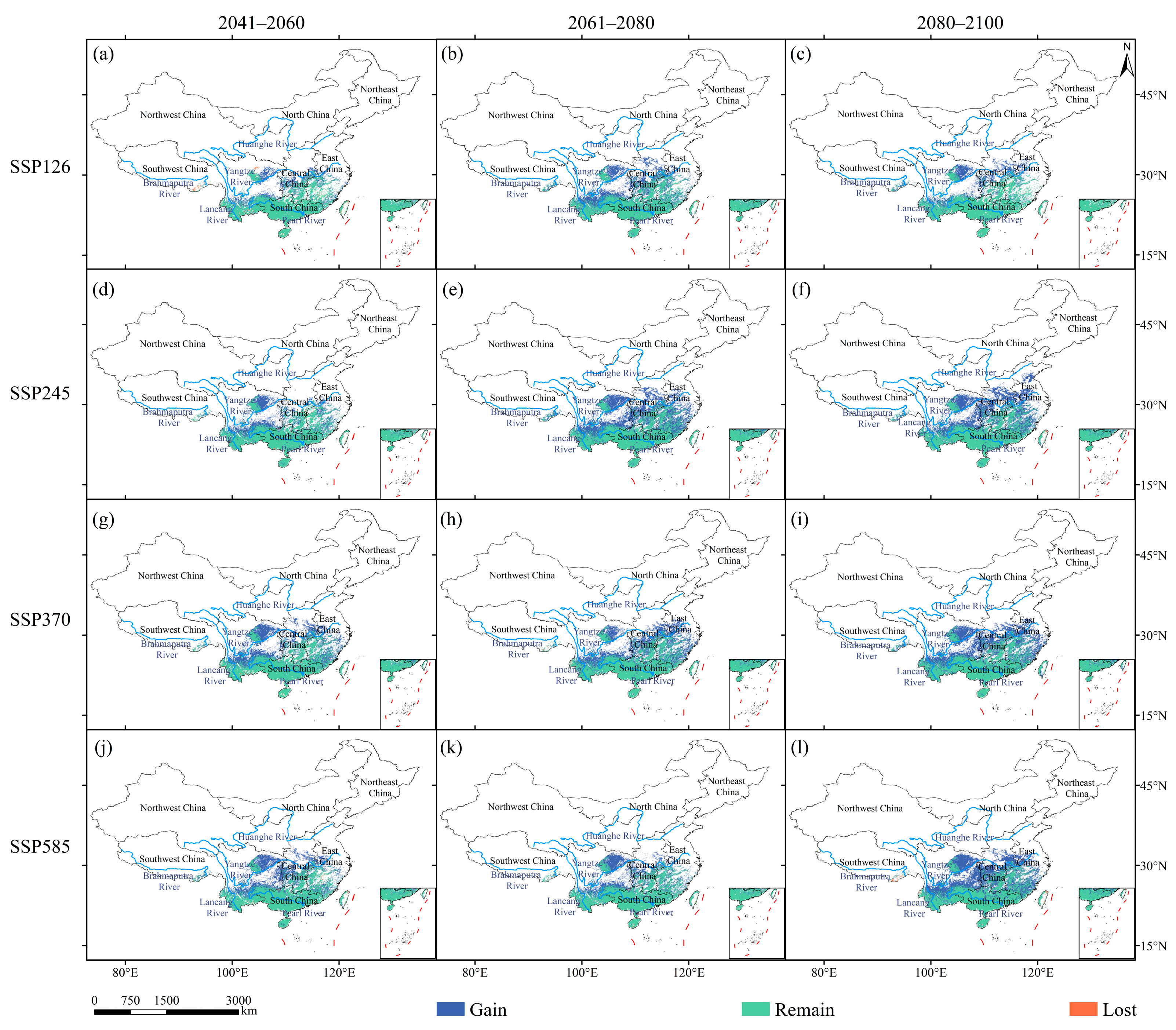
3.2. Dynamics of Suitable Habitats under Global Climate Change
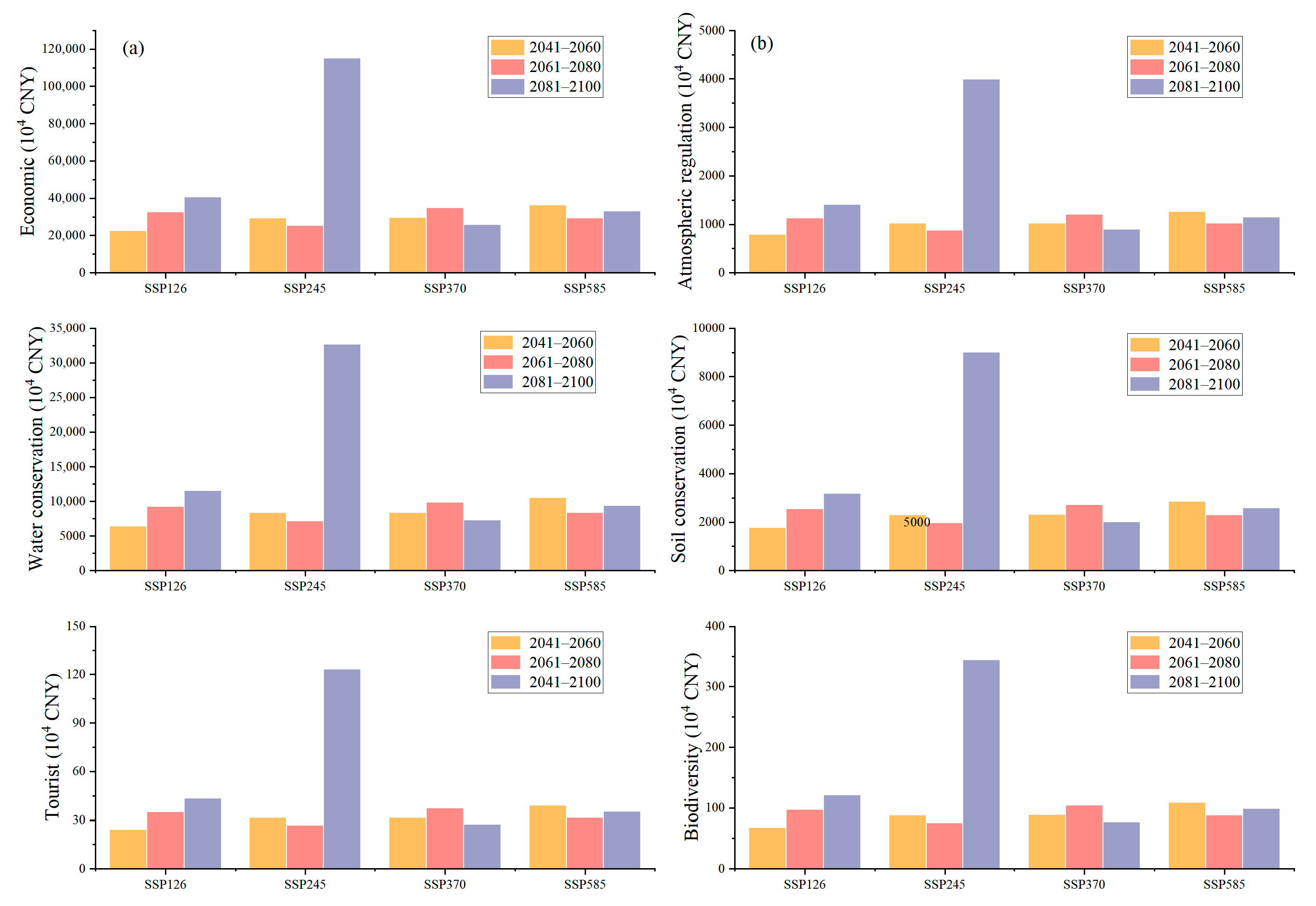
3.3. Ecological Service Value Assessments
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Malhi, Y.; Franklin, J.; Seddon, N.; Solan, M.; Turner, M.G.; Field, C.B.; Knowlton, N. Climate change and ecosystems: Threats, opportunities and solutions. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2020, 375, 20190104. [Google Scholar] [CrossRef]
- Fedele, G.; Donatti, C.I.; Harvey, C.A.; Hannah, L.; Hole, D.G. Transformative adaptation to climate change for sustainable social-ecological systems. Environ. Sci. Policy 2019, 101, 116–125. [Google Scholar] [CrossRef]
- Weiskopf, S.R.; Rubenstein, M.A.; Crozier, L.G.; Gaichas, S.; Griffis, R.; Halofsky, J.E.; Hyde, K.J.W.; Morelli, T.L.; Morisette, J.T.; Muñoz, R.C.; et al. Climate change effects on biodiversity, ecosystems, ecosystem services, and natural resource management in the United States. Sci. Total Environ. 2020, 733, 137782. [Google Scholar] [CrossRef]
- Barreto, J.R.; Pardini, R.; Metzger, J.P.; Silva, F.A.B.; Nichols, E.S. When forest loss leads to biodiversity gain: Insights from the Brazilian Atlantic Forest. Biol. Conserv. 2023, 279, 109957. [Google Scholar] [CrossRef]
- Bezeng, B.S.; Morales-Castilla, I.; van der Bank, M.; Yessoufou, K.; Daru, B.H.; Davies, T.J. Climate change may reduce the spread of non-native species. Ecosphere 2017, 8, e01694. [Google Scholar] [CrossRef]
- Bastien-Olvera, B.A.; Moore, F.C. Climate Impacts on Natural Capital: Consequences for the Social Cost of Carbon. Annu. Rev. Resour. Econ. 2022, 14, 515–532. [Google Scholar] [CrossRef]
- Chaalali, A.; Beaugrand, G.; Raybaud, V.; Lassalle, G.; Saint-Béat, B.; Le Loc’h, F.; Bopp, L.; Tecchio, S.; Safi, G.; Chifflet, M.; et al. From species distributions to ecosystem structure and function: A methodological perspective. Ecol. Model. 2016, 334, 78–90. [Google Scholar] [CrossRef]
- Ansley, R.J.; Rivera-Monroy, V.H.; Griffis-Kyle, K.; Hoagland, B.; Emert, A.; Fagin, T.; Loss, S.R.; McCarthy, H.R.; Smith, N.G.; Waring, E.F. Assessing impacts of climate change on selected foundation species and ecosystem services in the South-Central USA. Ecosphere 2023, 14, e4412. [Google Scholar] [CrossRef]
- Chi, X.; Zhang, Z.; Xu, X.; Zhang, X.; Zhao, Z.; Liu, Y.; Wang, Q.; Wang, H.; Li, Y.; Yang, G.; et al. Threatened medicinal plants in China: Distributions and conservation priorities. Biol. Conserv. 2017, 210, 89–95. [Google Scholar] [CrossRef]
- Zhang, B.Y.; Zou, H.; Chen, B.R.; Zhang, X.Y.; Kang, X.; Wang, C.; Zhang, X.X. Optimizing the distribution pattern of species under climate change: The protection and management of Phellodendron amurensein China. Front. Ecol. Evol. 2023, 11, 1186627. [Google Scholar] [CrossRef]
- De Kort, H.; Baguette, M.; Lenoir, J.; Stevens, V.M. Toward reliable habitat suitability and accessibility models in an era of multiple environmental stressors. Ecol. Evol. 2020, 10, 10937–10952. [Google Scholar] [CrossRef] [PubMed]
- Frans, V.F.; Augé, A.A.; Edelhoff, H.; Erasmi, S.; Balkenhol, N.; Engler, J.O. Quantifying apart what belongs together: A multi-state species distribution modelling framework for species using distinct habitats. Methods Ecol. Evol. 2018, 9, 98–108. [Google Scholar] [CrossRef]
- Damaneh, J.M.; Ahmadi, J.; Rahmanian, S.; Sadeghi, S.M.M.; Nasiri, V.; Borz, S.A. Prediction of wild pistachio ecological niche using machine learning models. Ecol. Inform. 2022, 72, 101907. [Google Scholar] [CrossRef]
- Latimer, A.M.; Wu, S.; Gelfand, A.E.; Silander, J.A., Jr. Building Statistical Models to Analyze Species Distributions. Ecol. Appl. 2006, 16, 33–50. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Li, J.J.; Fan, G.; He, Y. Predicting the current and future distribution of three Coptis herbs in China under climate change conditions, using the MaxEnt model and chemical analysis. Sci. Total Environ. 2020, 698, 134141. [Google Scholar] [CrossRef]
- Zhou, R.; Ci, X.; Xiao, J.; Cao, G.; Li, J. Effects and conservation assessment of climate change on the dominant group—The genus Cinnamomum of subtropical evergreen broad-leaved forests. Biodiv Sci. 2021, 29, 697–711. [Google Scholar] [CrossRef]
- Huang, Z.C.; Shu, J.P.; Yan, Y.H.; Chen, J.B. Simulation of potential distribution of Paphiopedilum in China based on the MaxEnt model. J. Trop. Subtrop. Bot. 2023, 32, 17–26. [Google Scholar]
- Velázquez-Hernández, J.M.; Ruíz-Corral, J.A.; Durán-Puga, N.; Macías, M.Á.; González-Eguiarte, D.R.; Santacruz-Ruvalcaba, F.; García-Romero, G.E.; Gallegos-Rodríguez, A. Ecogeography of Dioscorea remotiflora Kunth: An Endemic Species from Mexico. Plants 2023, 12, 3654. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, J.; Yong, Q.-Q.; Yuan, T.-H.; Wang, Q.; Li, M.-J.; Long, S.-W.; Bai, X.-X. Species Diversity and Geographical Distribution Patterns of Balsaminaceae in China. Diversity 2023, 15, 1012. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander, J.A. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Hernandez, P.A.; Graham, C.H.; Master, L.L.; Albert, D.L. The effect of sample size and species characteristics on performance of different species distribution modeling methods. Ecography 2006, 29, 773–785. [Google Scholar] [CrossRef]
- Ortega-Huerta, M.A.; Peterson, A.T. Modeling ecological niches and predicting geographic distributions: A test of six presence-only methods. Rev. Mex. Biodivers. 2008, 79, 205–216. [Google Scholar]
- Wang, Y.; Dai, E. Spatial-temporal changes in ecosystem services and the trade-off relationship in mountain regions: A case study of Hengduan Mountain region in Southwest China. J. Clean. Prod. 2020, 264, 121573. [Google Scholar] [CrossRef]
- Ying, Z.; Gao, M.X.; Liu, J.C.; Wen, Y.L.; Song, W.M. Green accounting for forest and green policies in China—A pilot national assessment. For. Policy Econ. 2011, 13, 513–519. [Google Scholar] [CrossRef]
- Wang, X.; Lv, Y.; Zhou, C.; Shen, J.Q. Value measurement and case study of development of traditional Chinese medicine resources and the protection of biodiversity. BIO Web Conf. 2017, 8, 03007. [Google Scholar] [CrossRef]
- Sharma, P.; Dwivedee, B.P.; Bisht, D.; Dash, A.K.; Kumar, D.D. The chemical constituents and diverse pharmacological importance of Tinospora cordifolia. Heliyon 2019, 5, e02437. [Google Scholar] [CrossRef]
- Novara, A.; Cerda, A.; Barone, E.; Gristina, L. Cover crop management and water conservation in vineyard and olive orchards. Soil. Tillage Res. 2021, 208, 104896. [Google Scholar] [CrossRef]
- Feng, Y. Research Advances on Production of Tinospora sagittata (Oliv.) Gagnep. or Tinospora capillipes Gagnep. Chin. J. Trop. Agric. 2021, 41, 38–43. [Google Scholar]
- Grover, J.K.; Vats, V.; Rathi, S.S. Anti-hyperglycemic effect of Eugenia jambolana and Tinospora cordifolia in experimental diabetes and their effects on key metabolic enzymes involved in carbohydrate metabolism. J. Ethnopharmacol. 2000, 73, 461–470. [Google Scholar] [CrossRef]
- Saha, S.; Ghosh, S. Tinospora cordifolia: One plant, many roles. Anc. Sci. Life 2012, 31, 151–159. [Google Scholar] [CrossRef]
- Udayabhanu; Nethravathi, P.C.; Kumar, M.A.P.; Suresh, D.; Lingaraju, K.; Rajanaika, H.; Nagabhushana, H.; Sharma, S.C. Tinospora cordifolia mediated facile green synthesis of cupric oxide nanoparticles and their photocatalytic, antioxidant and antibacterial properties. Mater. Sci. Semicond. Process. 2015, 33, 81–88. [Google Scholar] [CrossRef]
- Stanely, P.; Prince, M.; Menon, V.P. Hypoglycaemic and other related actions of Tinospora cordifolia roots in alloxan-induced diabetic rats. J. Ethnopharmacol. 2000, 70, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Weng, Y.-W.; CAI, W.-J.; Wang, C. The application and future directions of the Shared Socioeconomic Pathways (SSPs). Adv. Clim. Chang. Res. 2020, 16, 215–222. [Google Scholar]
- Zhang, Q.; Wu, M.-L.; Guo, J.; Zhang, D.-F.; Sun, C.-Z.; Xie, C.-X. Predicting the global areas for potential distribution of Gastrodia elata based on ecological niche models. Chin. J. Plant Ecol. 2017, 41, 770–778. [Google Scholar] [CrossRef]
- Luo, X.; Hu, Q.J.; Zhou, P.P.; Zhang, D.; Wang, Q.; Abbott, R.J.; Liu, J.Q. Chasing ghosts: Allopolyploid origin of Oxyria sinensis (Polygonaceae) from its only diploid congener and an unknown ancestor. Mol. Ecol. 2017, 26, 3037–3049. [Google Scholar] [CrossRef] [PubMed]
- Ranjitkar, S.; Xu, J.; Shrestha, K.K.; Kindt, R. Ensemble forecast of climate suitability for the Trans-Himalayan Nyctaginaceae species. Ecol. Model. 2014, 282, 18–24. [Google Scholar] [CrossRef]
- An, X.; Huang, T.; Zhang, H.; Yue, J.; Zhao, B. Prediction of Potential Distribution Patterns of Three Larix Species on Qinghai-Tibet Plateau under Future Climate Scenarios. Forests 2023, 14, 1058. [Google Scholar] [CrossRef]
- Cao, B.; Bai, C.; Zhang, L.; Li, G.; Mao, M. Modeling habitat distribution of Cornus officinalis with Maxent modeling and fuzzy logics in China. J. Plant Ecol. 2016, 9, 742–751. [Google Scholar] [CrossRef]
- Kumar, D.; Aggarwal, A.K.; Kumar, R. The effect of interrupted 5-day training on Integrated Management of Neonatal and Childhood Illness on the knowledge and skills of primary health care workers. Health Policy Plan. 2009, 24, 94–100. [Google Scholar] [CrossRef]
- Padalia, H.; Srivastava, V.; Kushwaha, S.P.S. Modeling potential invasion range of alien invasive species, Hyptis suaveolens (L.) Poit. in India: Comparison of MaxEnt and GARP. Ecol. Inform. 2014, 22, 36–43. [Google Scholar] [CrossRef]
- Rojas Briceño, N.B.; Cotrina Sánchez, D.A.; Barboza Castillo, E.; Barrena Gurbillón, M.Á.; Sarmiento, F.O.; Sotomayor, D.A.; Oliva, M.; Salas López, R. Current and Future Distribution of Five Timber Forest Species in Amazonas, Northeast Peru: Contributions towards a Restoration Strategy. Diversity 2020, 12, 305. [Google Scholar] [CrossRef]
- Rathore, P.; Roy, A.; Karnatak, H. Modelling the vulnerability of Taxus wallichiana to climate change scenarios in South East Asia. Ecol. Indic. 2019, 102, 199–207. [Google Scholar] [CrossRef]
- Gao, M.; Sa, R.; Tie, N.; Zhang, L. Distrribution Pattern of Betula platyphlla in China under Climate Change. J. Northeast. For. Univ. 2023, 51, 54–61+69. [Google Scholar] [CrossRef]
- Fand, B.B.; Kumar, M.; Kamble, A.L. Predicting the potential geographic distribution of cotton mealybug Phenacoccus solenopsis in India based on MAXENT ecological niche Model. J. Environ. Biol. 2014, 35, 973–982. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Li, M. Predicting the Potential Distribution of Oxalis debilis Kunth, an Invasive Species in China with a Maximum Entropy Model. Plants 2023, 12, 3999. [Google Scholar] [CrossRef]
- Jenks, G.F. The Data Model Concept in Statistical Mapping. Int. Yearb. Cartogr. 1967, 7, 186–190. [Google Scholar]
- Rong, Z.L.; Zhao, C.Y.; Liu, J.J.; Gao, Y.F.; Zang, F.; Guo, Z.X.; Mao, Y.H.; Wang, L. Modeling the Effect of Climate Change on the Potential Distribution of Qinghai Spruce (Picea crassifolia Kom.) in Qilian Mountains. Forests 2019, 10, 62. [Google Scholar] [CrossRef]
- Deng, F.; LI, X.B.; Wang, H.; Zhang, M.; Li, X.; Li, R.-H. The suitability of geographic distribution and the dominant factors of alfalfa based on MaxEnt model in Xilin Gol. Pratacultural Sci. 2014, 31, 1840–1847. [Google Scholar] [CrossRef]
- Zhang, Y.-B.; Liu, Y.-L.; Qin, H.; Meng, Q.-X. Prediction on spatial migration of suitable distribution of Elaeagnus mollis under climate change conditions in Shanxi Province, China. Chin. J. Appl. Ecol. 2019, 30, 496–502. [Google Scholar] [CrossRef]
- Wisz, M.S.; Pottier, J.; Kissling, W.D.; Pellissier, L.; Lenoir, J.; Damgaard, C.F.; Dormann, C.F.; Forchhammer, M.C.; Grytnes, J.A.; Guisan, A.; et al. The role of biotic interactions in shaping distributions and realised assemblages of species: Implications for species distribution modelling. Biol. Rev. 2013, 88, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Yao, L.; Meng, J.; Tao, J. Maxent modeling for predicting the potential geographical distribution of two peony species under climate change. Sci. Total Environ. 2018, 634, 1326–1334. [Google Scholar] [CrossRef]
- LY/T 1721-2008; Specifications for Assessment of Forest Ecosystem Services in China. Ecology and Nature Conservation Institute: Beijing, China, 2008.
- Zhou, Z.-J.; Zhao, Q.-J. Research on Natural Capital Accounting Method––Based on the Perspective of National Park. China For. Econ. 2020, 2, 6–10. [Google Scholar] [CrossRef]
- Cao, J. Research on Pricing the Value of Ecosystem Products and Services. Ecol. Econ. 2016, 32, 24–28. [Google Scholar] [CrossRef]
- Huang, M.; Shao, M.; Zhang, L.; Li, Y. Water use efficiency and sustainability of different long-term crop rotation systems in the Loess Plateau of China. Soil. Tillage Res. 2003, 72, 95–104. [Google Scholar] [CrossRef]
- Rabotyagov, S.S.; Tóth, S.F.; Ettl, G.J. Testing the Design Variables of ECOSEL: A Market Mechanism for Forest Ecosystem Services. For. Sci. 2013, 59, 303–321. [Google Scholar] [CrossRef]
- Wang, S.-m.; Tan, R.; Wu, L.-m. Assessing The Amenity Value of Agricultural Land: A Case Study in Jiangsu Province. Resour. Environ. Yangtze Basin 2005, 14, 720–724. [Google Scholar] [CrossRef]
- Hua, Y.-J.; Zhang, Z.-H.; Yong, S.-H. The Economic Analysis of Ecological Resource in Project Appraisal. Commer. Res. 2006, 10, 27–29+38. [Google Scholar] [CrossRef]
- Cong, M.; Li, Y.; Yang, W. Potential Distribution of Bryophyte, Entodon challengeri (Entodontaceae), under Climate Warming in China. Diversity 2023, 15, 871. [Google Scholar] [CrossRef]
- Zhou, X.; Lin, J.; Chen, T.; Yang, Y.; Zhang, M.; Li, X. Ecological suitability analysis of Tinospora sagittata. West. China J. Pharm. Sci. 2021, 36, 709–713. [Google Scholar] [CrossRef]
- Harrison, S.; Damschen, E.I.; Grace, J.B. Ecological contingency in the effects of climatic warming on forest herb communities. Proc. Natl. Acad. Sci. USA 2010, 107, 19362–19367. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, X.-P.; Zhang, Y.-F.; Hu, R.; Pan, Y.-X.; Chen, N. Responses of plant growth of different life forms to rainfall amount changes in an arid desert area. Chin. J. Ecol. 2015, 34, 1847–1853. [Google Scholar] [CrossRef]
- Ma, B.; Sun, J. Predicting the distribution of Stipa purpurea across the Tibetan Plateau via the MaxEnt model. BMC Ecol. 2018, 18, 10. [Google Scholar] [CrossRef] [PubMed]
- Diamond, S.E.; Frame, A.M.; Martin, R.A.; Buckley, L.B. Species’ traits predict phenological responses to climate change in butterflies. Ecology 2011, 92, 1005–1012. [Google Scholar] [CrossRef]
- Thuiller, W.; Lavorel, S.; Araújo, M.B. Niche properties and geographical extent as predictors of species sensitivity to climate change. Glob. Ecol. Biogeogr. 2005, 14, 347–357. [Google Scholar] [CrossRef]
- Wu, Z.; Zhang, H.; Krause, C.M.; Cobb, N.S. Climate change and human activities: A case study in Xinjiang, China. Clim. Chang. 2010, 99, 457–472. [Google Scholar] [CrossRef]
- Xu, C.; Chen, Y.; Yang, Y.; Hao, X.; Shen, Y. Hydrology and water resources variation and its response to regional climate change in Xinjiang. J. Geogr. Sci. 2010, 20, 599–612. [Google Scholar] [CrossRef]
- Guoyan, W.; Baskin, C.C.; Baskin, J.M.; Xuejun, Y.; Guofang, L.; Xuehua, Y.; Xinshi, Z.; Zhenying, H. Effects of climate warming and prolonged snow cover on phenology of the early life history stages of four alpine herbs on the southeastern Tibetan Plateau. Am. J. Bot. 2018, 105, 967–976. [Google Scholar] [CrossRef]
- Li, Y.C.; Li, M.Y.; Li, C.; Liu, Z.Z. Optimized Maxent Model Predictions of Climate Change Impacts on the Suitable Distribution of Cunninghamia lanceolata in China. Forests 2020, 11, 302. [Google Scholar] [CrossRef]
- Fang, J.; Zhu, J.; Yue, S. The responses of ecosystems to global warming. Chin. Sci. Bull. 2017, 63, 136–140. [Google Scholar] [CrossRef]
- Sekercioglu, C.H.; Schneider, S.H.; Fay, J.P.; Loarie, S.R. Climate change, elevational range shifts, and bird extinctions. Conserv. Biol. 2008, 22, 140–150. [Google Scholar] [CrossRef]
- Wang, J.R.; Hawkins, C.D.B.; Letchford, T. Photosynthesis, water and nitrogen use efficiencies of four paper birch (Betula papyrifera) populations grown under different soil moisture and nutrient regimes. For. Ecol. Manag. 1998, 112, 233–244. [Google Scholar] [CrossRef]
- Wu, Z.; Raven, P. Flora of China; Flora of China: Beijing, China, 1994. [Google Scholar]
- Dongliang, L.; Xiaolu, C.; Xuan, C.; Jun, Y.; Daojun, Z.; Yong, Y.; Ruikan, F. The survey and application research on lianas in Hainan Island, China. In Proceedings of the 2016 International Conference on Smart Grid and Electrical Automation (ICSGEA), Zhangjiajie, China, 11–12 August 2016; pp. 347–350. [Google Scholar]
- Liu, S.L.; Dong, Y.H.; Liu, H.; Wang, F.F.; Yu, L. Review of Valuation of Forest Ecosystem Services and Realization Approaches in China. Land. 2023, 12, 1102. [Google Scholar] [CrossRef]
- Xu, K.; Yang, Z.S. Research on the Value of Land Ecological Service in Yunnan Province Based on the Perspective of Spatial Pattern. Sustainability 2022, 14, 805. [Google Scholar] [CrossRef]
- Venegas-González, A.; Brancalion, P.H.S.; Albiero, A.; Chagas, M.P.; Anholetto, C.R.; Chaix, G.; Tomazello, M. What tree rings can tell us about the competition between trees and lianas? A case study based on growth, anatomy, density, and carbon accumulation. Dendrochronologia 2017, 42, 1–11. [Google Scholar] [CrossRef]
- Pei, S.; Xie, G.; Liu, C.; Zhang, C.; Li, S.; Chen, L. Dynamic Changes of Water Conservation Service of Typical Ecosystems in China within a Year Based on Data from CERN. Sustainability 2015, 7, 16513–16531. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
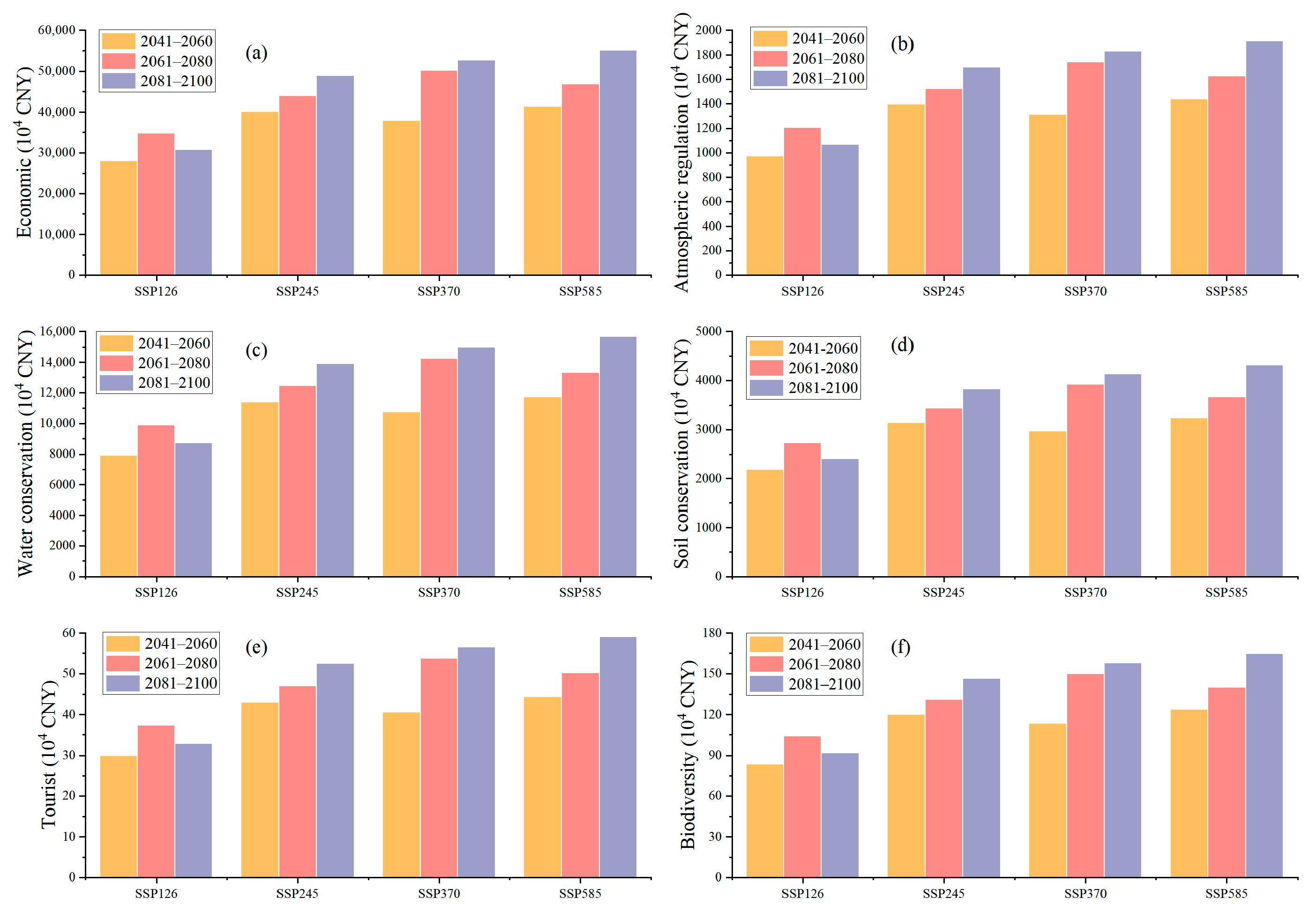{kind=link}
| Category | Variable | Description | Unit | Species 1 |
|---|---|---|---|---|
| Climate | Bio1 | Annual mean temperature | °C | b; c |
| Bio2 | Mean diurnal temperature range | °C | a; b | |
| Bio3 | Isothermality | °C | c | |
| Bio4 | Temperature seasonality | °C | a | |
| Bio6 | Min temperature of coldest month | °C | a | |
| Bio7 | Temperature annual range | °C | a; b; c | |
| Bio8 | Mean temperature of wettest quarter | °C | ||
| Bio9 | Mean temperature of driest quarter | °C | ||
| Bio10 | Mean temperature of warmest quarter | °C | ||
| Bio11 | Mean temperature of coldest quarter | °C | b | |
| Bio12 | Annual precipitation | mm | c | |
| Bio13 | Precipitation of wettest month | mm | c | |
| Bio14 | Precipitation of driest month | mm | ||
| Bio15 | Precipitation seasonality | % | a | |
| Bio16 | Precipitation of wettest quarter | mm | c | |
| Bio17 | Precipitation of driest quarter | mm | c | |
| Bio18 | Precipitation of warmest quarter | mm | ||
| Bio19 | Precipitation of coldest quarter | mm | a; b | |
| Topography | ASP | Aspect | degree | |
| SLO | Slope | degree | a; c | |
| ELE | Elevation | m | ||
| Soil | CEC | Cation exchange capacity of the soil | mmol/kg | a |
| SOC | Soil organic carbon content in the fine earth fraction | dg/kg | b | |
| OCS | Organic carbon stocks | t/ha | a | |
| CLAY | Proportion of clay particles | g/kg | b | |
| SAND | Proportion of sand particles | g/kg | b | |
| pH.H2O | Soil pH | pH × 10 | a; b; c | |
| BDOD | Bulk density of the fine earth fraction | cg/cm3 | ||
| CFVO | Volumetric fraction of coarse fragments (>2 mm) | cm3/dm3 (vol‰) | ||
| NITROGEN | Total nitrogen (N) | cg/kg | ||
| SILT | Proportion of silt particles (≥0.002 mm and ≤0.05 mm) in the fine earth fraction | g/kg | ||
| OCD | Organic carbon density | hg/dm3 |
| Species | Highly Suitable | Moderately Suitable | Marginally Suitable | Total Suitable | Unsuitable | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Area | Ratio | Area | Ratio | Area | Ratio | Area | Ratio | Area | Ratio | |
| T. craveniana | 11.90 | 1.24 | 19.78 | 2.06 | 53.66 | 5.59 | 85.34 | 8.89 | 874.66 | 91.11 |
| T. yunnanensis | 12.86 | 1.34 | 17.76 | 1.85 | 45.02 | 4.69 | 75.64 | 7.88 | 884.36 | 92.12 |
| T. sinensis | 10.18 | 1.06 | 18.43 | 1.92 | 51.65 | 5.38 | 80.26 | 8.36 | 879.74 | 91.64 |
| Category | Variable | Percent Contribution/% | ||
|---|---|---|---|---|
| T. craveniana | T. yunnanensis | T. sinensis | ||
| Climate | Bio1 | - | 6.3 | 47.8 |
| Bio2 | 32.1 | 24 | - | |
| Bio3 | - | - | 4 | |
| Bio4 | 0.4 | - | - | |
| Bio6 | 31.5 | - | - | |
| Bio7 | 1.3 | 13.9 | 2 | |
| Bio11 | - | 15.6 | - | |
| Bio13 | - | - | 0.7 | |
| Bio14 | - | - | 4.3 | |
| Bio15 | 1.1 | - | - | |
| Bio16 | - | - | 15.9 | |
| Bio17 | 15 | |||
| Bio19 | 9.1 | 6.3 | - | |
| Topography | SLO | 0.5 | - | 3.8 |
| Soil | CEC | 16.6 | - | - |
| SOC | - | 7.2 | - | |
| OCS | 0.6 | - | - | |
| CLAY | - | 8.6 | - | |
| SAND | - | 3.7 | - | |
| pH.H2O | 3.6 | 5.3 | 6.5 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Li, Z.; Zou, H.; Wang, Z.; Zhu, X.; Zhang, Y.; Liu, Z. Global Warming Drives Transitions in Suitable Habitats and Ecological Services of Rare Tinospora Miers Species in China. Diversity 2024, 16, 181. https://doi.org/10.3390/d16030181
Zhang H, Li Z, Zou H, Wang Z, Zhu X, Zhang Y, Liu Z. Global Warming Drives Transitions in Suitable Habitats and Ecological Services of Rare Tinospora Miers Species in China. Diversity. 2024; 16(3):181. https://doi.org/10.3390/d16030181
Chicago/Turabian StyleZhang, Huayong, Zhe Li, Hengchao Zou, Zhongyu Wang, Xinyu Zhu, Yihe Zhang, and Zhao Liu. 2024. "Global Warming Drives Transitions in Suitable Habitats and Ecological Services of Rare Tinospora Miers Species in China" Diversity 16, no. 3: 181. https://doi.org/10.3390/d16030181