Context-Dependence of Urban Forest Vegetation Invasion Level and Alien Species’ Ecological Success


1
Institute of Dendrology, Polish Academy of Sciences, Parkowa 5, 62-035 Kórnik, Poland
2
Department of Game Management and Forest Protection, Faculty of Forestry, Poznań University of Life Sciences, Wojska Polskiego 71c, 60-625 Poznań, Poland
*
Author to whom correspondence should be addressed.
Forests 2019, 10(1), 26; https://doi.org/10.3390/f10010026
Submission received: 2 November 2018
/
Revised: 29 December 2018
/
Accepted: 2 January 2019
/
Published: 3 January 2019
(This article belongs to the Special Issue Forest Invasive Species: Spread, Impact and Management)
Abstract
:Research Highlights: Urban ecosystems are claimed to be more invaded than natural vegetation. Despite numerous studies, the patterns of alien species occurrence in urban forests are rarely linked to invasion ecology hypotheses. Background and Objectives: We assumed that patterns of invasion level (i.e., neophyte richness) and neophyte ecological success (cover) are context-dependent, i.e., depend on the type of vegetation, and that hypotheses connected with empty niche and biotic acceptance will have the strongest support in urban forests. We also tested biotic resistance, habitat filtering, disturbance, resource availability, and environmental heterogeneity hypotheses. Materials and Methods: Using a random forest algorithm, we tested the importance of factors related to invasion ecology hypotheses in a dataset of urban forest vegetation plots (n = 120). We studied seven types of forest plant communities occurring in Poznań (W Poland) and we assessed the vegetation’s taxonomic and functional composition. Results: We found that models of alien species richness and cover explained 28.5% and 35.0% of variance, respectively. Vegetation type was of the highest importance in both cases, suggesting that the occurrence of alien plant species is context-dependent. Resource availability and disturbance ecological indicator values were also of high importance. Conclusions: Our study supported resource availability and habitat filtering hypotheses as explanations of the level of invasion and ecological success of alien species in an urban forest, with partial support for the disturbance hypothesis. Our study revealed that predictors of invasion level are context-dependent, as patterns of alien species richness and cover differed among vegetation types. We highlight context-dependence of alien species invasion patterns in different vegetation types due to the habitat-forming role of dominant tree species and different availability of resources and disturbance levels, as well as different pools of native species. Thus, prevention and management of biological invasions in urban forests should account for forest vegetation type.
1. Introduction
Biological invasions are one of the most important threats to biodiversity worldwide [1,2,3]. Their impact is especially severe in urban ecosystems, where human influence and habitat destruction are responsible for the retreat of numerous indigenous species and spread of alien species [4,5,6]. In cities, alien species often replace more specialized native species, which are locally extinct and would otherwise provide substantial ecosystem services [7]. However, urban forests are claimed to be especially vulnerable to alien species invasion [8,9], due to forest fragmentation and extinction of forest specialists, which would normally be the main competitors and habitat transformations [10,11,12]. Moreover, cities are places of numerous introductions of large pools of alien species, usually ornamentals, which are one of the main pathways of invasion [13,14].
Invasive species may achieve ecological success due to several mechanisms. These are usually divided into three main groups of factors: propagule pressure, habitat invasibility, and species invasiveness [15,16,17]. Successful spread of alien species in their exotic ranges is claimed to be an effect of interactions among all three groups of factors [17,18,19]. Propagule pressure defines the immigration of new specimens and depends both on distance to the propagule source [20,21] and propagule quantity and quality [22,23]. Habitat invasibility refers to ecosystem susceptibility to invasion [18,24]. Species invasiveness refers to their life history traits [24,25]. Abundances of alien species in plant communities are shaped by joint effects of these three factors. For example, alien species richness is co-determined by both propagule pressure and invasibility [26,27]. Ecological success of alien species (single or as a group) is expressed by their biomass [23,28,29], density [21,30,31], or cover [32,33,34].
Although invasion ecology is a young branch of ecology, its dynamic development since Elton’s [35] classic study has yielded numerous conceptual frameworks and hypotheses explaining the phenomenon of biological invasions [17,36]. Among these hypotheses, most of them refer to characteristics of recipient ecosystems [18,24,37] or life histories of alien species [38,39]. Moreover, alien species impacts in ecosystems are context-dependent [40,41,42], which result from different conditions driving the performance of alien species. Although numerous studies analyzed patterns of alien species spread in cities [4,5,43], little is known about drivers of alien species richness and cover in urban forests, in contrast to forests growing on natural sites [23,32,44,45]
We aimed to assess the importance of vegetation characteristics, related to earlier invasion ecology hypotheses (Table 1), in shaping the level of invasion (i.e., alien species richness) and alien species ecological success (i.e., their cover in the understory vegetation) of urban forests in Poznań, Poland. In detail, we examined whether the variances in the level of invasion and ecological success of alien species, may be explained by vegetation-related factors. We propose the following three hypotheses: (H1) Patterns of both level of invasion and alien species ecological success are context-dependent, i.e., depend mostly on type of vegetation, which considers the forest plant community type to be of utmost importance. (H2) Due to biotic homogenization and extinctions of specialized species in urban environments [4,5], factors related to empty niches and biotic acceptance hypotheses will have the strongest support in urban forests. (H3) Forest vegetation more transformed by humans will host a higher richness and cover of alien species, according to the invasional meltdown concept [46].
2. Materials and Methods
2.1. Study Area
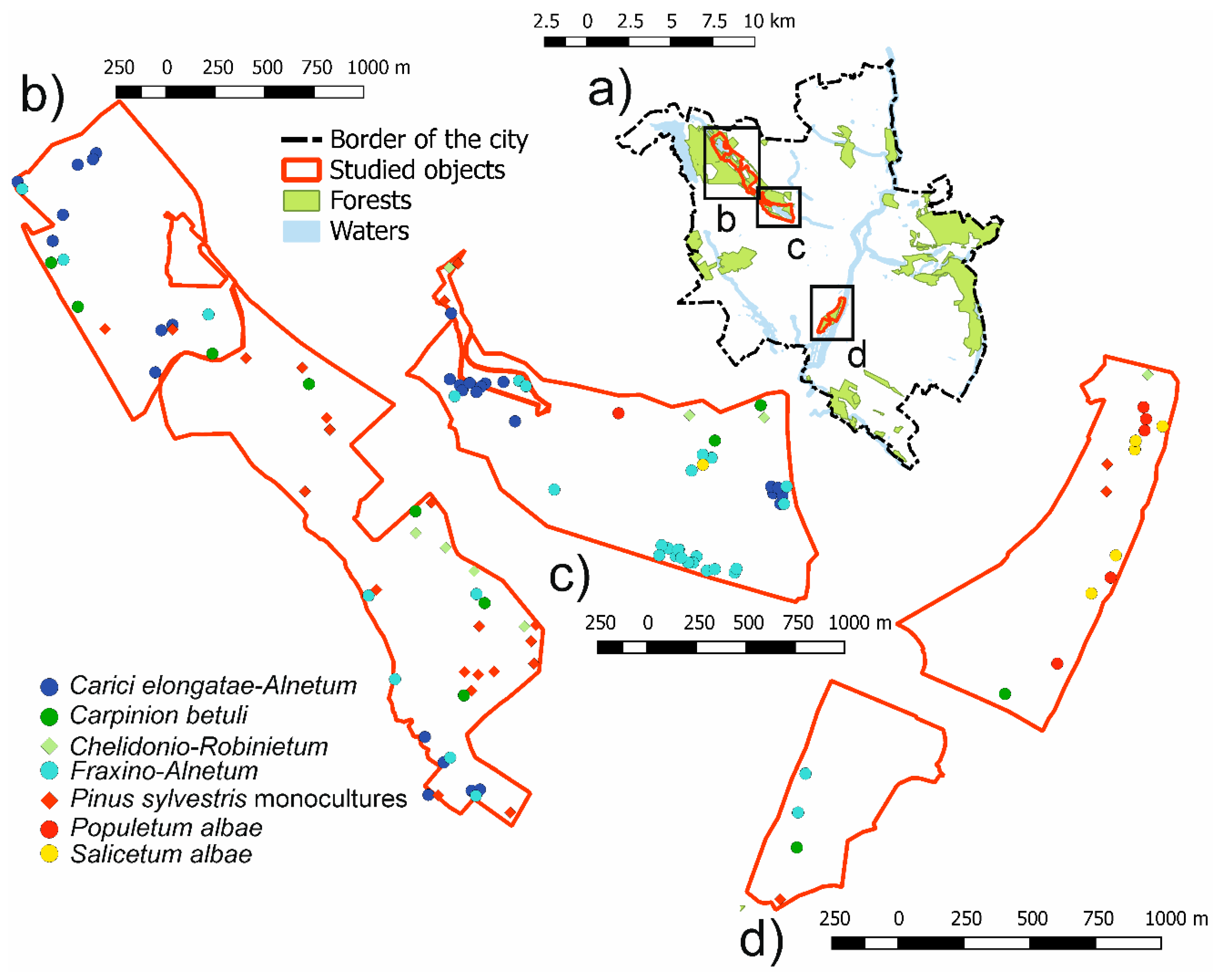
We conducted our study in urban forests of Poznań city (W Poland; 52°24′ N, 16°57′ E; 60–154 m a.s.l.). Poznań city has a population of 550,700 inhabitants and covers an area of 262 km2. Forests comprise 13% of the city area [54]. The climate of Poznań is temperate, with a mean annual temperature of 8.4 °C and mean annual precipitation of 521 mm in the years 1951–2010 [55]. We investigated forest vegetation in detail in four areas: three ecological lands (areas under low-restrictive nature conservation) and one complex of non-protected urban forest (Figure 1, Table S1) [56].
We sampled the main types of vegetation present in Poznań forests [54]. All these plant community types are related by successional and human impact dynamics (Tables S2 and S3). The most natural vegetation types comprise alder carrs (Carici elongatae-Alnetum W. Koch 1926 ex Schwickerath 1933) and riparian forests on organic soils (Fraxino-Alnetum W. Mat 1952). Both of these forests are dominated by Alnus glutinosa Gaertn. but Carici-elongatae-Alnetum comprises more meadow and wetland plant species, while Fraxino-Alnetum has more species typical of broadleaved forests. There are also two other types of riparian forests, connected with large river valleys, growing on mineral alluvial soils: Salicetum albae Issler 1926 and Populetum albae Br.-Bl. 1931. These plant associations differ in dominant tree species (Salix alba L. and Populus alba L., respectively), as well as flooding tolerance. Salicetum albae occurs in the bank zone and is tolerant of annual flooding, while Populetum albae occurs in the intermediate zone, usually flooded once per decade. Most of the pioneer species are found in understories of Salicetum albae. The most frequent type of broadleaved forest occurring on fertile soils are the remains of forests from the Carpinion betuli Issler 1931 em. Oberd. 1957 alliance (mostly degenerated forms of Galio-Carpinetum (R. Tx. 1937) Oberd. 1957), forest dominated by Quercus robus L., Carpinus betulus L., Tilia cordata Mill. and Acer platanoides L. We also sampled two types of human cultural forests: Chelidonio-Robinietum Jurko 1963 s.l. and Pinus sylvestris L. plantations. Chelidonio-Robinietum is a forest dominated by invasive Robinia pseudoacacia L. P. sylvestris plantations, which are usually replaced sites of Galio-Carpinetum.
2.2. Data Collection
We used data collected from 2012 to 2014 in the study area during vegetation inventories (Figure 1) [57,58,59,60]. Data for all vegetation plots were deposited in the Polish Vegetation Database [61]. From these plots, we selected plots sampled between July and September to account for only the summer part of seasonal dynamics. We did not account for spring aspects, as most of the alien species emerge during summer, and in temperate broadleaved forests, the peak of understory productivity is in summer [62,63]. For that reason, we may have overlooked early emerging native species. During each inventory, we first conducted field reconnaissance, where we recognized the vegetation types within study sites. After that, we located plots in the most frequently occurring types of forest vegetation to cover the widest range of variability possible. Although vegetation surveys conducted in this way are known to be biased by phytosociological attitudes and textbook standards [64,65,66], our study did not aim to describe vegetation types, thus we did not select plots in a preferential way. We did not use a randomized or systematic design due to the spatial heterogeneity of forest vegetation types in the study area. Thus, we established plots within the whole range of trophy and moisture to explore the relationships between alien species performance and ecosystem properties through the whole diversity of ecosystems present in the study area. Moreover, to eliminate observer effects, all vegetation surveys were conducted by one person (MKD).
During the vegetation surveys, we established 120 square or rectangular study plots (Figure 1), with an area of 80 to 200 m2, to cover homogenous vegetation patches (Table 2). Most of the plots had an area of 100 m2 (83.3%); smaller plots (2.5%) were used in cases of highly fragmented riparian forests and larger plots (14.2%) were used for homogenous Carpinion betuli forests. To separate plot size effects, we included plot size as a variable in our models. Within each plot, we noted covers of all plant species using the Braun-Blanquet scale. During the study, we tallied only the understory layer, as shrubs and trees in parts of the forests were artificially planted and thus did not account for spontaneous spread of alien species. However, we tallied their regeneration in the understory, which was considered as spontaneous spread. Among all study plots, we found 210 plant species, including six archaeophytes (alien species that arrived from prehistoric to medieval times), 24 neophytes (i.e., recent invaders), and 180 native species. We followed the nomenclature of Mirek et al. [67] and alien species status of Tokarska-Guzik et al. [68]. Due to the low abundance of archaeophytes (0–10.7% of cover, with an average of 0.5 ± 0.1% within the whole dataset) and their different patterns of invasion [13,26,68], we decided to exclude them from analyses and focus only on neophytes.
2.3. Data Analysis
We conducted all analyses in R software version 3.4.2 “Short Summer” (R Core Team, Vienna, Austria) [69]. For each vegetation plot we calculated all indices related to tested hypotheses (Table 1 and Table 3). Functional diversity components were calculated using the FD::dbFD() function [70] based on ecological indicator values [71] and functional traits from the BiolFlor [72] and LEDA [73] databases (Table S4). Functional richness describes the richness of functional plant types within a community and was standardized across the whole dataset (cover values of 0 to 1) and functional dispersion describes the mean distance to the centroid of a plant community within the functional trait hypervolume. These indices refer to niche filling by the species and whether plant community species composition is shaped by competition or environmental filtering [74,75]. We also calculated ecological indicator values (EIVs) for disturbance as proposed by Herben et al. [76]. These indices reflect disturbance severity, expressed as a fraction of community biomass being destroyed, and disturbance frequency, expressed as the logarithm of the mean time between disturbance events.
To assess joint effects of the plant community parameters studied and their importance in shaping cover and richness of alien species, we performed random forest analysis [68]. This model is a data-mining method, usually using randomly constructed regression trees, which predict explained values based on multiple predictors. Random forest analysis is suitable for analyzing cover and count data due to its robustness to non-normal distributions and collinearity of predictors [77]. To decrease overfitting of the models, we used repeated cross-validation (10 repeats, 10 times) in the caret::train() function [69]. To stabilize the variance and overcome the problem with uneven ranges of predictors, we centered and scaled predictors via reduction of the mean and dividing by the standard deviation. To assess the importance of variables and impacts of single predictors assuming constant levels of other predictors, we used two R packages for the machine learning model: DALEX [78] and ceterisParibus [79]. We assessed variable importance using the dropout loss of RMSE (root mean squared error), i.e. increase of prediction RMSE in the case of shuttling the particular variable within a dataset. To evaluate impacts of particular predictors on the modelled output, we used ceteris paribus plots, which is output modelled for each percentile of a particular predictor assuming a constant (mean) value of all remaining predictors. For comparison of alien species richness in vegetation types, we used generalized linear modeling (GLM) based on the Poisson distribution implemented in the stats::glm() function. For comparison of alien species cover, we used zero-inflated Beta regression, implemented in the gamlss::gamlss() function [80]. Both analyses were followed by Tukey’s posteriori tests.
3. Results
3.1. Predictors of Alien Species Richness
The model describing alien species richness explained 28.6% of the variability and had an RMSE of 0.97 (Figure 2). Vegetation type had the highest importance; predicted alien species richness was highest in Chelidonio-Robinietum, and lowest in Carici elongatae-Alnetum. We also found high importance of both functional richness and EIV for moisture and soil fertility. Although partial dependence plots for soil fertility EIV revealed a positive relationship with alien species richness, we did not find such a relationship with functional richness. The exception was in the lowest values of functional richness, where we found lower alien species richness. However, in plots with the highest functional richness we found lower alien species richness than in plots with intermediate functional richness. For moisture EIV, we found a negative relationship, but the effect size was low. Despite the low importance, we found decreasing alien species richness in plots with higher values of disturbance frequency EIV and increasing richness in plots with higher disturbance severity index. Plot size had the lowest importance and almost no impact on the predicted alien species richness.
3.2. Predictors of Alien Species Cover
The model describing alien species cover explained 35.0% of variability and had an RMSE of 9.15 (Figure 3). Vegetation type had the highest importance; predicted alien species cover was highest in Salicetum albae and lowest in Carpinion betuli and P. sylvestris monocultures. We also found high importance of both light EIV and disturbance frequency, both negatively correlated with alien species cover. Plot size also had high importance; however, this variable influenced less than 20% of study plots. We also found a weak positive relationship with alien species cover for soil fertility EIV, and negative relationships with EIVs for moisture. Despite the low importance, we found increasing alien species cover in plots with higher values of functional dispersion.
3.3. Differences among Vegetation Types
Our study revealed differences in mean alien species cover and richness among the vegetation types (Table 4 and Tables S5 and S6). We found the lowest alien species richness and cover in Carici elongatae-Alnetum and the highest in Salicetum albae and Chelidonio-Robinietum. Among 24 neophytes found in the vegetation plots, 11 occurred only in one vegetation type and four in two types. Only two alien species occurred in each vegetation type: Impatiens parviflora DC. and Prunus domestica L. (Table 5). However, the frequency of the former ranged from 74.2 to 90.0%, and the median cover ranged from 3 to 18%, while the latter was rarer (from 5.7 to 40.0% of plots) but occurred in a similar range of median cover (from 1 to 18%). We found the lowest number of alien species in Salicetum albae (seven species) and the highest in Fraxino-Alnetum (14). Over half of the alien species found were phanerophytes (15) and 20.8% (five species) were therophytes. According to the invasion status in Poland, we found 13 invasive species, with nine naturalized and two casual alien species (not naturalized).
4. Discussion
4.1. Patterns of Alien Species Invasions in Urban Forests
Our study revealed a moderate level of invasion (expressed as alien species richness) in urban forests of Poznań. In other urban forests, mean species richness of alien species is often higher, e.g., in spontaneous forests of Padova, mean alien species richness was 4.3 ± 2.1 [81]. The proportion of alien to native species in our study (14.3%) was lower than in neophyte communities in Riga and Kaunas (36.4–17.0%) [82], lower than proportion of alien species in European urban floras (40%) [83], and even lower than the proportion of alien species in the ecological lands studied (23.0%) [56]. This lower proportion may result from lower invasibility of forest ecosystems in comparison with ruderal and roadside vegetation [26,32] comprising important parts of alien species pools. As is typical of urban forests, most of the alien species in the understory were trees and shrubs [5,7,84]. This may be connected with proximity of gardens and urban green areas, which are important sources of alien species propagules [10,13,14].
4.2. Resource Availability and Habitat Filtering
Our study revealed that both alien species richness and cover depended on resource availability (light, moisture, and soil fertility EIV) and functional richness. These results support the increased resource availability and habitat filtering hypotheses. According to them, a high level of soil fertility supports invasive species encroachment into plant communities. Zerbe and Wirth [85] found more alien species in tree stands of P. sylvestris on fertile sites than on poor sites. Moreover, the most alien plant species are generalists, not occurring in ecosystems with nutrient limitations [52,86]. Thus, the pattern we found follows the continental pattern [32]: fewer alien species in poor habitats. On the other hand, species with high EIVs of soil fertility are usually good competitors: they invest in higher leaf area and growth rate [87]. For that reason, most alien species recorded in our study plots were functionally similar: they were phanerophytes or hemicryptophytes with usually competitive or mixed life strategies (Table 5). Although nine of them were annual plants (therophytes), which usually are ruderals, most of them had lower cover and frequencies than other alien species. The exceptions were two vines (Bilderdykia convolvulus and Echinocystis lobata) and Impatiens parviflora. In the case of B. convolvulus, the most probable key factor determining its frequency was residence time [39,88], and for E. lobata, its competitive life strategy is connected with high leaf area and seed mass [89].
The case of the most frequent alien species in the dataset, Impatiens parviflora, suggests support for the empty niche hypothesis. I. parviflora is functionally distinct from dominant species in the vegetation studied and has both ruderal (small seeds, autochory, and high growth rate) and competitive (high specific leaf area and canopy height) traits [90,91]. For that reason, this species is the most frequent alien species in European woodlands [32]. However, this species is a unique example of a functionally distinct alien species that reached ecological success.
Habitat filtering is manifested by moisture EIV, high levels of which usually prevent non-specialized alien species from encroaching into vegetation patches with high groundwater levels. As alien species are usually not adapted to such conditions, the wettest forest patches are the best-preserved sites in the city [12]. This is also a reason for low alien species richness and cover in the most extreme vegetation types [32,92,93]. Such extreme habitats, like poor wetlands or mountains, are usually characterized by low functional dispersion, i.e., low diversity of functional traits within plant communities (see Reference [93] for wetlands and Reference [75] for mountains). These sites are usually free from invasive species, with the exception of disturbed sites and roads [92,94].
4.3. Rich Does Not Get Richer?
Our study did not support the biotic resistance or the biotic acceptance hypotheses (Table 1). This result could be surprising due to numerous studies supporting both positive [10,47] and negative [44,95] relationships between native and alien species richness. Some studies also related the ecological success of single alien species to native species richness [37]. However, relationships between alien and native species are usually negative at small spatial scales and positive at larger scales [96,97]. In these studies, intermediate-size plots revealed either positive, negative, or no relationships. Herben et al. [96] revealed that relationship sign depends on size and proportions of both alien and native species pools. Thus, relatively small plots and small alien species pools would lead to a lack of distinct relationships. The size of species pools in urban environments is both decreased by local extinctions of specialized species and increased by alien species introductions. However, in urban forests, we may assume that woodland species are more likely to go extinct, decreasing native species pools [5,6,11].
4.4. Impact of Disturbance
Our results supported the disturbance hypothesis. Despite having a low importance, the effect sizes on partial dependence plots revealed higher alien species richness and cover in plots with higher disturbance severity EIV and lower richness with lower disturbance frequency EIV. Our dataset had only partial representation of the range of the disturbance gradient: from 8.3 to 50% of destroyed biomass in terms of severity and turnover from eight years to nine months. However, these ranges were the most frequent in the dataset prepared by Herben et al. [76] and even a small change of disturbance regime may create an opportunity for invasive species [53]. Our study is also in line with the hypotheses that alien species perform better in sites with intermediate disturbances [96,98]. Lower importance may also result from different responses of alien species guilds to disturbances: ruderal species benefit more from disturbances than competitors [99,100,101]. Another caveat is connected with indirect accounting for disturbances. Although our previous study [101] revealed that EIVs elaborated by Herben et al. [76] were useful in describing the disturbance gradient across different forest vegetation types, legacies of disturbance and resilience vary temporally [93,102]. For that reason, in snapshot-like studies (such as our study) only part of the dynamic states of an ecosystem may be recorded, which would decrease the importance of disturbances. However, this component is also included in the most important variable in our models, namely in vegetation types, due to their differences in disturbance regimes.
4.5. Differences among Vegetation Types
In our study, we expected the highest richness and cover of alien species in Chelidonio-Robinietum and P. sylvestris monocultures. In both vegetation types, we found the highest frequency and cover of alien species in shrub and tree layers (Tables S1 and S2), with two exceptions: we found more Acer negundo in Salicetum albae and more Cornus sericea in Fraxino-Alnetum. We found that alien tree and shrub species in the understory layer occurred in these vegetation types, where these species were present in tree and shrub layers, which highlights the importance of propagule pressure. Propagule pressure interacts with habitat invasibility [52], shaping the level of invasions, as the most invasive species emerge near propagule sources [21,22,103]. This pattern also seems to support the invasional meltdown hypothesis [46], as alien species modify habitats for other alien species, in addition to producing their own offspring. This resulted in the highest alien species richness in Chelidonio-Robinietum (Figure 2c).
The highest importance of vegetation type was connected with different alien species pools within different vegetation types. For example, Acer negundo invaded mainly riparian forests of Salicetum albae and Populetum albae, while Cornus sericea invaded mainly Fraxino-Alnetum. Such differentiation was reported earlier at continental [32,86], national [104,105], and regional [33,106] scales. The source of this pattern may be connected with the habitat-forming role of the dominant tree species, which are able to modify light availability [37,107] and nutrient cycling [108,109], and thus to shape dependent communities of different organismal groups [110,111], including understory vegetation [112,113]. On the other hand, tree species establishment is also driven by soil nutrients and moisture content [54,58,71]. The further explanation could be supported by observations of Jarošík et al. [43], who found that soil type was the most important factor influencing occurrence of neophytes. These impacts shape different opportunities for alien species, and as a consequence, cause the main source of context-dependency at the spatial scale studied. For that reason, we would expect that in, e.g., P. sylvestris plantations, increased light availability would increase alien species cover, in contrast to Carici elongatae-Alnetum, where due to high moisture, empty niches will be not be colonized by species of the local alien species pool.
4.6. Study Limitations
Despite the patterns revealed, our study may have been influenced by intercorrelation of predictors. Although data mining algorithms are resistant to that problem [77], collinearity of the variables may be a problem in interpretation of compound effects. For example, in broadleaved forests, soil nutrients EIV is usually correlated with moisture EIV [71]. These factors are crucial for tree stand species composition, leading to masking of the factors. Moreover, presence of some vegetation types is conditioned by disturbance, e.g., flood regime determines the presence of Salicetum albae and Populetum albae [10,54]. For that reason, our study design does not allow conclusions about single hypotheses. Our results may also be dependent on limiting the study to only the neophytes. Due to the low abundance and different responses to environmental factors compared to neophytes [5,26,68], we excluded archaeophytes. However, this group is established in cities and is sometimes also a threatened element of urban biodiversity [68].
5. Conclusions
Our study revealed that not only impacts, but also predictors, of invasion level and alien species ecological success, are context-dependent because patterns of alien species richness and cover differed among vegetation types. We did not confirm the significance of the biotic acceptance or the resistance hypotheses in an urban forest ecosystem at the scale of single study plots. We confirmed the important impact of resource availability and habitat filtering on alien species richness and cover. The most frequent alien species in the study area, Impatiens parviflora, can benefit from its functional distinctness from native competitors. This partially supports the empty niche hypothesis, and in conditions of increasing alien species pools, should warn us that new alien species may enter the forest via this mechanism in the future. Our analyses also partially supported the disturbance hypothesis, as we found higher alien species cover and numbers in plots with higher disturbance frequency and severity. These patterns were consistent across the vegetation types, which each provided unique contexts for alien species pools and rules of community assembly. However, we did not support the hypothesis of higher alien species richness and cover in the most transformed vegetation types. Instead, we highlight context-dependence of alien species invasion patterns in different vegetation types due to habitat-forming roles of dominant tree species. For that reason, both prevention and management of biological invasions in urban forests need to account for forest vegetation type. This context-dependence results from different resource availabilities and disturbance levels, as well as different pools of native species. Thus, understanding the factors limiting invasive species encroachment allows for maintaining less-invaded vegetation patches, e.g., by maintaining high groundwater levels in hydrogenic forests or maintaining closed canopies in forests where light availability limits alien species.
Supplementary Materials
The following are available online at https://www.mdpi.com/1999-4907/10/1/26/s1: Table S1. Overview of the forests studied in Poznań; Table S2. Tree layer species composition in eight vegetation types studied: frequency and median cover of species are listed in the table. Table S3. Shrub layer species composition in eight vegetation types studied: frequency and median cover of species are listed in the table. Table S4. Functional traits used for functional diversity components, their ecological significance, and completeness. Table S5. Result of Poisson GLM of alien species richness, estimated using the stats::glm() function. AIC0 refers to AIC of intercept-only model (null model). Table S6. Result of zero-inflated beta regression of alien species cover, estimated using the gamlss::gamlss() function. AIC0 refers to AIC of intercept-only model (null model).
Author Contributions
Conceptualization, methodology, study design, M.K.D., A.M.J.; data collection and analyses, draft preparation and editing, M.K.D.; supervision, A.M.J.
Funding
This research was supported by the Institute of Dendrology, Polish Academy of Sciences, Kórnik (Poland).
Acknowledgments
We kindly thank Lee E. Frelich (The University of Minnesota Center for Forest Ecology, USA) for valuable comments to the manuscript and linguistic revision of the text. We are thankful to the two anonymous reviewers for their thorough and valuable comments on the earlier draft of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Vitousek, P.M.; Mooney, H.A.; Lubchenco, J.; Melillo, J.M. Human domination of Earth’s ecosystems. Science 1997, 277, 494–499. [Google Scholar] [CrossRef]
- Chapin, F.S., III; Zavaleta, E.S.; Eviner, V.T.; Naylor, R.L.; Vitousek, P.M.; Reynolds, H.L.; Hooper, D.U.; Lavorel, S.; Sala, O.E.; Hobbie, S.E.; et al. Consequences of changing biodiversity. Nature 2000, 405, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, C.L.; Bacher, S.; Essl, F.; Hulme, P.E.; Jeschke, J.M.; Kühn, I.; Kumschick, S.; Nentwig, W.; Pergl, J.; Pyšek, P.; et al. Framework and guidelines for implementing the proposed IUCN Environmental Impact Classification for Alien Taxa (EICAT). Divers. Distrib. 2015, 21, 1360–1363. [Google Scholar] [CrossRef] [Green Version]
- Kowarik, I. Novel urban ecosystems, biodiversity, and conservation. Environ. Pollut. 2011, 159, 1974–1983. [Google Scholar] [CrossRef] [PubMed]
- Lososová, Z.; Chytrý, M.; Danihelka, J.; Tichý, L.; Ricotta, C. Biotic homogenization of urban floras by alien species: The role of species turnover and richness differences. J. Veg. Sci. 2016, 27, 452–459. [Google Scholar] [CrossRef]
- Chocholoušková, Z.; Pyšek, P. Changes in composition and structure of urban flora over 120 years: A case study of the city of Plzeň. Flora 2003, 198, 366–376. [Google Scholar] [CrossRef]
- Potgieter, L.J.; Gaertner, M.; Kueffer, C.; Larson, B.M.H.; Livingstone, S.W.; O’Farrell, P.J.; Richardson, D.M. Alien plants as mediators of ecosystem services and disservices in urban systems: A global review. Biol. Invasions 2017, 19, 3571–3588. [Google Scholar] [CrossRef]
- Godefroid, S.; Koedam, N. How important are large vs. small forest remnants for the conservation of the woodland flora in an urban context? Glob. Ecol. Biogeogr. 2003, 12, 287–298. [Google Scholar] [CrossRef]
- Kowarik, I. On the role of alien species in urban flora and vegetation. In Plant Invasions: General Aspects and Special Problems; Pyšek, P., Prach, K., Rejmánek, M., Wade, M., Eds.; SPB Academic Publishing: Amsterdam, Netherlands, 1995; pp. 85–103. [Google Scholar]
- Dyderski, M.K.; Gdula, A.K.; Jagodziński, A.M. “The rich get richer” concept in riparian woody species—A case study of the Warta River Valley (Poznań, Poland). Urban For. Urban Green. 2015, 14, 107–114. [Google Scholar] [CrossRef]
- Knapp, S.; Kühn, I.; Stolle, J.; Klotz, S. Changes in the functional composition of a Central European urban flora over three centuries. Perspect. Plant Ecol. Evol. Syst. 2010, 12, 235–244. [Google Scholar] [CrossRef]
- Dyderski, M.K.; Tyborski, J.; Jagodziński, A.M. The utility of ancient forest indicator species in urban environments: A case study from Poznań, Poland. Urban For. Urban Green. 2017, 27, 76–83. [Google Scholar] [CrossRef]
- Pergl, J.; Sádlo, J.; Petřík, P.; Danihelka, J.; Chrtek, J., Jr.; Hejda, M.; Moravcova, L.; Perglová, I.; Štajerová, K.; Pyšek, P. Dark side of the fence: Ornamental plants as a source of wild-growing flora in the Czech Republic. Preslia 2016, 88, 163–184. [Google Scholar]
- Haeuser, E.; Dawson, W.; Thuiller, W.; Dullinger, S.; Block, S.; Bossdorf, O.; Carboni, M.; Conti, L.; Dullinger, I.; Essl, F.; et al. European ornamental garden flora as an invasion debt under climate change. J. Appl. Ecol. 2018, 55, 2386–2395. [Google Scholar] [CrossRef]
- Richardson, D.M.; Pyšek, P.; Carlton, J.T. A compendium of esssential concepts and terminology in invasion ecology. In Fifty Years of Invasion Ecology: The Legacy of Charles Elton; Richardson, D.M., Ed.; Blackwell Publishing Ltd.: Oxford, UK, 2011; pp. 409–420. [Google Scholar]
- Blackburn, T.M.; Pyšek, P.; Bacher, S.; Carlton, J.T.; Duncan, R.P.; Jarošík, V.; Wilson, J.R.U.; Richardson, D.M. A proposed unified framework for biological invasions. Trends Ecol. Evol. 2011, 26, 333–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeschke, J.M. General hypotheses in invasion ecology. Divers. Distrib. 2014, 20, 1129–1134. [Google Scholar] [CrossRef]
- Davis, M.A.; Thompson, K.; Philip Grime, J. Invasibility: The local mechanism driving community assembly and species diversity. Ecography 2005, 28, 696–704. [Google Scholar] [CrossRef]
- Ricciardi, A.; Blackburn, T.M.; Carlton, J.T.; Dick, J.T.A.; Hulme, P.E.; Iacarella, J.C.; Jeschke, J.M.; Liebhold, A.M.; Lockwood, J.L.; MacIsaac, H.J.; et al. Invasion Science: A Horizon Scan of Emerging Challenges and Opportunities. Trends Ecol. Evol. 2017, 32, 464–474. [Google Scholar] [CrossRef] [Green Version]
- Pairon, M.; Jonard, M.; Jacquemart, A.-L. Modeling seed dispersal of black cherry, an invasive forest tree: How microsatellites may help? Can. J. For. Res. 2006, 36, 1385–1394. [Google Scholar] [CrossRef]
- Jagodziński, A.M.; Dyderski, M.K.; Rawlik, M.; Banaszczak, P. Plantation of coniferous trees modifies risk and size of Padus serotina (Ehrh.) Borkh. Invasion—Evidence from a Rogów Arboretum case study. For. Ecol. Manag. 2015, 357, 84–94. [Google Scholar] [CrossRef]
- Vanhellemont, M.; Verheyen, K.; Keersmaeker, L.; Vandekerkhove, K.; Hermy, M. Does Prunus serotina act as an aggressive invader in areas with a low propagule pressure? Biol. Invasions 2009, 11, 1451–1462. [Google Scholar] [CrossRef]
- Dyderski, M.K.; Jagodziński, A.M. Drivers of invasive tree and shrub natural regeneration in temperate forests. Biol. Invasions 2018, 20, 2363–2379. [Google Scholar] [CrossRef] [Green Version]
- Alpert, P.; Bone, E.; Holzapfel, C. Invasiveness, invasibility and the role of environmental stress in the spread of non-native plants. Perspect. Plant Ecol. Evol. Syst. 2000, 3, 52–66. [Google Scholar] [CrossRef]
- Richardson, D.M.; Pyšek, P. Plant invasions: Merging the concepts of species invasiveness and community invasibility. Prog. Phys. Geogr. 2006, 30, 409–431. [Google Scholar] [CrossRef]
- Chytrý, M.; Jarošík, V.; Pyšek, P.; Hájek, O.; Knollová, I.; Tichý, L.; Danihelka, J. Separating Habitat Invasibility by Alien Plants from the Actual Level of Invasion. Ecology 2008, 89, 1541–1553. [Google Scholar] [CrossRef] [PubMed]
- Pyšek, P.; Chytrỳ, M.; Jarošík, V. Habitats and land-use as determinants of plant invasions in the temperate zone of Europe. In Bioinvasions and Globalization: Ecology, Economics, Management and Policy; Perrings, C., Mooney, H., Williamson, M., Eds.; Oxford University Press: Oxford, UK, 2010; pp. 66–79. [Google Scholar]
- Annighöfer, P.; Mölder, I.; Zerbe, S.; Kawaletz, H.; Terwei, A.; Ammer, C. Biomass functions for the two alien tree species Prunus serotina Ehrh. and Robinia pseudoacacia L. in floodplain forests of Northern Italy. Eur. J. For. Res. 2012, 131, 1619–1635. [Google Scholar] [CrossRef]
- Franzese, J.; Urrutia, J.; García, R.A.; Taylor, K.; Pauchard, A. Pine invasion impacts on plant diversity in Patagonia: Invader size and invaded habitat matter. Biol. Invasions 2017, 19, 1015–1027. [Google Scholar] [CrossRef]
- Staska, B.; Essl, F.; Samimi, C. Density and age of invasive Robinia pseudoacacia modulate its impact on floodplain forests. Basic Appl. Ecol. 2014, 15, 551–558. [Google Scholar] [CrossRef]
- Riepšas, E.; Straigyté, L. Invasiveness and Ecological Effects of Red Oak (Quercus rubra L.) in Lithuanian Forests. Balt. For. 2008, 14, 122–130. [Google Scholar]
- Wagner, V.; Chytrý, M.; Jiménez-Alfaro, B.; Pergl, J.; Hennekens, S.; Biurrun, I.; Knollová, I.; Berg, C.; Vassilev, K.; Rodwell, J.S.; et al. Alien plant invasions in European woodlands. Divers. Distrib. 2017, 23, 969–981. [Google Scholar] [CrossRef] [Green Version]
- Chmura, D. Penetration and naturalisation of invasive alien plant species (neophytes) in woodlands of the Silesian Upland (southern Poland). Nat. Conserv. 2004, 60, 3–11. [Google Scholar]
- Catford, J.A.; Downes, B.J.; Gippel, C.J.; Vesk, P.A. Flow regulation reduces native plant cover and facilitates exotic invasion in riparian wetlands. J. Appl. Ecol. 2011, 48, 432–442. [Google Scholar] [CrossRef] [Green Version]
- Elton, C.S. The Ecology of Invasions by Animals and Plants; University of Chicago Press: Chicago, IL, USA, 1958. [Google Scholar]
- Catford, J.A.; Jansson, R.; Nilsson, C. Reducing redundancy in invasion ecology by integrating hypotheses into a single theoretical framework. Divers. Distrib. 2009, 15, 22–40. [Google Scholar] [CrossRef] [Green Version]
- Knight, K.S.; Oleksyn, J.; Jagodziński, A.M.; Reich, P.B.; Kasprowicz, M. Overstorey tree species regulate colonization by native and exotic plants: A source of positive relationships between understorey diversity and invasibility. Divers. Distrib. 2008, 14, 666–675. [Google Scholar] [CrossRef]
- Baker, H.G. The Evolution of Weeds. Ann. Rev. Ecol. Syst. 1974, 5, 1–24. [Google Scholar] [CrossRef]
- Pyšek, P.; Manceur, A.M.; Alba, C.; McGregor, K.F.; Pergl, J.; Štajerová, K.; Chytrý, M.; Danihelka, J.; Kartesz, J.; Klimešová, J.; et al. Naturalization of central European plants in North America: Species traits, habitats, propagule pressure, residence time. Ecology 2015, 96, 762–774. [Google Scholar] [CrossRef]
- Vilà, M.; Espinar, J.L.; Hejda, M.; Hulme, P.E.; Jarošík, V.; Maron, J.L.; Pergl, J.; Schaffner, U.; Sun, Y.; Pyšek, P. Ecological impacts of invasive alien plants: A meta-analysis of their effects on species, communities and ecosystems. Ecol. Lett. 2011, 14, 702–708. [Google Scholar] [CrossRef]
- Hulme, P.E.; Pyšek, P.; Jarošík, V.; Pergl, J.; Schaffner, U.; Vilà, M. Bias and error in understanding plant invasion impacts. Trends Ecol. Evol. 2013, 28, 212–218. [Google Scholar] [CrossRef]
- González-Moreno, P.; Diez, J.M.; Ibáñez, I.; Font, X.; Vilà, M. Plant invasions are context-dependent: Multiscale effects of climate, human activity and habitat. Divers. Distrib. 2014, 20, 720–731. [Google Scholar] [CrossRef]
- Jarošík, V.; Pyšek, P.; Kadlec, T. Alien plants in urban nature reserves: From red-list species to future invaders? NeoBiota 2011, 10, 27–46. [Google Scholar]
- Iannone, B.V.; Potter, K.M.; Hamil, K.-A.D.; Huang, W.; Zhang, H.; Guo, Q.; Oswalt, C.M.; Woodall, C.W.; Fei, S. Evidence of biotic resistance to invasions in forests of the Eastern USA. Landsc. Ecol. 2016, 31, 85–99. [Google Scholar] [CrossRef]
- Gaertner, M.; Wilson, J.R.U.; Cadotte, M.W.; MacIvor, J.S.; Zenni, R.D.; Richardson, D.M. Non-native species in urban environments: Patterns, processes, impacts and challenges. Biol. Invasions 2017, 19, 3461–3469. [Google Scholar] [CrossRef]
- Simberloff, D.; Von Holle, B. Positive interactions of nonindigenous species: Invasional meltdown? Biol. Invasions 1999, 1, 21–32. [Google Scholar] [CrossRef]
- Stohlgren, T.J.; Jarnevich, C.; Chong, G.W.; Evangelista, P.H.; Pyšek, P.; Kaplan, Z.; Richardson, D.M. Scale and plant invasions: A theory of biotic acceptance. Preslia 2006, 78, 405–426. [Google Scholar]
- Darwin, C. On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life; John Murray: London, UK, 1859; ISBN 978-83-235-0531-0. [Google Scholar]
- Melbourne, B.A.; Cornell, H.V.; Davies, K.F.; Dugaw, C.J.; Elmendorf, S.; Freestone, A.L.; Hall, R.J.; Harrison, S.; Hastings, A.; Holland, M.; et al. Invasion in a heterogeneous world: Resistance, coexistence or hostile takeover? Ecol. Lett. 2007, 10, 77–94. [Google Scholar] [CrossRef] [PubMed]
- Hierro, J.L.; Maron, J.L.; Callaway, R.M. A biogeographical approach to plant invasions: The importance of studying exotics in their introduced and native range. J. Ecol. 2005, 93, 5–15. [Google Scholar] [CrossRef]
- Sher, A.A.; Hyatt, L.A. The Disturbed Resource-Flux Invasion Matrix: A New Framework for Patterns of Plant Invasion. Biol. Invasions 1999, 1, 107–114. [Google Scholar] [CrossRef]
- Davis, M.A.; Grime, J.P.; Thompson, K. Fluctuating resources in plant communities: A general theory of invasibility. J. Ecol. 2000, 88, 528–534. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Huenneke, L.F. Disturbance, Diversity, and Invasion: Implications for Conservation. Conserv. Biol. 1992, 6, 324–337. [Google Scholar] [CrossRef]
- Jackowiak, B. Poznań. In Plants and Habitats of European Cities; Müller, N., Kelcey, J.G., Eds.; Springer: New York, NY, USA, 2011; pp. 363–405. ISBN 978-0-387-89683-0. [Google Scholar]
- Statistical Office in Poznań. Statistical Yearbook of Poznań City; Statistical Office in Poznań: Poznań, Poland, 2013.
- Dyderski, M.K.; Wrońska-Pilarek, D.; Jagodziński, A.M. Ecological lands for conservation of vascular plant diversity in the urban environment. Urban Ecosyst. 2016, 20, 639–650. [Google Scholar] [CrossRef] [Green Version]
- Dyderski, M.K.; Wrońska-Pilarek, D. Szata roślinna nowo powstałego użytku ekologicznego “Strzeszyn” w Poznaniu i stan jej zachowania. Nauk. Przyr. Technol. 2015, 9, 39. [Google Scholar]
- Dyderski, M.K.; Gdula, A.K.; Wrońska-Pilarek, D. Roślinność nowo utworzonych użytków ekologicznych Bogdanka I i Bogdanka II w Poznaniu. Acta Scientiarum Polonorum Silvarum Colendarum Ratio et Industria Lignaria 2014, 13, 22–37. [Google Scholar]
- Dyderski, M.K.; Wrońska-Pilarek, D. Szata roślinna nowo powstałych użytków ekologicznych “Dębina I” i “Dębina II” w Poznaniu. Nauk. Przyr. Technol. 2015, 9, 46. [Google Scholar]
- Dyderski, M.K.; Gdula, A.K.; Wrońska-Pilarek, D. Aktualny stan szaty roślinnej terenów wokół jeziora Rusałka w Poznaniu w warunkach antropopresji. Acta Scientiarum Polonorum Silvarum Colendarum Ratio et Industria Lignaria 2016, 15, 229–246. [Google Scholar] [CrossRef]
- Kącki, Z.; Śliwiński, M. The Polish Vegetation Database: Structure, resources and development. Acta Soc. Bot. Pol. 2012, 81, 75–79. [Google Scholar] [CrossRef]
- Tremblay, N.O.; Larocque, G.R. Seasonal dynamics of understory vegetation in four eastern Canadian forest types. Int. J. Plant Sci. 2001, 162, 271–286. [Google Scholar] [CrossRef]
- Jagodziński, A.M.; Dyderski, M.K.; Rawlik, K.; Kątna, B. Seasonal variability of biomass, total leaf area and specific leaf area of forest understory herbs reflects their life strategies. For. Ecol. Manag. 2016, 374, 71–81. [Google Scholar] [CrossRef]
- Czapiewska, N.; Paź, S.; Dyderski, M.K.; Jagodziński, A.M. Ciągłość kompozycji florystycznej pomiędzy dwoma zbiorowiskami roślinnymi—Carici elongatae-Alnetum oraz Fraxino-Alnetum. Leśn. Pr. Badaw. 2017, 78, 285–296. [Google Scholar]
- Holeksa, J.; Woźniak, G. Biased vegetation patterns and detection of vegetation changes using phytosociological databases. A case study in the forests of the Babia Góra National Park (the West Carpathians, Poland). Phytocoenologia 2005, 35, 1–18. [Google Scholar] [CrossRef]
- Lepš, J.; Hadincová, V. How reliable are our vegetation analyses? J. Veg. Sci. 1992, 3, 119–124. [Google Scholar] [CrossRef]
- Mirek, Z.; Piękoś-Mirkowa, H.; Zając, A.; Zając, M. Flowering Plants and Pteridophytes of Poland. A Checklist; W. Szafer Institute of Botany, PAN: Kraków, Poland, 2003. [Google Scholar]
- Tokarska-Guzik, B.; Dajdok, Z.; Zając, M.; Zając, A.; Urbisz, A.; Danielewicz, W.; Hołdyński, C. Rośliny Obcego Pochodzenia w Polsce ze Szczególnym Uwzględnieniem Gatunków Inwazyjnych; Generalna Dyrekcja Ochrony Środowiska: Warszawa, Poland, 2012; ISBN 978-83-62940-33-2.
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Laliberté, E.; Legendre, P.; Shipley, B. FD: Measuring Functional Diversity (FD) from Multiple Traits, and Other tools for Functional Ecology; 2014. Available online: https://cran.r-project.org/web/packages/FD/index.html (accessed on 28 December 2018).
- Ellenberg, H.; Leuschner, C. Vegetation Mitteleuropas Mit Den Alpen in Ökologischer, Dynamischer und Historischer Sicht; UTB: Stuttgart, Germany, 2010. [Google Scholar]
- Klotz, S.; Kühn, I.; Durka, W. BIOLFLOR—Eine Datenbank zu Biologisch-Ökologischen Merkmalen der Gefäßpflanzen in Deutschland. Schriftenreihe für Vegetationskunde; Schriftenreihe für Vegetationskunde; Bundesamt für Naturschutz: Bonn, Germany, 2002. [Google Scholar]
- Kleyer, M.; Bekker, R.M.; Knevel, I.C.; Bakker, J.P.; Thompson, K.; Sonnenschein, M.; Poschlod, P.; Van Groenendael, J.M.; Klimeš, L.; Klimešová, J.; et al. The LEDA Traitbase: A database of life-history traits of the Northwest European flora. J. Ecol. 2008, 96, 1266–1274. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P. A distance-based framework for measuring functional diversity from multiple traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Czortek, P.; Delimat, A.; Dyderski, M.K.; Zięba, A.; Jagodziński, A.M.; Jaroszewicz, B. Climate change, tourism and historical grazing influence the distribution of Carex lachenalii Schkuhr—A rare arctic-alpine species in the Tatra Mts. Sci. Total Environ. 2018, 618, 1628–1637. [Google Scholar] [CrossRef] [PubMed]
- Herben, T.; Chytrý, M.; Klimešová, J. A quest for species-level indicator values for disturbance. J. Veg. Sci. 2016, 27, 628–636. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Biecek, P. DALEX: Descriptive mAchine Learning EXplanations. 2018. Available online: https://cran.r-project.org/web/packages/DALEX/index.html (accessed on 28 December 2018).
- Biecek, P. CeterisParibus: Ceteris Paribus Plots (What-If Plots) for a Single Observation. 2018. Available online: https://cran.r-project.org/web/packages/ceterisParibus/index.html (accessed on 28 December 2018).
- Rigby, R.A.; Stasinopoulos, D.M. Generalized additive models for location, scale and shape. J. R. Stat. Soc. C (Appl. Stat.) 2005, 54, 507–554. [Google Scholar] [CrossRef]
- Sitzia, T.; Campagnaro, T.; Weir, R.G. Novel woodland patches in a small historical Mediterranean city: Padova, Northern Italy. Urban Ecosyst. 2016, 19, 475–487. [Google Scholar] [CrossRef]
- Marozas, V.; Cekstere, G.; Laivins, M.; Straigyte, L. Comparison of neophyte communities of Robinia pseudoacacia L. and Acer negundo L. in the eastern Baltic Sea region cities of Riga and Kaunas. Urban For. Urban Green. 2015, 14, 826–834. [Google Scholar] [CrossRef]
- Pyšek, P. Alien and native species in Central European urban floras: A quantitative comparison. J. Biogeogr. 1998, 25, 155–163. [Google Scholar] [CrossRef]
- Kowarik, I.; Lippe, M.; Cierjacks, A. Prevalence of alien versus native species of woody plants in Berlin differs between habitats and at different scales. Preslia 2013, 85, 113–132. [Google Scholar]
- Zerbe, S.; Wirth, P. Non-indigenous plant species and their ecological range in Central European pine (Pinus sylvestris L.) forests. Ann. For. Sci. 2006, 63, 189–203. [Google Scholar] [CrossRef]
- Lambdon, P.W.; Pyšek, P.; Basnou, C.; Hejda, M.; Arianoutsou, M.; Essl, F.; Jarošík, V.; Pergl, J.; Winter, M.; Anastasiu, P.; et al. Alien flora of Europe: Species diversity, temporal trends, geographical patterns and research needs. Preslia 2008, 80, 101–149. [Google Scholar]
- Pierce, S.; Brusa, G.; Vagge, I.; Cerabolini, B.E.L. Allocating CSR plant functional types: The use of leaf economics and size traits to classify woody and herbaceous vascular plants. Funct. Ecol. 2013, 27, 1002–1010. [Google Scholar] [CrossRef]
- Pyšek, P.; Křivánek, M.; Jarošík, V. Planting intensity, residence time, and species traits determine invasion success of alien woody species. Ecology 2009, 90, 2734–2744. [Google Scholar] [CrossRef]
- Dylewski, Ł.; Maćkowiak, Ł.; Myczko, Ł. Physical defence of the wild cucumber Echinocystis lobata in an invasive range changing seed removal by rodents. Plant Ecol. 2018, 219, 863–873. [Google Scholar] [CrossRef]
- Chmura, D. Biology and Ecology of an Invasion of Impatiens Parviflora DC in Natural and Semi-Natural Habitats; Wydawnictwo ATH: Bielsko-Biała, Poland, 2014. [Google Scholar]
- Godefroid, S.; Koedam, N. Comparative ecology and coexistence of introduced and native congeneric forest herbs: Impatiens parviflora and I. noli-tangere. Plant Ecol. Evol. 2010, 143, 119–127. [Google Scholar] [CrossRef]
- Lembrechts, J.J.; Pauchard, A.; Lenoir, J.; Nuñez, M.A.; Geron, C.; Ven, A.; Bravo-Monasterio, P.; Teneb, E.; Nijs, I.; Milbau, A. Disturbance is the key to plant invasions in cold environments. Proc. Natl. Acad. Sci. USA 2016, 113, 14061–14066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyderski, M.K.; Czapiewska, N.; Zajdler, M.; Tyborski, J.; Jagodziński, A.M. Functional diversity, succession, and human-mediated disturbances in raised bog vegetation. Sci. Total Environ. 2016, 562, 648–657. [Google Scholar] [CrossRef] [PubMed]
- Pauchard, A.; Kueffer, C.; Dietz, H.; Daehler, C.C.; Alexander, J.; Edwards, P.J.; Arévalo, J.R.; Cavieres, L.A.; Guisan, A.; Haider, S.; et al. Ain’t no mountain high enough: Plant invasions reaching new elevations. Front. Ecol. Environ. 2009, 7, 479–486. [Google Scholar] [CrossRef]
- Parker, J.D.; Richie, L.J.; Lind, E.M.; Maloney, K.O. Land use history alters the relationship between native and exotic plants: The rich don’t always get richer. Biol. Invasions 2010, 12, 1557–1571. [Google Scholar] [CrossRef]
- Herben, T.; Mandák, B.; Bímová, K.; Münzbergová, Z. Invasibility and species richness of a community: A neutral model and a survey of published data. Ecology 2004, 85, 3223–3233. [Google Scholar] [CrossRef]
- Davies, K.F.; Chesson, P.; Harrison, S.; Inouye, B.D.; Melbourne, B.A.; Rice, K.J. Spatial heterogeneity explains the scale dependence of the native-exotic diversity relationship. Ecology 2005, 86, 1602–1610. [Google Scholar] [CrossRef]
- Fox, J.F. Intermediate-Disturbance Hypothesis. Science 1979, 204, 1344–1345. [Google Scholar] [CrossRef] [Green Version]
- Chmura, D.; Sierka, E. The invasibility of deciduous forest communities after disturbance: A case study of Carex brizoides and Impatiens parviflora invasion. For. Ecol. Manag. 2007, 242, 487–495. [Google Scholar] [CrossRef]
- Chabrerie, O.; Verheyen, K.; Saguez, R.; Decocq, G. Disentangling relationships between habitat conditions, disturbance history, plant diversity, and American black cherry (Prunus serotina Ehrh.) invasion in a European temperate forest. Divers. Distrib. 2008, 14, 204–212. [Google Scholar] [CrossRef]
- Dyderski, M.K.; Jagodziński, A.M. Low impact of disturbance on ecological success of invasive tree and shrub species in temperate forests. Plant Ecol. 2018, 219, 1369–1380. [Google Scholar] [CrossRef]
- Gunderson, L.H. Ecological Resilience—In Theory and Application. Ann. Rev. Ecol. Syst. 2000, 31, 425–439. [Google Scholar] [CrossRef]
- Jagodziński, A.M.; Dyderski, M.K.; Horodecki, P.; Rawlik, K. Limited dispersal prevents Quercus rubra invasion in a 14-species common garden experiment. Divers. Distrib. 2018, 24, 403–414. [Google Scholar] [CrossRef]
- Medvecká, J.; Kliment, J.; Majekova, J.; Halada, L.; Zaliberova, M.; Gojdičová, E.; Ferakova, V.; Jarolimek, I. Inventory of the alien flora of Slovakia. Preslia 2012, 84, 257–309. [Google Scholar]
- Pyšek, P.; Danihelka, J.; Sádlo, J.; Chrtek, J.; Chytrỳ, M.; Jarošík, V.; Kaplan, Z.; Krahulec, F.; Moravcová, L.; Pergl, J.; et al. Catalogue of alien plants of the Czech Republic: Checklist update, taxonomic diversity and invasion patterns. Preslia 2012, 84, 155–255. [Google Scholar]
- Wulf, M. Preference of plant species for woodlands with differing habitat continuities. Flora 2003, 198, 444–460. [Google Scholar] [CrossRef]
- Niinemets, Ü. A review of light interception in plant stands from leaf to canopy in different plant functional types and in species with varying shade tolerance. Ecol. Res. 2010, 25, 693–714. [Google Scholar] [CrossRef]
- Reich, P.B.; Oleksyn, J.; Modrzynski, J.; Mrozinski, P.; Hobbie, S.E.; Eissenstat, D.M.; Chorover, J.; Chadwick, O.A.; Hale, C.M.; Tjoelker, M.G. Linking litter calcium, earthworms and soil properties: A common garden test with 14 tree species. Ecol. Lett. 2005, 8, 811–818. [Google Scholar] [CrossRef]
- Horodecki, P.; Jagodziński, A.M. Tree species effects on litter decomposition in pure stands on afforested post-mining sites. For. Ecol. Manag. 2017, 406, 1–11. [Google Scholar] [CrossRef]
- Dickie, I.A.; Oleksyn, J.; Reich, P.B.; Karolewski, P.; Zytkowiak, R.; Jagodziński, A.M.; Turzanska, E. Soil modification by different tree species influences the extent of seedling ectomycorrhizal infection. Mycorrhiza 2006, 16, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Mueller, K.E.; Eisenhauer, N.; Reich, P.B.; Hobbie, S.E.; Chadwick, O.A.; Chorover, J.; Dobies, T.; Hale, C.M.; Jagodziński, A.M.; Kałucka, I.; et al. Light, earthworms, and soil resources as predictors of diversity of 10 soil invertebrate groups across monocultures of 14 tree species. Soil Biol. Biochem. 2016, 92, 184–198. [Google Scholar] [CrossRef] [Green Version]
- Barbier, S.; Gosselin, F.; Balandier, P. Influence of tree species on understory vegetation diversity and mechanisms involved—A critical review for temperate and boreal forests. For. Ecol. Manag. 2008, 254, 1–15. [Google Scholar] [CrossRef]
- Augusto, L.; Dupouey, J.-L.; Ranger, J. Effects of tree species on understory vegetation and environmental conditions in temperate forests. Ann. For. Sci. 2003, 60, 823–831. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Localities of study sites within the study area (a), localities of study plots within study sites: Strzeszyn and Bogdanka ecological lands (b), forests around Rusałka lake (c), and ecological lands “Dębina I” and “Dębina II” (d). Colors of dots represent vegetation types assigned to each study plot.
Figure 1.
Localities of study sites within the study area (a), localities of study plots within study sites: Strzeszyn and Bogdanka ecological lands (b), forests around Rusałka lake (c), and ecological lands “Dębina I” and “Dębina II” (d). Colors of dots represent vegetation types assigned to each study plot.

Figure 2.
Alien species richness explained by a random forest model. (a) Importance of variables, expressed by drop-out loss in model RMSE when a particular variable is perturbed (baseline refers to permuted model responses, dashed line indicates value for the full model). (b) Partial dependence plots (ceteris paribus plots) showing changes of predictions when a particular predictor was changed while all remaining predictors were constant. (c) Partial group predictions for predicted alien species richness assuming constant levels of other predictors. Boxes represent interquartile range and median and whiskers represent minimum–maximum range in forest vegetation types: Car-Aln—Carici elongatae-Alnetum, Car bet—Carpinion betuli, Che-Rob—Chelidonio-Robinietum, Fra-Aln—Fraxino-Alnetum, Pinus—monocultures of Pinus sylvestris, Pop alb—Populetum albae, and Sal alb—Salicetum albae. For further details, see Tables S2 and S3. Vegetation types marked by the same letter did not differ statistically significantly in predicted alien species richness, according to the Tukey posteriori test at p = 0.05. According to data transformation, values were scaled into relative percentiles.
Figure 2.
Alien species richness explained by a random forest model. (a) Importance of variables, expressed by drop-out loss in model RMSE when a particular variable is perturbed (baseline refers to permuted model responses, dashed line indicates value for the full model). (b) Partial dependence plots (ceteris paribus plots) showing changes of predictions when a particular predictor was changed while all remaining predictors were constant. (c) Partial group predictions for predicted alien species richness assuming constant levels of other predictors. Boxes represent interquartile range and median and whiskers represent minimum–maximum range in forest vegetation types: Car-Aln—Carici elongatae-Alnetum, Car bet—Carpinion betuli, Che-Rob—Chelidonio-Robinietum, Fra-Aln—Fraxino-Alnetum, Pinus—monocultures of Pinus sylvestris, Pop alb—Populetum albae, and Sal alb—Salicetum albae. For further details, see Tables S2 and S3. Vegetation types marked by the same letter did not differ statistically significantly in predicted alien species richness, according to the Tukey posteriori test at p = 0.05. According to data transformation, values were scaled into relative percentiles.

Figure 3.
Alien species cover explained by a random forest model. (a) Importance of variables, expressed by drop-out loss in the model RMSE when a particular variable was perturbed (baseline refers to permuted model responses, dashed line indicates value for the full model). (b) Partial dependence plots (ceteris paribus plots) showing changes of predictions when a particular predictor was changed while all remaining predictors were constant. (c) Partial group predictions for predicted alien species cover assuming constant levels of other predictors. Boxes represent interquartile range and median and whiskers represent minimum–maximum range in forest vegetation types: Car-Aln—Carici elongatae-Alnetum, Car bet—Carpinion betuli, Che-Rob—Chelidonio-Robinietum, Fra-Aln—Fraxino-Alnetum, Pinus—monocultures of Pinus sylvestris, Pop alb—Populetum albae, and Sal alb—Salicetum albae. For further details, see Tables S2 and S3. Vegetation types marked by the same letter did not differ statistically significantly in predicted alien species cover, according to the Tukey posteriori test at p = 0.05. According to data transformation, values were scaled into relative percentiles.
Figure 3.
Alien species cover explained by a random forest model. (a) Importance of variables, expressed by drop-out loss in the model RMSE when a particular variable was perturbed (baseline refers to permuted model responses, dashed line indicates value for the full model). (b) Partial dependence plots (ceteris paribus plots) showing changes of predictions when a particular predictor was changed while all remaining predictors were constant. (c) Partial group predictions for predicted alien species cover assuming constant levels of other predictors. Boxes represent interquartile range and median and whiskers represent minimum–maximum range in forest vegetation types: Car-Aln—Carici elongatae-Alnetum, Car bet—Carpinion betuli, Che-Rob—Chelidonio-Robinietum, Fra-Aln—Fraxino-Alnetum, Pinus—monocultures of Pinus sylvestris, Pop alb—Populetum albae, and Sal alb—Salicetum albae. For further details, see Tables S2 and S3. Vegetation types marked by the same letter did not differ statistically significantly in predicted alien species cover, according to the Tukey posteriori test at p = 0.05. According to data transformation, values were scaled into relative percentiles.


{kind=link}
{kind=link}
{kind=link}
| Parameter Tested (Table 2) | Hypothesis | Explanation | Reference |
|---|---|---|---|
| Rich.nat | Biotic resistance | High species richness of native species prevents invasion of alien species, i.e., decreases level of invasion | [24,35,44] |
| Rich.nat | Biotic acceptance | High native species richness indicates habitats suitable to be colonized by both native and alien species, i.e., prone to invasion | [10,47] |
| FRic, FDis, EIV-N, EIV-L, EIV-M | Habitat filtering | Invaders may be successful only if their traits are adapted to existing environmental filters, i.e., high level of environmental filtering decreases community level of invasion | [48,49] |
| FRic, FDis | Empty niche | Due to limited pool of native species, plant communities are unsaturated and provide opportunities for functionally distinct alien species, i.e., increases level of invasion | [50] |
| FDis | Environmental heterogeneity | Higher variability of niches in plant communities provide opportunities for colonization by more alien species, i.e., increases level of invasion | [49,50] |
| EIV-N, EIV-L, EIV-M | Increased resources availability | Increase of resource availability increases level of invasion | [25,51,52] |
| Dist.freq, Dist.sev | Disturbance | Disturbance events increase resource availability and reduce competition for alien species, i.e., increase level of invasion | [51,53] |
Table 2.
Number and sizes of study plots representing each vegetation type.
| Vegetation Type | No. of Plots | Min Plot Area (m2) | Mean Plot Area (m2) | SE | Max Plot Area (m2) |
|---|---|---|---|---|---|
| Carici elongatae-Alnetum | 31 | 100 | 109.7 | 5.4 | 200 |
| Carpinion betuli | 10 | 100 | 140.0 | 16.3 | 200 |
| Chelidonio-Robinietum | 8 | 100 | 106.3 | 6.3 | 150 |
| Fraxino-Alentum | 35 | 100 | 115.7 | 5.7 | 200 |
| Pinus monocultures | 24 | 100 | 100.0 | 0.0 | 100 |
| Populetum albae | 6 | 80 | 95.0 | 3.4 | 100 |
| Salicetum albae | 6 | 80 | 110.0 | 10.3 | 150 |
Table 3.
Overview of parameters analyzed in the study. Parameters marked by an asterisk are dependent variables and others are predictors.
Table 3.
Overview of parameters analyzed in the study. Parameters marked by an asterisk are dependent variables and others are predictors.
| Parameter | Abbreviation | Details | Min | Max | Mean | SE |
|---|---|---|---|---|---|---|
| Plot area | Plot.area | Measurement of plot area (m2) | 80.0 | 200.0 | 111.1 | 2.8 |
| Alien species richness * | Rich.alien | Number of alien species | 0.0 | 5.0 | 2.1 | 0.1 |
| Cover of alien species * | Cov.alien | Sum of cover scores for each alien species | 0.0 | 77.0 | 11.5 | 1.0 |
| Disturbance frequency index | Dist.freq | Species cover weighted mean of Herben et al. [76] indicators | −0.899 | 0.137 | −0.591 | 0.024 |
| Disturbance severity index | Dist.sev | Species cover weighted mean of Herben et al. [67] indicators | 0.083 | 0.500 | 0.258 | 0.008 |
| Ecological indicator value—light | EIV-L | Species cover weighted mean of Ellenberg and Leuschner [71] indicators | 4.000 | 6.898 | 5.422 | 0.066 |
| Ecological indicator value—moisture | EIV-M | Species cover weighted mean of Ellenberg and Leuschner [71] indicators | 3.950 | 9.545 | 6.122 | 0.111 |
| Ecological indicator value—soil fertility | EIV-N | Species cover weighted mean of Ellenberg and Leuschner [71] indicators | 1.495 | 8.638 | 6.217 | 0.100 |
| Functional dispersion | FDis | Averaged distance to the centroid in functional traits hypervolume | 0.05955 | 0.24113 | 0.16500 | 0.00370 |
| Functional richness | FRic | The volume of plant community species convex hull in functional traits hypervolume | 0.00001 | 0.20894 | 0.02336 | 0.00278 |
| Native species richness | Rich.nat | Number of native species | 3.0 | 28.0 | 11.6 | 0.5 |
| Tree stand type | StType | Plant association name | (seven classes) | |||
*—dependent variables.
Table 4.
Mean alien species richness and cover in the vegetation types of urban forests assessed using Poisson GLM for richness and zero-inflated Beta GLM for cover (see Tables S5 and S6 for full models, respectively) and Tukey posteriori test. Vegetation types marked by the same letter did not differ statistically significantly at p = 0.05.
Table 4.
Mean alien species richness and cover in the vegetation types of urban forests assessed using Poisson GLM for richness and zero-inflated Beta GLM for cover (see Tables S5 and S6 for full models, respectively) and Tukey posteriori test. Vegetation types marked by the same letter did not differ statistically significantly at p = 0.05.
| Vegetation Type | Mean Alien Species Richness | SE | Tukey Posteriori Test | Mean Alien Species Cover (%) | SE | Tukey Posteriori Test |
|---|---|---|---|---|---|---|
| Carici elongatae-Alnetum | 1.3 | 0.2 | a | 7.8 | 1.6 | a |
| Carpinion betuli | 2.5 | 0.3 | ab | 9.2 | 2.1 | c |
| Chelidonio-Robinietum | 3.6 | 0.4 | b | 14.8 | 2.6 | cd |
| Fraxino-Alentum | 2.2 | 0.2 | ab | 13.0 | 1.9 | e |
| Pinus monocultures | 2.3 | 0.2 | ab | 8.6 | 1.1 | b |
| Populetum albae | 2.5 | 0.5 | ab | 10.8 | 3.6 | abcd |
| Salicetum albae | 2.8 | 0.4 | ab | 34.5 | 9.8 | d |
Table 5.
Frequency of alien species in the herbaceous layer (% of all plots within each vegetation type) and median cover by vegetation types.
Table 5.
Frequency of alien species in the herbaceous layer (% of all plots within each vegetation type) and median cover by vegetation types.
| Species 1 | Life Form 2 | Invasion Status in Poland 3 [68] | Carici elongatae-Alnetum | Meadian Cover (%) | Carpinion betuli | Meadian Cover (%) | Chelidonio-Robinietum | Meadian Cover (%) | Fraxino-Alentum | Meadian Cover (%) | Pinus Monocultures | Meadian Cover (%) | Populetum albae | Meadian Cover (%) | Salicetum albae | Meadian Cover (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Acer negundo L. | P | inv | 0.0 | 0.0 | 10.0 | 2.0 | 25.0 | 1.5 | 5.7 | 2.0 | 4.2 | 2.0 | 16.7 | 2.0 | 66.7 | 3.0 |
| Amaranthus retroflexus L. | T | inv | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 16.7 | 2.0 |
| Bidens frondosa L. | T | inv | 9.7 | 1.0 | 0.0 | 0.0 | 0.0 | 0.0 | 11.4 | 2.0 | 0.0 | 0.0 | 16.7 | 2.0 | 66.7 | 10.5 |
| Chenopodium strictum Roth | T | nat | 0.0 | 0.0 | 0.0 | 0.0 | 12.5 | 1.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Cornus alba L. | P | cas | 3.2 | 3.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Cornus sericea L. | P | inv | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 5.7 | 3.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Echinocystis lobata (Michx.) Torr. & A. Gray | T, L | inv | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 16.7 | 3.0 | 0.0 | 0.0 |
| Fraxinus pensylvanica Marshall | P | inv | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 16.7 | 1.0 | 0.0 | 0.0 |
| Impatiens parviflora DC. | T | inv | 74.2 | 4.0 | 90.0 | 4.0 | 75.0 | 6.0 | 82.9 | 4.0 | 83.3 | 3.0 | 83.3 | 3.0 | 83.3 | 18.0 |
| Ligustrum vulgare L. * | P | nat | 6.5 | 1.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Lonicera tatarica L. | P | nat | 0.0 | 0.0 | 0.0 | 0.0 | 12.5 | 2.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Parthenocissus inserta (A. Kern.) R. M. Fritsch | P, L | inv | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 8.6 | 1.0 | 4.2 | 2.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Prunus domestica L. | P | nat | 6.5 | 1.0 | 40.0 | 1.0 | 25.0 | 3.5 | 5.7 | 1.5 | 12.5 | 1.0 | 16.7 | 4.0 | 16.7 | 18.0 |
| Prunus mahaleb L. | P | nat | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.2 | 1.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Prunus serotina Ehrh. | P | inv | 12.9 | 2.0 | 60.0 | 2.5 | 62.5 | 3.0 | 22.9 | 1.0 | 79.2 | 3.0 | 16.7 | 1.0 | 0.0 | 0.0 |
| Quercus rubra L. | P | inv | 0.0 | 0.0 | 0.0 | 0.0 | 37.5 | 1.0 | 0.0 | 0.0 | 8.3 | 1.5 | 0.0 | 0.0 | 0.0 | 0.0 |
| Ribes alpinum L. * | P | nat | 0.0 | 0.0 | 20.0 | 3.5 | 50.0 | 8.0 | 2.9 | 8.0 | 16.7 | 8.0 | 16.7 | 8.0 | 0.0 | 0.0 |
| Ribes rubrum L. | P | nat | 12.9 | 2.5 | 10.0 | 2.0 | 0.0 | 0.0 | 42.9 | 4.0 | 0.0 | 0.0 | 33.3 | 6.0 | 16.7 | 18.0 |
| Ribes uva-crispa L. * | P | nat | 0.0 | 0.0 | 10.0 | 2.0 | 12.5 | 2.0 | 8.6 | 3.0 | 0.0 | 0.0 | 0.0 | 0.0 | 16.7 | 3.0 |
| Robinia pseudacacia L. | P | inv | 0.0 | 0.0 | 10.0 | 2.0 | 37.5 | 2.0 | 2.9 | 1.0 | 16.7 | 1.5 | 0.0 | 0.0 | 0.0 | 0.0 |
| Rumex thyrsiflorus Fingerh. | H | nat | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 16.7 | 2.0 | 0.0 | 0.0 |
| Salvia glutinosa L. * | H | cas | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 8.6 | 3.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Solidago canadensis L. | G, H | inv | 3.2 | 1.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.9 | 2.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Solidago gigantea Aiton | G, H | inv | 0.0 | 0.0 | 0.0 | 0.0 | 12.5 | 3.0 | 5.7 | 2.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
1—species marked by an asterisk are native in southern Poland and alien in central and northern Poland, thus not included in Tokarska-Guzik et al. [68]; 2—life form after BiolFlor [72]: G—geophyte, L—liana, H—hemicryptophyte, P—phanerophyte (woody plant), T—therophyte (annual plant); 3—invasion status in Poland: cas—casual, nat—naturalized, inv—invasive.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dyderski, M.K.; Jagodziński, A.M. Context-Dependence of Urban Forest Vegetation Invasion Level and Alien Species’ Ecological Success. Forests 2019, 10, 26. https://doi.org/10.3390/f10010026
AMA Style
Dyderski MK, Jagodziński AM. Context-Dependence of Urban Forest Vegetation Invasion Level and Alien Species’ Ecological Success. Forests. 2019; 10(1):26. https://doi.org/10.3390/f10010026
Chicago/Turabian StyleDyderski, Marcin K., and Andrzej M. Jagodziński. 2019. "Context-Dependence of Urban Forest Vegetation Invasion Level and Alien Species’ Ecological Success" Forests 10, no. 1: 26. https://doi.org/10.3390/f10010026
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.