
Analysis of microRNA Expression Profiles in Broiler Muscle Tissues by Feeding Different Levels of Guanidinoacetic Acid

Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Feeding Management and Sample Collection
2.3. RNA Isolation, Construction and Sequencing of Small RNA Libraries
2.4. Quality Control of Small RNA Sequencing
2.5. Differential Expression Analysis and RT-qPCR Validation
2.6. Differentially Expressed miRNA Target Gene Prediction, GO and KEGG Enrichment Analysis
3. Results
3.1. Quality Control of Small RNA Sequencing Data
3.2. Genome Comparison and Classification Notes
3.3. Screening and RT-qPCR Validation of Differentially Expressed miRNAs
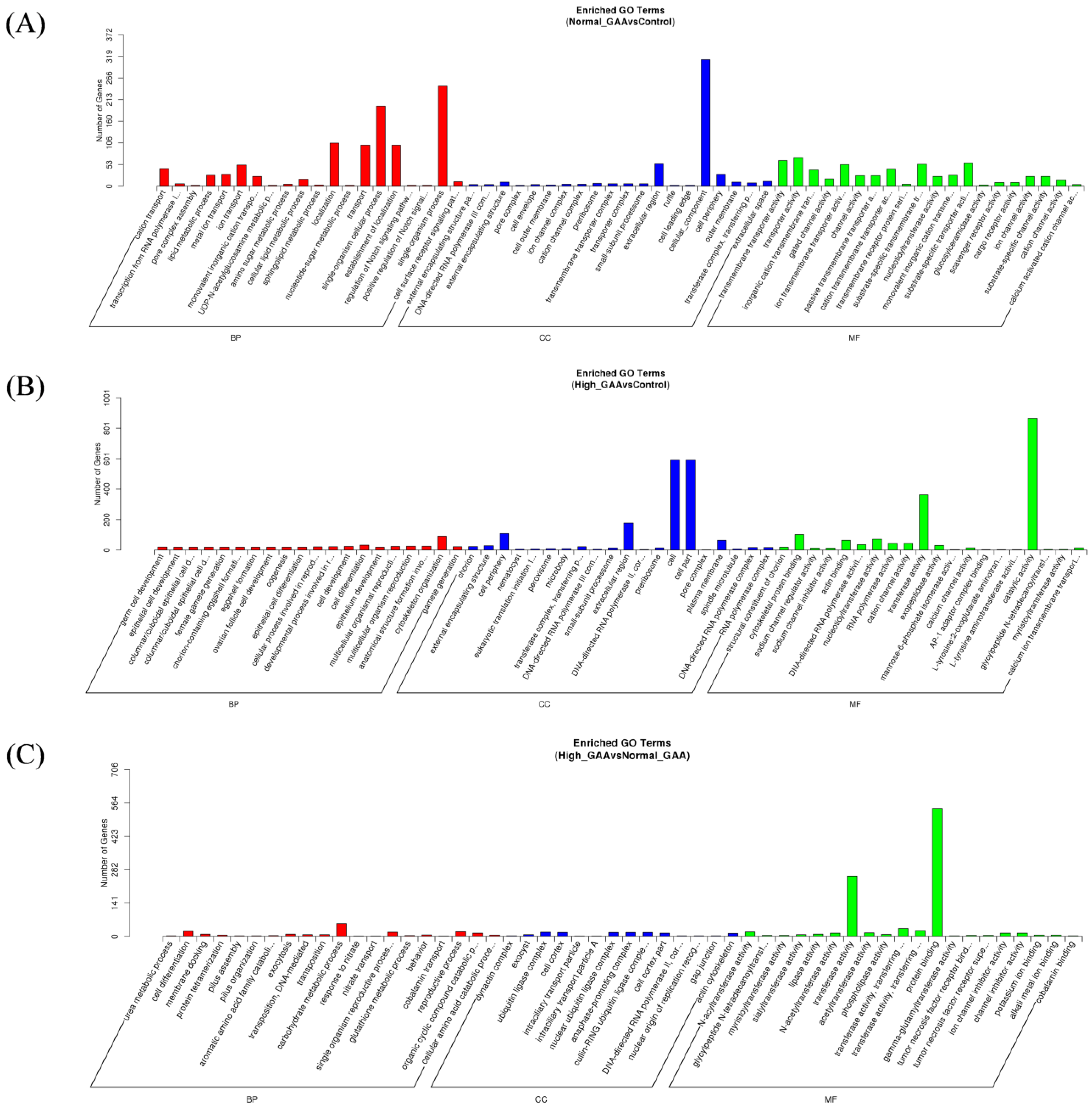
3.4. Prediction and Bioinformatics Analysis of Differentially Expressed miRNA Target Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Total_Reads | N% > 10% | Low Quality | 5_Adapter_Contamine | 3_Adapter_Null or Insert_Null | with ploya/T/G/C | Clean Reads |
|---|---|---|---|---|---|---|---|
| NC_1 | 9,875,151 (100.00%) | 394 (0.00%) | 18,995 (0.19%) | 105,833 (1.07%) | 312,038 (3.16%) | 13,139 (0.13%) | 9,424,752 (95.44%) |
| NC_2 | 14,441,618 (100.00%) | 365 (0.00%) | 38,177 (0.26%) | 31,738 (0.22%) | 651,214 (4.51%) | 52,777 (0.37%) | 13,667,347 (94.64%) |
| NC_3 | 15,600,912 (100.00%) | 360 (0.00%) | 33,805 (0.22%) | 75,301 (0.48%) | 228,177 (1.46%) | 24,754 (0.16%) | 15,238,515 (97.68%) |
| Normal_GAA_1 | 15,352,341 (100.00%) | 383 (0.00%) | 31,269 (0.20%) | 85,795 (0.56%) | 252,960 (1.65%) | 19,553 (0.13%) | 14,962,381 (97.46%) |
| Normal_GAA_2 | 15,560,784 (100.00%) | 542 (0.00%) | 33,473 (0.22%) | 35,518 (0.23%) | 434,904 (2.79%) | 16,993 (0.11%) | 15,039,354 (96.65%) |
| Normal_GAA_3 | 15,535,764 (100.00%) | 479 (0.00%) | 41,092 (0.26%) | 53,305 (0.34%) | 449,571 (2.89%) | 17,377 (0.11%) | 14,973,940 (96.38%) |
| High_GAA_1 | 15,287,814 (100.00%) | 373 (0.00%) | 38,844 (0.25%) | 39,613 (0.26%) | 167,238 (1.09%) | 27,765 (0.18%) | 15,013,981 (98.21%) |
| High_GAA_2 | 15,166,518 (100.00%) | 381 (0.00%) | 31,076 (0.20%) | 31,112 (0.21%) | 143,446 (0.95%) | 17,570 (0.12%) | 14,942,933 (98.53%) |
| High_GAA_3 | 15,296,267 (100.00%) | 345 (0.00%) | 30,692 (0.20%) | 32,201 (0.21%) | 183,827 (1.20%) | 19,186 (0.13%) | 15,030,016 (98.26%) |
| Sample | Reads | Bases | Error Rate | Q20 | Q30 | GC Content |
|---|---|---|---|---|---|---|
| NC_1 | 9,875,151 | 0.494 G | 0.01% | 99.57% | 98.26% | 48.60% |
| NC_2 | 14,441,618 | 0.722 G | 0.01% | 99.37% | 97.71% | 52.39% |
| NC_3 | 15,600,912 | 0.780 G | 0.01% | 99.54% | 98.21% | 46.58% |
| Normal_GAA_1 | 15,352,341 | 0.768 G | 0.01% | 99.55% | 98.30% | 46.04% |
| Normal_GAA_2 | 15,560,784 | 0.778 G | 0.01% | 99.53% | 98.07% | 45.44% |
| Normal_GAA_3 | 15,535,764 | 0.777 G | 0.01% | 99.54% | 98.19% | 45.46% |
| High_GAA_1 | 15,287,814 | 0.764 G | 0.01% | 99.47% | 97.71% | 45.92% |
| High_GAA_2 | 15,166,518 | 0.758 G | 0.01% | 99.53% | 98.27% | 45.39% |
| High_GAA_3 | 15,296,267 | 0.765 G | 0.01% | 99.59% | 98.43% | 45.36% |
| Sample | Total Reads | Total Bases (bp) | Uniq Reads | Uniq Bases (bp) |
|---|---|---|---|---|
| NC_1 | 6,294,787 | 144,153,981 | 292,159 | 6,966,123 |
| NC_2 | 8,083,155 | 171,818,221 | 358,265 | 7,735,712 |
| NC_3 | 14,262,690 | 324,483,031 | 358,329 | 8,775,554 |
| Normal_GAA_1 | 13,645,431 | 304,579,166 | 318,234 | 7,546,472 |
| Normal_GAA_2 | 13,575,735 | 301,417,656 | 362,851 | 8,403,654 |
| Normal_GAA_3 | 13,806,341 | 306,305,999 | 245,337 | 5,756,793 |
| High_GAA_1 | 14,263,796 | 320,833,708 | 453,565 | 10,991,480 |
| High_GAA_2 | 14,496,158 | 325,818,359 | 448,438 | 10,970,521 |
| High_GAA_3 | 14,185,836 | 315,122,981 | 342,272 | 8,174,591 |
References
- Yan, Z.; Yan, Z.; Liu, S.; Yin, Y.; Yang, T.; Chen, Q. Regulative Mechanism of Guanidinoacetic Acid on Skeletal Muscle Development and Its Application Prospects in Animal Husbandry: A Review. Front. Nutr. 2021, 8, 714567. [Google Scholar] [CrossRef] [PubMed]
- Faraji, M.; Dehkordi, S.K.; Moghadam, A.K.Z.; Ahmadipour, B.; Khajali, F. Combined effects of guanidinoacetic acid, coenzyme Q10 and taurine on growth performance, gene expression and ascites mortality in broiler chickens. J. Anim. Physiol. Anim. Nutr. 2018, 103, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Ahmadipour, B.; Naeini, S.Z.; Sharifi, M.; Khajali, F. Growth performance and right ventricular hypertrophy responses of broiler chickens to guanidinoacetic acid supplementation under hypobaric hypoxia. J. Poult. Sci. 2018, 55, 60–64. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.H.; Rahimnejad, S.; Wang, L.; Song, K.; Lu, K.; Zhang, C.X. Effects of guanidinoacetic acid supplementation in all-plant protein diets on growth, antioxidant capacity and muscle energy metabolism of bullfrog Rana (Lithobates) catesbeiana. Aquac. Res. 2018, 49, 748–756. [Google Scholar] [CrossRef]
- Tabatabaei Yazdi, F.; Golian, A.; Zarghi, H.; Varidi, M. Effect of wheat-soy diet nutrient density and guanidine acetic acid supplementation on performance and energy metabolism in broiler chickens. Ital. J. Anim. Sci. 2017, 16, 593–600. [Google Scholar] [CrossRef]
- Wyss, M.; Kaddurah-Daouk, R. Creatine and creatinine metabolism. Physiol. Rev. 2000, 80, 1107–1213. [Google Scholar] [CrossRef] [PubMed]
- Wallimann, T. Introduction–creatine: Cheap ergogenic supplement with great potential for health and disease. In Creatine and Creatine Kinase in Health and Disease; Springer: Berlin/Heidelberg, Germany, 2007; pp. 1–16. [Google Scholar]
- Balestrino, M.; Adriano, E. Presence of guanidinoacetate may compensate creatine absence and account for less statin-induced muscle damage in GAMT-deficient compared to AGAT-deficient mice. Amino Acids 2020, 52, 667–669. [Google Scholar] [CrossRef] [PubMed]
- Lygate, C.A.; Aksentijevic, D.; Dawson, D.; Ten Hove, M.; Phillips, D.; de Bono, J.P.; Medway, D.J.; Sebag-Montefiore, L.; Hunyor, I.; Channon, K.M.; et al. Living without creatine: Unchanged exercise capacity and response to chronic myocardial infarction in creatine-deficient mice. Circ. Res. 2013, 112, 945–955. [Google Scholar] [CrossRef]
- Ahmadipour, B.; Khajali, F.; Sharifi, M. Effect of guanidinoacetic acid supplementation on growth performance and gut morphology in broiler chickens. Poult. Sci. J. 2018, 6, 19–24. [Google Scholar]
- Amiri, M.; Ghasemi, H.A.; Hajkhodadadi, I.; Farahani, A.H.K. Efficacy of guanidinoacetic acid at different dietary crude protein levels on growth performance, stress indicators, antioxidant status, and intestinal morphology in broiler chickens subjected to cyclic heat stress. Anim. Feed Sci. Technol. 2019, 254, 114208. [Google Scholar] [CrossRef]
- Wong, R.R.; Abd-Aziz, N.; Affendi, S.; Poh, C.L. Role of microRNAs in antiviral responses to dengue infection. J. Biomed. Sci. 2020, 27, 4. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Guo, M.; Hay, B.A. MicroRNAs and the regulation of cell death. Trends Genet. 2004, 20, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Cheng, A.M.; Byrom, M.W.; Shelton, J.; Ford, L.P. Antisense inhibition of human miRNAs and indications for an involvement of miRNA in cell growth and apoptosis. Nucleic Acids Res. 2005, 33, 1290–1297. [Google Scholar] [CrossRef] [PubMed]
- O’Rourke, J.R.; Georges, S.A.; Seay, H.R.; Tapscott, S.J.; McManus, M.T.; Goldhamer, D.J.; Swanson, M.S.; Harfe, B.D. Essential role for Dicer during skeletal muscle development. Dev. Biol. 2007, 311, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-F.; Mandel, E.M.; Thomson, J.M.; Wu, Q.; E Callis, T.; Hammond, S.M.; Conlon, F.L.; Wang, D.-Z. The role of microRNA-1 and microRNA-133 in skeletal muscle proliferation and differentiation. Nat. Genet. 2006, 38, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Nakasa, T.; Ishikawa, M.; Shi, M.; Shibuya, H.; Adachi, N.; Ochi, M. Acceleration of muscle regeneration by local injection of muscle-specific microRNAs in rat skeletal muscle injury model. J. Cell. Mol. Med. 2010, 14, 2495–2505. [Google Scholar] [CrossRef] [PubMed]
- Dey, B.K.; Gagan, J.; Dutta, A. miR-206 and-486 induce myoblast differentiation by downregulating Pax7. Mol. Cell. Biol. 2011, 31, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Lee, Y.S.; Sivaprasad, U.; Malhotra, A.; Dutta, A. Muscle-specific microRNA miR-206 promotes muscle differentiation. J. Cell Biol. 2006, 174, 677–687. [Google Scholar] [CrossRef]
- Dong, X.; Cheng, Y.; Qiao, L.; Wang, X.; Zeng, C.; Feng, Y. Male-biased gga-miR-2954 regulates myoblast proliferation and differentiation of chicken embryos by targeting YY1. Genes 2021, 12, 1325. [Google Scholar] [CrossRef]
- Jia, X.; Ouyang, H.; Abdalla, B.A.; Xu, H.; Nie, Q.; Zhang, X. miR-16 controls myoblast proliferation and apoptosis through directly suppressing Bcl2 and FOXO1 activities. Biochim. Et Biophys. Acta-Gene Regul. Mech. 2017, 1860, 674–684. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Chen, F.; He, M.; Wu, P.; Zhou, K.; Zhang, T.; Chu, M.; Zhang, G. miR-7 regulates the apoptosis of chicken primary myoblasts through the KLF4 gene. Br. Poult. Sci. 2022, 63, 39–45. [Google Scholar] [CrossRef] [PubMed]
- GB/T 35892-2018; General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China (AQSIQ). Laboratory Animal-Guideline for Ethical Review of Animal Welfare. Standards Press of China: Beijing, China, 2018.
- Wen, M.; Shen, Y.; Shi, S.; Tang, T. miREvo: An integrative microRNA evolutionary analysis platform for next-generation sequencing experiments. BMC Bioinform. 2012, 13, 140. [Google Scholar] [CrossRef] [PubMed]
- Friedländer, M.R.; Mackowiak, S.D.; Li, N.; Chen, W.; Rajewsky, N. miRDeep2 accurately identifies known and hundreds of novel microRNA genes in seven animal clades. Nucleic Acids Res. 2012, 40, 37–52. [Google Scholar] [CrossRef]
- Wang, Y.; Ma, J.; Qiu, W.; Zhang, J.; Feng, S.; Zhou, X.; Wang, X.; Jin, L.; Long, K.; Liu, L.; et al. Guanidinoacetic acid regulates myogenic differentiation and muscle growth through miR-133a-3p and miR-1a-3p co-mediated Akt/mTOR/S6K signaling pathway. Int. J. Mol. Sci. 2018, 19, 2837. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zou, T.; Wang, Z.; Yang, J.; Li, L.; Guo, X.; He, Q.; Chen, L.; You, J. Dietary guanidinoacetic acid improves the growth performance and skeletal muscle development of finishing pigs through changing myogenic gene expression and myofibre characteristics. J. Anim. Physiol. Anim. Nutr. 2020, 104, 1875–1883. [Google Scholar] [CrossRef]
- Michiels, J.; Maertens, L.; Buyse, J.; Lemme, A.; Rademacher, M.; Dierick, N.; De Smet, S. Supplementation of guanidinoacetic acid to broiler diets: Effects on performance, carcass characteristics, meat quality, and energy metabolism. Poult. Sci. 2012, 91, 402–412. [Google Scholar] [CrossRef]
- Lemme, A.; Ringel, J.; Rostagno, H.S.; Redshaw, M.S. Supplemental guanidino acetic acid improved feed conversion, weight gain, and breast meat yield in male and female broilers. In Proceedings of the 16th European Symposium on Poultry Nutrition, Strasbourg, France, 26–30 August 2007. [Google Scholar]
- Ringel, J.; Lemme, A.; Redshaw, M.S.; Damme, K. The effects of supplemental guanidino acetic acid as a precursor of creatine in vegetable broiler diets on performance and carcass parameters. In Poultry Science; Poultry Science Assoc Inc.: Savoy, IL, USA, 2008; Volume 87, p. 72. [Google Scholar]
- Li, S.Y.; Wang, C.; Wu, Z.Z.; Liu, Q.; Guo, G.; Huo, W.J.; Zhang, J.; Chen, L.; Zhang, Y.L.; Pei, C.X.; et al. Effects of guanidinoacetic acid supplementation on growth performance, nutrient digestion, rumen fermentation and blood metabolites in Angus bulls. Animal 2020, 14, 2535–2542. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Wang, C.; Zhang, J.; Liu, Q.; Guo, G.; Huo, W.J.; Pei, C.X.; Chen, L.; Zhang, Y.L. Guanidinoacetic acid and betaine supplementation have positive effects on growth performance, nutrient digestion and rumen fermentation in Angus bulls. Anim. Feed Sci. Technol. 2021, 276, 114923. [Google Scholar] [CrossRef]
- Chao, Y.; Liu, B.; Kou, Q.; Niu, W.; Jiang, H.; Yang, L.; Zhang, G. Effects of guanidine acetic acid on growth performance, slaughter performance, fat deposition and nutritional components in muscle of stabling Tan sheep. Chin. J. Anim. Nutr. 2019, 31, 388–394. [Google Scholar]
- Li, W.-J.; Wu, Q.-C.; Cui, Z.-Y.; Jiang, Y.-W.; Aisikaer, A.; Zhang, F.; Chen, H.-W.; Wang, W.-K.; Wang, Y.-L.; Lv, L.-K.; et al. Guanidine acetic acid exhibited greater growth performance in younger (13–30 kg) than in older (30–50 kg) lambs under high-concentrate feedlotting pattern. Front. Vet. Sci. 2022, 9, 954675. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Li, M.; Huang, Y.; Duan, Y.; Jia, J.; Ge, C.; Cao, W. Effects of Guanidinoacetic Acid on Growth Performance, Slaughter Performance, Immune Organ Index and Antioxidant Capacity of Broilers. China Poult. 2023, 45, 21–26. [Google Scholar]
- Zhang, L.; Liu, J.; Cheng, P.; Lv, F. Correlation between miRNA target site polymorphisms in the 3′ UTR of AVPR1A and the risk of hypertension in the Chinese Han population. Biosci. Rep. 2019, 39, BSR20182232. [Google Scholar] [CrossRef] [PubMed]
- Pinzón, N.; Li, B.; Martinez, L.; Sergeeva, A.; Presumey, J.; Apparailly, F.; Seitz, H. microRNA target prediction programs predict many false positives. Genome Res. 2017, 27, 234–245. [Google Scholar] [CrossRef] [PubMed]
- Motohashi, N.; Alexander, M.S.; Shimizu-Motohashi, Y.; Myers, J.A.; Kawahara, G.; Kunkel, L.M. Regulation of IRS1/Akt insulin signaling by microRNA-128a during myogenesis. J. Cell Sci. 2013, 126, 2678–2691. [Google Scholar] [CrossRef]
- Alexander, M.S.; Kawahara, G.; Motohashi, N.; Casar, J.C.; Eisenberg, I.; A Myers, J.; Gasperini, M.J.; A Estrella, E.; Kho, A.T.; Mitsuhashi, S.; et al. MicroRNA-199a is induced in dystrophic muscle and affects WNT signaling, cell proliferation, and myogenic differentiation. Cell Death Differ. 2013, 20, 1194–1208. [Google Scholar] [CrossRef]
- Cardinali, B.; Castellani, L.; Fasanaro, P.; Basso, A.; Alema, S.; Martelli, F.; Falcone, G. Microrna-221 and microrna-222 modulate differentiation and maturation of skeletal muscle cells. PLoS ONE 2009, 4, e7607. [Google Scholar] [CrossRef]
- Song, C.; Yang, J.; Jiang, R.; Yang, Z.; Li, H.; Huang, Y.; Lan, X.; Lei, C.; Ma, Y.; Qi, X.; et al. miR-148a-3p regulates proliferation and apoptosis of bovine muscle cells by targeting KLF6. J. Cell. Physiol. 2019, 234, 15742–15750. [Google Scholar] [CrossRef] [PubMed]
- Feng, K.; Wu, Y.; Li, J.; Sun, Q.; Ye, Z.; Li, X.; Guo, X.; Kang, J. Critical Role of miR-130b-5p in Cardiomyocyte Proliferation and Cardiac Repair in Mice After Myocardial Infarction. Stem Cells 2024, 42, 29–41. [Google Scholar] [CrossRef]
- Liu, C.; Wang, L.; Chen, W. Preliminary study on miRNA-133a-3p and hypoxia in rat cardiomyocytes. J. Reg. Anat. Oper. Surg. 2016, 25, 784–788. [Google Scholar]
- Robinson, M.J.; Cobb, M.H. Mitogen-activated protein kinase pathways. Curr. Opin. Cell Biol. 1997, 9, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Bennett, A.M.; Tonks, N.K. Regulation of distinct stages of skeletal muscle differentiation by mitogen-activated protein kinases. Science 1997, 278, 1288–1291. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Liu, H.; Wang, M.; Li, R.; Zeng, J.; Mo, D.; Cong, P.; Liu, X.; Chen, Y.; He, Z. Disruption of the ZBED6 binding site in intron 3 of IGF2 by CRISPR/Cas9 leads to enhanced muscle development in Liang Guang Small Spotted pigs. Transgenic Res. 2019, 28, 141–150. [Google Scholar] [CrossRef]
- Dykstra, P.B.; Rando, T.A.; Smolke, C.D. Modulating myoblast differentiation with RNA-based controllers. PLoS ONE 2022, 17, e0275298. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.B.; Ruan, C.C.; Lin, J.R.; Xu, L.; Chen, X.H.; Du, Y.N.; Fu, M.X.; Kong, L.R.; Zhu, D.L.; Gao, P.J. Perivascular adipose tissue–derived PDGF-D contributes to aortic aneurysm formation during obesity. Diabetes 2018, 67, 1549–1560. [Google Scholar] [CrossRef] [PubMed]
- Kurasawa, K.; Arai, S.; Owada, T.; Maezawa, R.; Kumano, K.; Fukuda, T. Autoantibodies against platelet-derived growth factor receptor alpha in patients with systemic lupus erythematosus. Mod. Rheumatol. 2010, 20, 458–465. [Google Scholar] [CrossRef]
- Saravanamuthu, S.S.; Gao, C.Y.; Zelenka, P.S. Notch signaling is required for lateral induction of Jagged1 during FGF-induced lens fiber differentiation. Dev. Biol. 2009, 332, 166–176. [Google Scholar] [CrossRef]





| miRNA Name | Primer | Primer Sequence |
|---|---|---|
| gga-miR-130b-5p | Loop | GTTGGCTCTGGTGCAGGGTCCGAGGTATTCGCACCAGAGCCAACAGTAGT |
| F | TGTGTTTTCCTCTTTCCCTGTTG | |
| R | GTGCAGGGTCCGAGGT | |
| gga-miR-1a-3p | Loop | GTTGGCTCTGGTGCAGGGTCCGAGGTATTCGCACCAGAGCCAACTACATA |
| F | TGTTGTGGGTGGAATGTAAAGAAG | |
| R | GTGCAGGGTCCGAGGT | |
| gga-miR-19a-3p | Loop | GTTGGCTCTGGTGCAGGGTCCGAGGTATTCGCACCAGAGCCAACTCAGTT |
| F | GGTTTTTTTTTTGTGCAAATCTATGCAA | |
| R | GTGCAGGGTCCGAGGT |
| Sample | Total sRNA | Mapped sRNA | +Mapped sRNA | −Mapped sRNA |
|---|---|---|---|---|
| NC_1 | 6,294,787 (100.00%) | 5,835,551 (92.70%) | 3,938,059 (62.56%) | 1,897,492 (30.14%) |
| NC_2 | 8,083,155 (100.00%) | 6,821,769 (84.39%) | 5,655,641 (69.97%) | 1,166,128 (14.43%) |
| NC_3 | 14,262,690 (100.00%) | 135,553,165 (95.03%) | 10,017,664 (70.24%) | 3,535,501 (24.79%) |
| Normal_GAA_1 | 13,645,431 (100.00%) | 12,975,568 (95.09%) | 9,757,778 (71.51%) | 3,217,790 (23.58%) |
| Normal_GAA_2 | 13,575,735 (100.00%) | 12,976,213 (95.58%) | 10,679,295 (78.66%) | 2,296,918 (16.92%) |
| Normal_GAA_3 | 13,806,341 (100.00%) | 13,193,544 (95.56%) | 10,582,156 (76.65%) | 2,611,388 (18.91%) |
| High_GAA_1 | 14,263,796 (100.00%) | 13,609,746 (95.41%) | 11,012,433 (77.21%) | 2,597,313 (18.21%) |
| High_GAA_2 | 14,496,158 (100.00%) | 13,803,787 (95.22%) | 11,036,143 (76.13%) | 2,767,644 (19.09%) |
| High_GAA_3 | 14,185,836 (100.00%) | 13,770,873 (97.07%) | 11,467,097 (80.83%) | 2,303,776 (16.24%) |
| Types | NC_1 | NC_2 | NC_3 | Normal_GAA_1 | Normal_GAA_2 | Normal_GAA_3 | High_GAA_1 | High_GAA_2 | High_GAA_3 |
|---|---|---|---|---|---|---|---|---|---|
| total | 5,835,551 (100.00%) | 6,821,769 (100.00%) | 13,553,165 (100.00%) | 12,975,568 (100.00%) | 12,976,213 (100.00%) | 13,193,544 (100.00%) | 13,609,746 (100.00%) | 13,803,787 (100.00%) | 13,770,873 (100.00%) |
| known_miRNA | 2,520,080 (43.18%) | 2,099,008 (30.77%) | 9,152,611 (67.53%) | 8,735,596 (67.32%) | 10,154,260 (78.25%) | 9,971,723 (75.58%) | 9,975,678 (73.30%) | 10,387,778 (75.25%) | 10,926,503 (79.35%) |
| rRNA | 1,425,155 (24.42%) | 1,898,055 (27.82%) | 1,204,404 (8.89%) | 1,369,240 (10.55%) | 490,065 (3.78%) | 836,735 (6.34%) | 961,446 (7.06%) | 717,176 (5.20%) | 821,914 (5.97%) |
| tRNA | 144,481 (2.48%) | 73,725 (1.08%) | 257,837 (1.90%) | 48,019 (0.37%) | 104,579 (0.81%) | 33,326 (0.25%) | 179,490 (1.32%) | 141,007 (1.02%) | 56,612 (0.41%) |
| snRNA | 1633 (0.03%) | 13,109 (0.19%) | 3240 (0.02%) | 2433 (0.02%) | 2091 (0.02%) | 1459 (0.01%) | 4170 (0.03%) | 3218 (0.02%) | 2712 (0.02%) |
| snoRNA | 20,339 (0.35%) | 78,328 (1.15%) | 49,895 (0.37%) | 27,276 (0.21%) | 30,450 (0.23%) | 18,055 (0.14%) | 52,642 (0.39%) | 51,549 (0.37%) | 33,589 (0.24%) |
| repeat | 26,939 (0.46%) | 12,204 (0.18%) | 17,813 (0.13%) | 18,937 (0.15%) | 13,861 (0.11%) | 8890 (0.07%) | 13,933 (0.10%) | 12,478 (0.09%) | 9691 (0.07%) |
| novel_miRNA | 2385 (0.04%) | 5318 (0.08%) | 7863 (0.06%) | 3611 (0.03%) | 5041 (0.04%) | 3508 (0.03%) | 6634 (0.05%) | 10,407 (0.08%) | 7171 (0.05%) |
| exon | 592,596 (10.15%) | 220,250 (3.23%) | 441,720 (3.26%) | 543,749 (4.19%) | 232,465 (1.79%) | 251,753 (1.91%) | 297,656 (2.19%) | 314,672 (2.28%) | 216,897 (1.58%) |
| intron | 196,410 (3.37%) | 1,492,176 (21.87%) | 471,223 (3.48%) | 419,346 (3.23%) | 281,334 (2.17%) | 285,090 (2.16%) | 348,324 (2.56%) | 243,553 (1.76%) | 235,124 (1.71%) |
| other | 905,533 (15.52%) | 929,596 (13.63%) | 1,946,559 (14.36%) | 1,807,361 (13.93%) | 1,662,067 (12.81%) | 1,783,005 (13.51%) | 1,769,773 (13.00%) | 1,921,949 (13.92%) | 1,460,660 (10.61%) |
| Types | NC_1 | NC_2 | NC_3 | Normal_GAA_1 | Normal_GAA_2 | Normal_GAA_3 | High_GAA_1 | High_GAA_2 | High_GAA_3 |
|---|---|---|---|---|---|---|---|---|---|
| total | 219,401 (100.00%) | 284,669 (100.00%) | 278,624 (100.00%) | 239,364 (100.00%) | 284,463 (100.00%) | 186,060 (100.00%) | 358,971 (100.00%) | 342,354 (100.00%) | 268,206 (100.00%) |
| known_miRNA | 1453 (0.66%) | 2362 (0.83%) | 2304 (0.83%) | 1931 (0.81%) | 2068 (0.73%) | 2050 (1.10%) | 2443 (0.68%) | 2491 (0.73%) | 2411 (0.90%) |
| rRNA | 22,849 (10.41%) | 46,812 (16.44%) | 37,576 (13.49%) | 33,722 (14.09%) | 32,757 (11.52%) | 31,463 (16.91%) | 48,641 (13.55%) | 44,012 (12.86%) | 43,990 (16.40%) |
| tRNA | 1895 (0.86%) | 4136 (1.45%) | 3312 (1.19%) | 2534 (1.06%) | 3012 (1.06%) | 2,308 (1.24%) | 3841 (1.07%) | 4032 (1.18%) | 3053 (1.14%) |
| snRNA | 318 (0.14%) | 1434 (0.50%) | 605 (0.22%) | 482 (0.20%) | 560 (0.20%) | 345 (0.19%) | 910 (0.25%) | 724 (0.21%) | 686 (0.26%) |
| snoRNA | 1240 (0.57%) | 4672 (1.64%) | 2114 (0.76%) | 1612 (0.67%) | 1909 (0.67%) | 1343 (0.72%) | 2803 (0.78%) | 2458 (0.72%) | 2240 (0.84%) |
| repeat | 7059 (3.22%) | 5532 (1.94%) | 6842 (2.46%) | 6287 (2.63%) | 7356 (2.59%) | 4184 (2.25%) | 7813 (2.18%) | 6900 (2.02%) | 5275 (1.97%) |
| novel_miRNA | 75 (0.03%) | 123 (0.04%) | 98 (0.04%) | 82 (0.03%) | 90 (0.03%) | 78 (0.04%) | 114 (0.03%) | 98 (0.03%) | 95 (0.04%) |
| exon | 71,761 (32.71%) | 85,408 (30.00%) | 86,346 (30.99%) | 74,266 (31.03%) | 79,096 (27.81%) | 57,626 (30.97%) | 111,909 (31.17%) | 101,894 (29.76%) | 84,741 (31.60%) |
| intron | 42,222 (19.24%) | 35,702 (12.54%) | 46,795 (16.80%) | 38,953 (16.27%) | 62,547 (21.99%) | 24,670 (13.26%) | 63,943 (17.81%) | 59,326 (17.33%) | 37,840 (14.11%) |
| other | 70,529 (32.15%) | 98,488 (34.60%) | 92,632 (33.25%) | 79,495 (33.21%) | 95,068 (33.42%) | 61,993 (33.32%) | 116,554 (32.47%) | 120,419 (35.17%) | 87,875 (32.76%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, M.; Li, M.; Ruan, J.; Jia, J.; Ge, C.; Cao, W. Analysis of microRNA Expression Profiles in Broiler Muscle Tissues by Feeding Different Levels of Guanidinoacetic Acid. Curr. Issues Mol. Biol. 2024, 46, 3713-3728. https://doi.org/10.3390/cimb46040231
Liu M, Li M, Ruan J, Jia J, Ge C, Cao W. Analysis of microRNA Expression Profiles in Broiler Muscle Tissues by Feeding Different Levels of Guanidinoacetic Acid. Current Issues in Molecular Biology. 2024; 46(4):3713-3728. https://doi.org/10.3390/cimb46040231
Chicago/Turabian StyleLiu, Mengqian, Mengyuan Li, Jinrui Ruan, Junjing Jia, Changrong Ge, and Weina Cao. 2024. "Analysis of microRNA Expression Profiles in Broiler Muscle Tissues by Feeding Different Levels of Guanidinoacetic Acid" Current Issues in Molecular Biology 46, no. 4: 3713-3728. https://doi.org/10.3390/cimb46040231
APA StyleLiu, M., Li, M., Ruan, J., Jia, J., Ge, C., & Cao, W. (2024). Analysis of microRNA Expression Profiles in Broiler Muscle Tissues by Feeding Different Levels of Guanidinoacetic Acid. Current Issues in Molecular Biology, 46(4), 3713-3728. https://doi.org/10.3390/cimb46040231