Biotechnology Applications of Tethered Lipid Bilayer Membranes


Abstract
:
1. Introduction

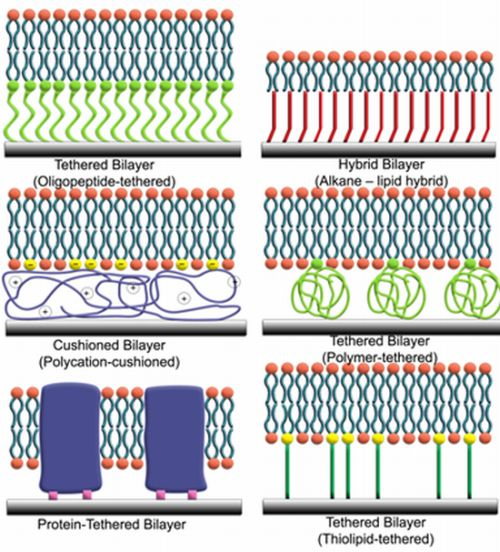{kind=link}
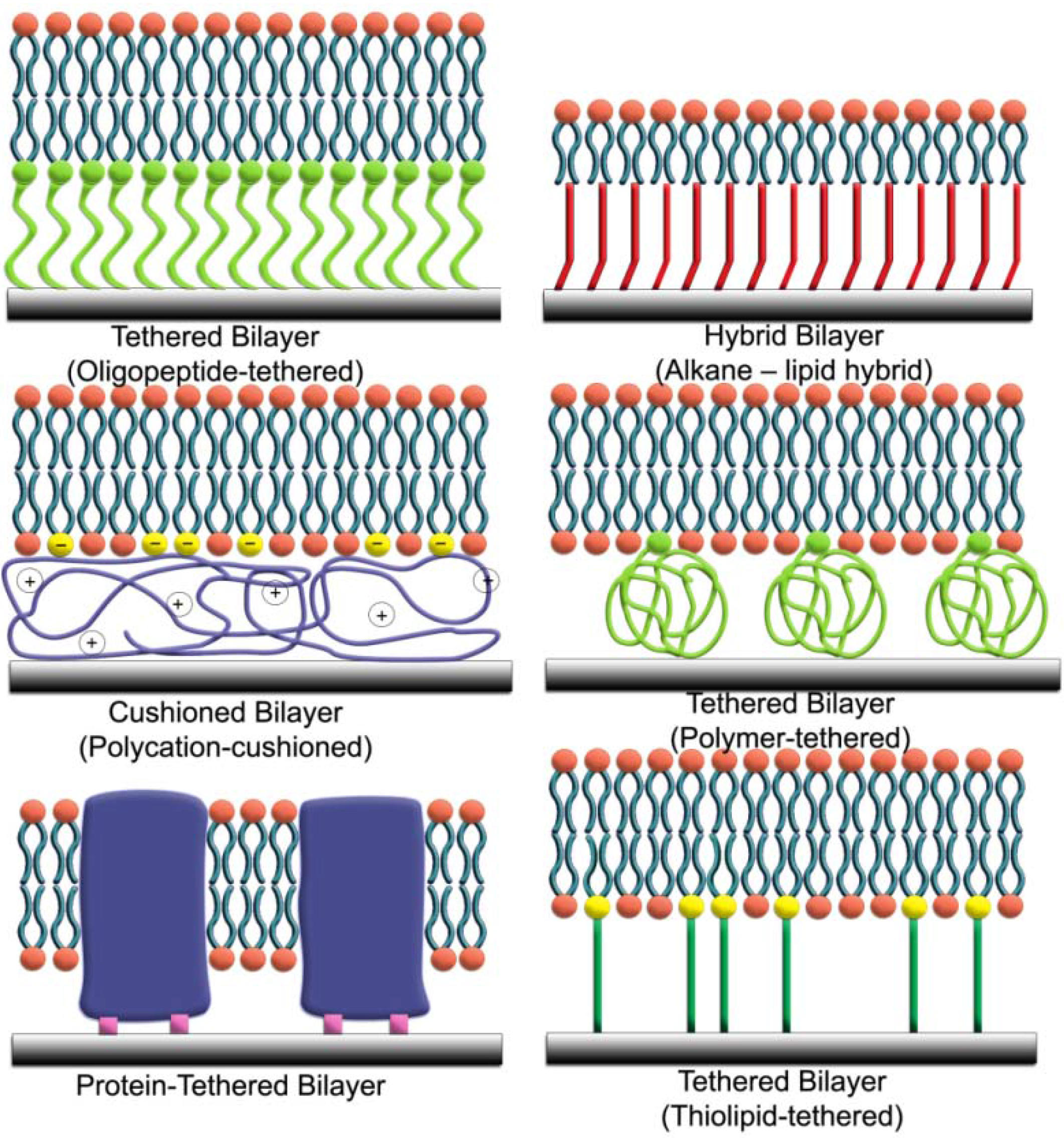{kind=link}
{kind=link}
{kind=link}
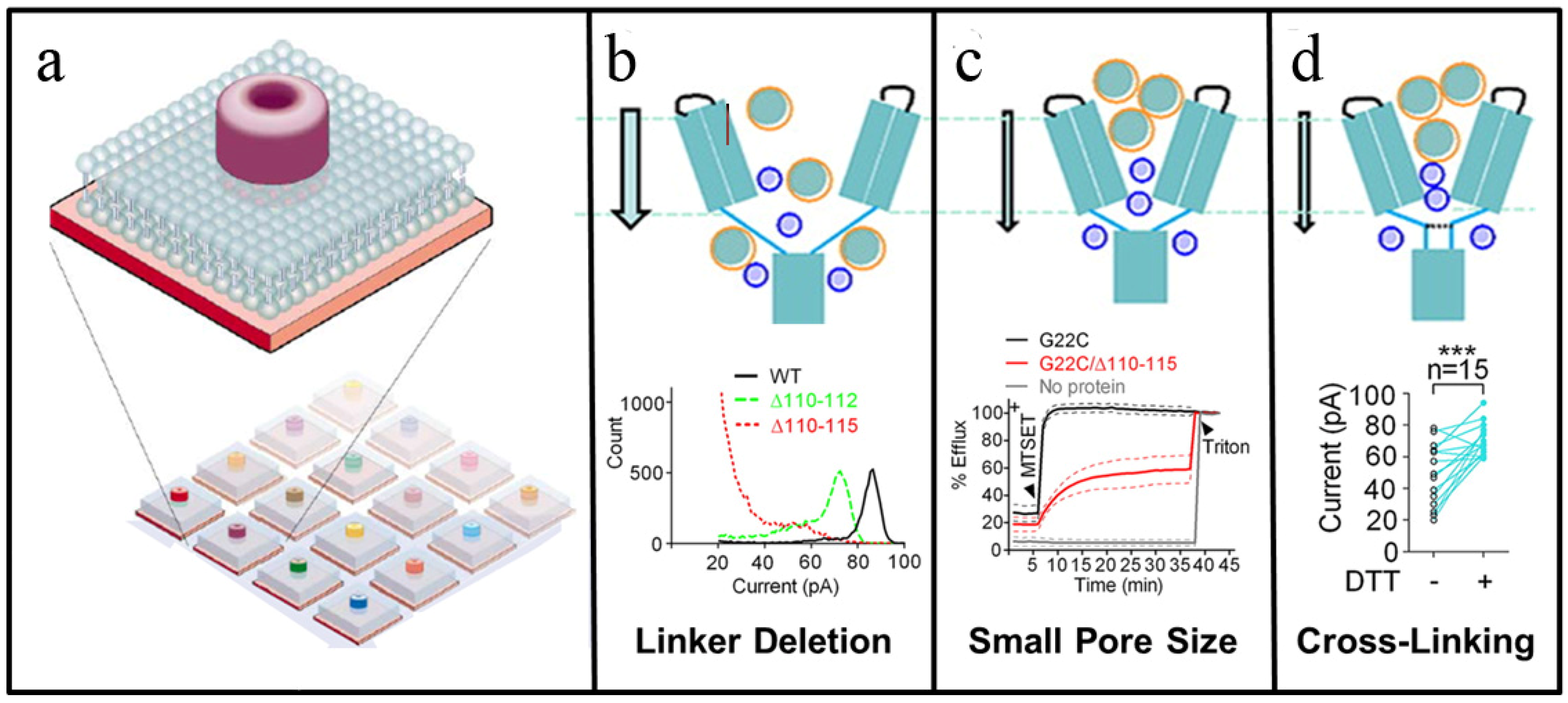{kind=link}
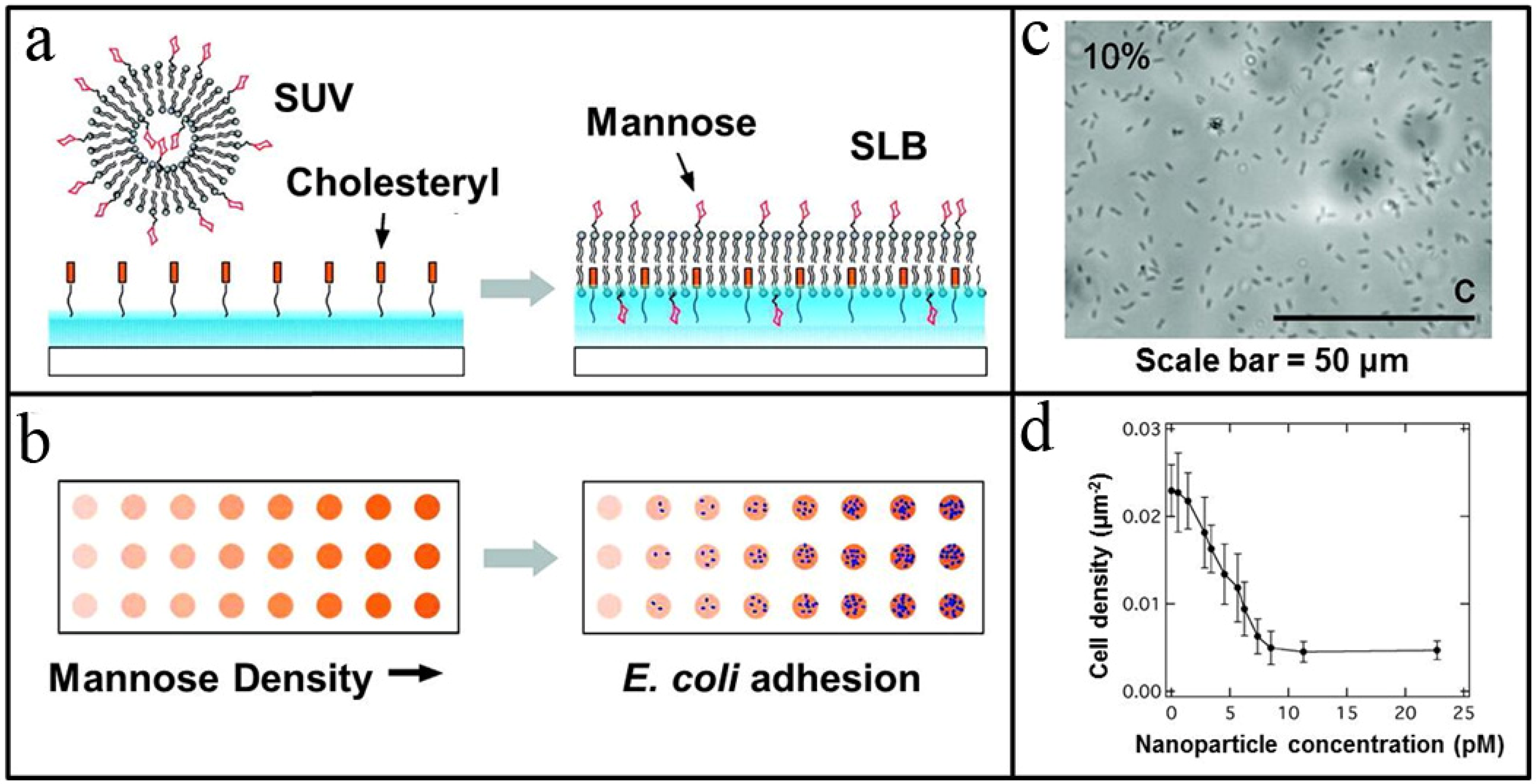{kind=link}
| Model System | Description | Surface-Sensitive Characterization | Electrical Sealing | Membrane Proteins | Air Stability |
|---|---|---|---|---|---|
| Lipid vesicles | Freely diffusing vesicles suspended in aqueous solution | No | Yes | Yes | No |
| Black lipid membrane | Lipid bilayer spans an open aperture | No | Yes | Yes | No |
| Supported lipid bilayer | Two-dimensional lipid bilayer physisorbed on solid support | Yes | Yes | No | Yes |
| Tethered lipid bilayer | Planar bilayer immobilized on solid support or soft cushion | Yes | Yes | Yes | Yes |
| Intact vesicle layer | Layer of intact vesicle physisorbed on solid support | Yes | No | Yes | No |
| Tethered vesicles | Intact vesicles immobilized to solid support or planar bilayer | Yes | No | Yes | No |
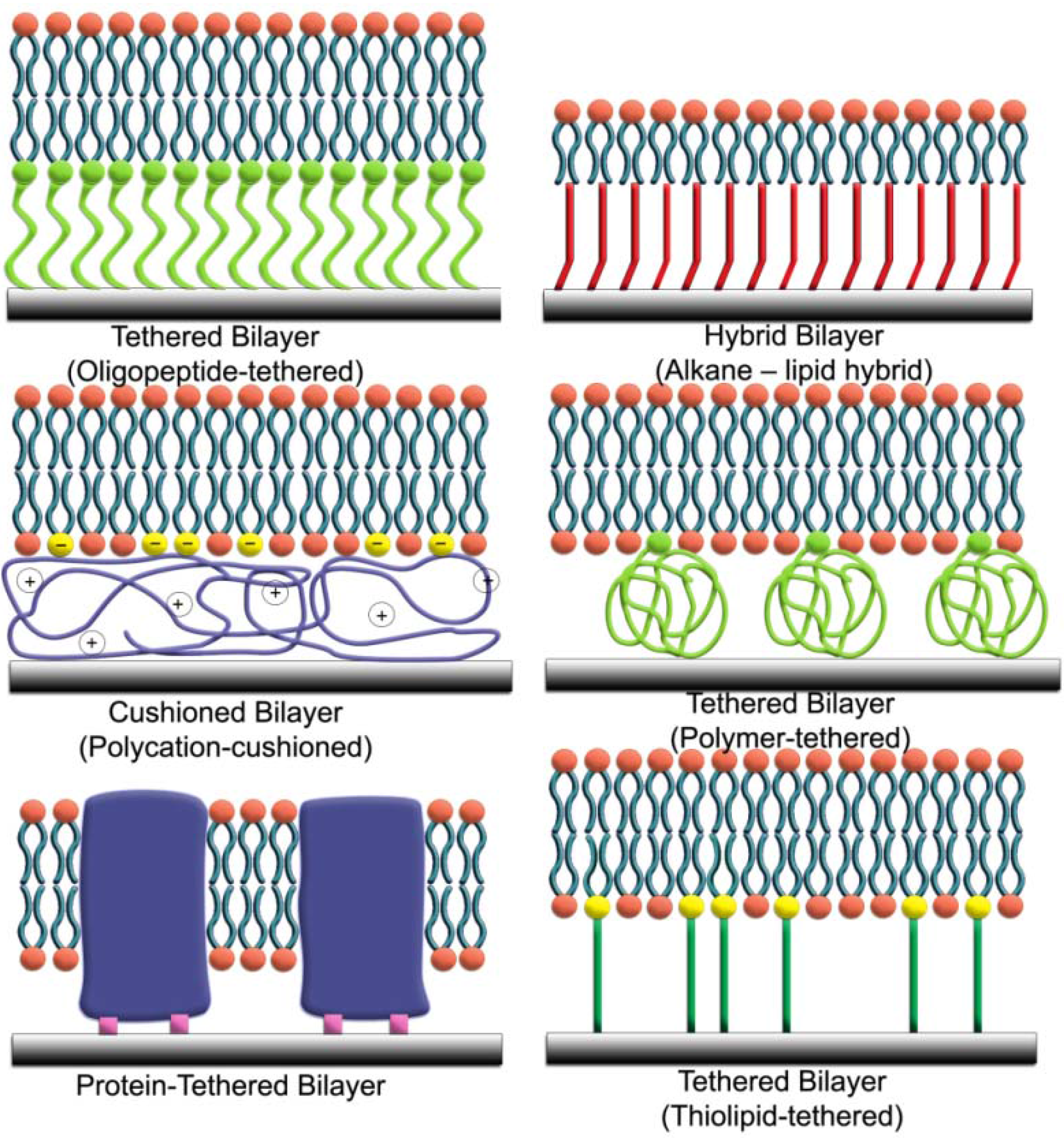
2. Characterization Methods
2.1. Functional Analysis of Membrane-Associated Peptides

2.2. Functional Analysis of Membrane-Associated Proteins
3. Biotechnology Applications
3.1. Ion Channel Biosensors
3.1.1. Influenza Virus Detection
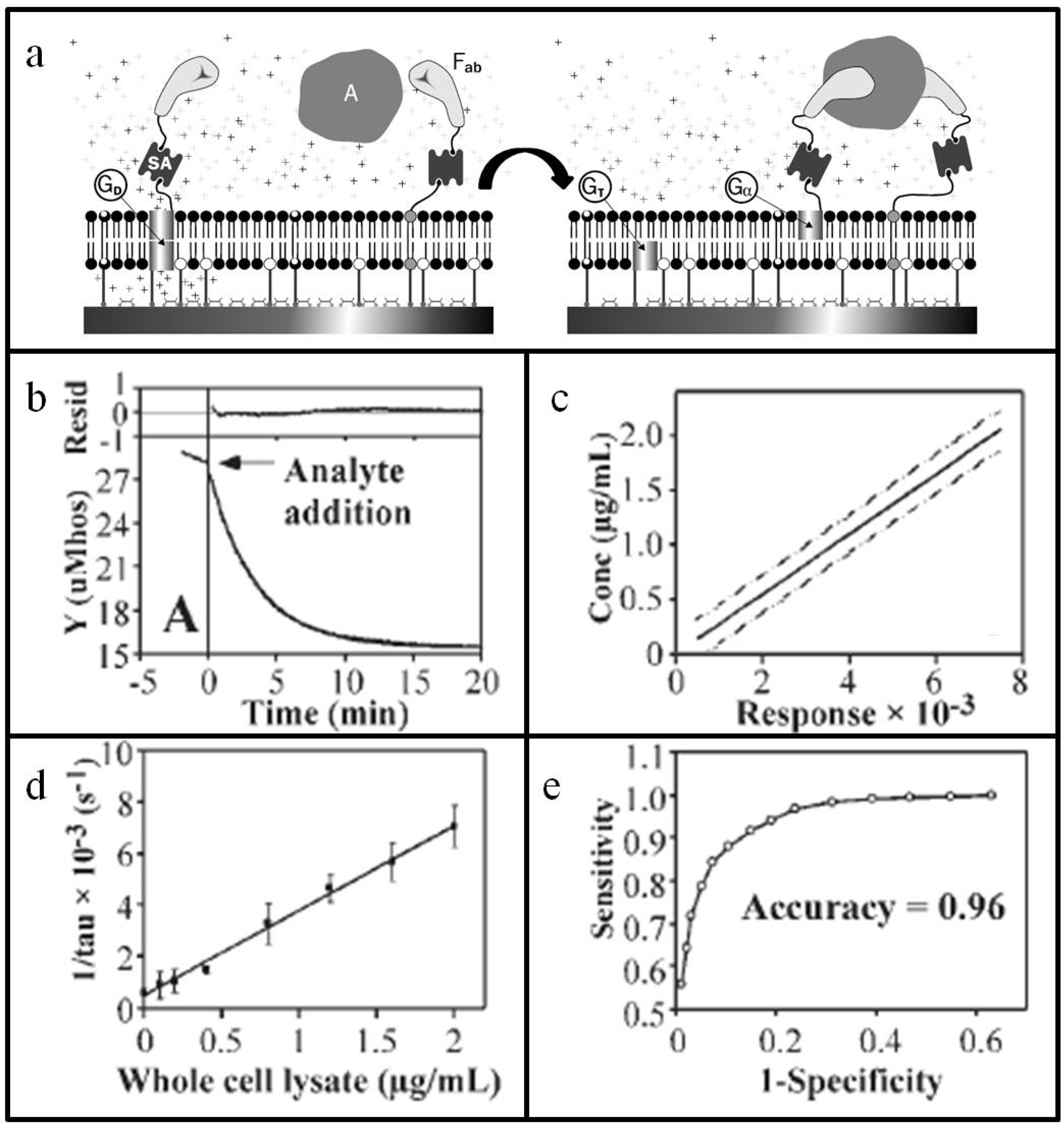
3.1.2. Stochastic Sensor Arrays

3.2. Air-Stable Bilayers

4. Perspectives
Acknowledgments
References
- Engelman, D.M. Membranes are more mosaic than fluid. Nature 2005, 438, 578–580. [Google Scholar] [CrossRef] [PubMed]
- Simons, K.; Toomre, D. Lipid rafts and signal transduction. Nat. Rev. Mol. Cell Biol. 2000, 1, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Singer, S.J.; Nicolson, G.L. The fluid mosaic model of the structure of cell membranes. Science 1972, 175, 720–731. [Google Scholar] [CrossRef] [PubMed]
- Israelachvili, J.N.; Marcelja, S.; Horn, R.G. Physical principles of membrane organization. Q. Rev. Biophys. 1980, 13, 121–200. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.H.; Boxer, S.G. Model membrane systems and their applications. Curr. Opin. Chem. Biol. 2007, 11, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Sackmann, E. Polymer-supported membranes as models of the cell surface. Nature 2005, 437, 656–663. [Google Scholar] [CrossRef] [PubMed]
- Sackmann, E. Supported membranes: Scientific and practical applications. Science 1996, 271, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Florin, E.L.; Gaub, H.E. Painted supported lipid membranes. Biophys. J. 1993, 64, 275–383. [Google Scholar] [CrossRef]
- Castellana, E.; Cremer, P. Solid supported lipid bilayers: From biophysical studies to sensor design. Surf. Sci. Rep. 2006, 61, 429–444. [Google Scholar] [CrossRef]
- Vontscharner, V.; McConnell, H. Physical properties of lipid monolayers on alkylated planar glass surfaces. Biophys. J. 1981, 36, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Plant, A.L. Self-assembled phospholipid/alkanethiol biomimetic bilayers on gold. Langmuir 1993, 9, 2764–2767. [Google Scholar] [CrossRef]
- Guidelli, R.; Becucci, L.; Dolfi, A.; Moncelli, M.R.; Buoninsegni, F.T. Some bioelectrochemical applications of self-assembled films on mercury. Solid States Ionics 2002, 150, 13–26. [Google Scholar] [CrossRef]
- Love, J.; Estroff, L.; Kriebel, J.; Nuzzo, R.; Whitesides, G. Self-assembled monolayers of thiolates on metals as a form of nanotechnology. Chem. Rev. 2005, 105, 1103–1169. [Google Scholar] [CrossRef] [PubMed]
- Demoz, A.; Harrison, D.J. Characterization and extremely low defect density hexadecanethiol monolayers on mercury surfaces. Langmuir 1993, 9, 1046–1050. [Google Scholar] [CrossRef]
- Silin, V.I.; Wieder, H.; Woodward, J.T.; Valincius, G.; Offenhausser, A.; Plant, A.L. The role of surface free energy on the formation of hybrid bilayer membranes. J. Am. Chem. Soc. 2002, 124, 14676–14683. [Google Scholar] [CrossRef] [PubMed]
- Naumann, C.A.; Prucker, O.; Lehmann, T.; Rühe, J.; Knoll, W.; Frank, C.W. The polymer-supported phospholipid bilayer: Tethering as a new approach to substrate−membrane stabilization. Biomacromolecules 2001, 3, 27–35. [Google Scholar] [CrossRef]
- Kügler, R.; Knoll, W. Polyelectrolyte-supported lipid membranes. Bioelectrochemistry 2002, 56, 175–178. [Google Scholar] [CrossRef] [PubMed]
- Naumann, R.; Schmidt, E.K.; Jonczyk, A.; Fendler, K.; Kadenbach, B.; Liebermann, T.; Offenhäusser, A.; Knoll, W. The peptide-tethered lipid membrane as a biomimetic system to incorporate cytochrome c oxidase in a functionally active form. Biosens. Bioelectron. 1999, 14, 641–662. [Google Scholar] [CrossRef]
- Schmidt, E.K.; Liebermann, T.; Kreiter, M.; Jonczyk, A.; Naumann, R.; Offenhäusser, A.; Neumann, E.; Kukol, A.; Maelicke, A.; Knoll, W. Incorporation of the acetylcholine receptor dimer from Torpedo californica in a peptide supported lipid membrane investigated by surface plasmon and fluorescence spectroscopy. Biosen. Bioelectron. 1998, 13, 585–591. [Google Scholar] [CrossRef]
- Naumann, R.; Baumgart, T.; Graber, P.; Jonczyk, A.; Offenhäusser, A.; Knoll, W. Proton transport through a peptide-tethered bilayer lipid membrane by the H+-ATP synthase from chloroplasts measured by impedance spectroscopy. Biosen. Bioelectron. 2002, 17, 25–34. [Google Scholar] [CrossRef]
- Raguse, B.; Braach-Maksvytis, V.; Cornell, B.A.; King, L.G.; Osman, P.D.J.; Pace, R.J.; Wieczorek, L. Tethered lipid bilayer membranes: Formation and ionic reservoir characterization. Langmuir 1998, 14, 648–659. [Google Scholar] [CrossRef]
- Woodhouse, G.; King, L.; Wieczorek, L.; Cornell, B. Kinetics of the competitive response of receptors immobilised to ion-channels which have been incorporated into a tethered bilayer. Faraday Discuss. 1998, 111, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Vanderah, D.; Pham, C.; Springer, S.; Silin, V.; Meuse, C. Characterization of a series of self-assembled monolayers of alkylated 1-thiaoligo (ethylene oxides)(4–8) on Gold. Langmuir 2000, 16, 6527–6532. [Google Scholar] [CrossRef]
- Cornell, B.; Krishna, G.; Osman, P.; Pace, R.; Wieczorek, L. Tethered-bilayer lipid membranes as a support for membrane-active peptides. Biochem. Soc. Trans. 2001, 29, 613–617. [Google Scholar] [CrossRef] [PubMed]
- Krishna, G.; Schulte, J.; Cornell, B.A.; Pace, R.; Wieczorek, L.; Osman, P.D. Tethered bilayer membranes containing ionic reservoirs: The interfacial capacitance. Langmuir 2001, 17, 4858–4866. [Google Scholar] [CrossRef]
- Atanasov, V.; Knorr, N.; Duran, R.S.; Ingebrandt, S.; Offenhäusser, A.; Knoll, W.; Köper, I. Membrane on a chip: A functional tethered lipid bilayer membrane on silicon oxide surfaces. Biophys. J. 2005, 89, 1780–1788. [Google Scholar] [CrossRef] [PubMed]
- Roskamp, R.; Vockenroth, I.; Eisenmenger, N.; Braunagel, J.; Koper, I. Functional tethered bilayer lipid membranes on aluminum oxide. Chemphyschem 2008, 9, 1920–1924. [Google Scholar] [CrossRef] [PubMed]
- Tien, H.T. Thickness and molecular organization of bimolecular lipid membranes in aqueous media. J. Mol. Biol. 1966, 16, 577–580. [Google Scholar] [CrossRef] [PubMed]
- Rothenhausler, B.; Knoll, W. Surface plasmon microscopy. Nature 1988, 332, 615–617. [Google Scholar] [CrossRef]
- Keller, C.; Kasemo, B. Surface specific kinetics of lipid vesicle adsorption measured with a quartz crystal microbalance. Biophys. J. 1998, 75, 1397–1402. [Google Scholar] [CrossRef]
- Mingeot-Leclercq, M.; Deleu, M.; Brasseur, R.; Dufrene, Y. Atomic force microscopy of supported lipid bilayers. Nat. Protoc. 2008, 3, 1654–1659. [Google Scholar] [CrossRef] [PubMed]
- Knoll, W.; Frank, C.W.; Heibela, C.; Naumann, R.; Offenhäusser, A.; Rühe1, J.; Schmidt, E.K.; Shenb, W.W.; Sinnera, A. Functional tethered lipid bilayers. Rev. Mol. Biotechnol. 2000, 74, 137–158. [Google Scholar] [CrossRef]
- Vanderah, D.; Meuse, C.; Silin, V.; Plant, A. Synthesis and characterization of self-assembled monolayers of alkylated 1-thiahexa(ethylene oxide) compounds on gold. Langmuir 1998, 14, 6916–6923. [Google Scholar] [CrossRef]
- Cornell, B.A.; Braach-Maksvytis, V.L.B.; King, L.G.; Osman, P.D.J.; Raguse, B.; Wieczorek, L.; Pace, R.J. A biosensor that uses ion-channel switches. Nature 1997, 387, 580–583. [Google Scholar] [CrossRef] [PubMed]
- Williams, L.M.; Evans, S.D.; Flynn, T.M.; Marsh, A.; Knowles, P.F.; Bushby, R.J.; Boden, N. Kinetics of the unrolling of small unilamellar phospholipid vesicles onto self-assembled monolayers. Langmuir 1997, 13, 751–757. [Google Scholar] [CrossRef]
- Lang, H.; Duschl, C.; Vogal, H. A new class of thiolipids for the attachment of lipid bilayers on gold surfaces. Langmuir 1994, 10, 197–210. [Google Scholar] [CrossRef]
- Vockenroth, I.K.; Atanasova, P.P.; Long, J.R.; Jenkins, A.T.; Knoll, W.; Köper, I. Functional incorporation of the pore forming segment of AChR M2 into tethered bilayer lipid membranes. Biochim. Biophys. Acta Biomembr. 2007, 1768, 1114–1120. [Google Scholar] [CrossRef]
- Naumann, R.L.C.; Nowak, C.; Knoll, W. Proteins in biomimetic membranes: Promises and facts. Soft Matter. 2011, 7, 9535–9548. [Google Scholar] [CrossRef]
- Giess, F.; Friedrich, M.; Heberle, J.; Naumann, R.; Knoll, W. The protein-tethered lipid bilayer: A novel mimic of the biological membrane. Biophys. J. 2004, 87, 3213–3220. [Google Scholar] [CrossRef] [PubMed]
- Nowak, C.; Laredo, T.; Gebert, J.; Lipkowski, J.; Gennis, R.B.; Ferguson-Miller, S.; Knoll, W.; Naumann, R.L.C. 2D-SEIRA spectroscopy to highlight conformational changes of the cytochrome c oxidase induced by direct electron transfer. Metallomics 2011, 3, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Hladky, S.B.; Haydon, D.A. Discreteness of conductance change in bimolecular lipid membranes in the presence of certain antibiotics. Nature 1970, 225, 451–453. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.K.; Cascão-Pereira, L.G.; Sala, R.F.; Holmes, S.P.; Ryan, K.J.; Becker, T. Ion channel switch array: A biosensor for detecting multiple pathogens. Ind. Biotechnol. 2005, 1, 26–31. [Google Scholar] [CrossRef]
- Oh, S.Y.; Cornell, B.A.; Smith, D.; Higgins, G.; Burrell, C.J.; Kok, T.W. Rapid detection of influenza a virus in clinical samples using an ion channel switch biosensor. Biosens. Bioelectron. 2008, 23, 1161–1165. [Google Scholar] [CrossRef] [PubMed]
- Avenet, P.; Lindemann, B. Amiloride-blockable sodium currents in isolated taste receptor cells. J. Membr. Biol. 1988, 105, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Bayley, H.; Cremer, P.S. Stochastic sensors inspired by biology. Nature 2001, 413, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Keizer, H.M.; Dorvel, B.R.; Andersson, M.; Fine, D.; Price, R.B.; Long, J.R.; Dodabalapur, A.; Köper, I.; Knoll, W.; Anderson, P.A.; Duran, R.S. Functional ion channels in tethered bilayer membranes—Implications for biosensors. ChemBioChem 2007, 8, 1246–1250. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M.; Keizer, H.M.; Zhu, C.; Fine, D.; Dodabalapur, A.; Duran, R.S. Detection of single ion channel activity on a chip using tethered bilayer membranes. Langmuir 2007, 23, 2924–2927. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M.; Okeyo, G.; Wilson, D.; Keizer, H.; Moe, P.; Blount, P.; Fine, D.; Dodabalapur, A.; Duran, R.S. Voltage-induced gating of the mechanosensitive MscL ion channel reconstituted in a tethered lipid bilayer membrane. Biosens. Bioelectron. 2008, 23, 919–923. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.M.; Wray, R.; Parker, J.; Wilson, D.; Duran, R.S.; Blount, P. Three routes to modulate the pore size of the MscL Channel/Nanovalve. ACS Nano 2011, 6, 1134–1141. [Google Scholar] [CrossRef]
- Plant, A.L. Supported hybrid bilayer membranes as rugged cell membrane mimics. Langmuir 1999, 15, 5128–5135. [Google Scholar] [CrossRef]
- Daniel, S.; Albertorio, F.; Cremer, P. Making lipid membranes rough, tough, and ready to hit the road. MRS Bull. 2006, 31, 536–540. [Google Scholar] [CrossRef]
- Cremer, P.; Boxer, S. Formation and spreading of lipid bilayers on planar glass supports. J. Phys. Chem. B 1999, 103, 2554–2559. [Google Scholar] [CrossRef]
- Ross, E.; Ross, E.E.; Rozanski, L.J.; Spratt, T.; Liu, S.; O’Brien, D.F.; Saavedra, S.S. Planar supported lipid bilayer polymers formed by vesicle fusion. 1. Influence of diene monomer structure and polymerization method on film properties. Langmuir 2003, 19, 1752–1765. [Google Scholar] [CrossRef]
- Holden, M.; Jung, S.; Yang, T.; Castellana, E.; Cremer, P. Creating fluid and air-stable solid supported lipid bilayers. J. Am. Chem. Soc. 2004, 126, 6512–6513. [Google Scholar] [CrossRef] [PubMed]
- Albertorio, F.; Diaz, A.J.; Yang, T.; Chapa, V.A.; Kataoka, S.; Castellana, E.T.; Cremer, P.S. Fluid and air-stable lipopolymer membranes for biosensor applications. Langmuir 2005, 21, 7476–7482. [Google Scholar] [CrossRef] [PubMed]
- Albertorio, F.; Chapa, V.; Chen, X.; Diaz, A.; Cremer, P. The alpha,alpha-(1→1) linkage of trehalose is key to anhydrobiotic preservation. J. Am. Chem. Soc. 2007, 129, 10567–10574. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Wang, Y.; Holtz, B.; Li, J.; Traaseth, N.; Veglia, G.; Stottrup, B.J.; Elde, R.; Pei, D.; Guo, A.; Zhu, X.Y. Fluidic and air-stable supported lipid bilayer and cell-mimicking microarrays. J. Am. Chem. Soc. 2008, 130, 6267–6271. [Google Scholar] [CrossRef] [PubMed]
- Henriksen, J.; Rowat, A.C.; Brief, E.; Hsueh, Y.W.; Thewalt, J.L.; Zuckermann, M.J.; Ipsen, J.H. Universal behavior of membranes with sterols. Biophys. J. 2006, 90, 1639–1649. [Google Scholar] [CrossRef] [PubMed]
- Simons, K.; Vaz, W. Model systems, lipid rafts, and cell membranes. Ann. Rev. Biophys. Biomol. Struct. 2004, 33, 269–295. [Google Scholar] [CrossRef]
- Lauer, S.; Goldstein, B.; Nolan, R.; Nolan, J. Analysis of cholera toxin-ganglioside interactions by flow cytometry. Biochemistry 2002, 41, 1742–1751. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.Y.; Holtz, B.; Wang, Y.; Wang, L.X.; Orndorff, P.E.; Guo, A. Quantitative glycomics from fluidic glycan microarrays. J. Am. Chem. Soc. 2009, 131, 13646–13650. [Google Scholar] [CrossRef] [PubMed]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Jackman, J.A.; Knoll, W.; Cho, N.-J. Biotechnology Applications of Tethered Lipid Bilayer Membranes. Materials 2012, 5, 2637-2657. https://doi.org/10.3390/ma5122637
Jackman JA, Knoll W, Cho N-J. Biotechnology Applications of Tethered Lipid Bilayer Membranes. Materials. 2012; 5(12):2637-2657. https://doi.org/10.3390/ma5122637
Chicago/Turabian StyleJackman, Joshua A., Wolfgang Knoll, and Nam-Joon Cho. 2012. "Biotechnology Applications of Tethered Lipid Bilayer Membranes" Materials 5, no. 12: 2637-2657. https://doi.org/10.3390/ma5122637