Stocks and Stoichiometry of Soil Organic Carbon, Total Nitrogen, and Total Phosphorus after Vegetation Restoration in the Loess Hilly Region, China



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Soil Sampling and Laboratory Analyses
2.3. Calculation of SOC, Soil TN, and Soil TP Stocks
2.4. Statistical Analyses
3. Results
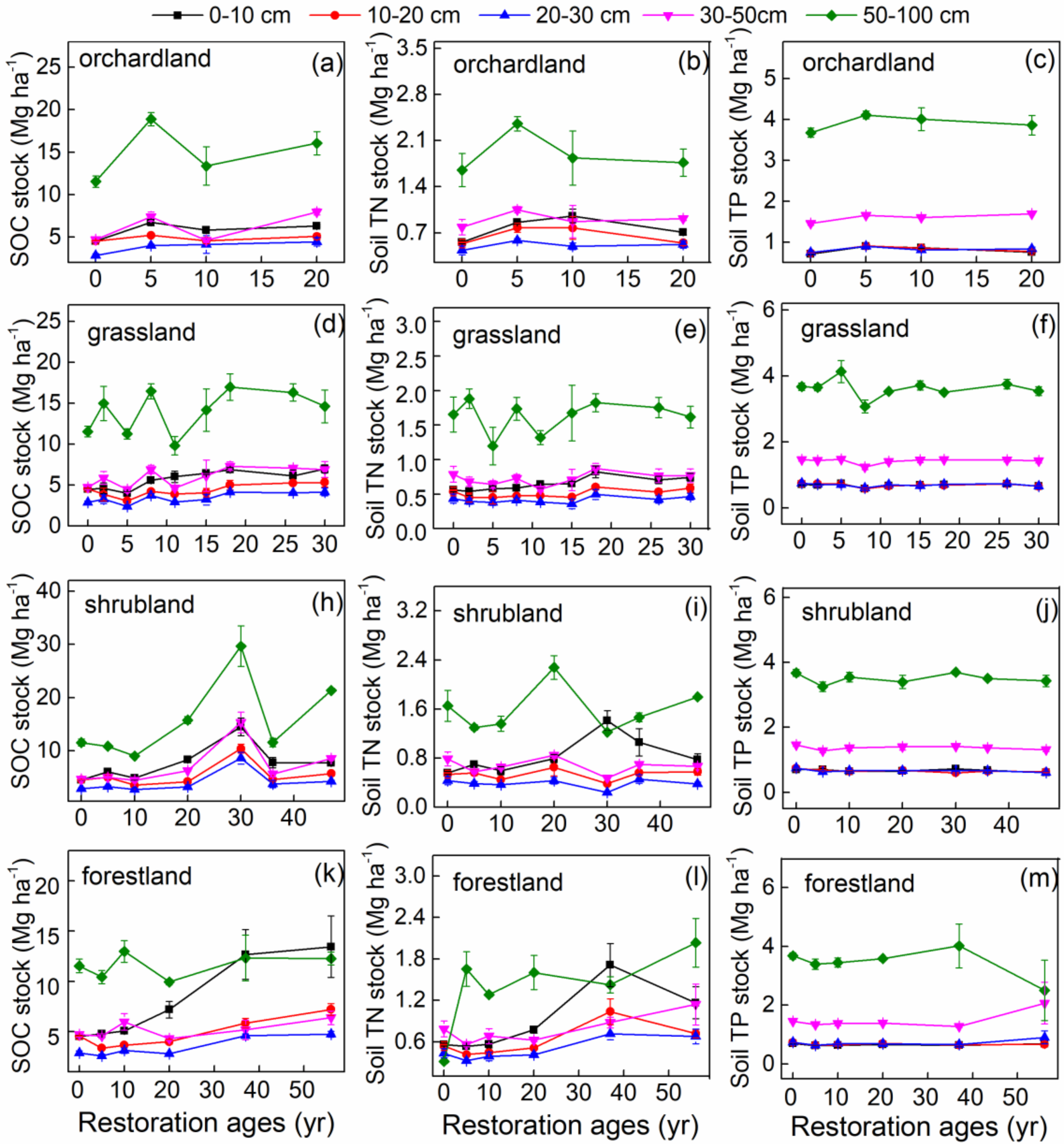
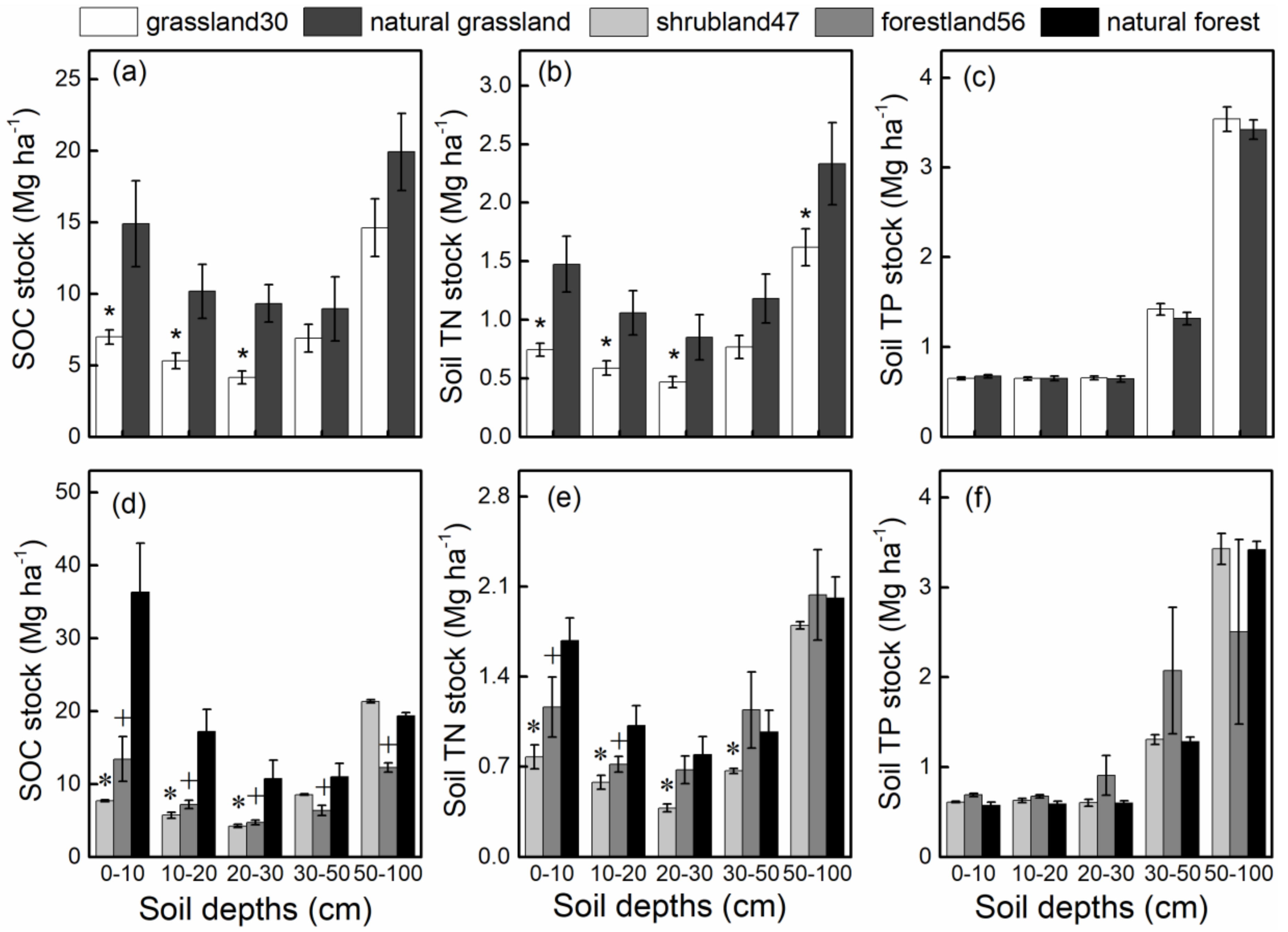
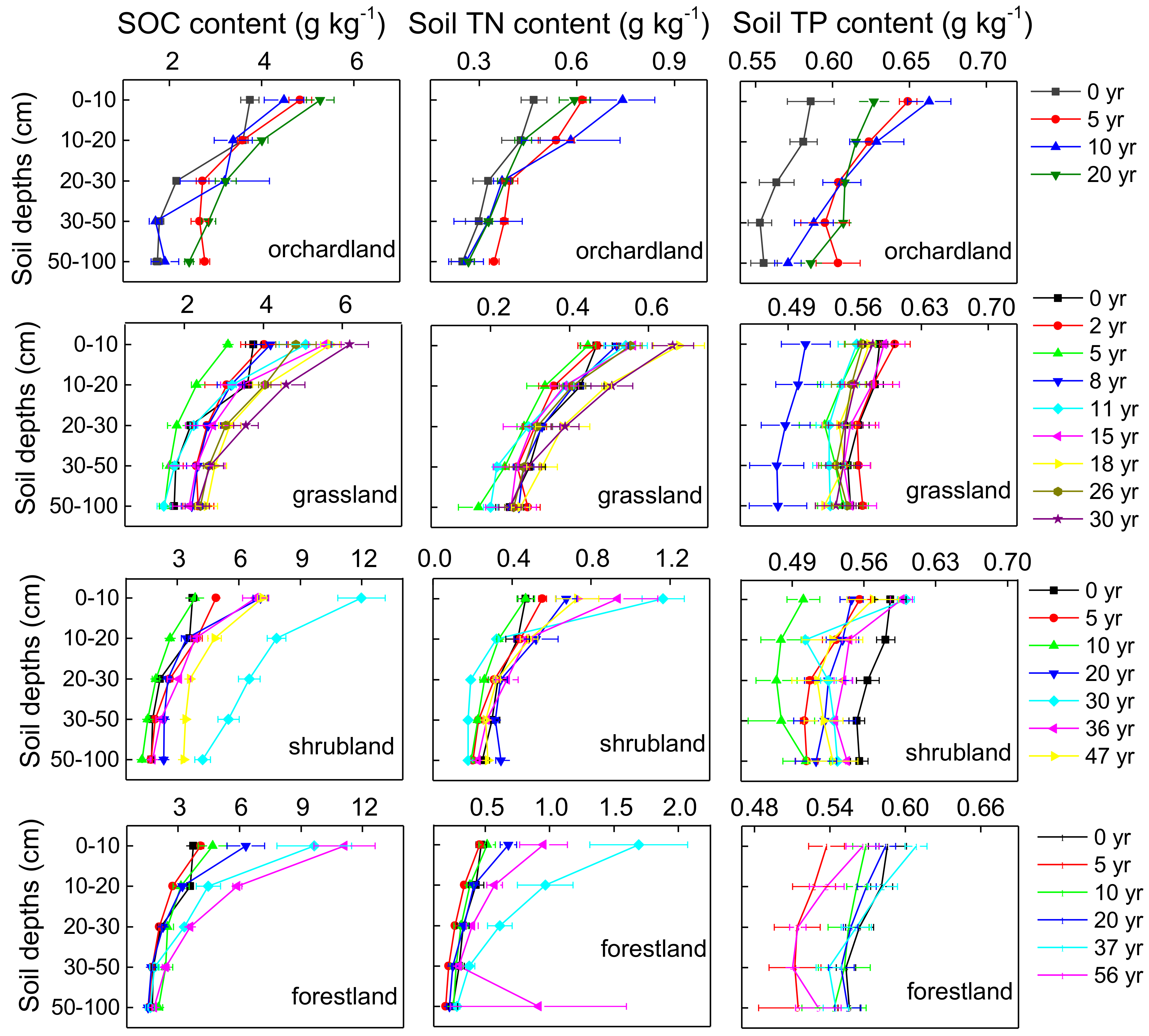
3.1. SOC, Soil TN, and Soil TP Stocks in Different Restoration Types
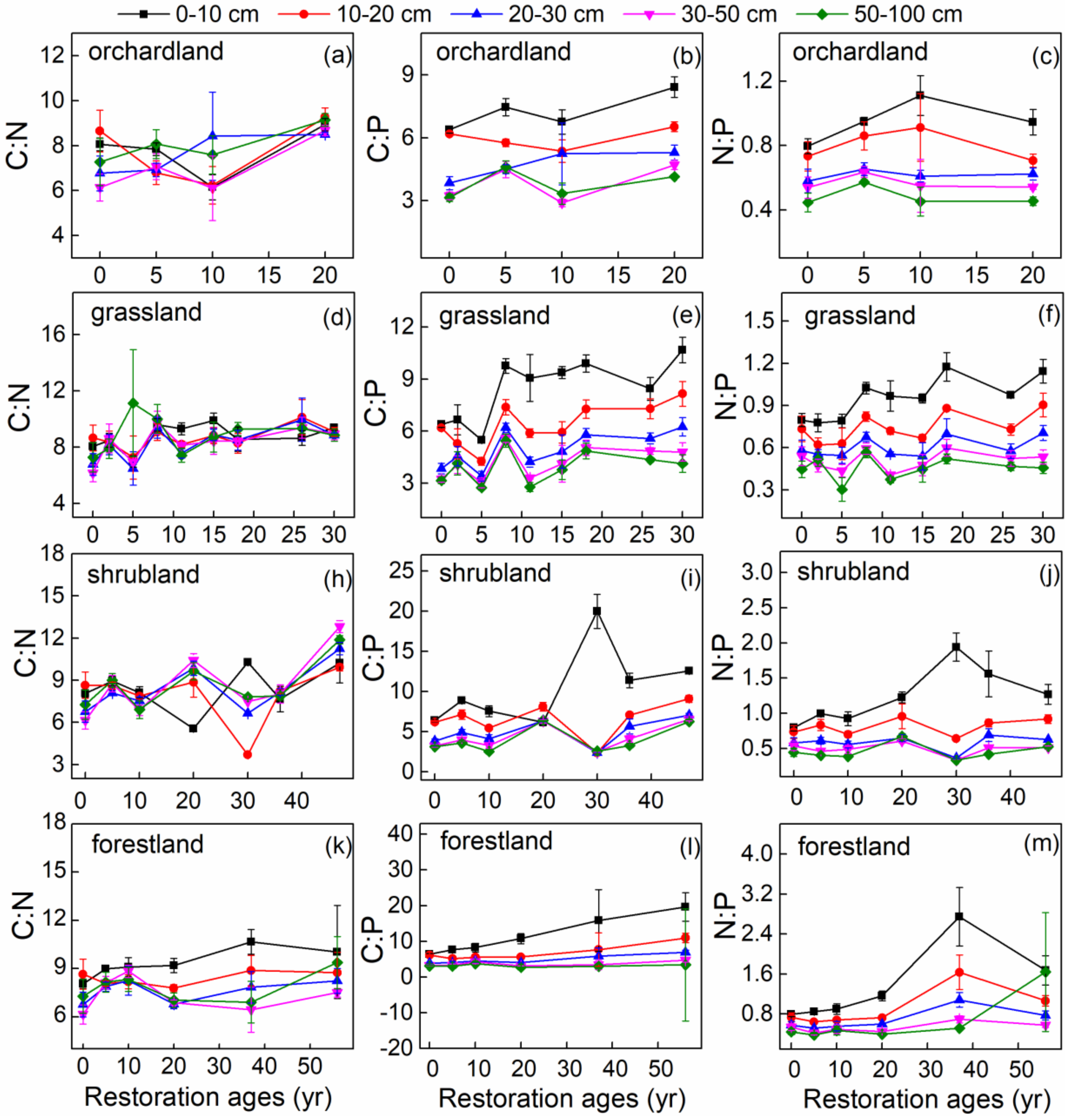
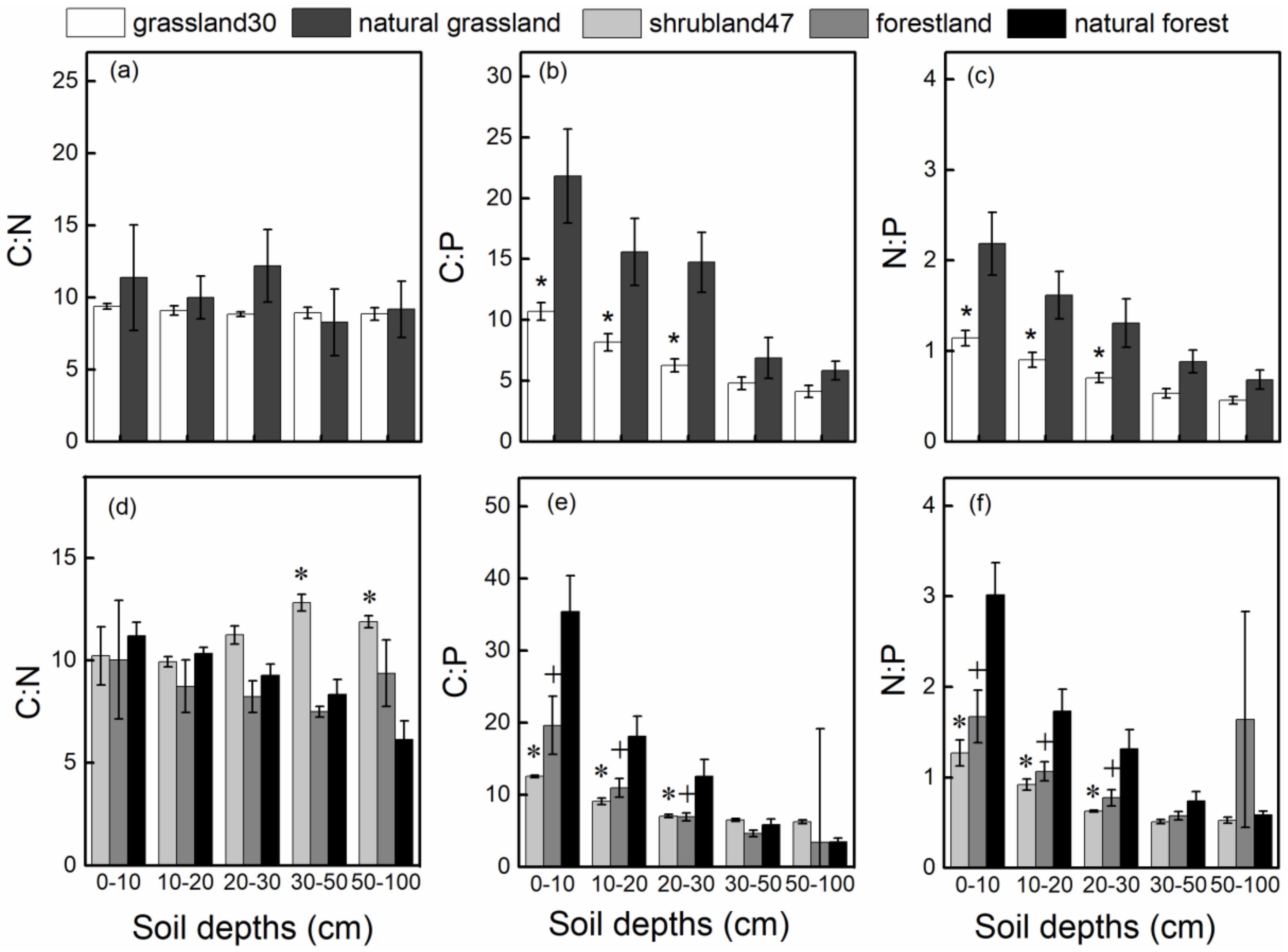
3.2. Changes in SOC, Soil TN, and Soil TP Stoichiometry
4. Discussion
4.1. Restoration Ages Altered SOC, Soil TN, and Soil TP Stocks and Stoichiometry
4.2. Vertical Distribution of Stocks and Stoichiometry of SOC, Soil TN, and Soil TP
4.3. Effect of Restoration Type on SOC, Soil TN, and Soil TP Stocks and Stoichiometry.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Reed, S.C.; Yang, X.; Thornton, P.E. Incorporating phosphorus cycling into global modeling efforts: A worthwhile, tractable endeavor. New Phytol. 2015, 208, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.L.; Yan, M.J.; Song, B.L. Ecological stoichiometry characteristics of soil carbon, nitrogen, and phosphorus in an oak forest and a black locust plantation in the Loess hilly region. Acta Ecol. Sin. 2017, 10, 3385–3393, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Rodríguez, E.S.; Lozano, Y.M.; Bardgett, R.D. Influence of soil microbiota in nurse plant systems. Funct. Ecol. 2016, 30, 30–40. [Google Scholar] [CrossRef]
- Rudel, T.K.; Coomes, O.T.; Moran, E.; Achard, F.; Angelsen, A.; Xu, J.; Lambin, E. Forest transitions: Towards a global understanding of land use change. Glob. Environ. 2005, 15, 23–31. [Google Scholar] [CrossRef]
- Winans, K.; Whalen, J.; Cogliastro, A.; Rivest, D.; Ribaudo, L. Soil Carbon Stocks in Two Hybrid Poplar-Hay Crop Systems in Southern Quebec, Canada. Forests 2014, 5, 1952–1966. [Google Scholar] [CrossRef]
- Jiang, P.; Chen, Y.; Cao, Y. C:N:P Stoichiometry and Carbon Storage in a Naturally-Regenerated Secondary Quercus variabilis Forest Age Sequence in the Qinling Mountains, China. Forests 2017, 8, 281. [Google Scholar] [CrossRef]
- Wang, W.; Sardans, J.; Zeng, C.; Zhong, C.; Li, Y.; Peñuelas, J. Responses of soil nutrient concentrations and stoichiometry to different human land uses in a subtropical tidal wetland. Geoderma 2014, 232–234, 459–470. [Google Scholar] [CrossRef]
- Gao, Y.; He, N.; Yu, G.; Chen, W.; Wang, Q. Long-term effects of different land use types on C, N, and P stoichiometry and storage in subtropical ecosystems: A case study in China. Ecol. Eng. 2014, 67, 171–181. [Google Scholar] [CrossRef]
- Zhao, F.; Sun, J.; Ren, C.; Kang, D.; Deng, J.; Han, X.; Yang, G.; Feng, Y.; Ren, G. Land use change influences soil C, N and P stoichiometry under ‘Grain-to-Green Program’ in China. Sci. Rep. 2015, 5, 10195. [Google Scholar] [CrossRef]
- Fang, J.; Chen, A.; Peng, C.; Zhao, S.; Ci, L. Changes in Forest Biomass Carbon Storage in China Between 1949 and 1998. Science 2001, 292, 2320–2322. [Google Scholar] [CrossRef]
- Post, W.M.; Kwon, K.C. Soil carbon sequestratrion and land-use change: Processes and potential. Glob. Environ. 2010, 6, 317–327. [Google Scholar] [CrossRef]
- Xiao, H.; Li, Z.; Dong, Y.; Chang, X.; Deng, L.; Huang, J.; Nie, X.; Liu, C.; Liu, L.; Wang, D.; et al. Changes in microbial communities and respiration following the revegetation of eroded soil. Agric. Ecosyst. Environ. 2017, 246, 30–37. [Google Scholar] [CrossRef]
- Yang, W.; Cheng, H.; Hao, F.; Ouyang, W.; Liu, S.; Lin, C. The influence of land-use change on the forms of phosphorus in soil profiles from the Sanjiang Plain of China. Geoderma 2012, 189–190, 207–214. [Google Scholar] [CrossRef]
- Liu, X.; Ma, J.; Ma, Z.; Li, L. Soil nutrient contents and stoichiometry as affected by land-use in an agro-pastoral region of northwest China. Catena 2017, 150, 146–153. [Google Scholar] [CrossRef]
- Li, C.; Zhao, L.; Sun, P.; Zhao, F.Z.; Kang, D.; Yang, G.H. Deep Soil C, N, and P Stocks and Stoichiometry in Response to Land Use Patterns in the Loess Hilly Region of China. PLoS ONE 2016, 11, e0159075. [Google Scholar] [CrossRef] [PubMed]
- Fei, W.; Rui, L.I.; Zhong, W. Problems and Proposals on Policy of Converting Cropland Into Forest and Grassland: A Case-based Study. Nat. Sci. Ed. 2003, 3, 60–65, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Xin, Z.; Ran, L.; Lu, X.X. Soil Erosion Control and Sediment Load Reduction in the Loess Plateau: Policy Perspectives. Int. J. Water Resour. Dev. 2012, 28, 325–341. [Google Scholar] [CrossRef]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon and organic matter. In Methods of Soil Analysis Part 3—Chemical Methods. Chem. Microbiol. Prop. 1982. [Google Scholar] [CrossRef]
- Bremner, J.; Mulvaney, C. Nitrogen—total. Methods of Soil Analysis Chemical Methods Part. 1996, 72, 532–535. [Google Scholar] [CrossRef]
- Schimel, J.P.; Weintraub, M.N. The implications of exoenzyme activity on microbial carbon and nitrogen limitation in soil: A theoretical model. Soil Biol. Biochem. 2003, 35, 549–563. [Google Scholar] [CrossRef]
- Clemmensen, K.E.; Bahr, A.; Ovaskainen, O.; Dahlberg, A.; Ekblad, A.; Wallander, H.; Stenlid, J.; Finlay, R.D.; Wardle, D.A.; Lindahl, B.D. Roots and Associated Fungi Drive Long-Term Carbon Sequestration in Boreal Forest. Science 2013, 339, 1615–1618. [Google Scholar] [CrossRef] [PubMed]
- Fontaine, S.; Barot, S.; Barré, P.; Bdioui, N.; Mary, B.; Rumpel, C. Stability of organic carbon in deep soil layers controlled by fresh carbon supply. Nature 2007, 450, 277–280. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Luo, Y. Carbon:nitrogen stoichiometry in forest ecosystems during stand development. Glob. Ecol. Biogeogr. 2011, 20, 354–361. [Google Scholar] [CrossRef]
- Li, Z.; Liu, C.; Dong, Y.; Chang, X.; Nie, X.; Liu, L.; Xiao, H.; Lu, Y.; Zeng, G. Response of soil organic carbon and nitrogen stocks to soil erosion and land use types in the Loess hilly–gully region of China. Soil Tillage Res. 2017, 166, 1–9. [Google Scholar] [CrossRef]
- Leon, D.G.D.; Moora, M.; Öpik, M.; Neuenkamp, L.; Gerz, M.; Jairus, T.; Vasar, M.; Bueno, C.G.; Davison, J.; Zobel, M. Symbiont dynamics during ecosystem succession: Co-occurring plant and arbuscular mycorrhizal fungal communities. FEMS Microbiol. Ecol. 2016, 92, 1–9. [Google Scholar] [CrossRef]
- Hao, W.F.; Liang, Z.S.; Chen, C.G.; Tang, L. Study of species diversity evolvement process during vegetation restoration of abandoned farmland in the hilly loess plateau. Pratac. Sci. 2005, 22, 1–8, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Novara, A.; Mantia, T.L.; Rühl, J.; Badalucco, L.; Kuzyakov, Y.; Gristina, L.; Laudicina, V.A. Dynamics of soil organic carbon pools after agricultural abandonment. Geoderma 2014, 235–236, 191–198. [Google Scholar] [CrossRef]
- O’Brien, S.L.; Jastrow, J.D. Physical and chemical protection in hierarchical soil aggregates regulates soil carbon and nitrogen recovery in restored perennial grasslands. Soil Biol. Biochem. 2013, 61, 1–13. [Google Scholar] [CrossRef]
- Jia, H.E.; Yu-Fu, H.U.; Shu, X.Y.; Wang, Q.; Jia, A.D.; Yan, X.; University, S.A. Effect of Salix cupularis plantations on soil stoichiometry and stocks in the alpine-cold desert of northwestern. Sin. Acta Agrestia Sin. 2018. (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Lane, P.N.J.; Noske, P.J.; Sheridan, G.J. Phosphorus enrichment from point to catchment scale following fire in eucalypt forests. Catena 2011, 87, 157–162. [Google Scholar] [CrossRef]
- Cheng, Y.; Li, P.; Xu, G.; Gao, H. Spatial distribution of soil total phosphorus in Yingwugou watershed of the Dan River, China. Catena 2016, 136, 175–181. [Google Scholar] [CrossRef]
- Kooijman, A.M.; Jongejans, J.; Sevink, J. Parent material effects on Mediterranean woodland ecosystems in NE Spain. Catena 2005, 59, 55–68. [Google Scholar] [CrossRef]
- Comino, J.R.; Sinoga, J.D.R.; González, J.M.S.; Guerra-Merchán, A.; Seeger, M.; Ries, J.B. High variability of soil erosion and hydrological processes in Mediterranean hillslope vineyards (Montes de Málaga, Spain). Catena 2016, 145, 274–284. [Google Scholar] [CrossRef]
- Duan, X.N.; Wang, X.K.; Lu, F.; Ouyang, Z.Y. Soil carbon sequestration and its potential by grassland ecosystems in China. Acta Ecol. Sin. 2008. (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Reich, P.B.; Oleksyn, J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proc. Natl. Acad. Sci. USA 2004, 101, 11001–11006. [Google Scholar] [CrossRef] [PubMed]
- An, H.; Li, G.Q. Effects of grazing on carbon and nitrogen in plants and soils in a semiarid desert grassland, China. J. Arid Land 2015, 7, 341–349. [Google Scholar] [CrossRef]
- Yu, Y.F.; Peng, W.X.; Song, T.Q. Stoichiometric characteristics of plant and soil C, N and P in different forest types in depressions between karst hills, southwest China. Chin. J. Appl. Ecol. 2014, 25, 947–954, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Chen, L.; Li, C.L. Research advances in ecological stoichiometry of marine plankton. Chin. J. Appl. Ecol. 2014, 25, 3047–3055, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Berger, T.W.; Neubauer, C.; Glatzel, G. Factors controlling soil carbon and nitrogen stores in pure stands of Norway spruce (Picea abies) and mixed species stands in Austria. For. Ecol. Manag. 2002, 159, 3–14. [Google Scholar] [CrossRef]
- Liu, X.Z.; Zhou, G.Y.; Zhang, D.Q.; Liu, S.Z.; Chu, G.W.; Yan, J.H. N and P stoichiometry of plant and soil in lower subtropical forest successional series in southern China. J. Plant Ecol. 2010, 34, 64–71, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Zeng, Q.; Liu, Y.; Fang, Y.; Ma, R.; Lal, R.; An, S.; Huang, Y. Impact of vegetation restoration on plants and soil C:N:P stoichiometry on the Yunwu Mountain Reserve of China. Ecol. Eng. 2017, 109, 92–100. [Google Scholar] [CrossRef]
- Barreto, P.A.B.; Gama-Rodrigues, E.F.; Fontes, A.G.; Polidoro, J.C.; Moço, M.K.S.; Machado, R.C.R.; Baligar, V.C. Distribution of oxidizable organic C fractions in soils under cacao agroforestry systems in Southern Bahia, Brazil. Agrofor. Syst. 2011, 81, 213–220. [Google Scholar] [CrossRef]
- Zhang, X. Ecological stoichiometry characteristics of robinia pseudoacacia forest soil in different latitudes of loess plateau. Acta Ecol. Sin. 2013, 50, 818–825. [Google Scholar] [CrossRef]
- Han, L.; Li, Z.; Ceng, Y. Carbon, nitrogen, and phosphorous stoichiometry of herbaceous plant leaf and soil in riparian zone of Taihu Lake basin, East China under effects of different land use types. Chin. J. Ecol. 2013, 32, 3281–3288, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Vesterdal, L.; Ritter, E.; Gundersen, P. Change in soil organic carbon following afforestation of former arable land. For. Ecol. Manag. 2002, 169, 137–147. [Google Scholar] [CrossRef]
- Guo, S.L.; Ma, Y.H.; Che, S.G.; Sun, W.Y. Effects of artificial and natural vegetations on litter production and soil organic carbon change in loess hilly areas. Sci. Silvae Sin. 2009, 45, 14–18, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Pizzeghello, D.; Francioso, O.; Concheri, G.; Muscolo, A.; Nardi, S. Land Use Affects the Soil C Sequestration in Alpine Environment, NE Italy. Forests 2017, 8, 197. [Google Scholar] [CrossRef]
- De Feudis, M.; Cardelli, V.; Massaccesi, L.; Lagomarsino, A.; Fornasier, F.; Westphalen, D.J.; Coccco, S.; Corti, G.; Agnelli, A. Influence of Altitude on Biochemical Properties of European Beech (Fagus sylvatica L.) Forest Soils. Forests 2017, 8, 213. [Google Scholar] [CrossRef]
- Cesarz, S.; Fender, A.C.; Beyer, F.; Valtanen, K.; Pfeiffer, B.; Gansert, D.; Hertel, D.; Leuschner, C. Roots from beech (Fagus sylvatica L.) and ash (Fraxinus excelsior L.) differentially affect soil microorganisms and carbon dynamics. Soil Biol. Biochem. 2013, 61, 23–32. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, H.; Qu, Q.; Li, P.; Guo, Z.; Wulan, E.; Xue, S. Stocks and Stoichiometry of Soil Organic Carbon, Total Nitrogen, and Total Phosphorus after Vegetation Restoration in the Loess Hilly Region, China. Forests 2019, 10, 27. https://doi.org/10.3390/f10010027
Xu H, Qu Q, Li P, Guo Z, Wulan E, Xue S. Stocks and Stoichiometry of Soil Organic Carbon, Total Nitrogen, and Total Phosphorus after Vegetation Restoration in the Loess Hilly Region, China. Forests. 2019; 10(1):27. https://doi.org/10.3390/f10010027
Chicago/Turabian StyleXu, Hongwei, Qing Qu, Peng Li, Ziqi Guo, Entemake Wulan, and Sha Xue. 2019. "Stocks and Stoichiometry of Soil Organic Carbon, Total Nitrogen, and Total Phosphorus after Vegetation Restoration in the Loess Hilly Region, China" Forests 10, no. 1: 27. https://doi.org/10.3390/f10010027
APA StyleXu, H., Qu, Q., Li, P., Guo, Z., Wulan, E., & Xue, S. (2019). Stocks and Stoichiometry of Soil Organic Carbon, Total Nitrogen, and Total Phosphorus after Vegetation Restoration in the Loess Hilly Region, China. Forests, 10(1), 27. https://doi.org/10.3390/f10010027