Variability of Non-Polar Secondary Metabolites in the Red Alga Portieria

Abstract
:1. Introduction
2. Results and Discussion
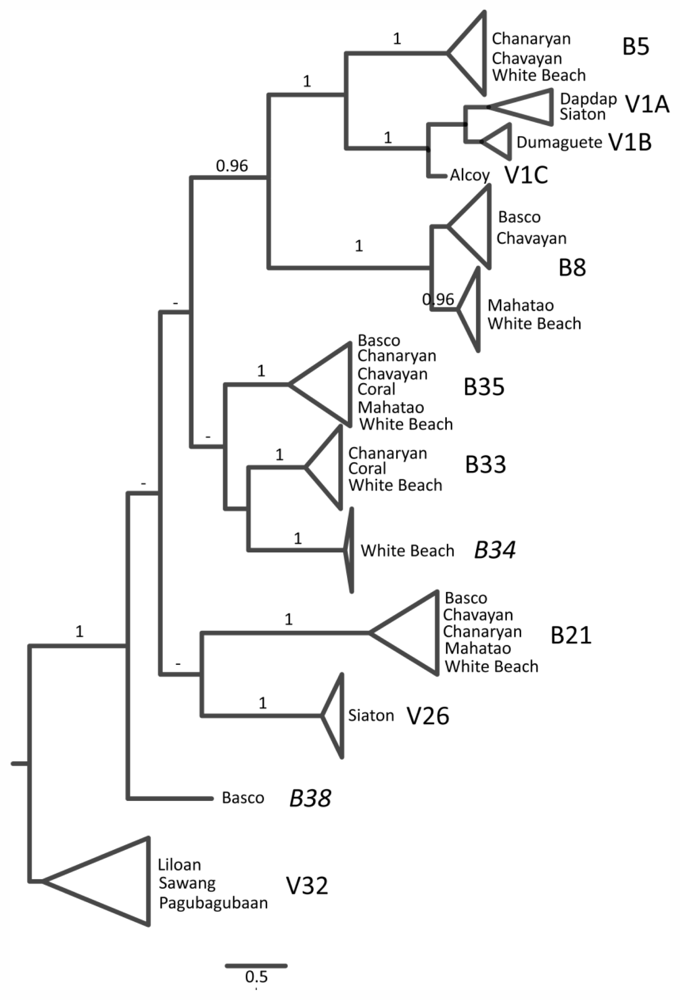
2.1. Cryptic Diversity
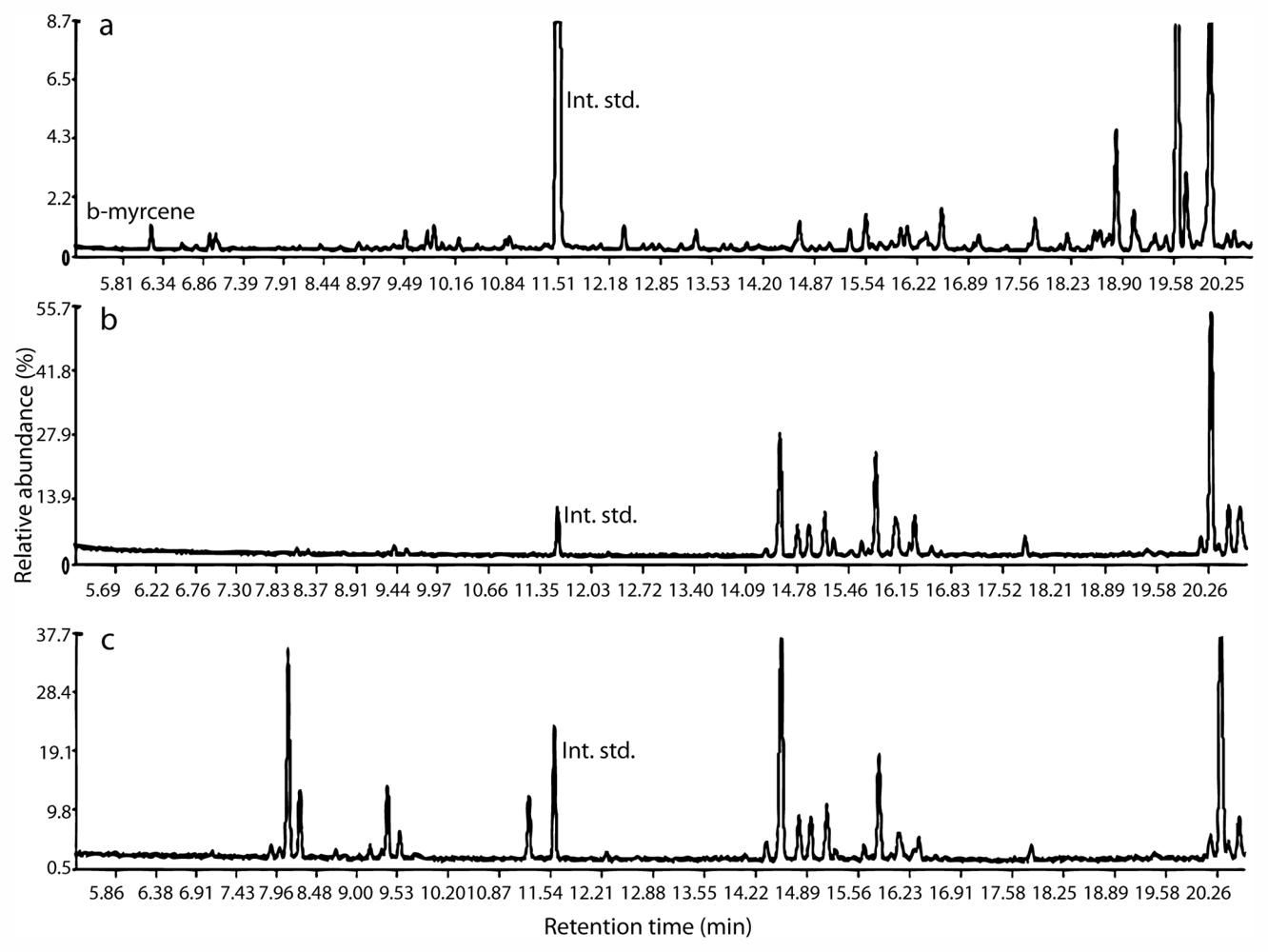
2.2. Identification of Compounds in Non-Polar Extracts
2.3. Metabolite Fingerprinting
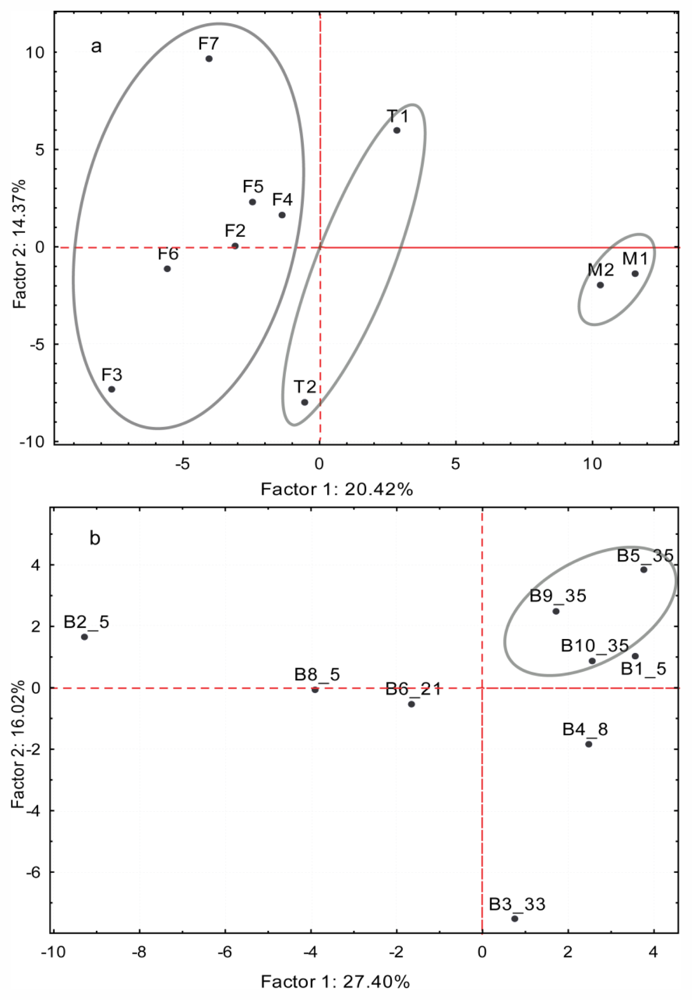
2.3.1. Variation Between Life Stages
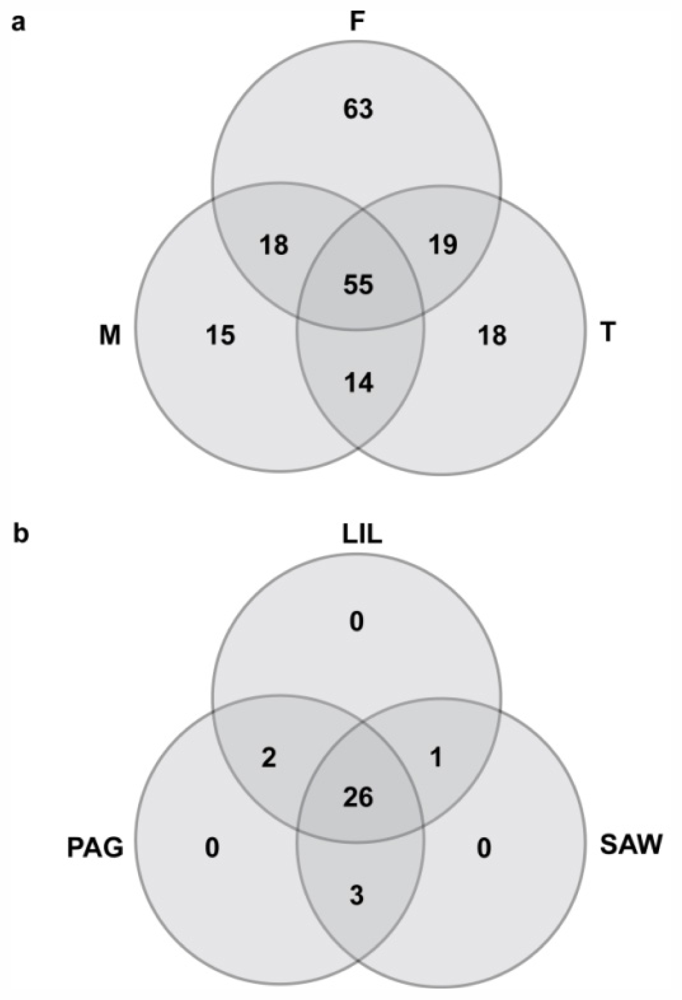
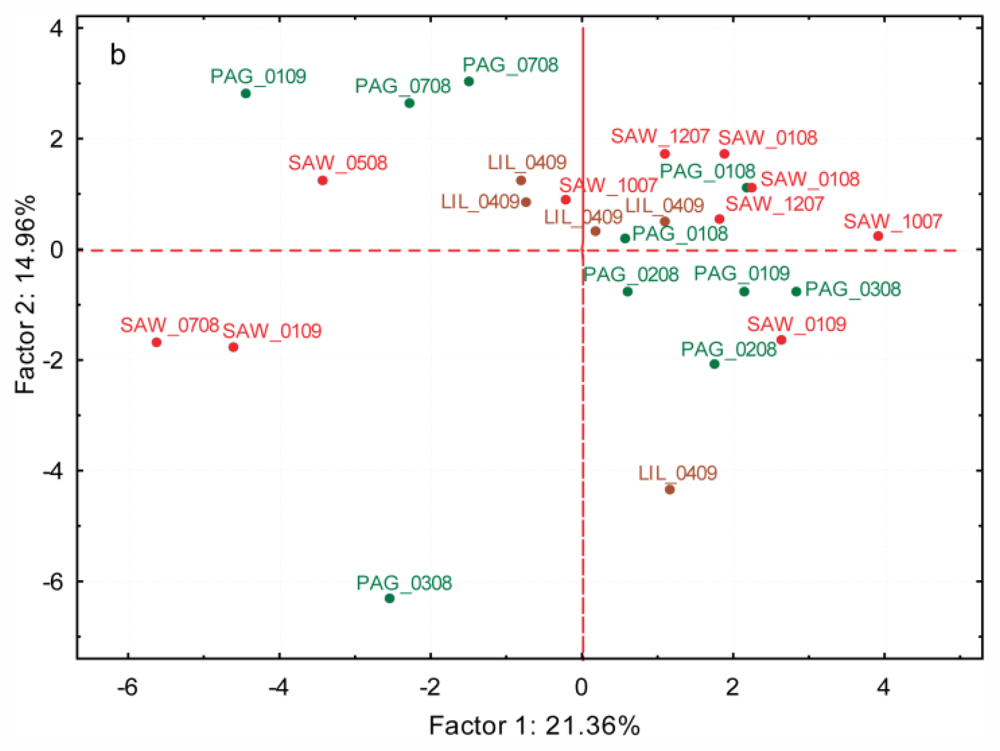
2.3.2. Variation Between Cryptic Species
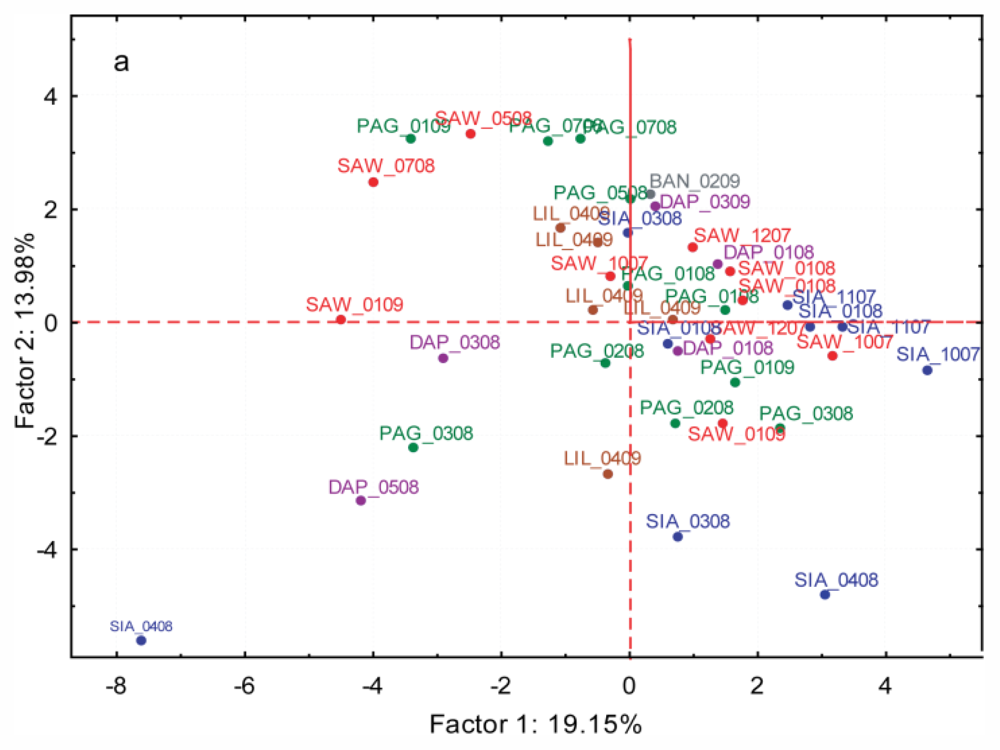
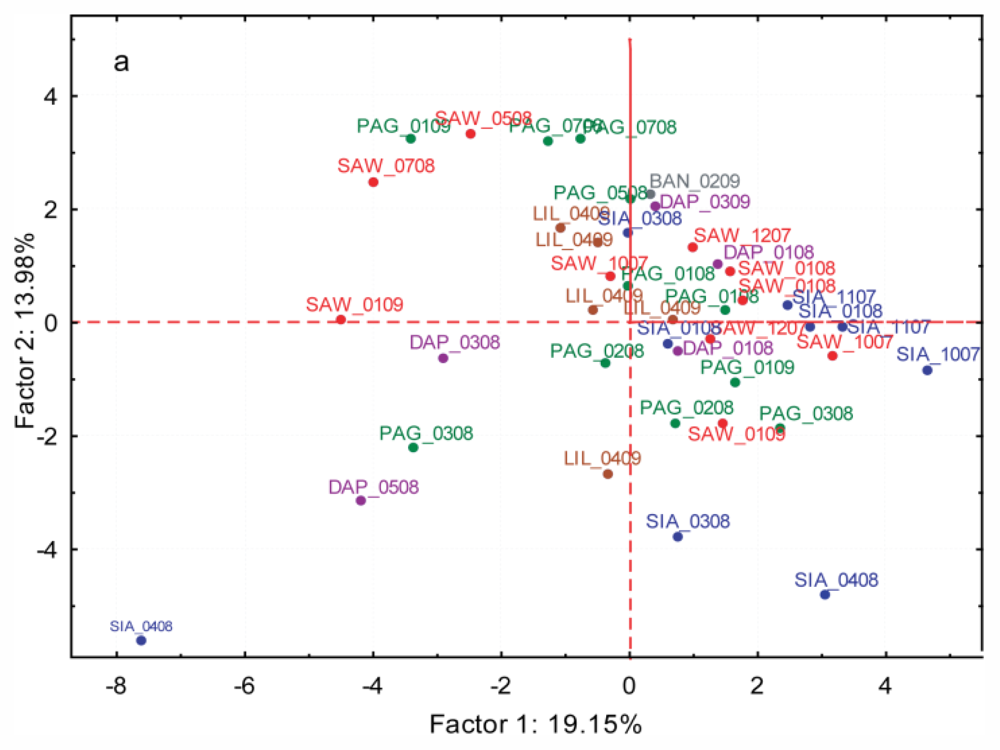
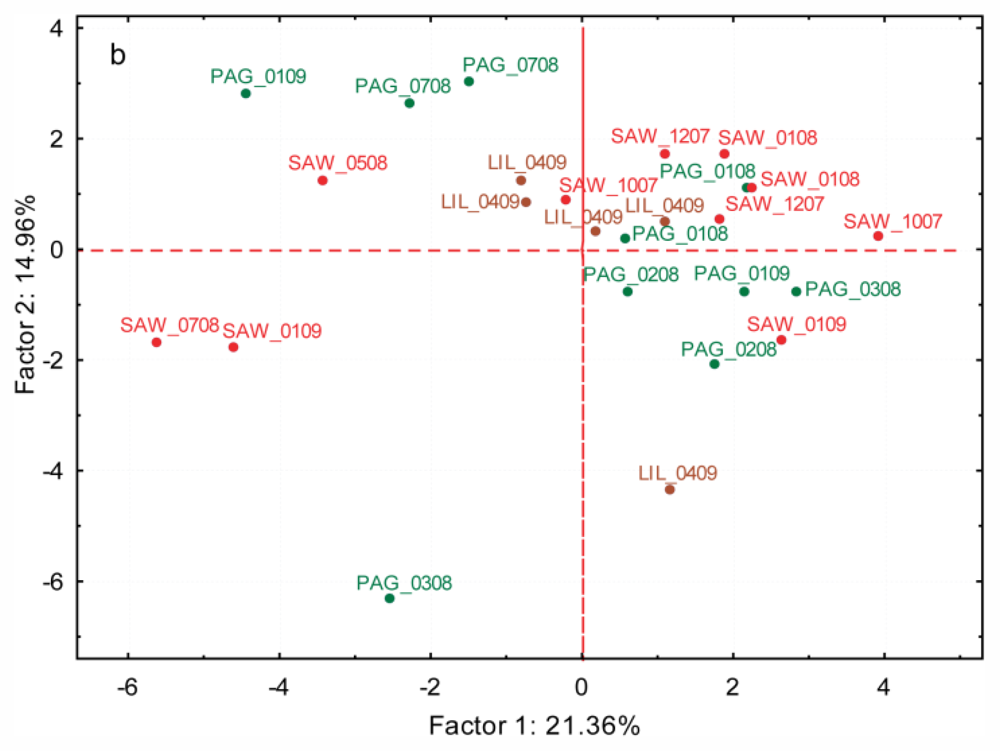
2.3.3. Spatial and Temporal Variation
2.4. Patterns of Secondary Metabolite Distribution
3. Experimental Section
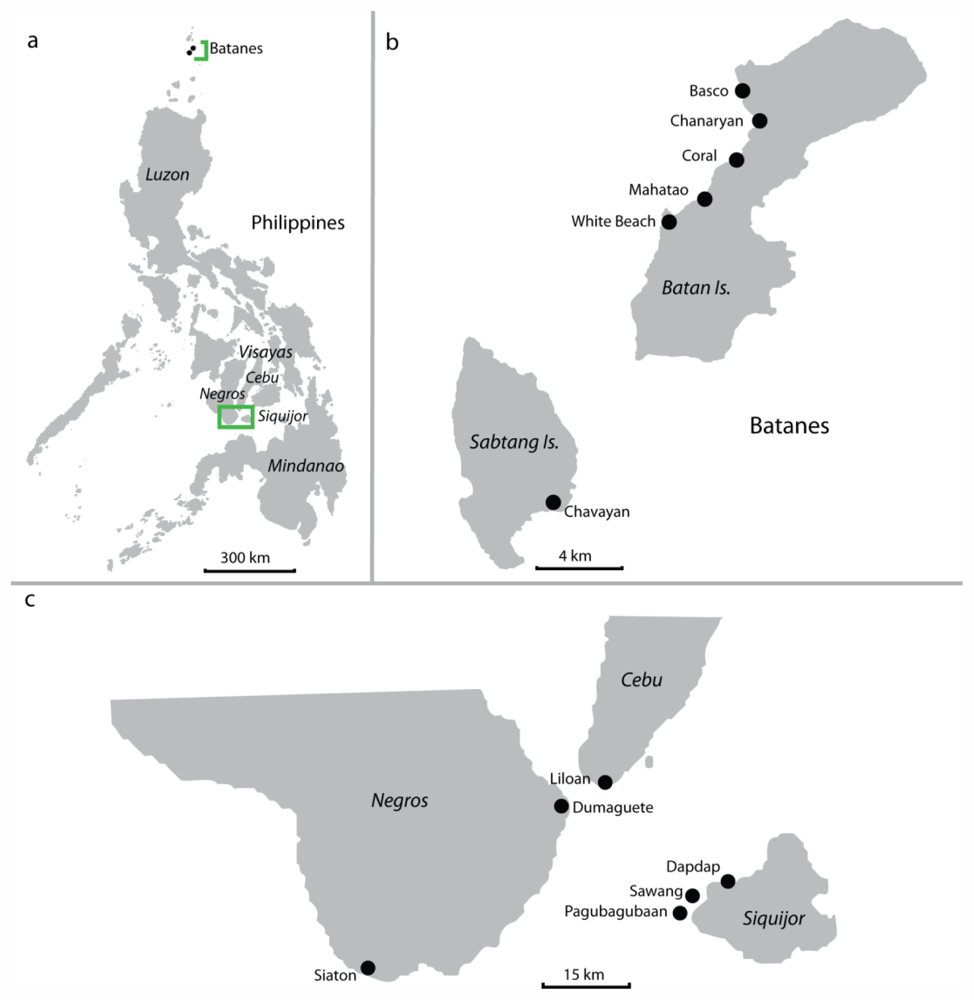
3.1. Collection and Storage
3.2. Extraction
3.2.1. Life-History Stages Comparison
3.2.2. Species Comparison
3.2.3. Seasonal and Spatial Comparison
3.3. Phylogenetic Analysis
3.4. Metabolite Analysis, Data Processing and Multivariate Analysis
3.4.1. GC-MS Analysis
3.4.2. GC-MS Data Processing
3.4.3. Compound Identification
3.4.4. Data Standardization, Metabolite Fingerprinting and Multivariate Analyses
4. Conclusions
Supplementary

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference | Cpd # | Name | Mol. Formula | Description | Specimen Source |
|---|---|---|---|---|---|
| [18] | 1 | Myrcene | C10H16 | acyclic halogenated monoterpene | Amami Is., Japan |
| 2 | 7-chloro-myrcene | C10H15Cl | " | " | |
| 3 | 7-bromo-myrcene | C10H15Br | " | " | |
| 4 | (Z)-10-bromo-myrcene | C10H15Br | " | " | |
| 5 | (E)-10-bromo-myrcene | C10H15Br | " | " | |
| 6 | (Z)-10-bromo-7-chloro-myrcene | C10H14BrCl | " | " | |
| 7 | (E)-10-bromo-7-chloro-myrcene | C10H14BrCl | " | " | |
| 8 | 3-chloro-7, (Z)-10-dibromo-myrcene | C10H13Br2Cl | " | " | |
| 9 | (Z)-10-chloro-3,7-dibromo-myrcene | C10H13Br2Cl | " | " | |
| 10 | 3-bromo-7-chloro-myrcene | C10H14BrCl | " | " | |
| 11 | 7-bromo-10-chloro-myrcene | C10H14BrCl | " | " | |
| 12 | - | C10H15Br | cyclic halogenated monoterpene | " | |
| [19] | 1 | chondrocole A | C10H14BrClO | " | Hawaii |
| 2 | chondrocole B | C10H14BrClO | " | Hawaii | |
| [46] | 3 | - | C10H15Cl3Br2 | acyclic halogenated monoterpene | Black Point, Oahu, Hawaii |
| 4 | - | C10H16Cl3Br | " | " | |
| 5 | - | C10H15Cl2Br | " | " | |
| 6 | - | C10H16Cl2Br2 | " | Halona Blowhole Is, Oahu, Hawaii | |
| 10 | - | C10H16Cl2Br2 | acyclic halogenated monoterpene | " | |
| 11 | 6-bromo-2-chloromyrcene | C10H15ClBr | " | Black Point, Oahu, Hawaii | |
| 12 | chondrocole c | C10H14Br2O | cyclic halogenated monoterpene | " | |
| 13 | - | C10H15Cl3Br2 | acyclic halogenated monoterpene | Hawaii | |
| 14 | Z-3-bromomethylene-7-methyl-1,6-octadiene | C10H16ClBr2 | " | Halona Blowhole Is., Oahu, Hawaii | |
| 15 | - | C10H14Br2Cl2 | cyclic halogenated monoterpene | " | |
| [47] | 1 | myrcene | C10H16 | acyclic halogenated monoterpene | Kada Coast, Wakayama Pref., Japan |
| 2 | 7-chloro-myrcene | C10H15Cl | " | " | |
| 3 | 7-bromo-myrcene | C10H15Br | " | " | |
| 4 | 3,7-dichloro-myrcene | C10H14Cl2 | " | " | |
| 5 | (Z)-10-bromo-3-methoxy-α-myrcene | C11H17OBr | " | " | |
| 6 | (E)-10-bromo-3-methoxy-α-myrcene | C11H17OBr | " | " | |
| 7 | 3-bromo-7-chloro-myrcene | C10H14BrCl | " | " | |
| 8 | (Z)-10-bromo-1-methoxy-myrcene | C11H17OBr | " | " | |
| 9 | (E)-10-bromo-1-methoxy-myrcene | C11H17OBr | " | " | |
| [48] | (−)-3-bromomethyl-3-chloro-7-methyl-1,6-octadiene | C10H16BrCl | " | Trincomalee (Foul Point) Sri Lanka | |
| [49] | 2-(1-chloro-2-hydroxyethyl)-4,4-dimethylcyclohexa-2, 5-dienone: a precursor of 4,5-dimethylbenzo [b] furan | - | - | - | |
| [20] | 1 | (2Z,6E)-1,8-dichloro-3-chloromethyl-7-methylocta-2,6-diene | C10H15Cl3 | acyclic halogenated monoterpene | Rib Reef, Great Barrier Reef, Australia |
| 2 | (E)-1,2-dibromo-3-chloromethylene-7-methyloct-6-ene | C10H15Br2Cl | " | " | |
| 3 | (Z)-1-chloro-3-chloromethyl-7-methylocta-2,6-diene | C10H16Cl2 | " | " | |
| 4 | (Z)-1,6-dichloro-3-chloromethyl-7-methylocta-2,7-diene | C10H15Cl3 | " | " | |
| 5 | (2R*,3(8)E,4S*,6R*)-6-bromo-2-chloro-1,4-oxido-3,(8)-ochtodene | C10H14BrClO | epimeric bicyclic monoterpene | " | |
| 6 | (2S*,3(8)E,4S*,6R*)-6-bromo-2-chloro-1,4-oxido-3,(8)-ochtodene | C10H14BrClO | " | " | |
| [50] | (Z)-3-bromo-8-chloro-6-chloromethyl-2-methylocta-1,6-diene (9,6-hydroxymethyl-2-methylocta-2,8-dien-6-ol | - | - | - | |
| [21] | 1 | (2Z)-6-bromo-3-chloromethyl-1,7-dichloro-7-methylocta-2-ene | C10H16Cl3Br | acyclic halogenated monoterpene | Nelly Bay, Magnetic Island, Queensland, Australia |
| 2 | (2Z,6E)-3-chloromethyl-1-chloroocta-2,6-dien-8-al | C10H14OCl2 | " | " | |
| 3 | 3-methoxymethyl-6-methoxyl-7-methylocta-1,7(10)-dien-3-ol | C12H22 O3 | " | " | |
| 4 | (2Z,6S)-3-chloromethyl-1-methylocta-2,7(10)-dien-6-ol | C11H19 O2Cl | " | " | |
| 5 | (2Z,6S)-3-chloromethyl-6-methylocta-2,7(10)-dien-1-ol | C11H19 O2Cl | " | " | |
| [22] | 1 | halomon/6(R)-bromo-3(S)-(bromomethyl)-7-methyl-2,3,7-trichloro-1-octene | C10H15Br2Cl3 | acyclic halogenated monoterpene | Chanaryan, Batan Is., Batanes, Philippines |
| 2 | - | C10H13Cl | cyclic halogenated monoterpene | Banilad, Bacong, Negros Oriental, Philippines | |
| [23] | 2 | isohalomon | C10H15Br2Cl3 | acyclic halogenated monoterpene; isomeric with halomon | Chanaryan, Batan Is., Batanes, Philippines |
| 3 | - | C10H14BrCl3 | acyclic halogenated monoterpene | " | |
| 4 | - | C10H14Br2Cl2 | " | " | |
| 5 | - | C10H14Br3Cl | cyclic halogenated monoterpene | " | |
| 6 | - | C10H14BrCl3 | " | " | |
| 7 | - | C10H13Cl3 | " | " | |
| 8 | - | C10H15BrCl2 | acyclic halogenated monoterpene | " | |
| 9 | - | C10H16Br2Cl2 | " | " | |
| 10 | - | C10H16BrCl | " | Halona Blowhole Is, Oahu, Hawaii | |
| 11 | - | C10H15BrCl | " | " | |
| 12 | - | C10H14BrCl | cyclic halogenated monoterpene | " | |
| [8] | 1 | apakaochtodene A/6(S*)-bromo-1,4(S*),8(R*)-trichloro-2(Z)-ochtodene | C10H14Cl3Br | cyclic halogenated monoterpene | Apaka Point Beach, Guam |
| 2 | apakaochtodene B/6(S)-bromo-1,4(S), 8(R*)-trichloro-2(E)-ochtodene | C10H14Cl3Br | cyclic halogenated monoterpene | Gun Beach; Double Reef; Pago Bay; Guam | |
| [11] | 1 | myrcene/7-methyl-3-methylene-1,6-octadiene | C10 H16O | acyclic non-halogenated monoterpene | microplantlet culture, Double Reef NW Guam |
| 2 | 10E-bromomyrcene/E-3-bromomethylene-7-methyl-1,6-octadiene | C10H15Br | " | " | |
| 3 | 10Z-bromomyrcene/Z-3-bromomethylene-7-methyl-1,6-octadiene | C10H15Br | " | " | |
| 4 | 10E-bromo-3-chloro-α-myrcene/E-3-bromomethylene-6-chloro-1,7-octadiene | C10H14BrCl | " | " | |
| 5 | - | C10H14Br2 | " | " | |
| 6 | apakaochtodene B/6(S)-bromo-1,4(S), 8(R*)-trichloro-2(E)-ochtodene | C10H14BrCl3 | cyclic halogenated monoterpene | " | |
| 7 | - | C10 H16O | non-halogenated monoterpene | " | |
| 8 | bromomyrcene isomer | C10H15Br | halogenated monoterpene | " | |
| 9 | 7-chloromyrcene/2-chloro-3-methylene-7-methyl-1,6-octadiene | C10H15Cl | " | " | |
| 10 | C10H14Cl | " | " | ||
| 11 | chloromyrcene derivative | C10H16Cl | " | " | |
| 12 | bromomyrcene isomer | C10H15Br | " | " | |
| 13 | chloromyrcene derivative | C10H14Cl | " | " | |
| 14 | - | C10H18Cl2 | " | " | |
| 15 | - | C15H18 O4 | sesquiterpene | " | |
| [13] | 5 | 1,2-dibromoochtoda-3(8),5-dien-4-one | C22H12Br2O | cyclic halogenated monoterpene | Cape Zampa, Okinawa, Japan |
| 6 | 1-bromo-2-chloroochtoda-3(8),5-dien-4-one | C10H12BrClO | " | " | |
| 7 | 1,2-dichloroochtoda-3(8),5-dien-4-one | C10H12Cl2O | " | Gushichan coast, Okinawa, Japan | |
| 8 | (1Z)-1-bromoochtoda-1,3(8),5-trien-4-one | C10H11BrO | " | Cape Zampa, Okinawa, Japan | |
| 9 | (1Z)-1-chloroochtoda-1,3(8),5-trien-4-one | C10H11ClO | " | " | |
| [12] | 1 | halomon/6(R)-bromo-3(S)-(bromomethyl)-7-methyl-2,3,7-trichloro-1-octene | C10H15Br2Cl3 | acyclic halogenated monoterpene | Tolagniaro, Fort Dauphin, Madagascar |
| 2 | - | C10H13ClBr | " | " | |
| 3 | - | C10H14Cl2Br | " | " | |
| 4 | C10H15Br2Cl3 | " | " |
| # | Sample | Base Peak | KI | KI Based ID | Mass Spectra Based ID | |||
|---|---|---|---|---|---|---|---|---|
| RT (min) | Nearest KI | Lucero et al. [37] | NIST | MassBase | ||||
| 1 | F6_2 | 93 | 6.1863 | 994 | 993/994 | beta-myrcene/6-methyl-5-hepten-2-ol | b-myrcene | b-myrcene |
| 2 | T1_1 | 104 | 6.5874 | 1008 | 1007 | α-phellandrene | - | - |
| 3 | T1_2 | 104 | 6.5909 | 1008 | 1007 | α-phellandrene | - | - |
| 4 | M1_1 | 91 | 6.7685 | 1016 | 1017 | α-terpinene | Benzene, tert-butyl- | 4-methylacetophenone |
| 5 | F3_3 | 79 | 6.9501 | 1023 | 1024 | r-cymene | - | - |
| 6 | M1_2 | 79 | 6.9532 | 1023 | 1024 | r-cymene | - | - |
| 7 | M1_3 | 91 | 7.017 | 1026 | 1025 | p-cymene | carbonic acid | 1-phenylpropan-2-one |
| 8 | M2_2 | 91 | 7.021 | 1026 | 1025 | p-cymene | - | protopine |
| 9 | F3_2 | 91 | 7.0387 | 1027 | 1025 | p-cymene | - | N-Methyl-Npropagylbenzylamine |
| 10 | T2_1 | 57 | 8.0381 | 1063 | 1062 | y-terpinene | - | - |
| 11 | M2_2 | 132 | 8.8894 | 1089 | 1089 | p-mentha-2,4(8)diene | - | - |
| 12 | T2_1 | 117 | 9.5127 | 1110 | 1099 | linalool | - | - |
| 13 | F4_3 | 117 | 9.8874 | 1124 | 1123 | chrysanthenone | - | - |
| 14 | M1_3 | 117 | 9.9855 | 1128 | 1127 | α-campholenal | - | - |
| 15 | M2_2 | 91 | 10.2042 | 1136 | 1127 | α-campholenal | - | - |
| 16 | F2_2 | 91 | 10.2132 | 1136 | 1127 | α-campholenal | - | Dimethirimol |
| 17 | T2_1 | 119 | 10.4565 | 1145 | 1145 | camphor | - | Benzimidazole |
| 18 | M1_1 | 68 | 10.8826 | 1160 | 1158 | isobomeol | - | - |
| 19 | F7_3 | 68 | 10.8845 | 1160 | 1158 | isobomeol | - | - |
| 20 | T2_1 | 68 | 10.8866 | 1160 | 1158 | isobomeol | - | - |
| 21 | F3_3 | 68 | 10.8873 | 1160 | 1158 | isobomeol | - | - |
| 22 | F4_1 | 117 | 11.3242 | 1174 | 1171 | ethyl-benzoate | - | - |
| 23 | F4_1 | 50 | 11.5177 | 1180 | 1180 | m-cymen-8-ol | - | - |
| 24 | T2_2 | 127 | 11.5317 | 1180 | 1180 | m-cymen-8-ol | - | - |
| 25 | T2_3 | 134 | 11.7619 | 1188 | 1187 | p-cymen-8-ol | - | - |
| 26 | M2_2 | 105 | 12.0654 | 1197 | 1196 | methylchavicol | - | 3-cyanopyridine |
| 27 | F3_2 | 57 | 12.1827 | 1200 | 1203 | n-decanal | 3-Hexanone | - |
| 28 | F7_3 | 113 | 12.3674 | 1207 | 1207 | verbenone | - | - |
| 29 | M1_2 | 113 | 12.3831 | 1208 | 1207 | verbenone | - | - |
| 30 | T1_3 | 91 | 12.3874 | 1208 | 1207 | verbenone | - | - |
| 31 | M2_2 | 93 | 12.6294 | 1217 | 1217 | trans-carveol | - | - |
| 32 | M1_2 | 69 | 12.7588 | 1222 | 1219 | trans-carveol | - | - |
| 33 | M1_1 | 55 | 12.8397 | 1225 | 1229 | nerol | - | - |
| 34 | F7_3 | 67 | 13.1203 | 1236 | 1236 | thymol methyl ether | - | - |
| 35 | M1_1 | 79 | 13.1266 | 1236 | 1236 | thymol methyl ether | - | - |
| 36 | F5_2 | 93 | 13.127 | 1236 | 1236 | thymol methyl ether | - | Beta-pinene; y-eudesmol |
| 37 | M2_2 | 93 | 13.32 | 1243 | 1243 | carvone | - | - |
| 38 | T1_1 | 93 | 13.327 | 1243 | 1243 | carvone | 2-chloropropionyl chloride | - |
| 39 | T2_1 | 93 | 13.3305 | 1243 | 1243 | carvone | - | - |
| 40 *(Br) | F6_2 | 69 | 13.6916 | 1256 | 1256 | geraniol | - | - |
| 41 | M1_1 | 91 | 13.6953 | 1256 | 1256 | geraniol | - | |
| 42 | F2_1 | 69 | 13.6967 | 1256 | 1256 | geraniol | - | |
| 43 | F5_1 | 69 | 13.6975 | 1256 | 1256 | geraniol | - | - |
| 44 | M2_1 | 91 | 13.9837 | 1266 | 1263 | (E)-2-decenal | - | N-Methyl-Npropagylbenzylamine |
| 45 | F7_3 | 119 | 14.131 | 1271 | 1271 | geranial | - | - |
| 46 | F2_3 | 119 | 14.1354 | 1271 | 1271 | geranial | - | - |
| 47 | F3_2 | 69 | 14.4252 | 1281 | 1282 | a-terpinen-7-al | - | - |
| 48 | M2_3 | 69 | 14.4368 | 1281 | 1282 | a-terpinen-7-al | - | - |
| 49 | F5_1 | 91 | 14.6745 | 1289 | 1289 | p-cymen-7-ol | - | - |
| 50 | T2_1 | 91 | 14.68 | 1289 | 1289 | p-cymen-7-ol | - | - |
| 51 | F6_3 | 149 | 14.9157 | 1297 | 1297 | perilla alcohol | - | Benzoylcholine |
| 52 * (Cl) | F6_3 | 131 | 15.0641 | 1302 | 1302 | trans-ascaridole | - | 2-chloro-1-phenyl-2-butene |
| 53 | F6_1 | 166 | 15.3344 | 1313 | 1313 | 2E,4E-decadienal | - | 2-chloro-1-phenyl-2-butene |
| 54 | F6_3 | 81 | 15.5368 | 1320 | 1313 | 2E,4E-decadienal | - | - |
| 55 | M2_2 | 81 | 15.5381 | 1321 | 1313 | 2E,4E-decadienal | - | - |
| 56 | T1_2 | 81 | 15.5481 | 1321 | 1313 | 2E,4E-decadienal | - | - |
| 57 | M2_2 | 67 | 15.6269 | 1324 | 1313 | 2E,4E-decadienal | - | 1-Adamantanamine |
| 58 | M1_2 | 67 | 15.6354 | 1324 | 1313 | 2E,4E-decadienal | - | - |
| 59 | F3_1 | 57 | 15.7447 | 1329 | 1339 | d-elemene | - | - |
| 60 | F2_1 | 57 | 15.7649 | 1329 | 1339 | d-elemene | - | - |
| 61 | M2_2 | 91 | 15.8827 | 1334 | 1339 | d-elemene | - | - |
| 62 | M1_2 | 91 | 15.8885 | 1334 | 1339 | d-elemene | - | - |
| 63 | M2_2 | 139 | 16.5157 | 1357 | 1357 | eugenol | - | - |
| 64 | F6_1 | 115 | 16.5278 | 1358 | 1357 | eugenol | - | - |
| 65 | M2_3 | 166 | 16.5392 | 1358 | 1357 | eugenol | - | - |
| 66 | M2_2 | 91 | 17.02 | 1375 | 1373 | a-ylangene | - | - |
| 67 | M2_3 | 91 | 17.0254 | 1376 | 1373 | a-ylangene | - | - |
| 68 | M1_2 | 91 | 17.0311 | 1376 | 1373 | a-ylangene | - | - |
| 69 | T2_1 | 127 | 17.403 | 1389 | 1389 | isolongifolene | - | Imidazole-4-acetate |
| 70 | T2_1 | 57 | 17.7476 | 1400 | 1399 | 1,7-di-epi-a-cedrene | - | - |
| 71 | M2_2 | 79 | 17.7625 | 1401 | 1402 | methyleugenol | - | - |
| 72 | M2_3 | 127 | 17.7782 | 1402 | 1402 | methyleugenol | - | - |
| 73 | T2_2 | 67 | 18.1041 | 1415 | 1415 | cis-a-bergarnotene | - | - |
| 74 | T1_3 | 79 | 18.18 | 1418 | 1419 | β-caryophyllene | - | - |
| 75 | F6_1 | 79 | 18.1866 | 1419 | 1419 | β-caryophyllene | - | - |
| 76 | T1_1 | 79 | 18.1928 | 1419 | 1419 | β-caryophyllene | Pyridine | - |
| 77 | F5_1 | 79 | 18.1934 | 1419 | 1419 | β-caryophyllene | - | camphene |
| 78 | M1_2 | 79 | 18.1939 | 1419 | 1419 | β-caryophyllene | - | - |
| 79 * (Br) | F7_2 | 212 | 18.3193 | 1424 | 1420 | β-caroyophyllene | - | - |
| 80 | F6_2 | 210 | 18.4564 | 1429 | 1420 | β-caroyophyllene | - | - |
| 81 | M2_3 | 131 | 18.4673 | 1430 | 1420 | β-caroyophyllene | - | - |
| 82 | M1_1 | 67 | 18.5462 | 1433 | 1435 | trans-a-bergamotene | - | - |
| 83 | M1_2 | 67 | 18.5503 | 1433 | 1435 | trans-a-bergamotene | - | - |
| 84 | F2_2 | 133 | 18.6084 | 1435 | 1435 | trans-a-bergamotene | - | - |
| 85 | T2_2 | 69 | 18.6925 | 1439 | 1438 | trans-a-bergamotene | - | - |
| 86 | F3_3 | 69 | 18.6982 | 1439 | 1440 | a-guaiene | - | - |
| 87 | M2_3 | 69 | 18.7018 | 1439 | 1440 | a-guaiene | - | - |
| 88 | M1_1 | 91 | 18.7475 | 1441 | 1440 | a-guaiene | - | - |
| 89 | F4_2 | 69 | 18.8243 | 1444 | 1440 | a-guaiene | - | - |
| 90 | F7_2 | 69 | 18.8273 | 1444 | 1440 | a-guaiene | - | - |
| 91 | T1_3 | 69 | 18.8306 | 1444 | 1440 | a-guaiene | - | - |
| 92 | F2_2 | 210 | 19.1226 | 1456 | 1455 | a-humulene | - | - |
| 93 | F3_3 | 133 | 19.3339 | 1464 | 1464 | a-acoradiene | - | |
| 94 | T2_2 | 105 | 19.4843 | 1469 | 1469 | drima-7,9(11)-diene | - | - |
| 95 | T2_1 | 105 | 19.4852 | 1469 | 1469 | drima-7,9(11)-diene | - | - |
| 96 | F7_2 | 131 | 19.4915 | 1470 | 1469 | drima-7,9(11)-diene | - | - |
| 97 | F5_1 | 91 | 19.6255 | 1475 | 1474 | b-cadinene | - | - |
| 98 | F6_3 | 91 | 19.7468 | 1479 | 1479 | y-curcurnene | - | |
| 99 | F2_1 | 91 | 19.7581 | 1480 | 1480 | germacrene D | - | - |
| 100 | F4_1 | 153 | 20.0567 | 1491 | 1491 | cis-β-guaiene | - | - |
| 101 | F4_1 | 153 | 20.0567 | 1491 | 1491 | cis-β-guaiene | - | - |
| 102 | F3_2 | 71 | 20.2111 | 1496 | 1495 | a-zingiberene | - | |
| 103 | F7_2 | 135 | 20.3621 | 1502 | 1499 | a-muurolene | - | - |
| 104 | F5_1 | 91 | 20.3727 | 1503 | 1506 | d-selinene | - | - |
| 105 | T2_1 | 91 | 20.3766 | 1503 | 1506 | d-selinene | - | - |
| 106 | F2_2 | 91 | 20.3829 | 1503 | 1506 | d-selinene | -- | - |
| 107 | F6_1 | 205 | 20.6221 | 1513 | 1513 | g-cadinene | - | - |
| 108 | F4_3 | 205 | 20.6237 | 1513 | 1513 | g-cadinene | - | - |
| 109 | F7_2 | 205 | 20.6252 | 1513 | 1513 | g-cadinene | - | 5-bromo-4-methoxyoct-1-ene |
| 110 | T2_1 | 205 | 20.6278 | 1513 | 1513 | g-cadinene | p-Benzoquinone | - |
| 111 | F3_2 | 133 | 20.6781 | 1516 | 1514 | sesquicineole | - | - |
| 112 | F3_3 | 133 | 20.6803 | 1516 | 1514 | sesquicineole | - | - |
| 113 | F6_3 | 133 | 20.685 | 1516 | 1514 | sesquicineole | - | - |
| 114 | F5_1 | 69 | 20.8293 | 1522 | 1523 | d-cadinene | - | -- |
| 115 | F5_1 | 69 | 20.9154 | 1526 | 1525 | eugenyl acetate | - | - |
| 116 | F6_3 | 69 | 20.9158 | 1526 | 1525 | eugenyl acetate | - | - |
| 117 | F7_2 | 121 | 20.9204 | 1526 | 1525 | eugenyl acetate | - | - |
| 118 | T1_3 | 69 | 20.9227 | 1526 | 1525 | eugenyl acetate | - | - |
| 119 | F7_2 | 167 | 21.1103 | 1534 | 1533 | cadina-1,4-diene1 | - | - |
| 120 | T2_2 | 91 | 21.1498 | 1536 | 1538 | a-cadinene | - | - |
| 121 | F6_1 | 167 | 21.43 | 1547 | 1548 | elemol | - | - |
| 122 | F7_2 | 167 | 21.4346 | 1547 | 1548 | elemol | - | - |
| 123 | T1_3 | 69 | 21.467 | 1549 | 1549 | elemol | - | -- |
| 124 | F3_2 | 69 | 21.4709 | 1549 | 1549 | elemol | - | - |
| 125 | F7_2 | 91 | 21.598 | 1554 | 1552 | elemicin | - | - |
| 126 | F6_2 | 91 | 21.6852 | 1557 | 1557 | germacrene B | - | - |
| 127 | F5_1 | 91 | 21.6859 | 1558 | 1557 | germacrene B | - | - |
| 128 | F6_3 | 91 | 21.6869 | 1558 | 1557 | germacrene B | - | - |
| 129 | M1_1 | 91 | 21.6895 | 1558 | 1557 | germacrene B | - | - |
| 130 | F6_3 | 67 | 21.7268 | 1559 | 1557 | germacrene B | - | - |
| 131 | F7_2 | 133 | 21.8253 | 1563 | 1564 | β-calacorene | - | l-Asparagine |
| 132 | T1_1 | 91 | 21.8405 | 1564 | 1564 | β-calacorene | - | - |
| 133 | F7_2 | 167 | 22.0012 | 1570 | 1574 | prenopsan-8-ol | - | - |
| 134 | T1_3 | 213 | 22.018 | 1571 | 1574 | prenopsan-8-ol | - | - |
| 135 | F6_3 | 132 | 22.0226 | 1571 | 1574 | prenopsan-8-ol | - | - |
| 136 | F6_3 | 153 | 22.0226 | 1571 | 1574 | prenopsan-8-ol | - | - |
| 137 | M1_1 | 91 | 22.3956 | 1586 | 1585 | gleenol | - | - |
| 138 | F2_2 | 133 | 22.5362 | 1591 | 1590 | viridflorol | - | - |
| 139 | F6_3 | 71 | 22.7179 | 1598 | 1590 | viridflorol | - | - |
| 140 | F3_2 | 71 | 22.7193 | 1598 | 1590 | viridflorol | - | - |
| 141 | F2_3 | 135 | 22.722 | 1598 | 1590 | viridflorol | - | - |
| 142 | F3_3 | 57 | 22.7547 | 1600 | 1607 | b-oplopenone | - | - |
| 143 | F3_2 | 57 | 22.7603 | 1600 | 1607 | b-oplopenone | - | - |
| 144 | F3_3 | 57 | 22.761 | 1600 | 1607 | b-oplopenone | - | - |
| 145 | F6_3 | 67 | 22.8782 | 1605 | 1607 | b-oplopenone | - | - |
| 146 | T2_2 | 68 | 23.0541 | 1613 | 1612 | tetradecanal | - | - |
| 147 | F4_3 | 148 | 23.2435 | 1622 | 1623 | silphiperfol-6-en-5-one | - | - |
| 148 | F3_1 | 135 | 23.2485 | 1622 | 1623 | silphiperfol-6-en-5-one | - | - |
| 149 | F5_3 | 91 | 23.52 | 1634 | 1633 | y-eudesmol | - | - |
| 150 | F4_3 | 91 | 23.5226 | 1634 | 1633 | y-eudesmol | - | - |
| 151 | F7_2 | 68 | 23.5261 | 1634 | 1633 | y-eudesmol | - | - |
| 152 | F7_2 | 68 | 23.5261 | 1634 | 1633 | y-eudesmol | - | -- |
| 153 | T2_2 | 68 | 23.5278 | 1634 | 1633 | y-eudesmol | - | - |
| 154 | F6_1 | 167 | 23.5309 | 1634 | 1633 | y-eudesmol | - | |
| 155 | F6_2 | 67 | 23.7239 | 1643 | 1642 | cubenol | - | allylcy-anide |
| 156 | T1_3 | 91 | 23.8325 | 1647 | 1646 | a-muurolol | - | - |
| 157 | F7_2 | 132 | 23.8759 | 1649 | 1646 | a-muurolol | - | - |
| 158 | F7_2 | 132 | 23.9988 | 1654 | 1653 | a-cadinol | - | - |
| 159 | F6_2 | 149 | 24.3377 | 1669 | 1668 | bulnesol | - | - |
| 160 | F7_2 | 149 | 24.3428 | 1669 | 1668 | bulnesol | - | - |
| 161 | F6_3 | 167 | 24.6091 | 1680 | 1682 | a-bisabolol | - | - |
| 162 | F6_1 | 57 | 24.6252 | 1681 | 1682 | a-bisabolol | - | - |
| 163 | F7_2 | 69 | 24.8113 | 1689 | 1686 | 8-cedren-13-ol | - | - |
| 164 | F7_2 | 69 | 24.8207 | 1689 | 1686 | 8-cedren-13-ol | - | - |
| 165 | F2_3 | 67 | 24.9282 | 1693 | 1686 | 8-cedren-13-ol | - | - |
| 166 | F6_1 | 57 | 25.0854 | 1700 | 1686 | 8-cedren-13-ol | - | Heptadecane |
| 167 | F7_2 | 91 | 25.3006 | 1710 | 1686 | 8-cedren-13-ol | - | - |
| 168 | F3_2 | 71 | 25.3224 | 1711 | 1735 | oplopanone | - | - |
| 169 | F3_3 | 68 | 25.4814 | 1718 | 1735 | oplopanone | - | - |
| 170 | F6_3 | 67 | 25.7278 | 1730 | 1735 | oplopanone | - | - |
| 171 | F7_2 | 103 | 25.7292 | 1730 | 1735 | oplopanone | - | - |
| 172 | F2_1 | 67 | 25.7318 | 1730 | 1735 | oplopanone | - | - |
| 173 | F2_3 | 133 | 25.8305 | 1735 | 1735 | oplopanone | - | - |
| 174 | F3_1 | 117 | 25.8366 | 1735 | 1735 | oplopanone | - | - |
| 175 | F7_2 | 167 | 26.3223 | 1757 | 1761 | benzyl-benzoate | - | - |
| 176 | T2_1 | 167 | 26.5838 | 1769 | 1761 | benzyl-benzoate | - | 6-Methylmer-captopurine |
| 177 | F6_2 | 117 | 26.7333 | 1775 | 1789 | 8-a-acetoxyelemol | - | - |
| 178 | T2_2 | 103 | 26.8111 | 1779 | 1789 | 8-a-acetoxyelemol | - | - |
| 179 | F3_1 | 67 | 26.8577 | 1781 | 1789 | 8-a-acetoxyelemol | - | - |
| 180 | F6_1 | 67 | 26.8599 | 1781 | 1789 | 8-a-acetoxyelemol | - | - |
| 181 | F3_2 | 67 | 26.8671 | 1781 | 1789 | 8-a-acetoxyelemol | - | - |
| 182 | T2_1 | 67 | 26.8697 | 1781 | 1789 | 8-a-acetoxyelemol | - | - |
| 183 * (Cl2) | F6_1 | 103 | 26.8838 | 1782 | 1789 | 8-a-acetoxyelemol | - | - |
| 184 | F3_1 | 103 | 26.8852 | 1782 | 1789 | 8-a-acetoxyelemol | - | - |
| 185 * (Br) | F6_1 | 67 | 27.4286 | 1806 | 1798 | nootkatone | - | - |
| 186 * (Br) | F7_2 | 67 | 27.4342 | 1806 | 1798 | nootkatone | - | - |
| 187 | F2_1 | 67 | 27.4383 | 1807 | 1798 | nootkatone | - | - |
| 188 | F6_3 | 103 | 27.4961 | 1810 | 1798 | nootkatone | - | - |
| 189 | F6_1 | 67 | 27.5012 | 1810 | 1798 | nootkatone | - | - |
| 190 | F6_2 | 67 | 27.5676 | 1813 | 1827 | isopropyl tetradecanoate | - | - |
| 191 | F6_2 | 69 | 28.0351 | 1836 | 1827 | isopropyl tetradecanoate | - | - |
| 192 | F3_2 | 69 | 28.0387 | 1836 | 1827 | isopropyl tetradecanoate | - | - |
| 193 | F6_1 | 91 | 28.3153 | 1849 | 1867 | flourensiadiol | - | - |
| 194 | F3_2 | 149 | 28.735 | 1869 | 1867 | flourensiadiol | - | - |
| 195 * (ClBr) | F6_2 | 69 | 29.2918 | 1895 | 1878 | hexadecanol | - | - |
| 196 | F5_3 | 67 | 29.4766 | 1903 | 1927 | methyl hexadecanoate | - | - |
| 197 | T2_1 | 67 | 29.4889 | 1904 | 1927 | methyl hexadecanoate | - | - |
| 198 | F7_2 | 91 | 30.7705 | 1968 | 1999 | eicosane | - | - |
| 199 | T2_2 | 91 | 30.7753 | 1969 | 1999 | eicosane | - | - |
| 200 | F3_2 | 91 | 30.7855 | 1969 | 1999 | eicosane | - | - |
| 201 | T2_2 | 64 | 31.6475 | - | - | - | - | - |
| 202 | T2_2 | 129 | 38.661 | - | - | - | - | - |
| # | Sample | Base Peak | KI | KI Based ID | Mass Spectra Based ID | |||
|---|---|---|---|---|---|---|---|---|
| RT (min) | Nearest KI | Lucero et al. [37] | NIST | Massbase | ||||
| 1 | B4B | 57 | 8.1122 | 1065 | 1062 | y-terpinene | - | - |
| 2 | B4C | 69 | 8.7386 | 1085 | 1085 | artemisia-alcohol | - | - |
| 3 | B4C | 57 | 9.1807 | 1098 | 1097 | linalool | - | - |
| 4 | B4B | 71 | 9.1841 | 1098 | 1097 | linalool | - | - |
| 5 | B6C | 57 | 9.4047 | 1106 | 1099 | linalool | 3-hexanone, 2,2-dimethyl- | 2-methylbutane |
| 6 | B4C | 69 | 11.2515 | 1172 | 1171 | ethyl-benzoate | - | - |
| 7- * (Cl) | B3A | 69 | 11.2560 | 1172 | 1171 | ethyl-benzoate | - | - |
| 8 | B4A | 57 | 14.3547 | 1279 | 1277 | trans-carvone-oxide | - | 2,2-dimethylbutane |
| 9 | B4B | 57 | 14.3593 | 1279 | 1277 | trans-carvone-oxide | - | 3-ethylhexane |
| 10 | B3B | 57 | 14.5560 | 1285 | 1286 | borneol-acetate | - | |
| 11 | B3A | 69 | 15.2678 | 1310 | 1306 | undecanal | - | trans-4-octene |
| 12 | B3A | 57 | 15.6497 | 1325 | 1314 | 2E,4E-decadienal | - | |
| 13 | B6B | 57 | 15.6518 | 1325 | 1314 | 2E,4E-decadienal | - | |
| 14 | B4A | 71 | 16.5884 | 1360 | 1357 | eugenol | - | 2-methylpentane |
| 15 | B6C | 57 | 17.8154 | 1403 | 1403 | italicene | - | 2,2-dimethylbutane |
| 16 | B3A | 57 | 17.8289 | 1404 | 1403 | italicene | - | |
| 17 | B10 | 71 | 20.306 | 1500 | 1499 | a-muurolene | - | Pentacosane |
| 18 | B3A | 57 | 20.4025 | 1504 | 1506 | d-selinene | - | tripropylamine |
| 19 | B4A | 191 | 20.715 | 1517 | 1514 | sesquicineole | - | |
| 20 | B6C | 57 | 21.0008 | 1529 | 1532 | cadina-1,4-diene | - | |
| 21- * (Br) | B3B | 57 | 21.0024 | 1529 | 1532 | cadina-1,4-diene | - | 1-octene |
| 22 | B4A | 69 | 21.5566 | 1552 | 1552 | elemicin | - | |
| 23 | B4A | 57 | 25.1819 | 1704 | 1686 | 8-cedren-13-ol | - | |
| 24 | B4B | 71 | 25.4142 | 1715 | 1735 | oplopanone | - | Hexacosane |
| 25 | B4B | 69 | 26.0498 | 1745 | 1747 | 6S-7R-bisabolone | - | 10-methyl-1-dodecanol |
| 26 | B4A | 57 | 27.4031 | 1805 | 1798 | nootkatone | - | 4-methyl-1-pentene |
| 27 | B3B | 57 | 27.9515 | 1832 | 1827 | isopropyl-tetradecanoate | - | 2,4-dimethylpentane |
| 28 | B3B | 71 | 29.9932 | 1930 | 1927 | methyl-hexadecanoate | - | docosane |
| 29 | B6B | 57 | 30.4182 | 1951 | 1927 | methyl-hexadecanoate | - | - |
| 30 | B10 | 69 | 30.5987 | 1960 | 1927 | methyl-hexadecanoate | - | - |
| 31 | B4C | 73 | 30.6949 | 1965 | 1999 | eicosane | - | - |
| 32 | B6B | 71 | 32.0267 | - | - | 3-hexanone, 2,2-dimethyl- | 2,2-dimethylbutane | |
| 33 | B3B | 71 | 32.0458 | - | - | - | 2,2-dimethylbutane | |
| 34 | B6B | 69 | 34.1452 | - | - | - | docosane | |
| 35 | B3B | 69 | 34.7128 | - | - | - | - | |
| 36 | B4C | 69 | 34.7233 | - | - | - | - | |
| 37 | B6C | 71 | 35.0713 | - | - | - | - | |
| 38 | B3C | 71 | 36.7162 | - | - | - | 3-ethylpentane | |
| 39 | B9B | 395 | 38.2697 | - | - | - | - | |
| 40 | B6C | 71 | 38.8875 | - | - | - | 3-ethylhexane | |
| 41 | B6B | 71 | 39.0147 | - | - | - | - | |
| 42 | B4C | 57 | 39.4792 | - | - | - | 3-ethylhexane | |
| 43 | B6B | 57 | 39.4822 | - | - | - | 3,3-dimethyl-1-butene | |
| 44 | B4C | 71 | 39.6266 | - | - | - | 3-ethylpentane | |
| 45 | B3C | 71 | 39.7884 | - | - | - | - | |
| 46 | B6B | 57 | 40.4997 | - | - | 3-hexanone, 2,2-dimethyl- | 2-methylbutane | |
| 47 | B6C | 57 | 40.7684 | - | - | - | 3-ethylpentane | |
| 48 | B3C | 71 | 40.7722 | - | - | - | - | |
| 49 | B4C | 57 | 43.09 | - | - | - | 3-ethylpentane | |
| 50 | B6B | 57 | 43.0982 | - | - | - | 2,2,4,6,6-pentamethylheptane | |
| 51 | B6C | 85 | 43.2865 | - | - | - | 2-methylnonane | |
| 52 | B6B | 71 | 43.3628 | - | - | - | 3-ethylpentane | |
| 53 | B3C | 71 | 43.3691 | - | - | - | - | |
| 54 | B3B | 85 | 43.7727 | - | - | - | 3,3-dimethyl-1-butene | |
| 55 | B4B | 85 | 44.3901 | - | - | - | Amitrole | |
| 56 | B6B | 71 | 44.3953 | - | - | - | 3-aminopropiononitrile | |
| 57 | B4A | 71 | 46.2638 | - | - | - | - | |
| 58 | B3C | 71 | 46.8143 | - | - | - | 2,3-dimethylbutane | |
| 59 | B6B | 71 | 46.822 | - | - | 3-hexanone, 2,2-dimethyl- | - | |
| 60 | B6C | 71 | 47.3544 | - | - | - | - | |
| 61 | B4A | 71 | 47.7467 | - | - | - | 3-ethylypentane | |
| 62 | B3A | 57 | 47.7529 | - | - | - | 3,3-dimethyl-1-butene | |
| 63 | B4A | 71 | 48.0072 | - | - | - | - | |
| 64 | B6B | 71 | 48.0159 | - | - | - | - | |
| 65 | B6C | 71 | 48.0169 | - | - | - | - | |
| 66 | B4A | 71 | 48.0321 | - | - | - | - | |
| 67 | B4B | 71 | 50.7731 | - | - | - | - | |
| # | Sample | Base Peak | KI | KI Based ID | Mass Spectra Based ID | ||
|---|---|---|---|---|---|---|---|
| RT | Nearest KI | Lucero et al. [37] | Massbase | ||||
| 1 | 3A | 57 | 8.1118 | 1065 | 1062 | y-terpinene | undecane |
| 2 | 15A | 57 | 14.556 | 1285 | 1286 | borneol acetate | 1-nonene |
| 3 | 9A | 57 | 14.556 | 1285 | 1286 | borneol acetate | pentacosane |
| 4 | 29A | 57 | 20.3128 | 1500 | 1499 | a-muurolene | tricosane |
| 5 | 15B | 71 | 20.5458 | 1510 | 1509 | β-bisabolene | 3-ethylpentane |
| 6 | 15A | 57 | 25.186 | 1705 | 1686 | 8-cedren-13-ol | - |
| 7 | 34A | 57 | 25.4206 | 1716 | 1735 | oplopanone | - |
| 8 | 15B | 71 | 26.6674 | 1772 | 1761 | benzyl-benzoate | 2-methylbutane |
| 9 | 15A | 57 | 27.8534 | 1827 | 1827 | isopropyl tetradecanoate | |
| 10 | 44BB | 71 | 30.0015 | 1930 | 1927 | methyl hexadecanoate | docosane |
| 11 | 8B | 57 | 30.4069 | 1950 | 1927 | methyl hexadecanoate | |
| 12 | 15B | 71 | 31.6111 | - | - | - | - |
| 13 | 15A | 57 | 31.8326 | - | - | - | - |
| 14 | 15B | 57 | 31.8342 | - | - | - | 1,4-dimethylhexane |
| 15 | 44BA | 71 | 34.1535 | - | - | - | docosane |
| 16 | 15B | 71 | 34.2967 | - | - | - | 3-ethylpentane |
| 17 | 15A | 57 | 34.76 | - | - | - | 1-octene |
| 18 | 15A | 71 | 35.4496 | - | - | - | 2-methylbutane |
| 19 | 15B | 57 | 35.5757 | - | - | - | - |
| 20 | 15B | 57 | 35.5757 | - | - | - | 2,2-dimethylbutane |
| 21 | 44BA | 71 | 37.9407 | - | - | - | docosane |
| 22 | 15B | 410 | 38.2743 | - | - | - | - |
| 23 | 15A | 71 | 39.3484 | - | - | - | 3-ethylhexane |
| 24 | 15A | 71 | 40.0877 | - | - | - | 2,2-dimethylbutane |
| 25 | 15A | 71 | 40.0981 | - | - | - | beta-Aminopropionitrile |
| 26 | 15B | 71 | 42.5358 | - | - | - | 2-methylbutane |
| 27 | 15B | 57 | 43.9626 | - | - | - | 4-methyloctane |
| 28 | 44C | 71 | 44.6454 | - | - | - | - |
| 29 | 15A | 57 | 45.2255 | - | - | - | 2,2-dimethylbutane |
| 30 | 15A | 71 | 47.3529 | - | - | - | - |
| 31 | 15B | 57 | 47.3534 | - | - | - | - |
| 32 | 15B | 57 | 47.3581 | - | - | - | 2,4-dimethylpentane |
| 33 | 15A | 71 | 47.6237 | - | - | - | 2,4-dimethylhexane |
{kind=link}
Acknowledgments
References
- Williams, D.H.; Stone, M.J.; Hauck, P.R.; Rahman, S.K. Why are secondary metabolites (natural products) biosynthesized? J. Nat. Prod 1989, 52, 1189–1208. [Google Scholar]
- Hay, M.E.; Fenical, W. Chemical ecology and marine biodiversity: Insights and products from the sea. Oceanography 1996, 9, 10–20. [Google Scholar]
- Wise, M.L. Monoterpene biosynthesis in marine algae. Phycologia 2003, 42, 370–377. [Google Scholar]
- Bhat, S.V.; Nagasampagi, B.A.; Sivakumar, M. Chemistry of Natural Products; Narosa Publishing House: New Delhi, India, 2005. [Google Scholar]
- Bourgaud, F.; Gravot, A.; Milesi, S.; Gontier, E. Production of plant secondary metabolites: A historical perspective. Plant Sci 2001, 161, 839–851. [Google Scholar]
- Gribble, G.W. The diversity of naturally produced organohalogens. Chemosphere 2003, 52, 289–297. [Google Scholar]
- Dworjanyn, S.A.; de Nys, R.; Steinberg, P.D. Localisation and surface quantification of secondary metabolites in the red alga Delisea pulchra. Mar. Biol 1999, 133, 727–736. [Google Scholar]
- Gunatilaka, A.A.L.; Paul, V.J.; Park, P.U.; Puglisi, M.P.; Gitler, A.D.; Eggleston, D.S.; Haltiwanger, R.C.; Kingston, D.G.I. Apakaochtodenes a and b: Two tetrahalogenated monoterpenes from the red marine alga Portieria hornemannii. J. Nat. Prod 1999, 62, 1376–1378. [Google Scholar]
- Kladi, M.; Vagias, C.; Roussis, V. Volatile halogenated metabolites from marine red algae. Phytochem. Rev 2004, 3, 337–366. [Google Scholar]
- Fenical, W. Halogenation in the Rhodophyta: A review. J. Phycol 1975, 11, 245–259. [Google Scholar]
- Barahona, L.F.; Rorrer, G.L. Isolation of halogenated monoterpenes from bioreactor-cultured microplantlets of the macrophytic red algae Ochtodes secundiramea and Portieria hornemannii. J. Nat. Prod 2003, 66, 743–751. [Google Scholar]
- Andrianasolo, E.H.; France, D.; Cornell-Kennon, S.; Gerwick, W.H. DNA methyl transferase inhibiting halogenated monoterpenes from the Madagascar red marine alga Portieria hornemannii. J. Nat. Prod 2006, 69, 576–579. [Google Scholar]
- Kuniyoshi, M.; Oshiro, N.; Miono, T.; Higa, T. Halogenated monoterpenes having a cyclohexadienone from the red alga Portieria hornemanni. J. Chin. Chem. Soc 2003, 50, 167–170. [Google Scholar]
- Smit, A.J. Medicinal and pharmaceutical uses of seaweed natural products: A review. J. Appl. Phycol 2004, 16, 245–262. [Google Scholar]
- Kamenarska, Z.; Ivanova, A.; Stancheva, R.; Stoyneva, M.; Stefanov, K.; Dimitrova-Konaklieva, S.; Popov, S. Volatile compounds from some Black Sea red algae and their chemotaxonomic application. Bot. Mar 2006, 49, 47–56. [Google Scholar]
- Blunt, J.W.; Copp, B.R.; Hu, W.P.; Munro, M.H.G.; Northcote, P.T.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep 2007, 24, 31–86. [Google Scholar]
- Bhadury, P.; Wright, P.C. Exploitation of marine algae: Biogenic compounds for potential antifouling applications. Planta 2004, 219, 561–578. [Google Scholar]
- Ichikawa, N.; Naya, Y.; Enomoto, S. New halogenated monoterpenes from Desmia (Chondrococcus) hornemanni. Chem. Lett 1974, 3, 1333–1336. [Google Scholar]
- Burreson, B.J.; Woolard, F.X.; Moore, R.E. Chondrocole a and b, two halogenated dimethylhexahydrobenzofurans from the red alga Chondrococcus hornemannii (Mertens) Schmitz. Tetrahedron Lett 1975, 16, 2155–2158. [Google Scholar]
- Coll, J.C.; Wright, A.D. Tropical marine algae.1. New halogenated monoterpenes from Chondrococcus hornemannii (Rhodophyta, Gigartinales, Rhizophyllidaceae). Aust. J. Chem 1987, 40, 1893–1900. [Google Scholar]
- Wright, A.D.; Konig, G.M.; Sticher, O.; de Nys, R. Five new monoterpenes from the marine red alga Portieria hornemannii. Tetrahedron 1991, 47, 5717–5724. [Google Scholar]
- Fuller, R.W.; Cardellina, J.H.; Kato, Y.; Brinen, L.S.; Clardy, J.; Snader, K.M.; Boyd, M.R. A pentahalogenated monoterpene from the red alga Portieria hornemannii produces a novel cytotoxicity profile against a diverse panel of human tumor cell lines. J. Med. Chem 1992, 35, 3007–3011. [Google Scholar]
- Fuller, R.W.; Cardellina, J.H.; Jurek, J.; Scheuer, P.J.; Alvaradolindner, B.; McGuire, M.; Gray, G.N.; Steiner, J.R.; Clardy, J.; Menez, E.; et al. Isolation and structure activity features of halomon-related antitumor monoterpenes from the red alga Portieria hornemannii. J. Med. Chem 1994, 37, 4407–4411. [Google Scholar]
- Matlock, D.B.; Ginsburg, D.W.; Paul, V.J. Spatial variability in secondary metabolite production by the tropical red alga Portieria hornemannii. Hydrobiologia 1999, 399, 267–273. [Google Scholar]
- Puglisi, M.P.; Paul, V.J. Intraspecific variation in the red alga Portieria hornemannii: Monoterpene concentrations are not influenced by nitrogen or phosphorus enrichment. Mar. Biol 1997, 128, 161–170. [Google Scholar]
- Miller, K.; Alvarez, B.; Battershill, C.; Northcote, P.; Parthasarathy, H. Genetic, morphological, and chemical divergence in the sponge genus Latrunculia (Porifera: Demospongiae) from New Zealand. Mar. Biol 2001, 139, 235–250. [Google Scholar]
- Pietra, F. Biodiversity and Natural Product Diversity; Pergamon Trento: Italy, 2002; Volume 21, p. 351. [Google Scholar]
- McGovern, T.M.; Hellberg, M.E. Cryptic species, cryptic endosymbionts, and geographical variation in chemical defences in the bryozoan Bugula neritina. Mol. Ecol 2003, 12, 1207–1215. [Google Scholar]
- Payo, D.A. Diversity of the Marine Red Alga Portieria in the Philippines, an Integrative Approach. Ph.D. Thesis, Ghent University, Gent, Belgium, 2011. [Google Scholar]
- Fiehn, O.; Kind, T. Metabolite Profiling in Blood Plasma. In Methods in Molecular Biology, Metabolomics: Methods and Protocols; Weckwerth, W., Ed.; Humana Press: Totowa, NJ, USA, 2006; Volume 358, pp. 3–17. [Google Scholar]
- Fiehn, O. Metabolomics—The link between genotypes and phenotypes. Plant Mol. Biol 2002, 48, 155–171. [Google Scholar]
- Lucero, M.; Estell, R.; Tellez, M.; Frederickson, E. A retention index calculator simplifies identification of plant volatile organic compounds. Phytochem. Anal 2009, 20, 378–384. [Google Scholar]
- Egorin, M.J.; Rosen, D.M.; Benjamin, S.E.; Callery, P.S.; Sentz, D.L.; Eiseman, J.L. In vitro metabolism by mouse and human liver preparations of halomon, an antitumor halogenated monoterpene. Cancer Chemother. Pharmacol 1997, 41, 9–14. [Google Scholar]
- Pelletreau, K.N.; Targett, N.M. New perspectives for addressing patterns of secondary metabolites in marine macroalgae. Algal Chem. Ecol 2008, 121–146. [Google Scholar]
- Wise, M.L.; Rorrer, G.L.; Polzin, J.J.; Croteau, R. Biosynthesis of marine natural products: Isolation and characterization of a myrcene synthase from cultured tissues of the marine red alga Ochtodes secundiramea. Arch. Biochem. Biophys 2002, 400, 125–132. [Google Scholar]
- Thornber, C.; Stachowicz, J.J.; Gaines, S. Tissue type matters: Selective herbivory on different life history stages of an isomorphic alga. Ecology 2006, 87, 2255–2263. [Google Scholar]
- Verges, A.; Paul, N.A.; Steinberg, P.D. Sex and life-history stage alter herbivore responses to a chemically defended red alga. Ecology 2008, 89, 1334–1343. [Google Scholar]
- Cavalcanti, D.N.; de Oliveira, M.A.R.; De-Paula, J.C.; Barbosa, L.S.; Fogel, T.; Pinto, M.A.; de Palmer Paixão, I.C.N.; Teixeira, V.L. Variability of a diterpene with potential anti-HIV activity isolated from the Brazilian brown alga Dictyota menstrualis. J. Appl. Phycol 2010, 5, 1–4. [Google Scholar]
- Huelsenbeck, J.P.; Ronquist, F. Mrbayes: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar]
- Posada, D. Jmodeltest: Phylogenetic model averaging. Mol. Biol. Evol 2008, 25, 1253–1256. [Google Scholar]
- Rambaut, A.; Drummond, A.J. Tracer v1.4. Available online: http://beast.bio.ed.ac.uk/Tracer (accessed on 11 October 2007).
- AMDIS. Available online: http://www.amdis.net (accessed on 7 March 2010).
- Styczynski, M.P.; Moxley, J.F.; Tong, L.V.; Walther, J.L.; Jensen, K.L.; Stephanopoulos, G.N. Systematic identification of conserved metabolites in GC/MS data for metabolomics and biomarker discovery. Anal. Chem. 2007, 79, pp. 966–973. Available online: http://spectconnect.mit.edu (accessed on 23 February 2010).
- Horai, H.; Arita, M.; Kanaya, S.; Nihei, Y.; Ikeda, T.; Suwa, K.; Ojima, Y.; Tanaka, K.; Tanaka, S.; Aoshima, K.; et al. Massbank: A public repository for sharing mass spectral data for life sciences. J. Mass Spectrometry. 2010, 45, pp. 703–714. Available online: http://www.massbank.jp (accessed on 18 March 2010).
- The Pherobase: Database of Pheromones and Semiochemicals. Available online: http://www.pherobase.com (accessed on 11 August 2010).
- Burreson, B.J.; Woolard, F.X.; Moore, R.E. Evidence for the biogenesis of halogenated myrcenes from the red alga Chondrococcus hornemanni. Chem. Lett 1975, 4, 1111–1114. [Google Scholar]
- Ichikawa, N.; Naya, Y.; Enomoto, S. Halogenated monoterpene derivatives from Desmia (Chondrococcus) japonicus. Proc. Jpn. Acad 1975, 51, 562–565. [Google Scholar]
- Woolard, F.X.; Moore, R.E.; Mahendran, M.; Sivapalan, A. (−)-3-bromomethyl-3-chloro-7- methyl-1 6-octadiene from Sri Lankan Chondrococcus hornemanni. Phytochemistry 1976, 15, 1069–1070. [Google Scholar]
- Higa, T. 2-(1-chloro-2-hydroxyethyl)-4,4-dimethylcyclohexa-2,5-dienone: A precursor of 4, 5-dimethylbenzo [β] furan from the red alga Desmia hornemanni. Tetrahedron Lett 1985, 26, 2335–2336. [Google Scholar]
- Coll, J.C.; Wright, A.D. Tropical marine-algae. 6. New monoterpenes from several collections of Chondrococcus hornemannii (Rhodophyta, Gigartinales, Rhizophyllidaceae). Aust. J. Chem 1989, 42, 1983–1993. [Google Scholar]




| B21 (49) | B33 (51) | B35 (60) | B8 (57) | B5 (57) | Frequency (67) |
|---|---|---|---|---|---|
| 0 | 0 | 0 | 1 | 0 | 1 |
| 0 | 0 | 1 | 1 | 0 | 3 |
| 0 | 0 | 1 | 1 | 1 | 3 |
| 0 | 1 | 0 | 0 | 0 | 1 |
| 0 | 1 | 0 | 1 | 0 | 2 |
| 0 | 1 | 1 | 1 | 0 | 1 |
| 0 | 1 | 1 | 1 | 1 | 7 |
| 1 | 0 | 1 | 0 | 1 | 3 |
| 1 | 0 | 1 | 1 | 0 | 2 |
| 1 | 0 | 1 | 1 | 1 | 4 |
| 1 | 1 | 0 | 0 | 1 | 2 |
| 1 | 1 | 0 | 1 | 1 | 1 |
| 1 | 1 | 1 | 0 | 1 | 4 |
| 1 | 1 | 1 | 1 | 1 | 33 |
© 2011 by the authors; licensee MDPI, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Payo, D.A.; Colo, J.; Calumpong, H.; Clerck, O.d. Variability of Non-Polar Secondary Metabolites in the Red Alga Portieria. Mar. Drugs 2011, 9, 2438-2468. https://doi.org/10.3390/md9112438
Payo DA, Colo J, Calumpong H, Clerck Od. Variability of Non-Polar Secondary Metabolites in the Red Alga Portieria. Marine Drugs. 2011; 9(11):2438-2468. https://doi.org/10.3390/md9112438
Chicago/Turabian StylePayo, Dioli Ann, Joannamel Colo, Hilconida Calumpong, and Olivier de Clerck. 2011. "Variability of Non-Polar Secondary Metabolites in the Red Alga Portieria" Marine Drugs 9, no. 11: 2438-2468. https://doi.org/10.3390/md9112438
APA StylePayo, D. A., Colo, J., Calumpong, H., & Clerck, O. d. (2011). Variability of Non-Polar Secondary Metabolites in the Red Alga Portieria. Marine Drugs, 9(11), 2438-2468. https://doi.org/10.3390/md9112438