Towards a Guided Regeneration of Renal Tubules at a Polyester Interstitium

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Limited regeneration of renal parenchyma
1.2. Sensitivity of renal cells

1.3. From cells to parenchyma
2. View to the Embryonic Parenchyma
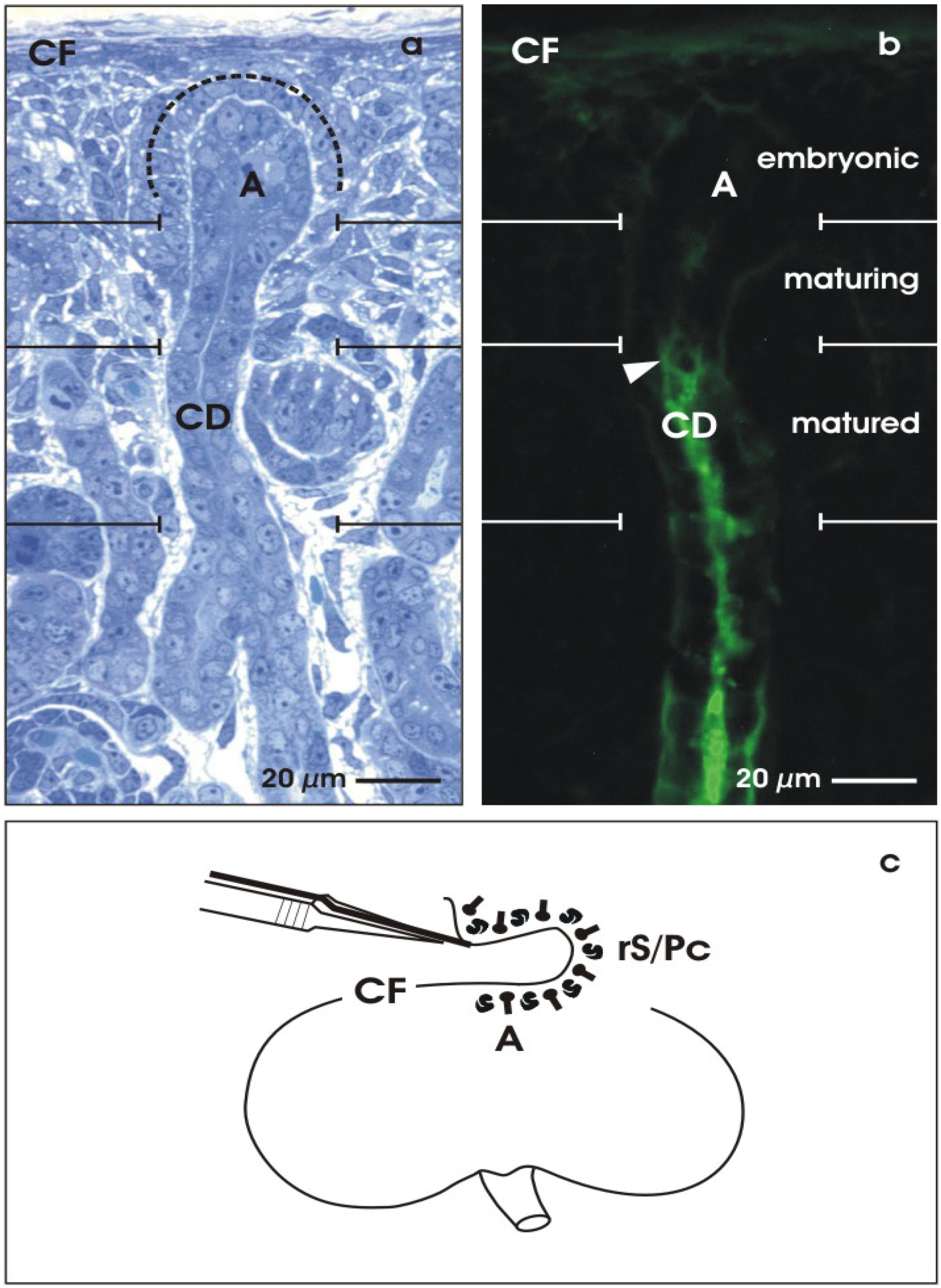
2.1. The renal stem cell niche

2.2. Maturation of stem/progenitor cells within the collecting duct ampulla
2.3. Isolation of renal stem/progenitor cells
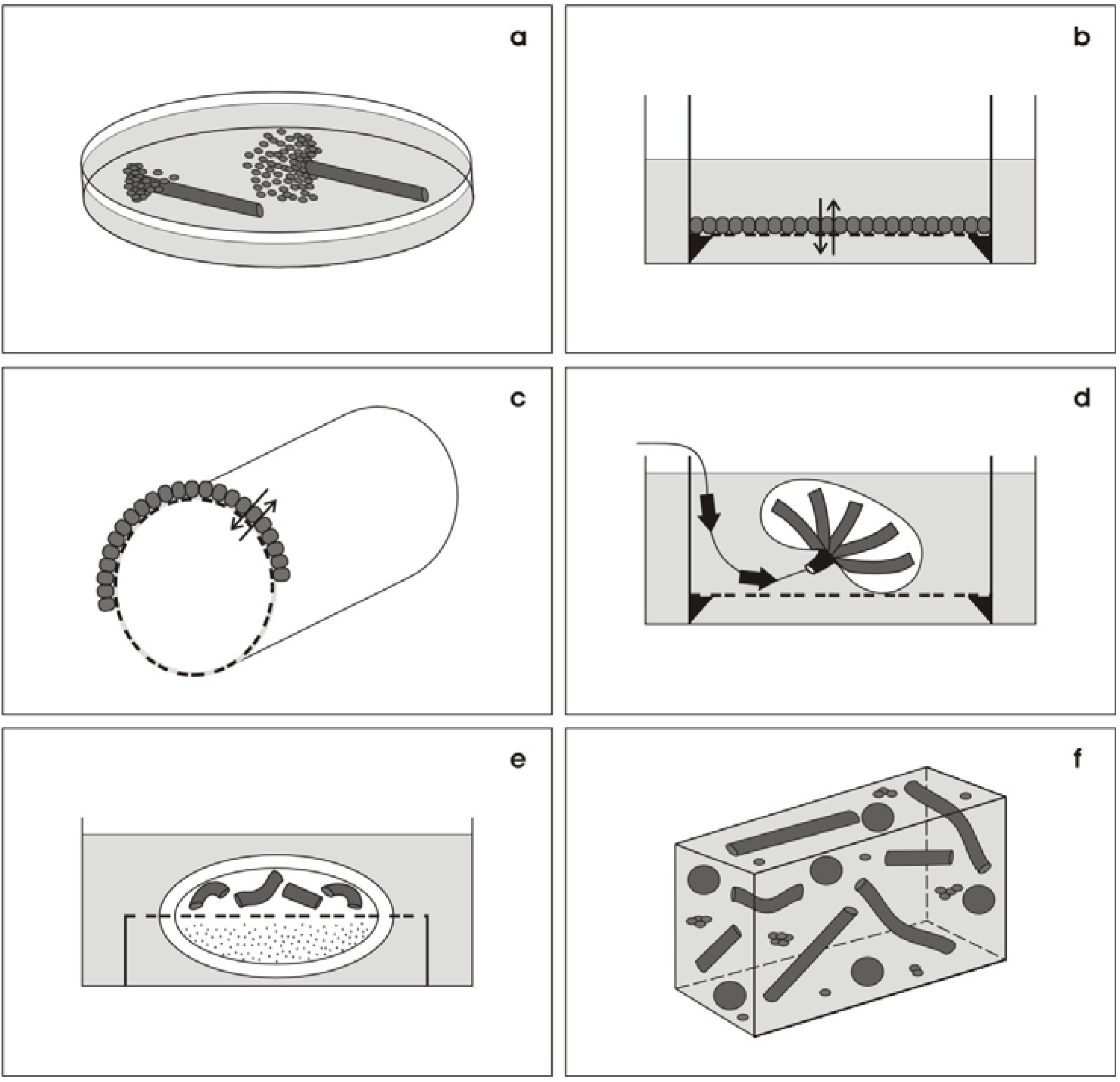
3. The New Concept—Offering an Artificial Interstitium
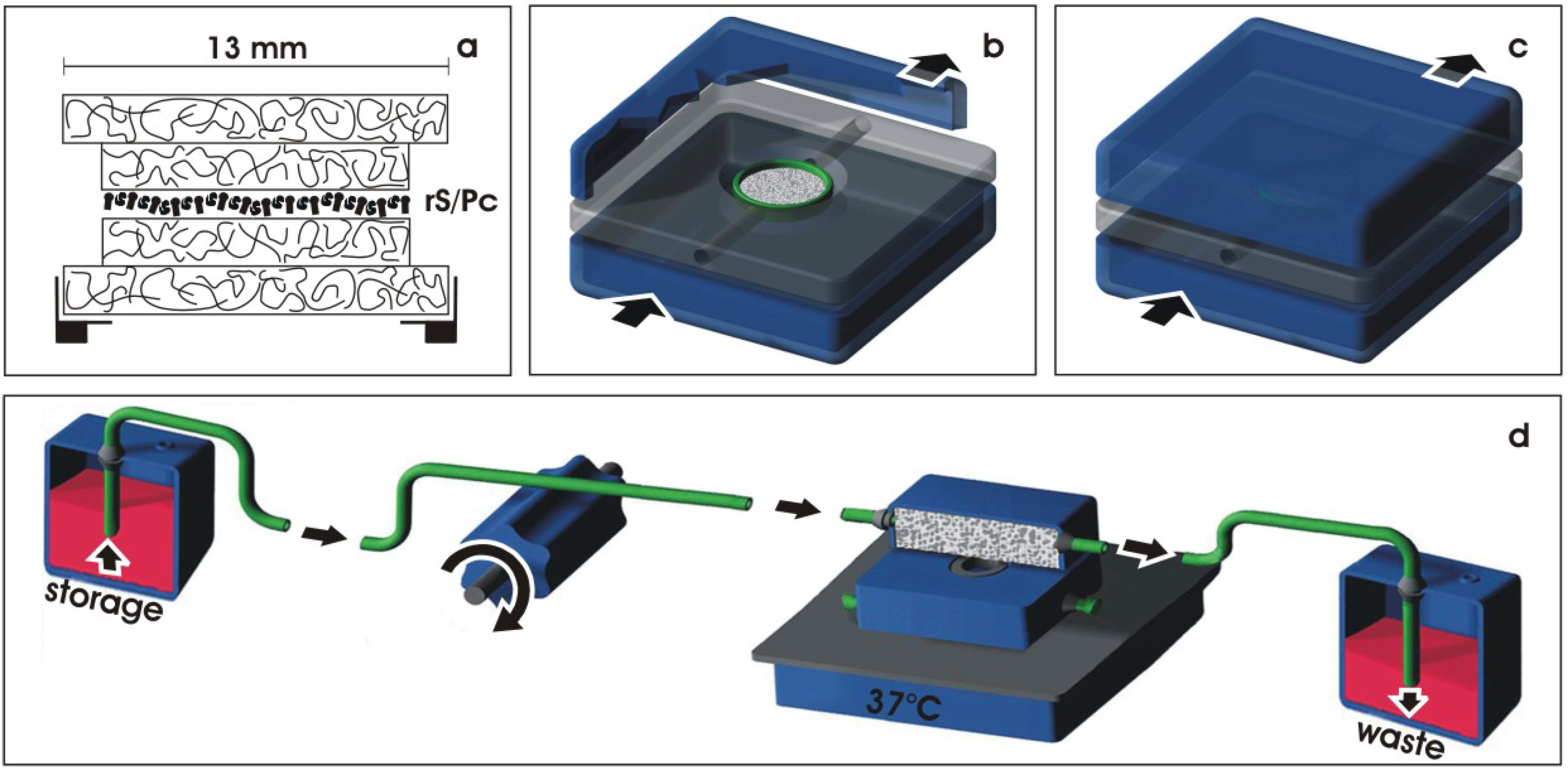
3.1. Environment for spatial development

3.2. Protecting the developing tissue

3.3. Transport of culture medium
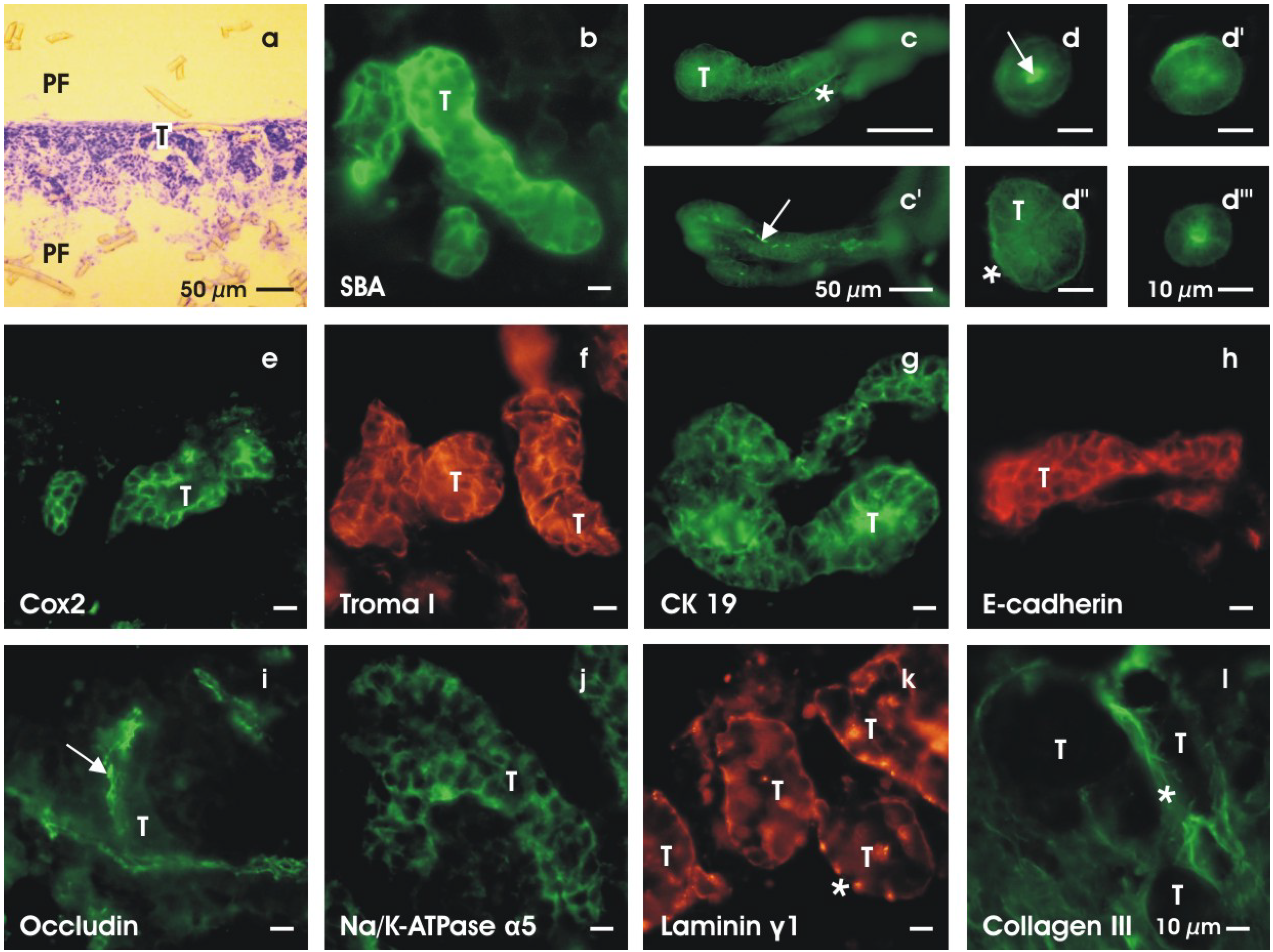
4. Experiments on Generated Tubules
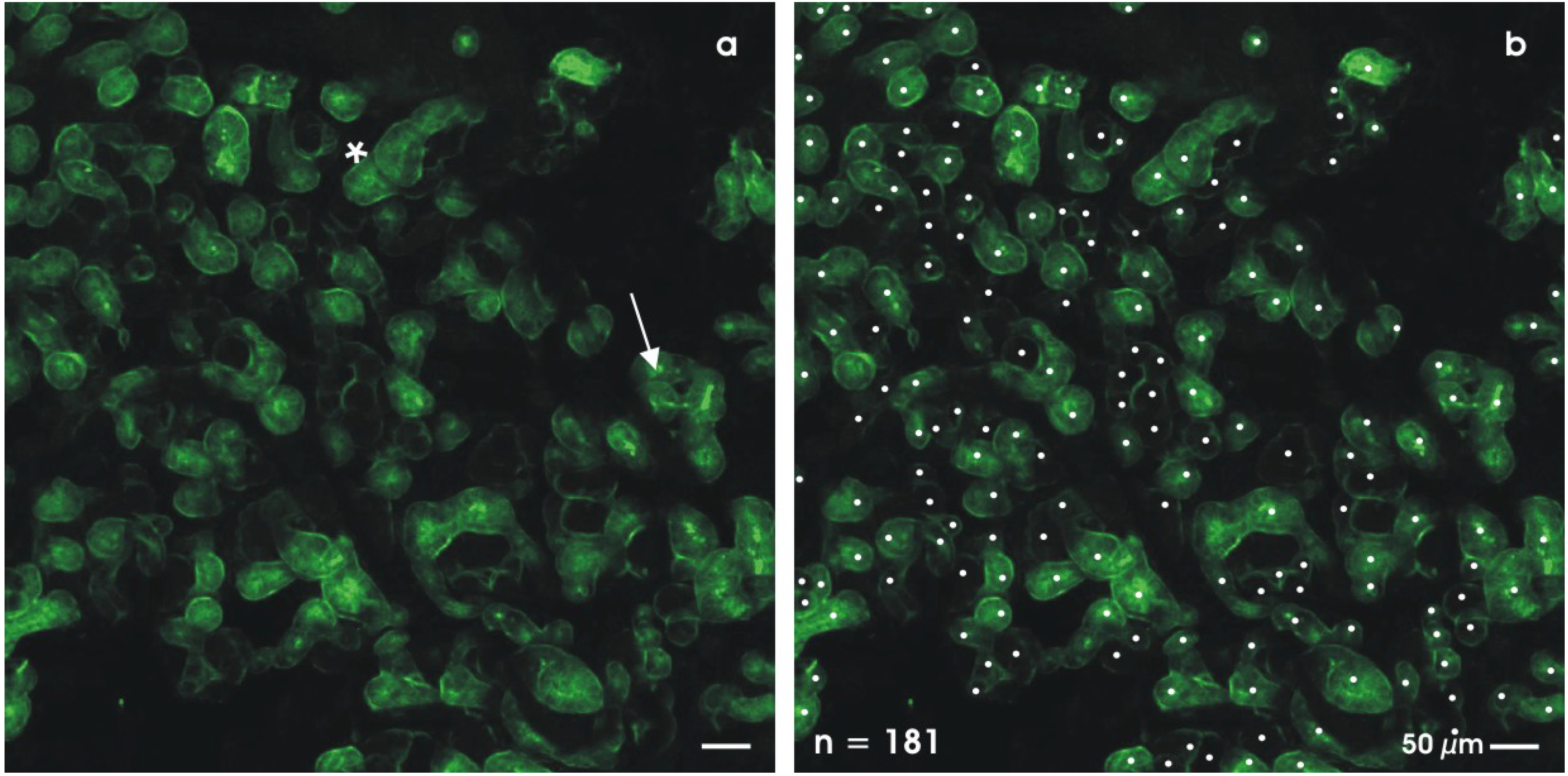
4.1. Labeling whole mount specimens

4.2. Detecting cell biological differentiation

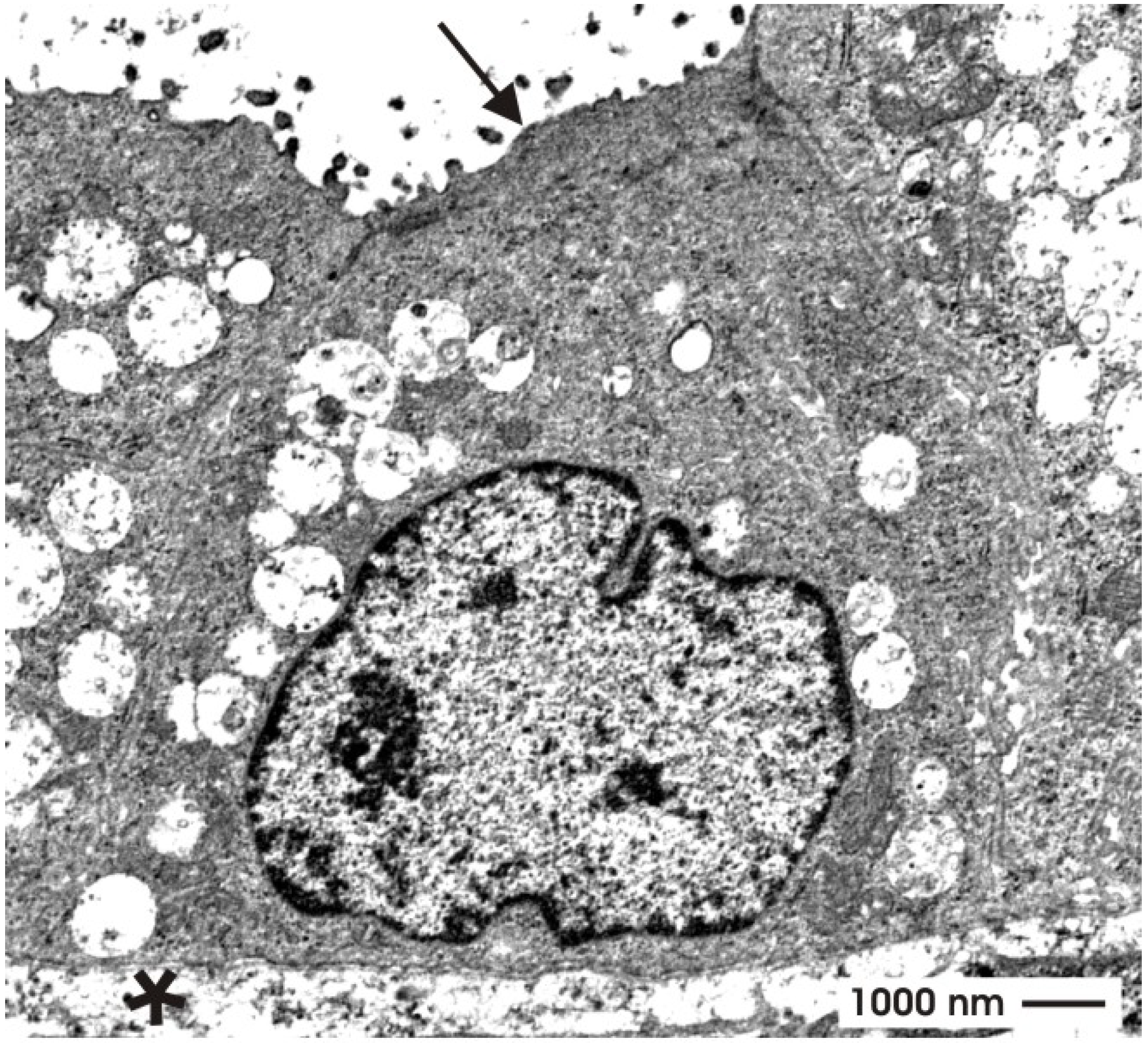
4.3. Regarding ultrastructural features


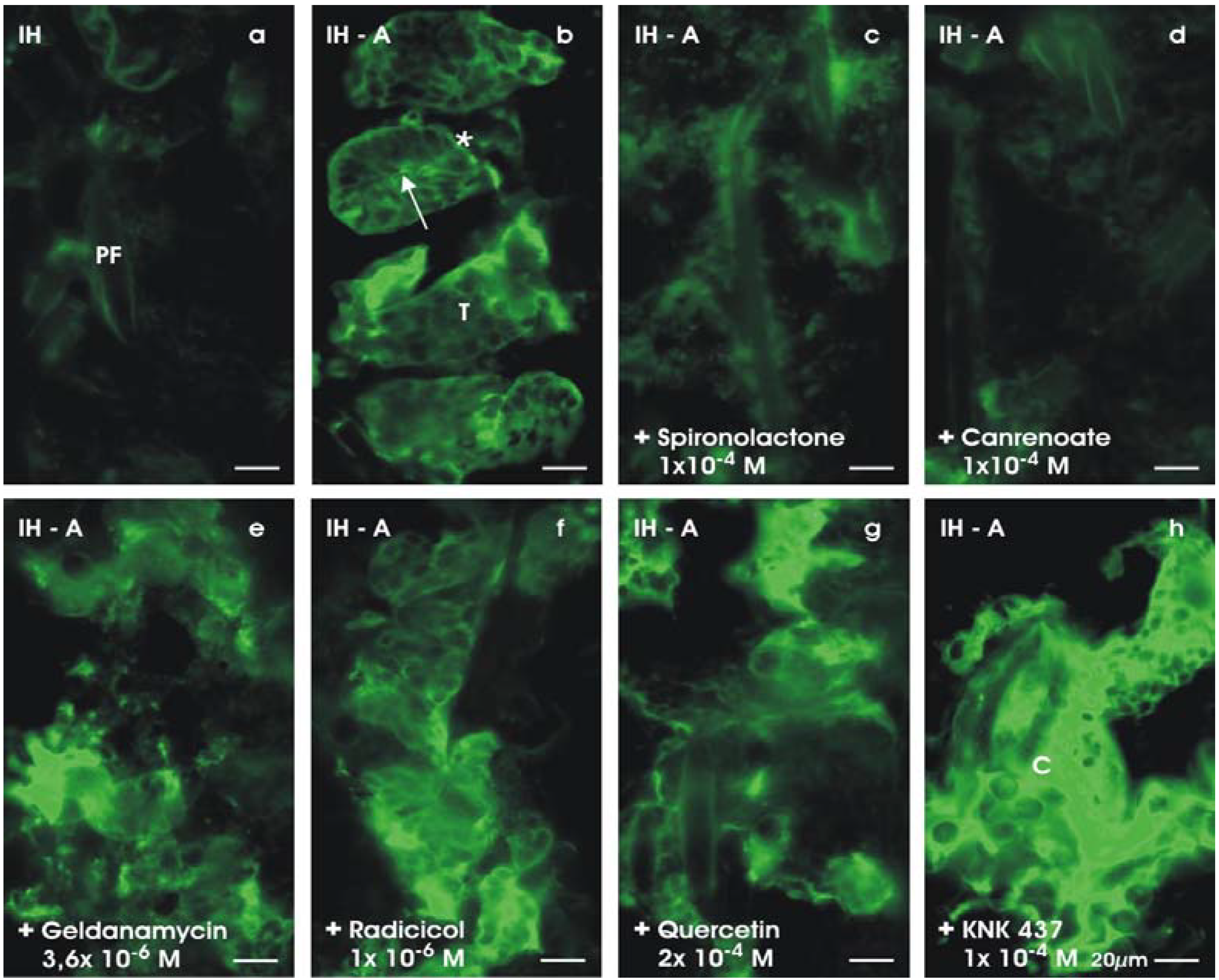
5. Triggering Formation of Tubules
5.1. Inducing tubulogenic development

5.2. Specifity of aldosterone action
5.3. Antagonizing the action of aldosterone
5.4. Mineralocorticoid receptor and chaperons
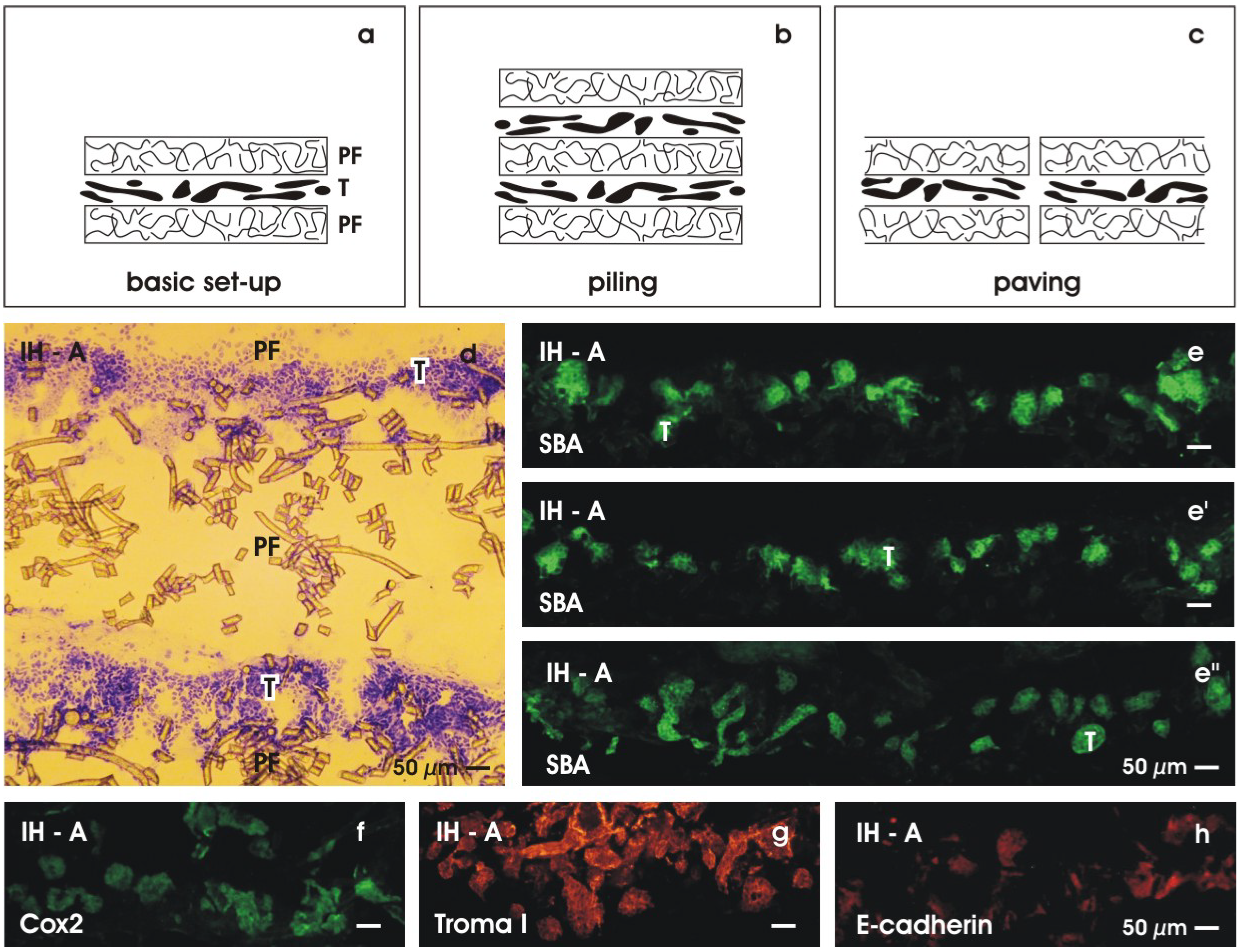
6. Considerations about Implantation
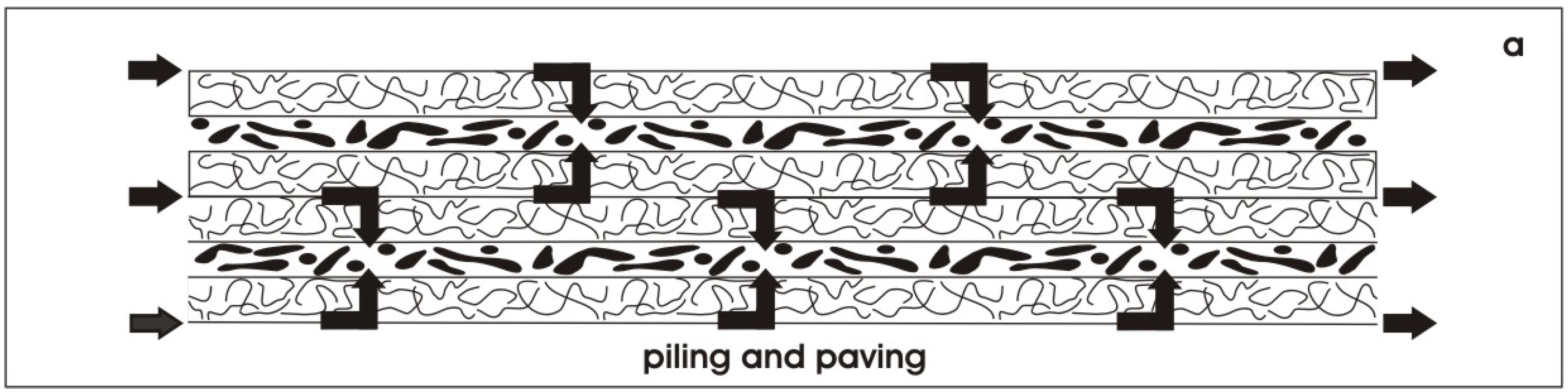
6.1. Piling of renal stem/progenitor cells

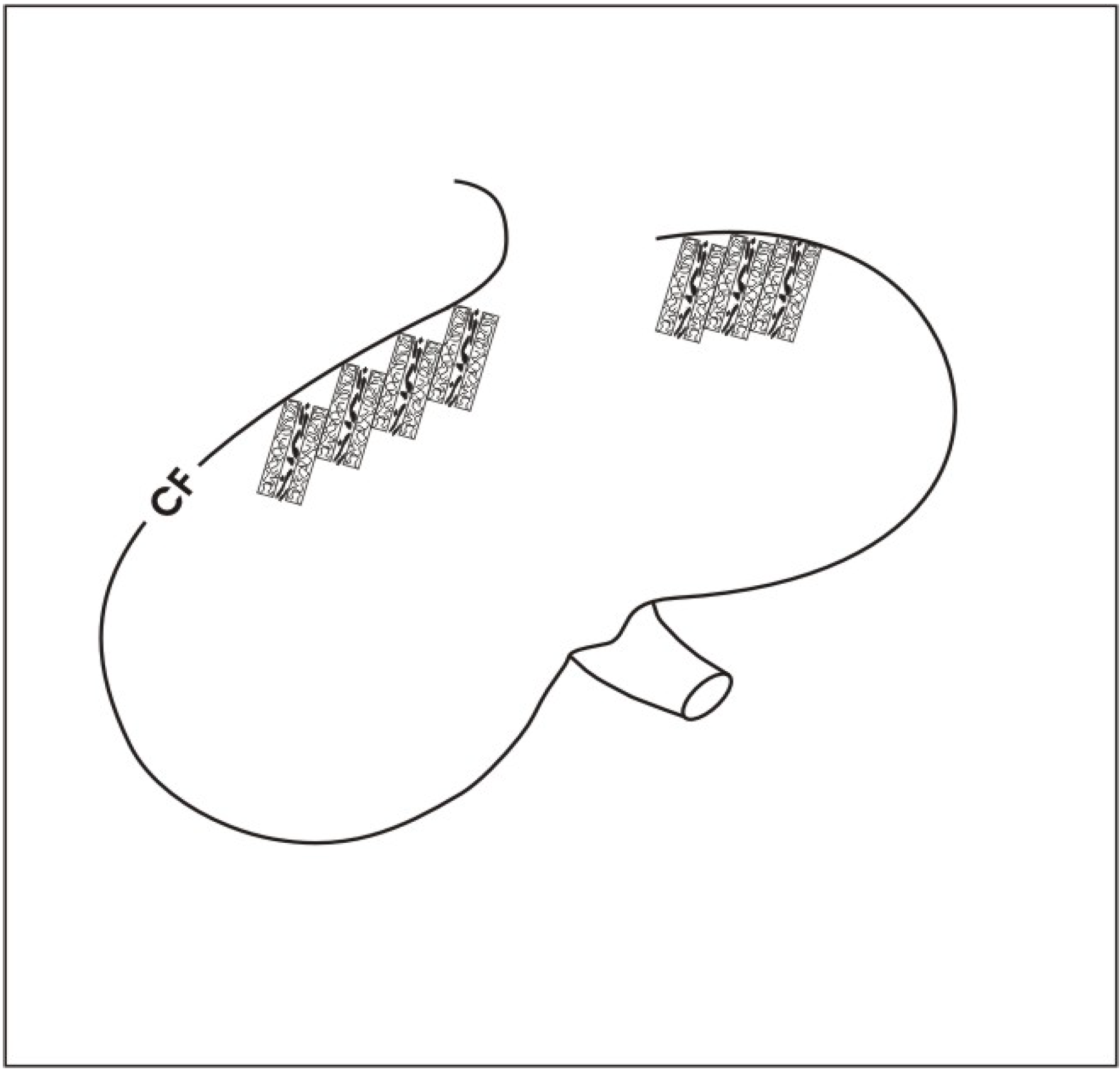
6.2. Finding a solution for rapid micro-vascularization

6.3. Using basic sandwich set-ups as a subcapsular implant

7. Conclusions
References and Notes
- Rookmaaker, M.B.; Verhaar, M.C.; van Zonneveld, A.J.; Rabelink, T.J. Progenitor cells in the kidney: Biology and therapeutic perspectives. Kidney Int. 2004, 2, 518–522. [Google Scholar] [CrossRef]
- Mansilla, E.; Drago, H.; Sturla, F.; Bossi, S.; Salas, E.; Marín, G.H.; Ibar, R.; Soratti, C. Matrix superhighways configurations: New concepts for complex organ regeneration. Transplant. Proc. 2007, 7, 2431–2433. [Google Scholar] [CrossRef]
- Humphreys, B.D.; Bonventre, J.V. The contribution of adult stem cells to renal repair. Nephrol. Ther. 2007, 1, 3–10. [Google Scholar] [CrossRef]
- McTaggart, S.J.; Atkinson, K. Mesenchymal stem cells: Immunobiology and therapeutic potential in kidney disease. Nephrology (Carlton) 2007, 1, 44–52. [Google Scholar] [CrossRef]
- Vigneau, C.; Polgar, K.; Striker, G.; Elliott, J.; Hyink, D.; Weber, O.; Fehling, H.-J.; Keller, G.; Burrow, C.; Wilson, P. Mouse embryonic stem cell-derived embryoid bodies generate progenitors that integrate long-term into renal proximal tubules in vivo. J. Am. Soc. Nephrol. 2007, 6, 1709–1720. [Google Scholar] [CrossRef]
- Steenhard, B.M.; Isom, K.S.; Cazcarro, P.; Dunmore, J.H.; Godwin, A.R.; St John, P.L.; Abrahamson, D.R. Integration of embryonic stem cells in metanephric kidney organ culture. J. Am. Soc. Nephrol. 2005, 6, 1623–1631. [Google Scholar] [CrossRef]
- Hammerman, M.R. Cellular therapies for kidney failure. Expert. Opin. Biol. Ther. 2006, 2, 87–97. [Google Scholar] [CrossRef]
- Maeshima, A.; Sakurai, H.; Nigam, S.K. Adult kidney tubular cell population showing phenotypic plasticity, tubulogenic capacity, and integration capability into developing kidney. J. Am. Soc. Nephrol. 2006, 1, 188–198. [Google Scholar]
- Atala, A. Bioengineered tissues for urogenital repair in children. Pediatr. Res. 2008, 5, 569–575. [Google Scholar] [CrossRef]
- Perin, L.; Giuliani, S.; Sedrakyan, S.; DA Sacco, S.; Filippo, R.E.de. Stem cell and regenerative science applications in the development of bioengineering of renal tissue. Pediatr. Res. 2008, 5, 467–471. [Google Scholar] [CrossRef]
- Loo, D.; Beltejar, C.; Hooley, J.; Xu, X. Primary and multipassage culture of human fetal kidney epithelial progenitor cells. Methods Cell Biol. 2008, 241–255. [Google Scholar]
- Giuliani, S.; Perin, L.; Sedrakyan, S.; Kokorowski, P.; Jin, D.; Filippo, R.de. Ex vivo whole embryonic kidney culture: A novel method for research in development, regeneration and transplantation. J. Urol. 2008, 1, 365–370. [Google Scholar] [CrossRef]
- Koh, C.J.; Atala, A. Tissue engineering, stem cells, and cloning: Opportunities for regenerative medicine. J. Am. Soc. Nephrol. 2004, 5, 1113–1125. [Google Scholar] [CrossRef]
- Atala, A. Tissue engineering, stem cells and cloning: Current concepts and changing trends. Expert Opin. Biol. Ther. 2005, 7, 879–892. [Google Scholar] [CrossRef]
- Little, M.H. Regrow or repair: Potential regenerative therapies for the kidney. J. Am. Soc. Nephrol. 2006, 9, 2390–2401. [Google Scholar] [CrossRef]
- Bussolati, B.; Camussi, G. Stem cells in acute kidney injury. Contrib. Nephrol. 2007, 156, 250–258. [Google Scholar] [PubMed]
- Sallustio, F.; Benedictis, L.; de Castellano, G.; Zaza, G.; Loverre, A.; Costantino, V.; Grandaliano, G.; Schena, F.P. TLR2 plays a role in the activation of human resident renal stem/progenitor cells. FASEB J. 2009. [Google Scholar] [CrossRef]
- Metsuyanim, S.; Harari-Steinberg, O.; Buzhor, E.; Omer, D.; Pode-Shakked, N.; Ben-Hur, H.; Halperin, R.; Schneider, D.; Dekel, B. Expression of stem cell markers in the human fetal kidney. PLoS ONE 2009, 8, e6709. [Google Scholar] [CrossRef]
- Hellman, N.E.; Greco, A.J.; Rogers, K.K.; Kanchagar, C.; Balkovetz, D.F.; Lipschutz, J.H. Activated extracellular signal-regulated kinases are necessary and sufficient to initiate tubulogenesis in renal tubular MDCK strain I cell cysts. Am. J. Physiol. Renal Physiol. 2005, 4, F777–F785. [Google Scholar] [CrossRef]
- Meyer, T.N.; Schwesinger, C.; Bush, K.T.; Stuart, R.O.; Rose, D.W.; Shah, M.M.; Vaughn, D.A.; Steer, D.L.; Nigam, S.K. Spatiotemporal regulation of morphogenetic molecules during in vitro branching of the isolated ureteric bud: Toward a model of branching through budding in the developing kidney. Dev. Biol. 2004, 1, 44–67. [Google Scholar] [CrossRef]
- Ash, S.R.; Cuppage, F.E.; Hoses, M.E.; Selkurt, E.E. Culture of isolated renal tubules: A method of assessing viability of normal and damaged cells. Kidney Int. 1975, 1, 55–60. [Google Scholar] [CrossRef]
- Wilson, P.D.; Horster, M.F. Differential response to hormones of defined distal nephron epithelia in culture. Am. J. Physiol. 1983, 3, C166–C174. [Google Scholar]
- Fissell, W.H.; Fleischman, A.J.; Humes, H.D.; Roy, S. Development of continuous implantable renal replacement: Past and future. Transl. Res. 2007, 6, 327–336. [Google Scholar] [CrossRef]
- Ding, F.; Humes, H.D. The bioartificial kidney and bioengineered membranes in acute kidney injury. Nephron Exp. Nephrol. 2008, 4, e118–e122. [Google Scholar] [CrossRef]
- Rebollar, E.; Frischauf, I.; Olbrich, M.; Peterbauer, T.; Hering, S.; Preiner, J.; Hinterdorfer, P.; Romanin, C.; Heitz, J. Proliferation of aligned mammalian cells on laser-nanostructured polystyrene. Biomaterials 2008, 12, 1796–1806. [Google Scholar] [CrossRef]
- Candia, O.; Mia, A.J.; Yorio, T. Influence of filter supports on transport characteristics of cultured A6 kidney cells. Am. J. Physiol. 1993, 6 (Pt 1), C1479–C1488. [Google Scholar]
- Genestie, I.; Morin, J.P.; Guery, C.; Bode, G.; Lorenzon, G. Morphological and biochemical characterization of primary culture of rabbit proximal kidney tubule cells grown on collagen-IV coated Millicell-CM. In Vitro Cell. Dev. Biol. Anim. 1997, 9, 692–702. [Google Scholar] [CrossRef]
- Cook, J.R.; Crute, B.E.; Patrone, L.M.; Gabriels, J.; Lane, M.E.; van Buskirk, R.G. Microporosity of the substratum regulates differentiation of MDCK cells in vitro. In Vitro Cell. Dev. Biol. 1989, 10, 914–922. [Google Scholar] [CrossRef]
- Hu, M.; Deng, R.; Schumacher, K.M.; Kurisawa, M.; Ye, H.; Purnamawati, K.; Ying, J.Y. Hydrodynamic spinning of hydrogel fibers. Biomaterials 2010, 5, 863–869. [Google Scholar] [CrossRef]
- Hardman, P.; Klement, B.J.; Spooner, B.S. Growth and morphogenesis of embryonic mouse organs on non-coated and extracellular matrix-coated Biopore membrane. Dev. Growth Differ. 1993, 6, 683–690. [Google Scholar] [CrossRef]
- Ueland, J.; Yuan, A.; Marlier, A.; Gallagher, A.R.; Karihaloo, A. A novel role for the chemokine receptor Cxcr4 in kidney morphogenesis: An in vitro study. Dev. Dyn. 2009, 5, 1083–1091. [Google Scholar] [CrossRef]
- Grobstein, C. Trans-filter induction of tubules in mouse metanephrogenic mesenchyme. Exp. Cell Res. 1956, 2, 424–440. [Google Scholar] [CrossRef]
- Saxén, L.; Lehtonen, E. Embryonic kidney in organ culture. Differentiation 1987, 1, 2–11. [Google Scholar] [CrossRef]
- Michos, O. Kidney development: From ureteric bud formation to branching morphogenesis. Curr. Opin. Genet. Dev. 2009, 5, 484–490. [Google Scholar] [CrossRef]
- Shan, J.; Jokela, T.; Skovorodkin, I.; Vainio, S. Mapping of the fate of cell lineages generated from cells that express the Wnt4 gene by time-lapse during kidney development. Differentiation 2010, 1, 57–64. [Google Scholar] [CrossRef]
- Inoue, C.N.; Sunagawa, N.; Morimoto, T.; Ohnuma, S.; Katsushima, F.; Nishio, T.; Kondo, Y.; Iinuma, K. Reconstruction of tubular structures in three-dimensional collagen gel culture using proximal tubular epithelial cells voided in human urine. In Vitro Cell. Dev. Biol. Anim. 2003, 8–9, 364–367. [Google Scholar] [CrossRef]
- Chung, I.-M.; Enemchukwu, N.O.; Khaja, S.D.; Murthy, N.; Mantalaris, A.; García, A.J. Bioadhesive hydrogel microenvironments to modulate epithelial morphogenesis. Biomaterials 2008, 17, 2637–2645. [Google Scholar] [CrossRef]
- Kong, T.; Xu, D.; Yu, W.; Takakura, A.; Boucher, I.; Tran, M.; Kreidberg, J.A.; Shah, J.; Zhou, J.; Denker, B.M. G alpha 12 inhibits alpha2 beta1 integrin-mediated Madin-Darby canine kidney cell attachment and migration on collagen-i and blocks tubulogenesis. Mol. Biol. Cell 2009, 21, 4596–4610. [Google Scholar] [CrossRef]
- Brakeman, P.R.; Liu, K.D.; Shimizu, K.; Takai, Y.; Mostov, K.E. Nectin proteins are expressed at early stages of nephrogenesis and play a role in renal epithelial cell morphogenesis. Am. J. Physiol. Renal Physiol. 2009, 3, F564–F574. [Google Scholar]
- Rosines, E.; Sampogna, R.V.; Johkura, K.; Vaughn, D.A.; Choi, Y.; Sakurai, H.; Shah, M.M.; Nigam, S.K. Staged in vitro reconstitution and implantation of engineered rat kidney tissue. Proc. Natl. Acad. Sci. USA 2007, 52, 20938–20943. [Google Scholar] [CrossRef]
- Schumacher, K.; Strehl, R.; de Vries, U.; Groene, H.J.; Minuth, W.W. SBA-positive fibers between the CD ampulla, mesenchyme, and renal capsule. J. Am. Soc. Nephrol. 2002, 10, 2446–2453. [Google Scholar] [CrossRef]
- Minuth, W.W.; Schumacher, K. From the renal stem cell niche to functional tubule. Med. Klin. (Munich) 2003, 31–35. [Google Scholar]
- Hartman, H.A.; Lai, H.L.; Patterson, L.T. Cessation of renal morphogenesis in mice. Dev. Biol. 2007, 2, 379–387. [Google Scholar] [CrossRef]
- Strehl, R.; Minuth, W.W. Nephron induction—the epithelial mesenchymal interface revisited. Pediatr. Nephrol. 2001, 1, 38–40. [Google Scholar] [CrossRef]
- Dressler, G. Tubulogenesis in the developing mammalian kidney. Trends Cell Biol. 2002, 8, 390–395. [Google Scholar] [CrossRef]
- Walid, S.; Eisen, R.; Ratcliffe, D.R.; Dai, K.; Hussain, M.M.; Ojakian, G.K. The PI 3-kinase and mTOR signaling pathways are important modulators of epithelial tubule formation. J. Cell. Physiol. 2008, 2, 469–479. [Google Scholar] [CrossRef]
- Hogan, B.L.M.; Kolodziej, P.A. Organogenesis: Molecular mechanisms of tubulogenesis. Nat. Rev. Genet. 2002, 7, 513–523. [Google Scholar] [CrossRef]
- Lubarsky, B.; Krasnow, M.A. Tube morphogenesis: Making and shaping biological tubes. Cell 2003, 1, 19–28. [Google Scholar] [CrossRef]
- Karihaloo, A.; Nickel, C.; Cantley, L.G. Signals which build a tubule. Nephron Exp. Nephrol. 2005, 1, e40–e45. [Google Scholar] [CrossRef]
- Karner, C.M.; Chirumamilla, R.; Aoki, S.; Igarashi, P.; Wallingford, J.B.; Carroll, T.J. Wnt9b signaling regulates planar cell polarity and kidney tubule morphogenesis. Nat. Genet. 2009, 7, 793–799. [Google Scholar] [CrossRef]
- Minuth, W.W. Neonatal rabbit kidney cortex in culture as tool for the study of collecting duct formation and nephron differentiation. Differentiation 1987, 1, 12–22. [Google Scholar] [CrossRef]
- Avner, E.D.; Ellis, D.; Temple, T.; Jaffe, R. Metanephric development in serum-free organ culture. In Vitro 1982, 8, 675–682. [Google Scholar] [CrossRef]
- Heber, S.; Denk, L.; Hu, K.; Minuth, W.W. Modulating the development of renal tubules growing in serum-free culture medium at an artificial interstitium. Tissue Eng. 2007, 2, 281–292. [Google Scholar] [CrossRef]
- Hu, K.; Denk, L.; Vries, U. de; Minuth, W.W. Chemically defined medium environment for the development of renal stem cells into tubules. Biotechnol. J. 2007, 8, 992–995. [Google Scholar]
- Minuth, W.W. Vorrichtung zum Kultivieren und/oder Differenzieren und/oder Halten von Zellen und/oder Geweben. DE 199 52 847, 2006. [Google Scholar]
- Minuth, W.W.; Denk, L.; Heber, S. Growth of embryonic renal parenchyme at the interphase of a polyester artificial interstitium. Biomaterials 2005, 33, 6588–6598. [Google Scholar] [CrossRef]
- Minuth, W.W.; Denk, L.; Hu, K. The role of polyester interstitium and aldosterone during structural development of renal tubules in serum-free medium. Biomaterials 2007, 30, 4418–4428. [Google Scholar] [CrossRef]
- Blattmann, A.; Denk, L.; Strehl, R.; Castrop, H.; Minuth, W.W. The formation of pores in the basal lamina of regenerated renal tubules. Biomaterials 2008, 18, 2749–2756. [Google Scholar] [CrossRef]
- Minuth, W.W.; Denk, L.; Meese, C.; Rachel, R.; Roessger, A. Ultrastructural insights in the interface between generated renal tubules and a polyester interstitium. Langmuir 2009, 8, 4621–4627. [Google Scholar] [CrossRef]
- Minuth, W.W.; Denk, L. Aldosterone-dependent generation of tubules derived from renal stem/progenitor cells. Transplantationsmedizin 2008, 20, 42–47. [Google Scholar]
- Minuth, W.W.; Denk, L.; Hu, K.; Castrop, H.; Gomez-Sanchez, C. The tubulogenic effect of aldosterone is attributed to intact binding and intracellular response of the mineralocorticoid receptor. Cent. Eur. J. Biol. 2007, 2, 3307–3325. [Google Scholar] [CrossRef]
- Pratt, W.B.; Galigniana, M.D.; Harrell, J.M.; DeFranco, D.B. Role of hsp90 and the hsp90-binding immunophilins in signalling protein movement. Cell. Signal. 2004, 8, 857–872. [Google Scholar] [CrossRef]
- Bamberger, C.M.; Wald, M.; Bamberger, A.M.; Schulte, H.M. Inhibition of mineralocorticoid and glucocorticoid receptor function by the heat shock protein 90-binding agent geldanamycin. Mol. Cell. Endocrinol. 1997, 2, 233–240. [Google Scholar] [CrossRef]
- Batulan, Z.; Taylor, D.M.; Aarons, R.J.; Minotti, S.; Doroudchi, M.M.; Nalbantoglu, J.; Durham, H.D. Induction of multiple heat shock proteins and neuroprotection in a primary culture model of familial amyotrophic lateral sclerosis. Neurobiol. Dis. 2006, 2, 213–225. [Google Scholar] [CrossRef]
- Minuth, W.W.; Blattmann, A.; Denk, L.; Castrop, H. Mineralocorticoid Receptor, Heat Shock Proteins and Immunophilins Participate in the Transmission of the Tubulogenic Signal of Aldosterone. J. Epithel. Biol. Pharmacol. 2008, 11, 24–34. [Google Scholar] [CrossRef]
- Manwell, L.A.; Heikkila, J.J. Examination of KNK437- and quercetin-mediated inhibition of heat shock-induced heat shock protein gene expression in Xenopus laevis cultured cells. Comp. Biochem. Physiol., A Mol. Integr. Physiol. 2007, 3, 521–530. [Google Scholar] [CrossRef]
- Yokota, S.; Kitahara, M.; Nagata, K. Benzylidene lactam compound, KNK437, a novel inhibitor of acquisition of thermotolerance and heat shock protein induction in human colon carcinoma cells. Cancer Res. 2000, 11, 2942–2948. [Google Scholar]
- Burst, V.R.; Gillis, M.; Pütsch, F.; Herzog, R.; Fischer, J.H.; Heid, P.; Müller-Ehmsen, J.; Schenk, K.; Fries, J.W.U.; Baldamus, C.A.; Benzing, T. Poor Cell Survival Limits the Beneficial Impact of Mesenchymal Stem Cell Transplantation on Acute Kidney Injury. Nephron Exp. Nephrol. 2009, 3, e107–e116. [Google Scholar]
- Minuth, W.W.; Denk, L.; Castrop, H. Generation of tubular superstructures by piling of renal stem/progenitor cells. Tissue Eng. C Methods 2008, 1, 3–13. [Google Scholar] [CrossRef]
- Curtis, L.M.; Chen, S.; Chen, B.; Agarwal, A.; Klug, C.A.; Sanders, P.W. Contribution of intrarenal cells to cellular repair after acute kidney injury: Subcapsular implantation technique. Am. J. Physiol. Renal Physiol. 2008, 1, F310–F314. [Google Scholar] [CrossRef]
- Ohashi, K.; Koyama, F.; Tatsumi, K.; Shima, M.; Park, F.; Nakajima, Y.; Okano, T. Functional life-long maintenance of engineered liver tissue in mice following transplantation under the kidney capsule. J. Tissue Eng. Regen. Med. 2009, 4, 141–148. [Google Scholar] [CrossRef]
- Roessger, A.; Denk, L.; Minuth, W.W. Potential of stem/progenitor cell cultures within polyester fleeces to regenerate renal tubules. Biomaterials 2009, 30, 3723–3732. [Google Scholar] [CrossRef] [PubMed]
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an-open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Minuth, W.W.; Denk, L.; Glashauser, A. Towards a Guided Regeneration of Renal Tubules at a Polyester Interstitium. Materials 2010, 3, 2369-2392. https://doi.org/10.3390/ma3042369
Minuth WW, Denk L, Glashauser A. Towards a Guided Regeneration of Renal Tubules at a Polyester Interstitium. Materials. 2010; 3(4):2369-2392. https://doi.org/10.3390/ma3042369
Chicago/Turabian StyleMinuth, Will W., Lucia Denk, and Anne Glashauser. 2010. "Towards a Guided Regeneration of Renal Tubules at a Polyester Interstitium" Materials 3, no. 4: 2369-2392. https://doi.org/10.3390/ma3042369