
Targeted Metabolite Fingerprints of Thirteen Gambierdiscus, Five Coolia and Two Fukuyoa Species

 , ,
, ,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
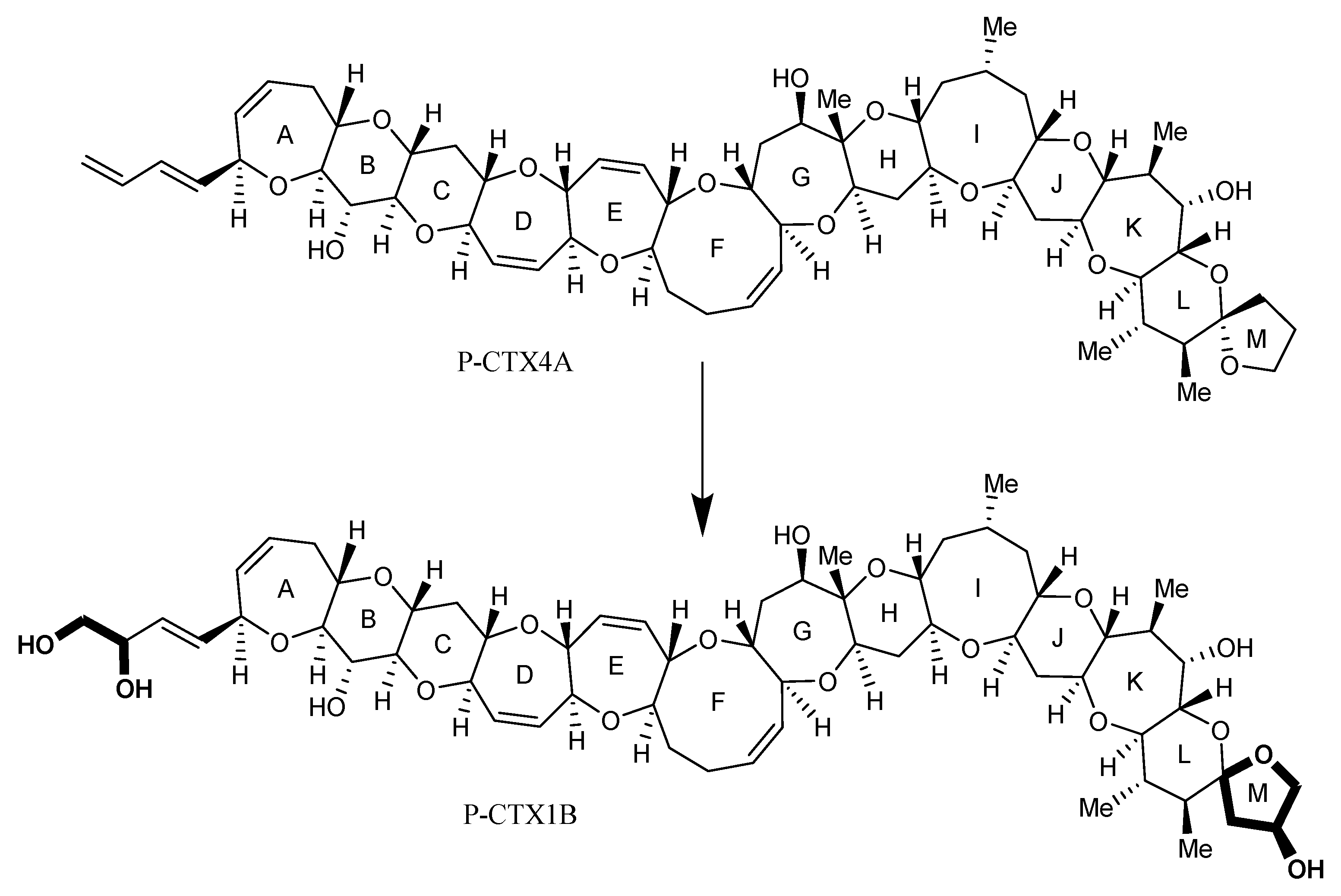
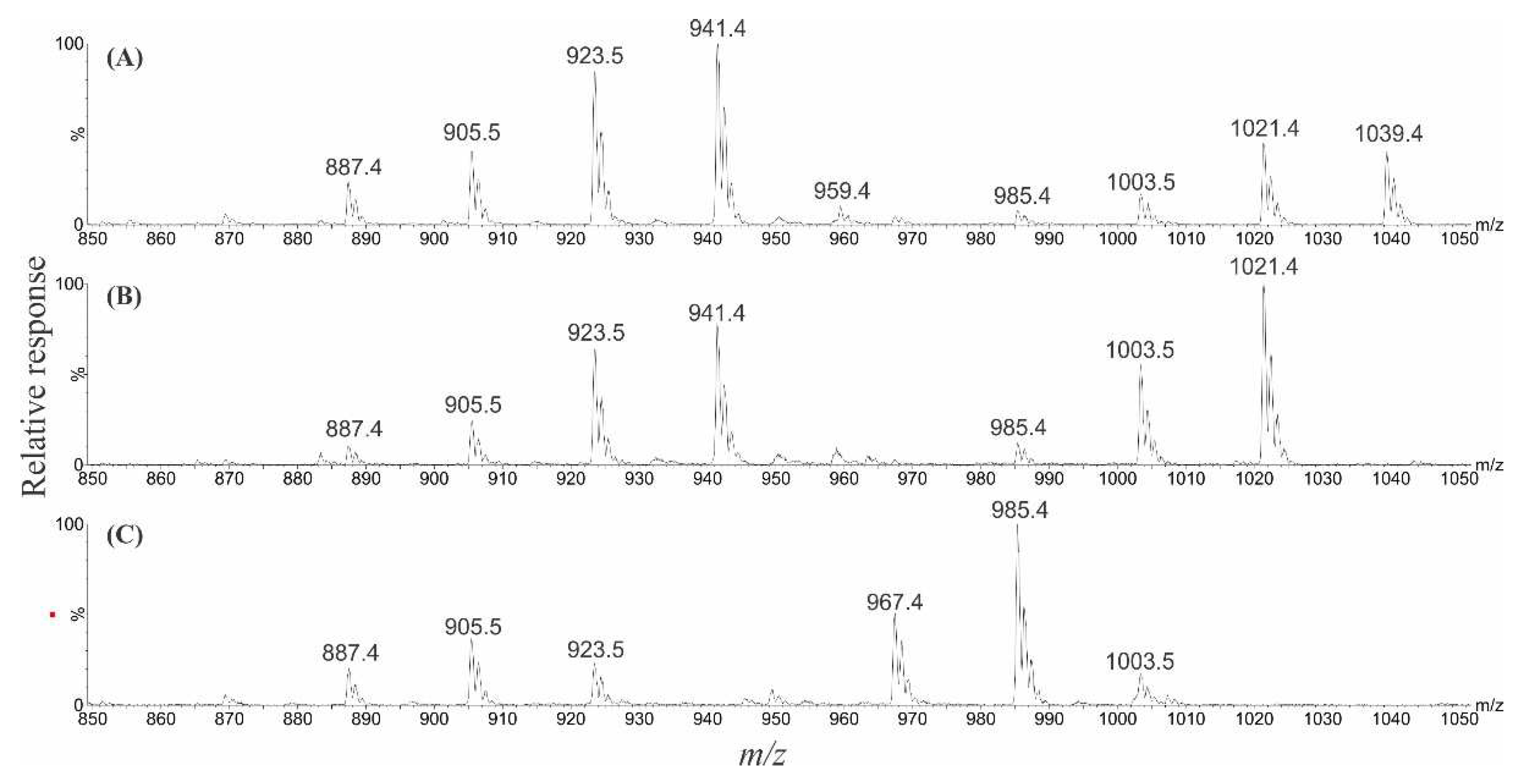
2.1. Ciguatoxins
2.2. Maitotoxins
2.3. Gambierones
2.4. Other Metabolites
2.5. Metabolite Spiking Experiments
2.6. Liquid Chromatography–Mass Spectrometry Analysis of Gambierones
3. Discussion
4. Materials and Methods
4.1. Mapping the Metabolite Fingerprint
4.1.1. Microalgal Isolates
4.1.2. Targeted Liquid Chromatography–Mass Spectrometry
Ciguatoxins
Maitotoxins and Gambieroxide
Gambierones
Gambierol and Gambieric Acids
4.2. Tentative Gambierone Analogues
4.2.1. Culturing
4.2.2. Extraction and Isolation
4.2.3. Liquid Chromatography–Mass Spectrometry Analysis of Gambierone Analogues
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Botana, L.M. Toxicology of Seafood Toxins: A Critical Review; CRC Press: Boca Raton, FL, USA, 2014; pp. 100–196. [Google Scholar]
- Bagnis, R.; Chanteau, S.; Chungue, E.; Hurtel, J.M.; Yasumoto, T.; Inoue, A. Origins of ciguatera fish poisoning: A new dinoflagellate, Gambierdiscus toxicus Adachi and Fukuyo, definitively involved as a causal agent. Toxicon 1980, 18, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Chinain, M.; Darius, H.T.; Ung, A.; Fouc, M.T.; Revel, T.; Cruchet, P.; Pauillac, S.; Laurent, D. Ciguatera risk management in French Polynesia: The case study of Raivavae Island (Australes Archipelago). Toxicon 2010, 56, 674–690. [Google Scholar] [CrossRef]
- Friedman, M.A.; Fernandez, M.; Backer, L.C.; Dickey, R.W.; Bernstein, J.; Schrank, K.; Kibler, S.; Stephan, W.; Gribble, M.O.; Bienfang, P.; et al. An updated review of ciguatera fish poisoning: Clinical, epidemiological, environmental, and public health management. Mar. Drugs 2017, 15, 72. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organisation of the United Nations. Marine Biotoxins; Food and Agriculture Organisation of the United Nations: Rome, Italy, 2004; p. 217. [Google Scholar]
- Swift, A.E.; Swift, T.R. Ciguatera. J. Toxicol. Clin. Toxicol. 1993, 31, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Institute Loius Malarde. Ciguatera Online. 2014. Available online: http://www.ciguatera-online.com/index.php/en/ (accessed on 11 November 2022).
- Urdaneta, A. Relación de los Sucesos de la Armada del Comendador Loaiza a Las Islas de la Especiería o Molucas en 1525 y Sucesos Acaecidos en Ellas Hasta el 1536; Real Biblioteca del Palacio Real de Madrid: Madrid, Spain, 1580; Volume II, p. 1465. [Google Scholar]
- Tester, P.A.; Litaker, R.W.; Soler-Onís, E.; Fernández-Zabala, J.; Berdalet, E. Using artificial substrates to quantify Gambierdiscus and other toxic benthic dinoflagellates for monitoring purposes. Harmful Algae 2022, 120, 102351. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Richlen, M.L.; Liefer, J.D.; Robertson, A.; Kulis, D.; Smith, T.B.; Parsons, M.L.; Anderson, D.M. Influence of environmental variables on Gambierdiscus spp. (Dinophyceae) growth and distribution. PLoS ONE 2016, 11, 490–504. [Google Scholar] [CrossRef] [PubMed]
- Rongo, T.; van Woesik, R. The effects of natural disturbances, reef state, and herbivorous fish densities on ciguatera poisoning in Rarotonga, southern Cook Islands. Toxicon 2013, 64, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Loeffler, C.R.; Tartaglione, L.; Friedemann, M.; Spielmeyer, A.; Kappenstein, O.; Bodi, D. Ciguatera mini review: 21st century environmental challenges and the interdisciplinary research efforts rising to meet them. Int. J. Environ. Res. Public Health 2021, 18, 3027. [Google Scholar] [CrossRef]
- Chinain, M.; Gatti, C.M.i.; Darius, H.T.; Quod, J.P.; Tester, P.A. Ciguatera poisonings: A global review of occurrences and trends. Harmful Algae 2021, 102, 101873. [Google Scholar] [CrossRef]
- Smith, K.F.; Rhodes, L.; Verma, A.; Curley, B.G.; Harwood, D.T.; Kohli, G.S.; Solomona, D.; Rongo, T.; Munday, R.; Murray, S.A. A new Gambierdiscus species (Dinophyceae) from Rarotonga, Cook Islands: Gambierdiscus cheloniae sp. nov. Harmful Algae 2016, 60, 45–56. [Google Scholar] [CrossRef]
- Rhodes, L.; Smith, K.F.; Verma, A.; Curley, B.G.; Harwood, D.T.; Murray, J.S.; Kohli, G.S.; Solomona, D.; Rongo, T.; Munday, R.; et al. A new species of Gambierdiscus (Dinophyceae) from the south-west Pacific: Gambierdiscus honu sp. nov. Harmful Algae 2017, 65, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Kretzschmar, A.L.; Verma, A.; Harwood, T.; Hoppenrath, M.; Murray, S. Characterization of Gambierdiscus lapillus sp. nov. (Gonyaulacales, Dinophyceae): A new toxic dinoflagellate from the Great Barrier Reef (Australia). J. Phycol. 2017, 53, 283–297. [Google Scholar] [CrossRef] [PubMed]
- Kretzschmar, A.L.; Larsson, M.E.; Hoppenrath, M.; Doblin, M.A.; Murray, S.A. Characterisation of two toxic Gambierdiscus spp. (Gonyaulacales, Dinophyceae) from the Great Barrier Reef (Australia): G. lewisii sp. nov. and G. holmesii sp. nov. Protist 2019, 170, 125699. [Google Scholar] [CrossRef] [PubMed]
- Fraga, S.; Rodríguez, F.; Riobó, P.; Bravo, I. Gambierdiscus balechii sp. nov (Dinophyceae), a new benthic toxic dinoflagellate from the Celebes Sea (SW Pacific Ocean). Harmful Algae 2016, 58, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Nguyen-Ngoc, L.; Larsen, J.; Doan-Nhu, H.; Nguyen, X.-V.; Chomérat, N.; Lundholm, N.; Phan-Tan, L.; Dao, H.V.; Nguyen, N.-L.; Nguyen, H.-H.; et al. Gambierdiscus (gonyaulacales, dinophyceae) diversity in Vietnamese waters with description of G. vietnamensis sp. nov. J. Phycol. 2023, 59, 496–517. [Google Scholar] [CrossRef] [PubMed]
- Gómez, F.; Qiu, D.; Lopes, R.M.; Lin, S. Fukuyoa paulensis sp. nov., a new genus for the globular species of the dinoflagellate Gambierdiscus (Dinophyceae). PLoS ONE 2015, 10, e0119676. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Park, J.S.; Kang, N.S.; Chomérat, N.; Mertens, K.N.; Gu, H.; Lee, K.-W.; Kim, K.H.; Baek, S.H.; Shin, K.; et al. A new potentially toxic dinoflagellate Fukuyoa koreansis sp. nov. (Gonyaulacales, Dinophyceae) from Korean coastal waters: Morphology, phylogeny, and effects of temperature and salinity on growth. Harmful Algae 2020, 109, 102107. [Google Scholar] [CrossRef] [PubMed]
- Leung, P.T.; Yan, M.; Lam, V.T.; Yiu, S.K.; Chen, C.-Y.; Murray, J.S.; Harwood, D.T.; Rhodes, L.L.; Lam, P.K.; Wai, T.-C.J.H.a. Phylogeny, morphology and toxicity of benthic dinoflagellates of the genus Fukuyoa (Goniodomataceae, Dinophyceae) from a subtropical reef ecosystem in the South China Sea. Harmful Algae 2018, 74, 78–97. [Google Scholar] [CrossRef]
- Murray, J.S.; Finch, S.C.; Puddick, J.; Rhodes, L.L.; Harwood, D.T.; van Ginkel, R.; Prinsep, M.R. Acute toxicity of gambierone and quantitative analysis of gambierones produced by cohabitating benthic dinoflagellates. Toxins 2021, 13, 333. [Google Scholar] [CrossRef]
- Tibiriçá, C.E.J.d.A.; Sibat, M.; Fernandes, L.F.; Bilien, G.; Chomérat, N.; Hess, P.; Mafra Jr, L.L. Diversity and Toxicity of the Genus Coolia Meunier in Brazil, and Detection of 44-methylgambierone in Coolia tropicalis. Toxins 2020, 12, 327. [Google Scholar] [CrossRef]
- Roué, M.; Darius, H.T.; Picot, S.; Ung, A.; Viallon, J.; Gaertner-Mazouni, N.; Sibat, M.; Amzil, Z.; Chinain, M. Evidence of the bioaccumulation of ciguatoxins in giant clams (Tridacna maxima) exposed to Gambierdiscus spp. cells. Harmful Algae 2016, 57, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Kelly, A.M.; Kohler, C.C.; Tindall, D.R. Are crustaceans linked to the ciguatera food chain? Environ. Biol. Fishes 1992, 33, 275–286. [Google Scholar] [CrossRef]
- Silva, M.; Rodriguez, I.; Barreiro, A.; Kaufmann, M.; Neto, A.I.; Hassouani, M.; Sabour, B.; Alfonso, A.; Botana, L.M.; Vasconcelos, V. First report of ciguatoxins in two starfish species: Ophidiaster ophidianus and Marthasterias glacialis. Toxins 2015, 7, 3740–3757. [Google Scholar] [CrossRef] [PubMed]
- Darius, H.T.; Roué, M.; Sibat, M.; Viallon, J.; Gatti, C.M.i.; Vandersea, M.W.; Tester, P.A.; Litaker, R.W.; Amzil, Z.; Hess, P.; et al. Toxicological investigations on the sea urchin Tripneustes gratilla (Toxopneustidae, Echinoid) from Anaho Bay (Nuku Hiva, French Polynesia): Evidence for the presence of Pacific ciguatoxins. Mar. Drugs 2018, 16, 122. [Google Scholar] [CrossRef] [PubMed]
- Gatti, C.M.I.; Lonati, D.; Darius, H.T.; Zancan, A.; Roué, M.; Schicchi, A.; Locatelli, C.A.; Chinain, M. Tectus niloticus (Tegulidae, Gastropod) as a novel vector of ciguatera poisoning: Clinical characterization and follow-up of a mass poisoning event in Nuku Hiva Island (French Polynesia). Toxins 2018, 10, 102. [Google Scholar] [CrossRef] [PubMed]
- Mak, Y.L.; Wai, T.-C.; Murphy, M.B.; Chan, W.H.; Wu, J.J.; Lam, J.C.W.; Chan, L.L.; Lam, P.K.S. Pacific ciguatoxins in food web components of coral reef systems in the Republic of Kiribati. Environ. Sci. Technol. 2013, 47, 14070–14079. [Google Scholar] [CrossRef] [PubMed]
- Rongo, T.; van Woesik, R. Ciguatera poisoning in Rarotonga, southern Cook Islands. Harmful Algae 2011, 10, 345–355. [Google Scholar] [CrossRef]
- Yogi, K.; Sakugawa, S.; Oshiro, N.; Ikehara, T.; Sugiyama, K.; Yasumoto, T. Determination of toxins involved in ciguatera fish poisoning in the Pacific by LC/MS. J. AOAC Int. 2014, 97, 398–402. [Google Scholar] [CrossRef]
- Ikehara, T.; Kuniyoshi, K.; Oshiro, N.; Yasumoto, T. Biooxidation of ciguatoxins leads to species-specific toxin profiles. Toxins 2017, 9, 205. [Google Scholar] [CrossRef]
- Zhang, F.; Xu, X.; Li, T.; Liu, Z. Shellfish toxins targeting voltage-gated sodium channels. Mar. Drugs 2013, 11, 4698. [Google Scholar] [CrossRef]
- Rayner, M.D.; Szekerczes, J. Ciguatoxin: Effects on the sodium-potassium activated adenosine triphosphatase of human erythrocyte ghosts. Toxicol. Appl. Pharmacol. 1973, 24, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organisation of the United Nations. Report of the Expert Meeting on Ciguatera Poisoning. Rome, 19–23 November 2018; Food and Agriculture Organisation of the United Nations: Rome, Italy, 2020; p. 156. [Google Scholar]
- Vernoux, J.P.; Lewis, R.J. Isolation and characterisation of Caribbean ciguatoxins from the horse-eye jack (Caranx latus). Toxicon 1997, 35, 889–900. [Google Scholar] [CrossRef] [PubMed]
- Kryuchkov, F.; Robertson, A.; Miles, C.O.; Mudge, E.M.; Uhlig, S. LC–HRMS and chemical derivatization strategies for the structure elucidation of Caribbean ciguatoxins: Identification of C-CTX-3 and -4. Mar. Drugs 2020, 18, 182. [Google Scholar] [CrossRef] [PubMed]
- Mudge, E.M.; Miles, C.O.; Ivanova, L.; Uhlig, S.; James, K.S.; Erdner, D.L.; Fæste, C.K.; McCarron, P.; Robertson, A. Algal ciguatoxin identified as source of ciguatera poisoning in the Caribbean. Chemosphere 2023, 330, 138659. [Google Scholar] [CrossRef]
- Hamilton, B.; Hurbungs, M.; Jones, A.; Lewis, R.J. Multiple ciguatoxins present in Indian Ocean reef fish. Toxicon 2002, 40, 1347–1353. [Google Scholar] [CrossRef]
- Murata, M.; Naoki, H.; Iwashita, T.; Matsunaga, S.; Sasaki, M.; Yokoyama, A.; Yasumoto, T. Structure of maitotoxin. J. Am. Chem. Soc. 1993, 115, 2060–2062. [Google Scholar] [CrossRef]
- Xi, D.; Van Dolah, F.; Ramsdell, J. Maitotoxin induces a calcium-dependent membrane depolarization in GH4C1 pituitary cells via activation of type L voltage-dependent calcium channels. J. Biol. Chem. 1992, 267, 25025–25031. [Google Scholar] [CrossRef]
- Kobayashi, M.; Ohizumi, Y.; Yasumoto, T. The mechanism of action of maitotoxin in relation to Ca2+ movements in guinea pig and rat cardiac muscles. Br. J. Pharmacol. 1985, 86, 385–391. [Google Scholar] [CrossRef]
- Seymour, B.; Andreosso, A.; Seymour, J. Cardiovascular Toxicity from Marine Envenomation; Academic Press: Cambridge, MA, USA, 2015; pp. 203–223. [Google Scholar] [CrossRef]
- Blumenthal, K.M. Ion Channels as Targets for Toxins; Academic Press: Cambridge, MA, USA, 2012; pp. 509–524. [Google Scholar] [CrossRef]
- Yokoyama, A.; Murata, M.; Oshima, Y.; Iwashita, T.; Yasumoto, T. Some chemical properties of maitotoxin, a putative calcium channel agonist isolated from a marine dinoflagellate. J. Biochem. 1988, 104, 184–187. [Google Scholar] [CrossRef]
- Yasumoto, T.; Bagnis, R.; Vernoux, J.P. Toxicity of the Surgeonfishes-II. Properties of the Principal Water-soluble Toxin. Nippon Suisan Gakkaishi 1976, 42, 359–365. [Google Scholar] [CrossRef]
- Lewis, R.J.; Holmes, M.J.; Alewood, P.F.; Jones, A. Ionspray mass spectrometry of ciguatoxin-1, maitotoxin-2 and -3, and related marine polyether toxins. Nat. Toxins 1994, 2, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Murray, J.S.; Selwood, A.I.; Harwood, D.T.; van Ginkel, R.; Puddick, J.; Rhodes, L.L.; Rise, F.; Wilkins, A.L. 44-Methylgambierone, a new gambierone analogue isolated from Gambierdiscus australes. Tetrahedron Lett. 2019, 60, 621–625. [Google Scholar] [CrossRef]
- Boente-Juncal, A.; Álvarez, M.; Antelo, Á.; Rodríguez, I.; Calabro, K.; Vale, C.; Thomas, O.P.; Botana, L.M. Structure elucidation and biological evaluation of maitotoxin-3, a homologue of gambierone, from Gambierdiscus belizeanus. Toxins 2019, 11, 79. [Google Scholar] [CrossRef] [PubMed]
- Pisapia, F.; Sibat, M.; Herrenknecht, C.; Lhaute, K.; Gaiani, G.; Ferron, P.-J.; Fessard, V.; Fraga, S.; Nascimento, S.M.; Litaker, R.W. Maitotoxin-4, a novel MTX analog produced by Gambierdiscus excentricus. Mar. Drugs 2017, 15, 220. [Google Scholar] [CrossRef] [PubMed]
- Estevez, P.; Castro, D.; Leão-Martins, J.M.; Sibat, M.; Tudó, A.; Dickey, R.; Diogene, J.; Hess, P.; Gago-Martinez, A. Toxicity screening of a Gambierdiscus australes strain from the Western Mediterranean Sea and identification of a novel maitotoxin analogue. Mar. Drugs 2021, 19, 460. [Google Scholar] [CrossRef] [PubMed]
- Murray, J.S.; Finch, S.C.; Mudge, E.M.; Wilkins, A.L.; Puddick, J.; Harwood, D.T.; Rhodes, L.L.; van Ginkel, R.; Rise, F.; Prinsep, M.R. Structural characterization of maitotoxins produced by toxic Gambierdiscus species. Mar. Drugs 2022, 20, 453. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, I.s.; Genta-Jouve, G.g.; Alfonso, C.; Calabro, K.; Alonso, E.; Sánchez, J.A.; Alfonso, A.; Thomas, O.P.; Botana, L.M. Gambierone, a ladder-shaped polyether from the dinoflagellate Gambierdiscus belizeanus. Org. Lett. 2015, 17, 2392–2395. [Google Scholar] [CrossRef]
- Yon, T.; Sibat, M.; Robert, E.; Lhaute, K.; Holland, W.C.; Litaker, R.W.; Bertrand, S.; Hess, P.; Réveillon, D. Sulfo-gambierones, two new analogs of gambierone Produced by Gambierdiscus excentricus. Mar. Drugs 2021, 19, 657. [Google Scholar] [CrossRef]
- Liu, X.; Ma, Y.; Wu, J.; Yin, Q.; Wang, P.; Zhu, J.; Chan, L.L.; Wu, B. Characterization of New Gambierones Produced by Gambierdiscus balechii 1123M1M10. Mar. Drugs 2023, 21, 3. [Google Scholar] [CrossRef]
- Murray, J.S.; Nishimura, T.; Finch, S.C.; Rhodes, L.L.; Puddick, J.; Harwood, D.T.; Larsson, M.E.; Doblin, M.A.; Leung, P.; Yan, M.; et al. The role of 44-methylgambierone in ciguatera fish poisoning: Acute toxicity, production by marine microalgae and its potential as a biomarker for Gambierdiscus spp. Harmful Algae 2020, 97, 101853. [Google Scholar] [CrossRef]
- Stuart, J.; Smith, K.F.; Rhodes, L.; Murray, J.S.; Viallon, J.; Henry, K.; Darius, H.T.; Murray, S.A.; De Azevedo, C.D.; Argyle, P.; et al. Geographical distribution, molecular and toxin diversity of the dinoflagellate species Gambierdiscus honu in the Pacific region. Harmful Algae 2022, 118, 102308. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, R.; Uchida, H.; Suzuki, T.; Matsushima, R.; Nagae, M.; Toyohara, Y.; Satake, M.; Oshima, Y.; Inoue, A.; Yasumoto, T. Gambieroxide, a novel epoxy polyether compound from the dinoflagellate Gambierdiscus toxicus GTP2 strain. Tetrahedron Lett. 2013, 69, 10299–10303. [Google Scholar] [CrossRef]
- Morohashi, A.; Satake, M.; Yasumoto, T. The absolute configuration of gambierol, a toxic marine polyether from the dinoflagellate, Gambierdiscus toxicus. Tetrahedron Lett. 1999, 40, 97–100. [Google Scholar] [CrossRef]
- Cuypers, E.; Abdel-Mottaleb, Y.; Kopljar, I.; Rainier, J.D.; Raes, A.L.; Snyders, D.J.; Tytgat, J. Gambierol, a toxin produced by the dinoflagellate Gambierdiscus toxicus, is a potent blocker of voltage-gated potassium channels. Toxicon 2008, 51, 974–983. [Google Scholar] [CrossRef] [PubMed]
- Ghiaroni, V.; Sasaki, M.; Fuwa, H.; Rossini, G.P.; Scalera, G.; Yasumoto, T.; Pietra, P.; Bigiani, A. Inhibition of voltage-gated potassium currents by gambierol in mouse taste cells. Toxicol. Sci. 2005, 85, 657–665. [Google Scholar] [CrossRef] [PubMed]
- Fuwa, H.; Kainuma, N.; Satake, M.; Sasaki, M. Synthesis and biological evaluation of gambierol analogues. Bioorganic Med. Chem. Lett. 2003, 13, 2519–2522. [Google Scholar] [CrossRef] [PubMed]
- Fuwa, H.; Kainuma, N.; Tachibana, K.; Tsukano, C.; Satake, M.; Sasaki, M. Diverted total synthesis and biological evaluation of gambierol analogues: Elucidation of crucial structural elements for potent toxicity. Chem. A Eur. J. 2004, 10, 4894–4909. [Google Scholar] [CrossRef] [PubMed]
- Morohashi, A.; Satake, M.; Nagai, H.; Oshima, Y.; Yasumoto, T. The absolute configuration of gambieric acids A-D, potent antifungal polyethers, isolated from the marine dinoflagellate Gambierdiscus toxicus. Tetrahedron 2000, 56, 8995–9001. [Google Scholar] [CrossRef]
- Diogene, J.; Campas, M. Recent Advances in the Analysis of Marine Toxins; Wilson and Wilsons: Amsterdam, The Netherlands, 2018; Volume 78, p. 466. [Google Scholar]
- Soliño, L.; Costa, P.R. Differential toxin profiles of ciguatoxins in marine organisms: Chemistry, fate and global distribution. Toxicon 2018, 150, 124–143. [Google Scholar] [CrossRef]
- Food and Agriculture Organisation of the United Nations. Application of Risk Assessment in the Fish Industry; Food and Agriculture Organisation of the United Nations: Rome, Italy, 2004; p. 221. [Google Scholar]
- Munday, R.; Murray, J.S.; Rhodes, L.L.; Larsson, M.E.; Harwood, D.T. Ciguatoxins and maitotoxins in extracts of sixteen Gambierdiscus isolates and one Fukuyoa isolate from the south Pacific and their toxicity to mice by intraperitoneal and oral administration. Mar. Drugs 2017, 15, 208. [Google Scholar] [CrossRef]
- Mudge, E.M.; Robertson, A.; Leynse, A.K.; McCarron, P.; Miles, C.O. Selective extraction of gambierone and related metabolites in Gambierdiscus silvae using m-aminophenylboronic acid–agarose gel and liquid chromatography–high-resolution mass spectrometric detection. J. Chromatogr. B 2022, 1188, 123014. [Google Scholar] [CrossRef]
- Verma, A.; Barua, A.; Ruvindy, R.; Savela, H.; Ajani, P.A.; Murray, S.A. The genetic basis of toxin biosynthesis in dinoflagellates. Microorganisms 2019, 7, 222. [Google Scholar] [CrossRef]
- Kohli, G.S.; Campbell, K.; John, U.; Smith, K.F.; Fraga, S.; Rhodes, L.L.; Murray, S.A. Role of modular polyketide synthases in the production of polyether ladder compounds in ciguatoxin-producing Gambierdiscus polynesiensis and G. excentricus (Dinophyceae). J. Eukaryot. Microbiol. 2017, 64, 691–706. [Google Scholar] [CrossRef]
- Kohli, G.S.; John, U.; Figueroa, R.I.; Rhodes, L.L.; Harwood, D.T.; Groth, M.; Bolch, C.J.S.; Murray, S.A. Polyketide synthesis genes associated with toxin production in two species of Gambierdiscus (Dinophyceae). BMC Genom. 2015, 16, 410. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Culture ID | Scientific Name | Location | pg/Cell | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P-CTX | iso-P-CTX | MTX | G | 44-MG | ||||||||||
| 3B | 3C | 4A | 4B a | 3B/C b | 4A/B a | −1 | −5 c,d | −6 d | −7 d | |||||
| CAWD149 | G. australes | Rarotonga | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | 6 | 0.2 | ◦ | ◦ | ◦ e | 259 e |
| CAWD381 | G. australes | Kermadec Islands | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | 9 | 0.1 | ◦ | ◦ | ◦ | 160 |
| CCMP401 | G. belizeanus | St. Barthelemy Island | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | 540 | 53 |
| CAWD301 | G. caribaeus | Pohnpei | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ e | 44 e |
| CAWD237 | G. carpenteri f | Australia | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | 87 e | 74 e |
| CAWD237 | G. carpenteri | Australia | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | 65 e | 45 e |
| CAWD364 | G. carpenteri | Australia | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | 441 |
| CAWD232 | G. cheloniae | Rarotonga | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | 4 | ◦ | 55 e | 26 e |
| CAWD236 | G. cheloniae | Rarotonga | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | 5 | ◦ | 358 | 43 |
| CAWD368 | G. holmesii | Australia | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | 20 e | 97 e |
| CAWD242 | G. honu | Rarotonga | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | 14 | 38 e | 182 e |
| CAWD250 | G. honu | Tonga | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | 2 | 42 | 54 |
| NIES-4120 | G. jejuensis | Japan | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | 71 |
| NIES-4120 | G. jejuensis g | Japan | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | 72 |
| CAWD336 | G. lapillus | Rarotonga | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ e | 46 e |
| CAWD338 | G. lapillus | Rarotonga | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ e | 270 e |
| CAWD369 | G. lewisii | Australia | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | 1 e | 68 e |
| CAWD227 | G. pacificus | Rarotonga | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | 8 | 65 |
| CAWD337 | G. pacificus | Rarotonga | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | 1 e | 100 e |
| CAWD212 | G. polynesiensis | Rarotonga | 1.1 | 0.19 | 0.13 | 0.04 | 5.1 | 1.3 | ◦ | ◦ | ◦ | ◦ | 13 e | 29 e |
| CAWD267 | G. polynesiensis | Rarotonga | 0.82 | 0.14 | 0.05 | 0.02 | 7.8 | 2.0 | ◦ | ◦ | ◦ | ◦ | 13 e | 44 e |
| CAWD429 | G. scabrosus | Japan | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | 131 | 17 |
| CAWD429 | G. scabrosus g | Japan | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | 102 | 17 |
| K070922_1 | G. scabrosus | Japan | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | 166 | 32 |
| CAWD385 | C. canariensis | Kermadec Islands | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ |
| CAWD387 | C. canariensis | Kermadec Islands | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ |
| CAWD154 | C. malayensis | New Zealand | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | 2 e | 9 e |
| CAWD175 | C. malayensis | New Zealand | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | 17 e | 24 e |
| CAWD60 | C. monotis | Spain | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ |
| UTS4 | C. palmyrensis | Australia | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ |
| UTS25 | C. palmyrensis | Australia | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ |
| CAWD384 | C. tropicalis | Kermadec Islands | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ e | 14 e |
| CAWD388 | C. tropicalis | Kermadec Islands | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ e | 15 e |
| CAWD238 | F. paulensis | New Zealand | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ e | 5 e |
| CAWD306 | F. paulensis | New Zealand | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ e | 65 e |
| S044 | F. ruetzleri | Hong Kong | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | 8 e | 12 e |
| S051 | F. ruetzleri | Hong Kong | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | 6 e | 13 e |
| Culture ID | Scientific Name | Location | Detected/Not Detected | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Anhydro G | DiH-sulfo-G a | 12,13-diH-44-MG a | 29-MG | 38-deOH-44-MG a | 38-deOH-12,13-diH-44-MG a | Gambieroxide a | Gambieric Acid a | ||||||
| A | B | C | D | ||||||||||
| CAWD149 | G. australes | Rarotonga | ◦ | ◦ | D | ◦ | D | ◦ | ◦ | D | D | ◦ | ◦ |
| CAWD381 | G. australes | Kermadec Islands | ◦ | ◦ | D | ◦ | D | ◦ | ◦ | D | D | D | D |
| CCMP401 | G. belizeanus | St. Barthelemy Island | D | ◦ | D | ◦ | ◦ | D | ◦ | D | D | ◦ | ◦ |
| CAWD301 | G. caribaeus | Pohnpei | ◦ | ◦ | D | D | D | ◦ | ◦ | D | ◦ | ◦ | ◦ |
| CAWD237 | G. carpenteri b | Australia | D | ◦ | D | ◦ | D | D | ◦ | D | ◦ | ◦ | ◦ |
| CAWD237 | G. carpenteri | Australia | D | ◦ | D | ◦ | D | D | ◦ | D | ◦ | ◦ | ◦ |
| CAWD364 | G. carpenteri | Australia | ◦ | ◦ | D | ◦ | ◦ | D | ◦ | D | ◦ | ◦ | ◦ |
| CAWD232 | G. cheloniae | Rarotonga | D | ◦ | D | ◦ | ◦ | D | ◦ | D | ◦ | ◦ | ◦ |
| CAWD236 | G. cheloniae | Rarotonga | D | ◦ | D | ◦ | ◦ | D | ◦ | ◦ | ◦ | ◦ | ◦ |
| CAWD368 | G. holmesii | Australia | D | ◦ | D | ◦ | ◦ | D | ◦ | D | ◦ | ◦ | ◦ |
| CAWD242 | G. honu | Rarotonga | D | ◦ | D | ◦ | D | D | ◦ | ◦ | ◦ | ◦ | ◦ |
| CAWD250 | G. honu | Tonga | D | ◦ | D | D | D | D | ◦ | ◦ | D | ◦ | ◦ |
| NIES-4120 | G. jejuensis | Japan | ◦ | ◦ | D | D | D | D | ◦ | ◦ | ◦ | ◦ | ◦ |
| NIES-4120 | G. jejuensis c | Japan | ◦ | ◦ | D | D | D | D | ◦ | ◦ | D | ◦ | ◦ |
| CAWD336 | G. lapillus | Rarotonga | ◦ | D | D | D | D | D | ◦ | D | ◦ | ◦ | ◦ |
| CAWD338 | G. lapillus | Rarotonga | ◦ | D | D | D | D | D | ◦ | D | ◦ | ◦ | ◦ |
| CAWD369 | G. lewisii | Australia | ◦ | ◦ | D | ◦ | D | D | D | D | ◦ | ◦ | ◦ |
| CAWD227 | G. pacificus | Rarotonga | D | ◦ | D | D | D | D | D | D | ◦ | ◦ | ◦ |
| CAWD337 | G. pacificus | Rarotonga | D | ◦ | D | D | D | D | D | D | ◦ | ◦ | ◦ |
| CAWD212 | G. polynesiensis | Rarotonga | D | ◦ | D | D | ◦ | D | ◦ | D | ◦ | ◦ | ◦ |
| CAWD267 | G. polynesiensis | Rarotonga | D | ◦ | D | D | ◦ | D | ◦ | D | ◦ | ◦ | ◦ |
| CAWD429 | G. scabrosus | Japan | D | ◦ | D | D | ◦ | D | ◦ | ◦ | ◦ | ◦ | ◦ |
| CAWD429 | G. scabrosus c | Japan | D | ◦ | D | D | ◦ | D | ◦ | ◦ | ◦ | ◦ | ◦ |
| K070922_1 | G. scabrosus | Japan | D | ◦ | D | D | ◦ | D | ◦ | ◦ | ◦ | ◦ | ◦ |
| CAWD385 | C. canariensis | Kermadec Islands | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ |
| CAWD387 | C. canariensis | Kermadec Islands | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ |
| CAWD154 | C. malayensis | New Zealand | D | ◦ | D | D | ◦ | D | ◦ | ◦ | ◦ | ◦ | ◦ |
| CAWD175 | C. malayensis | New Zealand | D | ◦ | D | D | ◦ | D | ◦ | ◦ | ◦ | ◦ | ◦ |
| CAWD60 | C. monotis | Spain | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ |
| UTS4 | C. palmyrensis | Australia | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ |
| UTS25 | C. palmyrensis | Australia | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ | ◦ |
| CAWD384 | C. tropicalis | Kermadec Islands | ◦ | ◦ | D | D | ◦ | D | ◦ | ◦ | ◦ | ◦ | ◦ |
| CAWD388 | C. tropicalis | Kermadec Islands | ◦ | ◦ | D | D | ◦ | D | ◦ | ◦ | ◦ | ◦ | ◦ |
| CAWD238 | F. paulensis | New Zealand | ◦ | ◦ | D | D | ◦ | D | ◦ | ◦ | ◦ | ◦ | ◦ |
| CAWD306 | F. paulensis | New Zealand | ◦ | ◦ | D | D | ◦ | D | ◦ | ◦ | ◦ | ◦ | ◦ |
| S044 | F. ruetzleri | Hong Kong | D | ◦ | D | ◦ | ◦ | D | ◦ | ◦ | ◦ | ◦ | ◦ |
| S051 | F. ruetzleri | Hong Kong | D | ◦ | D | ◦ | ◦ | D | ◦ | ◦ | ◦ | ◦ | ◦ |
| Isolates | Scientific Name | Metabolites Detected | Acute Toxicity (LD50; mg/kg) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTXs | MTXs | G | 44-MG | Other Gs | GOx | GA A-B | GA C-D | Isolate (CAWD) | i.p. | Orally | ||
| CAWD149 and CAWD381 | G. australes | X | X | X | X | X b | 149 a | 0.37 | 40 | |||
| CCMP401 | G. belizeanus | X | X | X | X | |||||||
| CAWD301 | G. caribaeus | X | X | X | – d | |||||||
| CAWD237 c and CAWD364 | G. carpenteri | X b | X | X | X | 237 a | 10 | >158 | ||||
| CAWD232 and CAWD236 | G. cheloniae | X | X | X | X | X | 232 a | 0.32 | 118 | |||
| CAWD368 | G. holmesii | X | X | X | X | – e | ||||||
| CAWD242 and CAWD250 | G. honu | X | X | X | X | X | 242 a | 0.2 | 100 | |||
| NIES-4120 c | G. jejuensis | X | X | X | ||||||||
| CAWD336 and CAWD338 | G. lapillus | X | X | X | – e | |||||||
| CAWD369 | G. lewisii | X | X | X | X | X | – e | |||||
| CAWD227 and CAWD337 | G. pacificus | X | X | X | X | X | 227 a | 0.4 | >400 | |||
| CAWD212 and CAWD267 | G. polynesiensis | X | X | X | X | X | 212 a | 1.88 | 3.2 | |||
| CAWD429 c and K070922_1 | G. scabrosus | X | X | X | ||||||||
| CAWD385 and CAWD387 | C. canariensis | – d | ||||||||||
| CAWD154 and CAWD175 | C. malayensis | X | X | X | – d | |||||||
| CAWD60 | C. monotis | – d | ||||||||||
| UTS4 and UTS25 | C. palmyrensis | – d | ||||||||||
| CAWD384 and CAWD388 | C. tropicalis | X | X | – d | ||||||||
| CAWD238 and CAWD306 | F. paulensis | X | X | 238 a | 10 | >790 | ||||||
| S044 and S051 | F. ruetzleri | X | X | X | ||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murray, J.S.; Passfield, E.M.F.; Rhodes, L.L.; Puddick, J.; Finch, S.C.; Smith, K.F.; van Ginkel, R.; Mudge, E.M.; Nishimura, T.; Funaki, H.; et al. Targeted Metabolite Fingerprints of Thirteen Gambierdiscus, Five Coolia and Two Fukuyoa Species. Mar. Drugs 2024, 22, 119. https://doi.org/10.3390/md22030119
Murray JS, Passfield EMF, Rhodes LL, Puddick J, Finch SC, Smith KF, van Ginkel R, Mudge EM, Nishimura T, Funaki H, et al. Targeted Metabolite Fingerprints of Thirteen Gambierdiscus, Five Coolia and Two Fukuyoa Species. Marine Drugs. 2024; 22(3):119. https://doi.org/10.3390/md22030119
Chicago/Turabian StyleMurray, J. Sam, Emillie M. F. Passfield, Lesley L. Rhodes, Jonathan Puddick, Sarah C. Finch, Kirsty F. Smith, Roel van Ginkel, Elizabeth M. Mudge, Tomohiro Nishimura, Hiroshi Funaki, and et al. 2024. "Targeted Metabolite Fingerprints of Thirteen Gambierdiscus, Five Coolia and Two Fukuyoa Species" Marine Drugs 22, no. 3: 119. https://doi.org/10.3390/md22030119