
Shift from Pro- to Anti-Inflammatory Phase in Pelvic Floor Muscles at Postpartum Matches Histological Signs of Regeneration in Multiparous Rabbits
 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Histology
2.3. Immunohistochemistry
2.4. Statistical Analyses
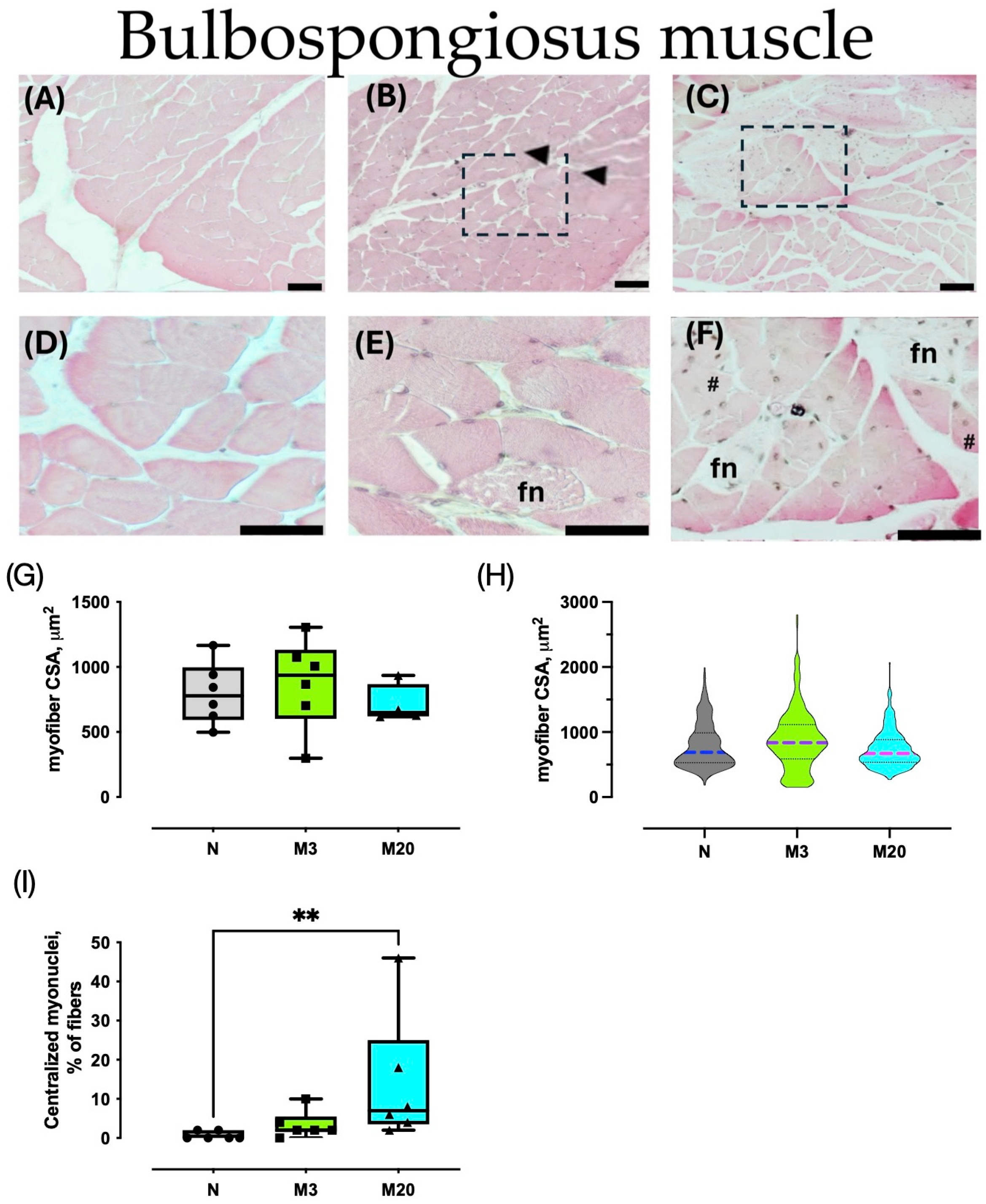
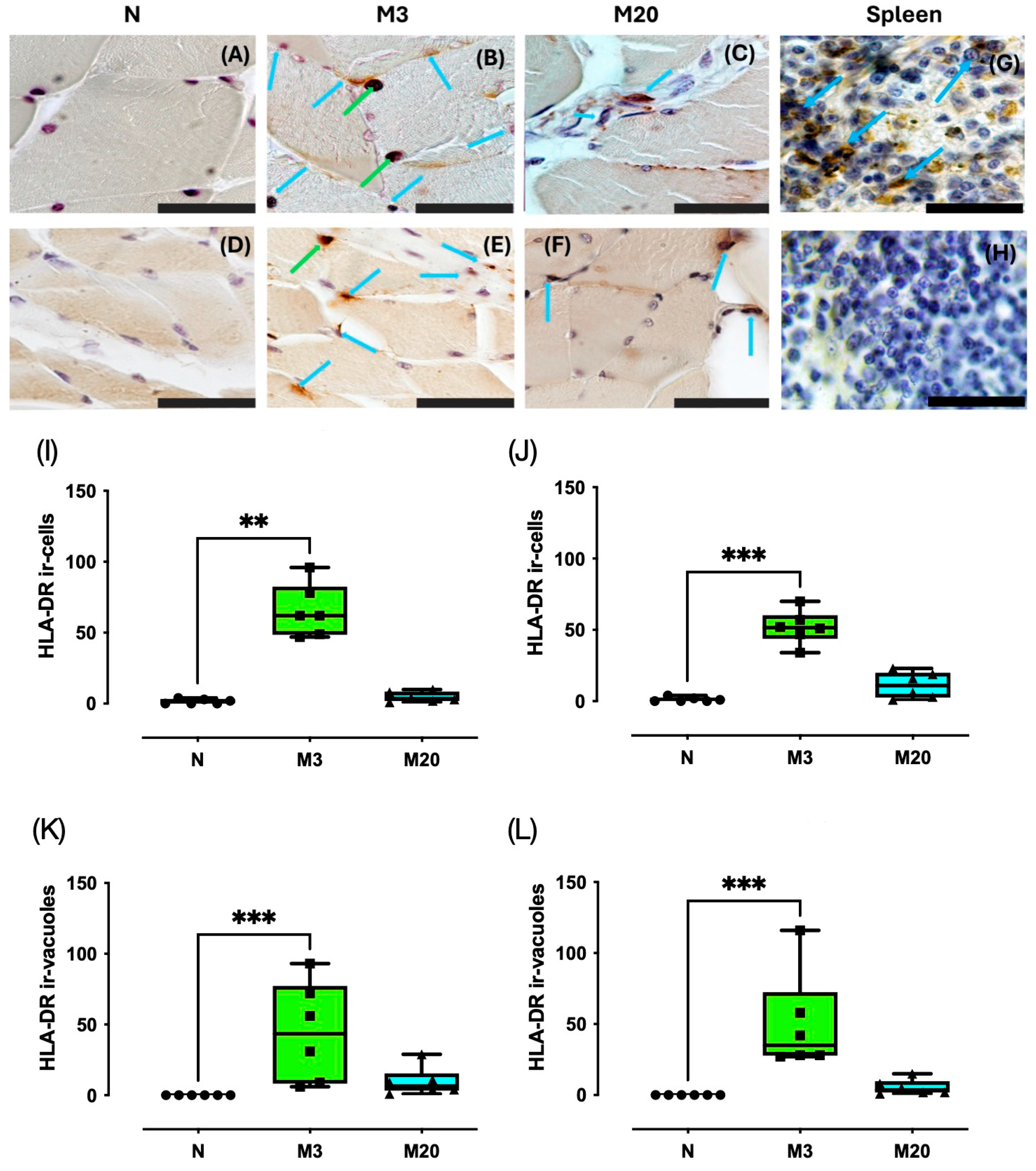
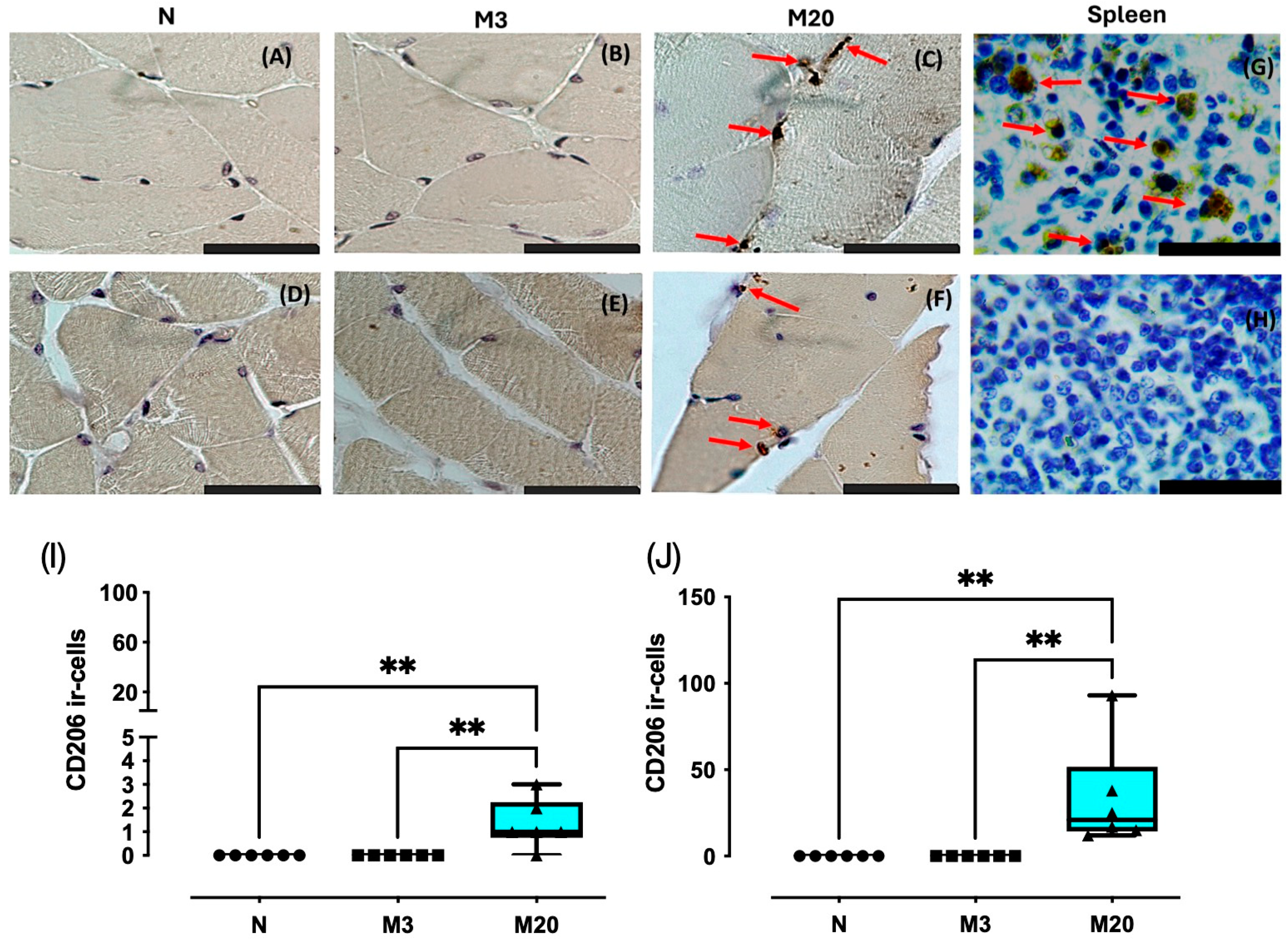
3. Results
3.1. Muscle Morphometry
3.2. HLA-DR Immunostaining
3.3. CD206 Immunostaining
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- DeLancey, J.O.L.; Masteling, M.; Pipitone, F.; LaCross, J.; Mastrovito, S.; Ashton-Miller, J.A. Pelvic floor injury during vaginal birth is life-altering and preventable: What can we do about it? Am. J. Obstet. Gynecol. 2024, 230, 279–294.e2. [Google Scholar] [CrossRef] [PubMed]
- Cassadó, J.; Simó, M.; Rodríguez, N.; Porta, O.; Huguet, E.; Mora, I.; Girvent, M.; Fernández, R.; Gich, I. Prevalence of levator ani avulsion in a multicenter study (PAMELA study). Arch. Gynecol. Obstet. 2020, 302, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Betschart, C.; Ramanah, R.; Ashton-Miller, J.A.; DeLancey, J.O.L. Anatomy of the pubovisceral muscle origin: Macroscopic and microscopic findings within the injury zone. Neurourol. Urodyn. 2015, 34, 774–780. [Google Scholar] [CrossRef] [PubMed]
- Youssef, A.; Brunelli, E.; Pilu, G.; Dietz, H.P. The maternal pelvic floor and labor outcome. Am. J. Obsterics. Gynecol. 2021, 3, 100452. [Google Scholar] [CrossRef] [PubMed]
- Nishibayashi, M.; Okagaki, R. Ultrasonographic evaluation of pelvic floor structure at antepartum and postpartum periods using three-dimensional transperineal ultrasound. J. Med. Ultrason. 2021, 48, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Regendova, A.; Samesova, A.; Zapletalova, K.; Horejskova, S.; Svata, Z.; Hrdouskova, M.; Zapletal, J.; Krofta, L.; Hajkova Hympanova, L. Comparison of Cross-Sectional Area of Pubovisceral Muscle in Nulliparous and Primiparous Women. Int. Urogynecol. J. 2024; advance online publication. [Google Scholar] [CrossRef]
- Li, Y.; Gong, T.; Lin, X.; Wei, X.; Cai, X.; Chen, X.; Lin, L.; Wang, G. Evaluating changes in the strength of the levator ani muscle after vaginal delivery using T2-parameter mapping. Eur. J. Radiol. 2023, 168, 111137. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Liu, B.; Lin, J.; Xue, T.; Han, Y.; Lu, C.; Zhou, S.; Gu, Y.; Xu, F.; Shen, Y.; et al. MuSCs and IPCs: Roles in skeletal muscle homeostasis, aging and injury. Cell. Mol. Life. Sci. 2024, 81, 67. [Google Scholar] [CrossRef] [PubMed]
- Callewaert, G.; Cunha, M.M.C.M.D.; Sindhwani, N.; Sampaolesi, M.; Albersen, M.; Deprest, J. Cell-based secondary prevention of childbirth-induced pelvic floor trauma. Nat. Rev. Urol. 2017, 14, 373–385. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhou, L. The multifaceted role of macrophages in homeostatic and injured skeletal muscle. Front. Immunol. 2023, 14, 1274816. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Peng, J.; Han, G.H.; Ding, X.; Wei, S.; Gao, G. Role of macrophages in peripheral nerve injury and repair. Neural. Regen. Res. 2019, 14, 1335–1342. [Google Scholar] [PubMed]
- Tidball, J.G. Regulation of muscle growth and regeneration by the immune system. Nat. Rev. Immunol. 2017, 17, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.S.; Carrier, S.; Morgan, D.M.; Lue, T.F. Effect of simulated birth trauma on the urinary continence mechanism in the rat. Urology 1998, 52, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Phull, H.S.; Pan, H.Q.; Butler, R.S.; Hansel, D.E.; Damaser, M.S. Vulnerability of continence structures to injury by simulated childbirth. Am. J. Physiol. Renal. Physiol. 2011, 301, F641–F649. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, S.; Sumino, Y.; Kwon, J.; Suzuki, T.; Kitta, T.; Miyazato, M. Effects of multiple simulated birth traumas on urethral continence function in rats. Am. J Physiol. Renal. Physiol. 2017, 313, F1089–F1096. [Google Scholar] [CrossRef] [PubMed]
- Alperin, M.; Lawley, D.M.; Esparza, M.C.; Lieber, R.L. Pregnancy-induced adaptations in the intrinsic structure of rat pelvic floor muscles. Am. J. Obst. Gynecol. 2015, 213, 191.e1–191.e1917. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Li, B.; Liu, C.; Hu, M.; Tang, J.; Min, J. M2 Macrophage-derived exosomal miR-501 contributes to pubococcygeal muscle regeneration. Int. Immunopharmacol. 2021, 101, 108223. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, A.J.; Sesillo, F.B.; Do, E.; Alperin, M. Effect of nonsteroidal anti-inflammatory drugs on pelvic floor muscle regeneration in a preclinical birth injury rat model. Am. J. Obst. Gynecol. 2023, 230, 432.e1–432.e14. [Google Scholar] [CrossRef] [PubMed]
- Corona-Quintanilla, D.L.; Castelán, F.; Fajardo, V.; Manzo, J.; Martínez-Gómez, M. Temporal coordination of pelvic and perineal striated muscle activity during micturition in female rabbits. J. Urol. 2009, 181, 1452–1458. [Google Scholar] [CrossRef] [PubMed]
- Thor, K.B.; de Groat, W.C. Neural control of the female urethral and anal rhabdosphincters and pelvic floor muscles. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 299, R416–R438. [Google Scholar] [CrossRef] [PubMed]
- López-García, K.; Cuevas, E.; Sánchez-García, O.; Pacheco, P.; Martínez-Gómez, M.; Castelán, F. Differential damage and repair responses of pubococcygeus and bulbospongiosus muscles in multiparous rabbits. Neurourol. Urodyn. 2016, 35, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Rajasekaran, M.R.; Fu, J.; Nguyen, M.U.L.; Wang, Y.; Albo, M.; Bhargava, V. Age and multiparity related urethral sphincter muscle dysfunction in a rabbit model: Potential roles of TGF-β and Wnt-β catenin signaling pathways. Neurourol. Urodyn. 2019, 38, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-García, O.; Hernández-Aragón, L.G.; López-García, K.; Juaréz, M.; Martínez-Gómez, M.; Castelán, F. Signs of damage in pelvic floor muscles at the end of pregnancy in rabbits. Int. Urogynecol. J. 2019, 30, 977–984. [Google Scholar] [CrossRef] [PubMed]
- López-García, K.; Cuevas, E.; Corona-Quintanilla, D.L.; Jiménez-Estrada, I.; Martínez-Gómez, M.; Castelán, F. Effect of multiparity on morphometry and oestrogen receptor expression of pelvic and perineal striated muscles in rabbits: Is serum oestradiol relevant? Eur. J. Obstet. Gyn. Reprod. Biol. 2013, 169, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Catanzarite, T.; Bremner, S.; Barlow, C.L.; Bou-Malham, L.; O’Connor, S.; Alperin, M. Pelvic muscles’ mechanical response to strains in the absence and presence of pregnancy-induced adaptations in a rat model. Am. J. Obstet. Gynecol. 2018, 218, 512.e1–512.e9. [Google Scholar] [CrossRef] [PubMed]
- Ema, R.; Nosaka, K.; Kawashima, R.; Kanda, A.; Ikeda, K.; Akagi, R. Muscle length influence on rectus femoris damage and protective effect in knee extensor eccentric exercise. Scand. J. Med. Sci. Sports. 2021, 31, 597–609. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Reynoso, A.G.; Corona-Quintanilla, D.L.; López-García, K.; Horbovetz, A.A.; Castelán, F.; Zimmern, P.; Martínez-Gómez, M.; Romero-Ortega, M. Targeted neuromodulation of pelvic floor nerves in aging and multiparous rabbits improves continence. Sci. Rep. 2021, 11, 10615. [Google Scholar] [CrossRef] [PubMed]
- Grigore, N.; Pirvut, V.; Mihai, I.; Ioan, S.; Mitariu, C.; Sava, M. Polymer Ligating Clips in Urologic Laparoscopic Surgery. Mat. Plas. 2017, 54, 295–297. [Google Scholar] [CrossRef]
- Zuk, J.A.; Fletcher, A. Skeletal muscle expression of class II histocompatibility antigens (HLA-DR) in polymyositis and other muscle disorders with an inflammatory infiltrate. J. Clin. Pathol. 1988, 41, 410. [Google Scholar] [CrossRef] [PubMed]
- Bernard, C.; Zavoriti, A.; Pucelle, Q.; Chazaud, B.; Gondin, J. Role of macrophages during skeletal muscle regeneration and hypertrophy-Implications for immunomodulatory strategies. Physiol. Rep. 2022, 10, e15480. [Google Scholar] [CrossRef] [PubMed]
- Inukai, A.; Kuru, S.; Liang, Y.; Takano, A.; Kobayashi, Y.; Sakai, M. Expression of HLA-DR and its enhancing molecules in muscle fibers in polymyositis. Muscle. Nerve. 2000, 23, 385–392. [Google Scholar] [CrossRef]
- Xu, Q.; Li, Q.X.; Bi, F.F.; Duan, H.Q.; Luo, Y.B.; Yang, H. The association between myositis-specific autoantibodies and muscle pathologies in idiopathic inflammatory myopathies. Clin. Rheumatol. 2021, 40, 613–624. [Google Scholar] [CrossRef] [PubMed]
- Duran, P.; Boscolo Sesillo, F.; Cook, M.; Burnett, L.; Menefee, S.A.; Do, E.; French, S.; Zazueta-Damian, G.; Dzieciatkowska, M.; Saviola, A.J.; et al. Proregenerative extracellular matrix hydrogel mitigates pathological alterations of pelvic skeletal muscles after birth injury. Sci. Trans. Med. 2023, 15, eabj3138. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Benítez, E.; López-García, K.; Xelhuantzi, N.; Corona-Quintanilla, D.L.; Castelán, F.; Martínez-Gómez, M. Shift from Pro- to Anti-Inflammatory Phase in Pelvic Floor Muscles at Postpartum Matches Histological Signs of Regeneration in Multiparous Rabbits. Medicina 2024, 60, 675. https://doi.org/10.3390/medicina60040675
Rodríguez-Benítez E, López-García K, Xelhuantzi N, Corona-Quintanilla DL, Castelán F, Martínez-Gómez M. Shift from Pro- to Anti-Inflammatory Phase in Pelvic Floor Muscles at Postpartum Matches Histological Signs of Regeneration in Multiparous Rabbits. Medicina. 2024; 60(4):675. https://doi.org/10.3390/medicina60040675
Chicago/Turabian StyleRodríguez-Benítez, Esteban, Kenia López-García, Nicte Xelhuantzi, Dora Luz Corona-Quintanilla, Francisco Castelán, and Margarita Martínez-Gómez. 2024. "Shift from Pro- to Anti-Inflammatory Phase in Pelvic Floor Muscles at Postpartum Matches Histological Signs of Regeneration in Multiparous Rabbits" Medicina 60, no. 4: 675. https://doi.org/10.3390/medicina60040675