The Positive Effect of Different 24-epiBL Pretreatments on Salinity Tolerance in Robinia pseudoacacia L. Seedlings


1
Co-Innovation Center for Sustainable Forestry in Southern China of Jiangsu Province, Key Laboratory of Soil and Water Conservation and Ecological Restoration, Nanjing Forestry University, 159 Longpan Road, Nanjing 21037, Jiangsu, China
2
Department of Forest and Conservation Sciences, Faculty of Forestry, University of British Columbia, 2424 Main Mall, Vancouver, BC V6T 1Z4, Canada
3
State Key Laboratory of Tree Genetics and Breeding & Key Laboratory of Tree Breeding and Cultivation, State Forestry Administration, Research Institute of Forestry, Chinese Academy of Forestry, Bejing 100091, China
*
Author to whom correspondence should be addressed.
Forests 2019, 10(1), 4; https://doi.org/10.3390/f10010004
Submission received: 28 November 2018
/
Revised: 16 December 2018
/
Accepted: 18 December 2018
/
Published: 20 December 2018
(This article belongs to the Special Issue Physiological Responses to Abiotic and Biotic Stress in Forest Trees)
Abstract
:As a brassinosteroid (BR), 24-epibrassinolide (24-epiBL) has been widely used to enhance the resistance of plants to multiple stresses, including salinity. Black locust (Robinia pseudoacacia L.) is a common species in degraded soils. In the current study, plants were pretreated with three levels of 24-epiBL (0.21, 0.62, or 1.04 µM) by either soaking seeds during the germination phase (Sew), foliar spraying (Spw), or root dipping (Diw) at the age of 6 months. The plants were exposed to salt stress (100 and 200 mM NaCl) via automatic drip-feeding (water content ~40%) for 45 days after each treatment. Increased salinity resulted in a decrease in net photosynthesis rate (Pn), stomatal conductance (Gs), intercellular:ambient CO2 concentration ratio (Ci/Ca), water-use efficiency (WUEi), and maximum quantum yield of photosystem II (PSII) (Fv/Fm). Non-photochemical quenching (NPQ) and thermal dissipation (Hd) were elevated under stress, which accompanied the reduction in the membrane steady index (MSI), water content (RWC), and pigment concentration (Chl a, Chl b, and Chl). Indicators of oxidative stress (i.e., malondialdehyde (MDA) and antioxidant enzymes (peroxidase (POD) and superoxide dismutase (SOD)) in leaves and Na+ content in chloroplasts increased accompanied by a reduction in chloroplastid K+ and Ca2+. At 200 mM NaCl, the chloroplast and thylakoid ultrastructures were severely disrupted. Exogenous 24-epiBL improved MSI, RWC, K+, and Ca2+ content, reduced Na+ levels, maintained chloroplast and thylakoid membrane structures, and enhanced the antioxidant ability in leaves. 24-epiBL also substantially alleviated stress-induced limitations of photosynthetic ability, reflected by elevated chlorophyll fluorescence, pigment levels, and Pn. The positive effects of alleviating salt stress in R. pseudoacacia seedlings in terms of treatment application was Diw > Sew > Spw, and the most positive impacts were seen with 1.04 µM 24-epiBL. These results provide diverse choice for 24-epiBL usage to defend against NaCl stress of a plant.
1. Introduction
A current report states that more than 1 billion hectares of land have been damaged by salinity as a result of anthropogenic activities [1]. With current practices, salinization could affect half of the currently cultivated land by 2050 [2]. China has the third largest area of saline soils (366,500 km2) in the world [3], resulting in significant agricultural losses and limiting economic development and quality of life. As the most densely populated province in China, located at the lower reaches of Yangtze River and along the coast of the Yellow Sea, Jiangsu province is at risk of soil salinity resulting from evaporation from the soil surface, especially in the ~15 km2 of degraded land that has been reclaimed along the coastline annually [4].
Black locust (Robinia pseudoacacia L.), an arboreal member of the Fabaceae indigenous to the eastern USA, is the third most-planted tree in reforestation schemes because of its ability to adapt to a range of environmental conditions in China [5]. As a salt-tolerant species, R. pseudoacacia is able to withstand salinity levels up to ~50 mM Na+ in laboratory conditions, or ~100 mM Na+ in field conditions [6]. It is also widely used as a major tree species to improve degraded soils [7], including saline soils, and has been widely cultivated in China since the early 20th century [8,9]. In Jiangsu, the soils most affected by salinity are along the coast, and these areas are also negatively impacted by hurricanes. It has been reported that stands of R. pseudoacacia could eliminate or reduce to <5% the impact of hurricanes on such stands and could also significantly increase soil nitrate concentrations along the coast [10]. Thus, there is a need to improve the adaptation of R. pseudoacacia to salinity-affected soils so that they can be planted along the coast in Jiangsu province.
Salt stress always results in water deficit, ion toxicities and imbalances, and oxidative stress, damaging plant cells and organs, limiting plant growth, and even causing death. For example, 85.73 mM NaCl increased the water saturation deficit in weak roots and limited the biomass accumulation of individual R. pseudoacacia [11]. The growth of the medicinal plant, L. japonica, was reduced by 200 mM NaCl because of negative effects on the ionic uptake and their distribution to plant organs [12]. Oxidative factors are prevalent in both lower and higher salinity conditions. For instance, 50 mM NaCl caused a significant reduction in plant growth and photosynthetic parameters accompanied by lipid peroxidation and hydrogen peroxide in Capsicum annuum L. [13]. In addition, 250 mM NaCl and 500 mM NaCl limited the growth of R. pseudoacacia by reducing the antioxidant enzyme activity, combined with a reduction in the photosynthetic pigment contents, damaging the chloroplast ultrastructure [14].
As a brassinosteroid (BR), 24-epibrassinolide (24-epiBL) has been used in many plant species to regulate their tolerance to stress; for example, against chilling stress in young grapevine seedlings (Vitis vinifera L.) [15], against Ca (NO3)2 stress in cucumber, Cucumis sativus (L., cv. ‘Jinyou No. 4′) [16], and against low temperatures and poor light intensities in tomato (Lycopersicon esculentum Mill.) [17]. In addition, 24-epiBl has also been used to modify salinity stress, such as, 24-epiBL ameliorated saline stress, and improved the productivity of wheat (Triticum aestivum L.) sprayed with 0.105 µM and 0.21 µM 24-epiBL by modifying the photosynthesis, chlorophyll, and nitrate levels in wheat leaves [18]. 24-epiBL ameliorated salinity-induced injuries by reducing the K+ efflux in barley seedlings (Hordeum vulgare cv. ‘Franklin′) when the seeds were soaked in 0.21, 0.53, or 1.05 µM 24-epiBL solutions [19].
Therefore, we hypothesized that soaking seeds, spraying leaves, and dipping roots of R. pseudoacacia in 0.21–1.04 µM 24-epiBL could improve the ability of seedlings to adapt to saline soils. This adaptation might result from response mechanisms, including maintaining the regular structure and function of chloroplasts and modifying photosynthesis. Our approach could also reveal the most suitable method for 24-epiBL application. Thus, we applied NaCl stress and 24-epiBL treatments to greenhouse seedlings and then measured: (a) Photosynthesis parameters; (b) chlorophyll fluorescence; (c) water content and membrane stability of leaves; (d) photosynthetic pigments in leaves; (e) malondialdehyde (MDA), and antioxidant enzyme activities (peroxidase (POD) and superoxide dismutase (SOD)) in leaves; (f) ions in chloroplasts; and (g) chloroplast ultrastructure. The results obtained could help shed light on the most appropriate treatment to use to improve the salinity tolerance of R. pseudoacacia seedlings for use in reforestation programs in areas of high salinity, both along the coastline in Jiangsu and elsewhere.
2. Materials and Methods
2.1. Plant Material and 24-epibrassinolide Treatments
The experiments ran from September 2014 to July 2015. 24-epiBL (Sigma-Aldrich, St. Louis, MO, USA) was applied at the concentrations of 0.21, 0.62, and 1.04 µM for each treatment type: Seed soaking, foliar spraying, and root dipping. Seeds were sown into trays containing quartz sand moistened with either distilled water or the relevant concentration 24-epiBL. When the cotyledons were fully expanded, the seedlings were transplanted into seedling bags. Seedlings were grown with no stress for 6 months until Spring 2015, when they were ~45 cm tall. All seedlings were then washed to remove the rooting medium. Seedlings grown from seeds pretreated with 24-epiBL (0.21, 0.62, or 1.04 µM) or distilled water were replanted directly into plastic pots (40 cm × 25 cm × 15 cm; 1 plant per pot) containing coarse sand and vermiculite 2:1 (v/v). After 1 week, seedlings were partly picked up of the seedlings from seeds with distilled water for foliar spraying with 0.21, 0.62, or 1.04 µM 24-epiBL every 7 days, each lasting 28 days. Other seedlings from untreated seeds were split into two groups and root dipped for 48 h in either distilled water or 0.21, 0.62, or 1.04 µM 24-epiBL (refreshed every 12 h) before being replanted using the methodology described earlier. Seedlings were watered to the drip-point every 7 days with a modified Hoagland’s nutrient solution [20]. The details of materials used to cultivate the seedlings were as described by our previous paper [21].
2.2. Groups and Salt Stress Treatments
After a further 2 weeks of growth, seedlings of uniform height (~50 cm) were selected for the salt stress treatments. There were four groups: (i) Seedlings never exposed to 24-epiBL (CK); (ii) seedlings from seeds treated with 24-epiBL (Sew); (iii) seedlings foliar sprayed with 24-epiBL (Spw); and (iv) seedlings root-dipped in 24-epiBL (Diw). Fifteen seedlings of each group were subjected to each of two levels of salt stress (100 and 200 mM NaCl, both in Hoagland’s solution) and non-stress (Hoagland’s solution). Pots were first flushed with their respective nutrient solutions and the water content was then maintained near to saturation (40%) by automatic drip-feeding with fresh solution delivered by an Intelligent Automatic Watering System (patent CN 201398356 Y). There were two salt stress treatments (100 and 200 mM NaCl, designated SS-S1 and SS-S2, respectively) plus a zero-salt control (CK), resulting in 21 treatment groups overall (Table 1).
Data for gas exchange, chlorophyll fluorescence, antioxidant systems, pigment content, ion content, and chloroplast ultrastructure were collected as described below, and measurements began 45 days after the treatments had started. Over the measurement period, the day/night temperature regime was 29 °C/18 °C, relative humidity varied from 45%–80%, and natural lighting provided a photoperiod of ~14 h and a mid-day photosynthetic photon flux density (PPFD) of ~1000 µmol m−2 s−1.
2.3. Photosynthesis
The net photosynthetic rate (Pn), stomatal conductance (Gs), and intercellular CO2 concentration (Ci) were measured on 0.7 cm × 3.0 cm (2.1 cm2) of the sunlight-exposed leaves using an ambient CO2 concentration (Ca) of 380 µmol mol−1 by gas exchange analyzer (LI-6400, LI-COR Inc., Lincoln, NE, USA). The ratio of intercellular to ambient CO2 concentration was calculated as Ci/Ca and the intrinsic water-use efficiency (WUEi) was calculated as Pn/Gs. The measurement details were as described in our previous paper [21].
Chlorophyll fluorescence was measured using a portable fluorometer (149 PAM-2500; Walz, Effeltrich, Germany). We report the maximum quantum yield of photosystem II (PSII) as Fv/Fm, thermal dissipation (Hd) as 1 − (Fv′/Fm′), and non-photochemical quenching (NPQ) as (Fm − Fm′)/Fm′ in this research. The measurement details were as described in our previous paper [21].
2.4. Estimation of Membrane Stability Index and Relative Water Content
The membrane stability index (MSI) and relative water content (RWC) were estimated following the method from Yue et al. [21]. MSI and RWC was calculated using the formula given by Sairam [22] and Hayat et al. [23] as follows:
MSI = [1 − C1/C2 × 100] and RWC = (FW − DW)/(TW − DW) × 100%. Electrical conductivity bridge measured at 40 °C for 30 min (C1), electrical conductivity bridge measured at 100 °C for 100 min (C2); fresh weight of leaves (FW), dry weight of leaves (DW) and turgor weight of leaves (TW).
2.5. Observation of Chloroplast Ultrastructure
The chloroplasts were observed using a Hitachi transmission electron microscope (Carl Zeiss, Göttingen, Germany) and the details are described by Yue et al. [21].
2.6. Assays for Malondialdehyde and Antioxidant Enzyme Activities
200 mg of fresh leaf samples (n = 3) taken from the plant were pulverized with a mortar and pestle in liquid nitrogen before adding 0.05M phosphate buffer (pH = 7.0) for further grinding. Then, the suspension was collected in a test tube and diluted with the same buffer to 10 mL for testing enzyme activity [24]. Superoxide dismutase (SOD) was assayed with the nitroblue tetrazolium (NBT) method of Fridovich (1975) [25] and expressed in terms of units min−1 g−1 fresh weight (FW). One unit (U) of SOD was defined as the amount of enzyme required to cause 50% inhibition of the reduction of NBT as monitored at 560 nm. Peroxidase (POD) was measured according to Hammerschmidt et al. (1982) [26] by monitoring the rate of guaiacol oxidation at 470 nm. The standard curve was constructed using 4-methoxyphenol and activity was expressed as μg oxidized min−1 g−1 FW. Lipid peroxidation was determined by measuring the amount of malondialdehyde (MDA) produced per g FW by the thiobarbituric acid reaction [27].
2.7. Assays for Chlorophyll Pigment Concentration
Chlorophyll pigment was obtained by 0.2 g of laminal tissue from three leaves ground in ice-cold 1:1 (v/v) ethanol: acetone using a mortar and pestle. The extracts were centrifuged at 6000g for 10 min, washed with extractant, then centrifuged again. The absorbance of the combined supernatants was at 645 nm and 663 nm. Total chlorophyll (Chl), chlorophyll a (Chl a), and chlorophyll b (Chl b) were calculated according to our previous paper [21].
2.8. Isolation of Chloroplasts for Testing
Chloroplasts were isolated from fully expanded leaves by differential and density gradient centrifugation as described by Cerović and Plesnicar (1984) [28] and Song and others (2006) [29] with some modifications. The details were the same as Yue et al. [21]. For chloroplast Na+, Ca2+, and K+ content, pelleted chloroplasts from a final 1 mL aliquot were extracted with 5 mL of 0.5 M HCl by shaking on a water-bath at 50 °C for 45 min. The volume was brought up to 10 mL with 0.5 M HCl before filtering and diluting for inductively coupled plasma optical spectroscopy (ICP-OES) as elemental analysis on a ICP-OES spectrometer (SPECTRO CIRO CCD, GmbH & Co KG, Kleve, Germany) at wavelengths of 226-502 nm [30].
2.9. Statistical Analysis
All analyses had at least three biological replicates. The data of control, salt stress, the seeds soaking, and roots dipping with 1.04 µM 24-epiBL in Pn, Gs, Fv/Fm, NPQ, RWC, and MSI was shown in our previous paper [21], and other data were initial. Data were statistically analyzed with the Statistical Package for the Social Sciences (SPSS) 19.0 (SPSS Inc., Chicago, IL, USA). One-way analyses of variance (ANOVA) were employed to test the effects of treatment with 24-epiBL under salt stress. Pairwise comparisons were by a Duncan test and considered significant at p < 0.05. Three-way analyses of variance (ANOVA) were employed to test the effects of salt stress, 24-epiBL application methods, 24-epiBL usage concentration, and their interaction, and means were separated using Duncan’s multiple range tests, with the significance considered as p < 0.05.
3. Results
3.1. Photosynthesis
Compared with the CK group, salt stress in the absence of any pretreatments with 24-epiBL obviously reduced photosynthesis (Pn) and stomatal conductance (Gs), 4.4 and 1.67 times under 100 NaCl mM stress (SS-S1) and 6.3 and 5.2 times under 200 mM NaCl stress (SS-S2), respectively. The change in Pn was not significant, but the difference in Gs was obvious between the two levels of NaCl (Figure 1A,B). Consequently, Ci/Ca was decreased at both salt stress levels, as was WUEi, although there were no significant difference in Ci/Ca between SS-S1 and CK and in WUEi between SS-S2 and CK (Figure 1C,D). There were no significant differences in Pn, Ci/Ca, or WUEi in the SS-S1 versus SS-S2. This inhibition of photosynthesis was partially prevented by pretreatment with 24-epiBL and the effect of 24-epiBL in SS-S1 was better than in SS-S2; Pn in Diw-1.04S1 > Diw-0.62S1 > Sew-1.04S1 > Spw-1.04S1 > Diw-0.21S1 > Sew-0.62S1 were significantly higher than in SS-S1 (Figure 1A), and Pn in SS-S2 was obviously increased only by Diw-1.04S2 and Sew-1.04S2 (Figure 1A). Under 100 mM NaCl stress, pretreatment with 24-epiBL did not significantly affect Gs relative to SS-S1 combined with the decrease in Ci/Ca (except for Sew-0.21S1, Spw-0.21S1, and Spw-0.62S1) and increase in WUEi (Figure 1B–D). The significant improvement in Gs following pretreatment with 24-epiBL was in contrast to the lack of an effect on Ci/Ca or WUEi in the 200 mM NaCl groups, and the significant difference of Gs existed in Diw-1.04S2 versus Spw-0.21/0.62S2; the increase of Ci/Ca was only in the Diw group (Figure 1B–D). All these changes were modified by increasing the concentration of 24-epiBL; the order of the concentration effect was 1.04 > 0.62 > 0.21 µM. The effects of salt concentration (Sc), the 24-epiBL application method (Am), and 24-epiBL application concentration (BRc) were obvious (p < 0.01), but the interactions in the above three factors were not significant (p > 0.05) in Pn, Gs, and WUEi. The obvious role of Ci/Ca was only shown in BRc and the interaction between the salt concentration and 24-epiBL application method (Sc*Am) (p < 0.01).
3.2. Chlorophyll Fluorescence
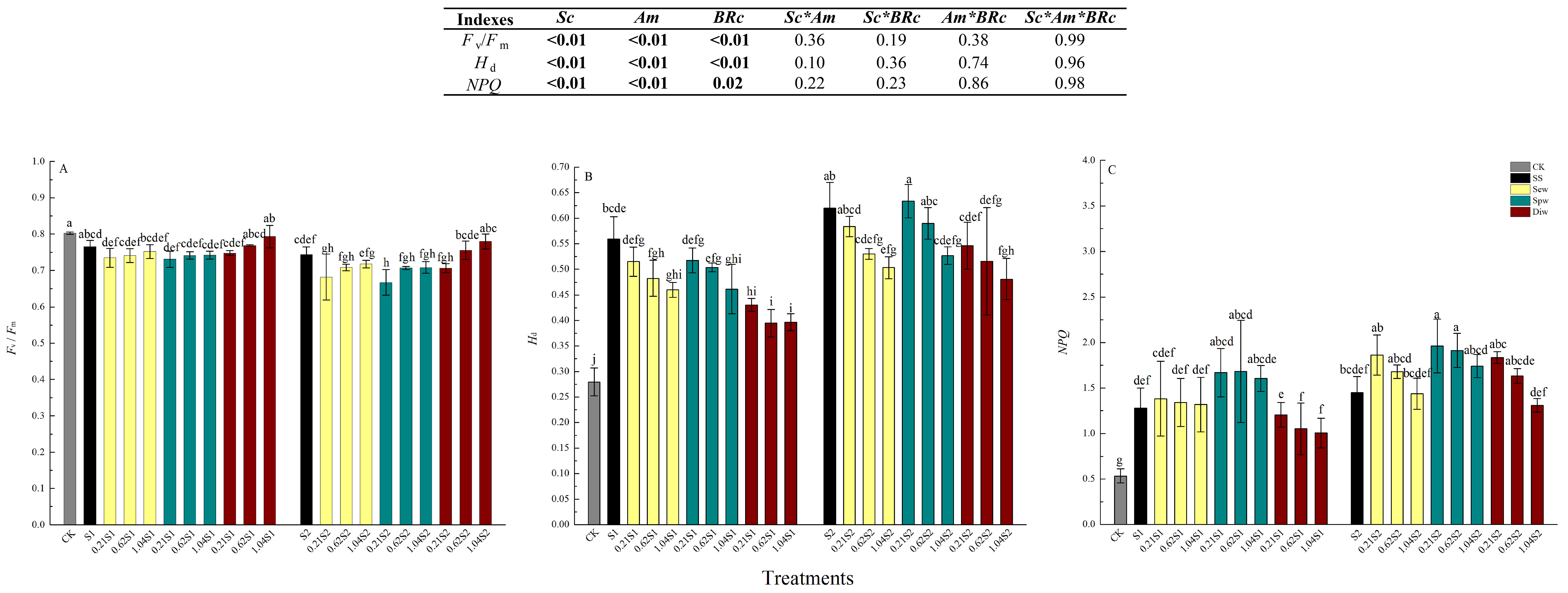
Maximum quantum yield of photosystem II (PSII) (Fv/Fm) decreased in response to saline treatments, whereas nonphotochemical quenching (NPQ) and thermal dissipation (Hd) significantly increased at both levels of salt stress compared with unstressed controls (Figure 2). There were no significant differences in Fv/Fm, NPQ, or Hd between the SS-S1 and SS-S2. Pretreatment with 24-epiBL by either Sew, Spw, or Diw had no effect on Fv/Fm, where Fv/Fm was equal to the CK level at both salinity stress levels (98.8% and 97.6% of CK) (Figure 2A). Hd improvement was reduced by all pretreatments with 24-epiBL, with the most significant effect in the Diw group at a 24-epiBL concentration of 1.04 µM (Figure 2B). NPQ decreased with increasing concentrations of 24-epiBL in the Sew, Spw, and Diw groups, although the values were higher than in SS-S1 and SS-S2. The treatment groups, Diw-0.21S1 (93.7% of SS-S1), Diw-0.62S1 (82.0% of SS-S1), Diw-1.04S1 (78.1% of SS-S1), Sew-1.04S2 (99.3% of SS-S2), and Diw-1.04S2 (90.9% of SS-0S2), showed a smaller decrease in NPQ in response to the two levels of NaCl (Figure 2C). The effects of Sc, Am, and BRc were obvious (p < 0.05), but the interactions in the above three factors were not significant (p > 0.05) in Fv/Fm, Hd, and NPQ.
3.3. RWC and MSI
Salinity stress resulted in reductions in RWC and MSI, in particular in MSI at 100 and 200 mM NaCl (Figure 3). Additionally, the significant reduction that compared 200 mM NaCl to 100 mM NaCl only appeared in MSI. In general, 24-epiBL pretreatments reversed these effects. At 100 mM NaCl, RWC was significantly increased in the Sew-1.04S1 and all Diw treatment groups compared with salt-stressed plants (SS-S1), whereas the mean RWC was similar to the CK group. Responses to 200 mM NaCl were similar and RWC was significantly greater than SS-S2 in the Sew-1.04S2 and Diw-1.04S2 groups (Figure 3A); the lower content of 24-epiBL in the Diw groups was obviously less effective in SS-S2 than in SS-S1. For MSI at 100 mM NaCl, only the Sew-0.21S1, Sew-0.62S1, and Spw-0.21S1 pretreatments showed no differences with the salt-stressed control, whereas, at 200 mM NaCl, all nine pretreatments were effective, and Sew-1.04S1, Diw-0.62S2, and Diw-1.04S2 showed values similar to the CK group (Figure 3B). The effects of Sc, Am, and BRc were obvious (p < 0.01), but the interactions in the above three factors were not significant (p > 0.05) in RWC and MSI.
3.4. Photosynthetic Pigments
Salinity stress had negative effects on pigment concentrations (Figure 4), although there were no statistically significant differences at different salinity stress levels. Relative to the unstressed control (CK), Chl a, Chl b, and total chlorophyll (Chl) were all reduced by 24%–28% under mild stress (SS-S1), and by 36%–40% at the higher stress level (SS-S2). All 24-epiBL pretreatments resulted in complete retention of these pigments at levels that were either statistically greater than the stressed control or even greater than the unstressed control (i.e., Diw-0.62 and Diw-1.04 at 100 mM NaCl) except for Chl b in the Sew-0.21S1 group, but the improvements were not obvious except Diw-1.04S1 in Chl a, Sew-1.04S1 and Diw-0.64/1.04S1 in Chl b, and Spw-1.04S1 and Diw-0.62/1.04S1 in Chl. Overall, the effects of regulation were Diw > Sew > Spw and the optimum concentration of 24-epiBL was 1.04 µM. The effect of Sc was obvious in Chl a, Chl b, and Chl (p < 0.01). The effects in Am (p = 0.03), BRc (p = 0.01), and Sc*Am (p = 0.01) were significant only in Chl b.
3.5. MDA, POD, and SOD in Leaves
Compared with the unstressed control group (CK), salt stress obviously increased the concentration of MDA by ~1.64-fold at 100 mM NaCl and by 2.04-fold at 200 mM NaCl (Figure 5A; SS-S1 and SS-S2 were significantly different from each other). Pretreatment with 24-epiBL was ineffective at reducing this increase to within just 1.18–1.92 times the unstressed value in all groups, with clear distinction between the concentration or developmental stage of application except the Sew-0.21S1,Spw-0.21S1, and Spw-0.21/0.62S2, which were no different to the salinity treatment separately; the optimal application method and 24-epiBL concentration were Diw and 1.04 µM, respectively, which resulted in a percentage modification of MDA of 72.4% of that of SS-S1 and 60.1% of that of SS-S2. The effect of Sc (p < 0.01), Am (p < 0.01), BRc (p < 0.01), and Sc*Am (p = 0.02) were significant in MDA (Figure 5A).
Salinity stress resulted in improved activities of POD, but these were only obvious in SS-S1 and the improvement of POD activities in SS-S1 was significantly better than SS-0S2. In general, 24-epiBL pretreatments increased POD activity and the regulated effect was much better in SS-S1 than SS-S2. The effective role of 24-epiBL pretreatments under 100 mM NaCl were Diw-1.04S1 > Spw-1.04S1 > Sew-1.04S1 > Diw-0.62S1, and only Diw-1.04S1 was significantly different than the other effective treatments. Under 200 mM NaCl, only Sew-0.21S2 and Spw-0.21S2 had no difference to SS-S2; other treatments in POD activities had no significant difference between the application method (Sew, Spw, Diw) and 24-epiBL contents (0.21, 0.62, 1.04 µM). The 24-epiBL pretreatment methods and concentrations in order of obvious impact on POD activity in SS-S2 were as follows: Sew-1.04S2 > Diw-1.04S2 > Sew-0.62S2 > Spw-1.04S2 > Spw-0.62S2 > Diw-0.62S2 > Diw-0.21S2. The effect of Sc (p < 0.01), Am (p < 0.01), BRc (p < 0.01), Sc*Am (p = 0.05), and the interaction in salt concentration and 24-epiBL application concentration (Sc*BRc) (p < 0.01) were significant in POD (Figure 5B).
NaCl treatment significantly increased SOD activity, although this was obviously lower in the SS-S2 than in the SS-S1, and the activities in SOD by 24-epiBL treatments in SS-S1 was higher than in SS-S2. Under 100 mM NaCl stress, the three 24-epiBL pretreatment approaches significantly increased SOD activity in contrast to CK, but only 1.04 µM 24-epiBL treatments resulted in higher SOD activity compared with SS-S1 (Diw-1.04S1 > Sew-1.04S1 > Spw-1.04S1). No significant effects were seen in the Sew-0.21S2, Spw-0.21S2, and Diw-0.21S2 groups compared with SS-S2. The optimum effects occurred in the Sew-1.04S2 (1.58-fold of SS-S2) and Diw-1.04S2 (1.74-fold of SS-S2) and they had no difference between each other. The effect of Sc (p < 0.01), Am (p < 0.01), BRc (p < 0.01), Sc*Am (p < 0.01), and the interaction in the 24-epiBL application method and 24-epiBL application concentration (Am*BRc) (p = 0.01) were significant in SOD (Figure 5C).
3.6. Variation in Na+, K+, and Ca2+ Content in Chloroplasts
There was a significant increase in Na+ content and decrease in K+ and Ca2+ accumulation in chloroplasts with increasing NaCl concentration (Figure 6). 24-epiBL reduced Na+ accumulation, although not significantly in each treatment, at both salinity levels, with Diw pretreatments resulting in more of a decrease in Na+ compared with Sew and Spw, especially at 200 mM NaCl. The significant effect only appeared in Sc in the Na+ increase (p < 0.01) (Figure 6A).
The higher the concentration of 24-epiBL, the greater the increase in K+, particularly in 200 mM NaCl, with the order of effect being Diw-1.04S2 > Sew-1.04S2 > Spw-1.04S2 > Diw-0.62S2. In 100 mM NaCl, although higher concentrations of 24-epiBL reduced the decrease in K+ under all 24-epiBL treatments, there was no significant difference compared with SS-S1. The effects of Sc (p < 0.01), Am (p = 0.03), BRc (p < 0.01), and Sc*BRc (p < 0.01) were significant in K+ reduction (Figure 6B).
The effect of 24-epiBL on regulating the decrease of Ca2+ was better in SS-S1 than in SS-S2. Under 100 mM NaCl stress, the three 24-epiBL application methods inhibited the reduction in Ca2+. The significantly effective inhibition was in the order compared to SS-S1: Diw-1.04S1 > Sew-1.04S1 > Diw-0.62S1. In the 200 mM NaCl treatment, most 24-epiBL treatment groups showed a reduction in the decrease of Ca2+ except Spw-0.21S2 and Spw-0.62S2, with the obvious effects compared to SS-S2 following the order: Diw-1.04S2 > Diw-0.62S2 > Sew-1.04S2 > Sew-0.62S2 > Diw-0.21S2 > Spw-1.04S2 (Figure 6C). The effects of Sc, Am, BRc, Sc*Am, and Sc*BRc were significant (p < 0.01 & p = 0.01) in regulating Ca2+ reduction.
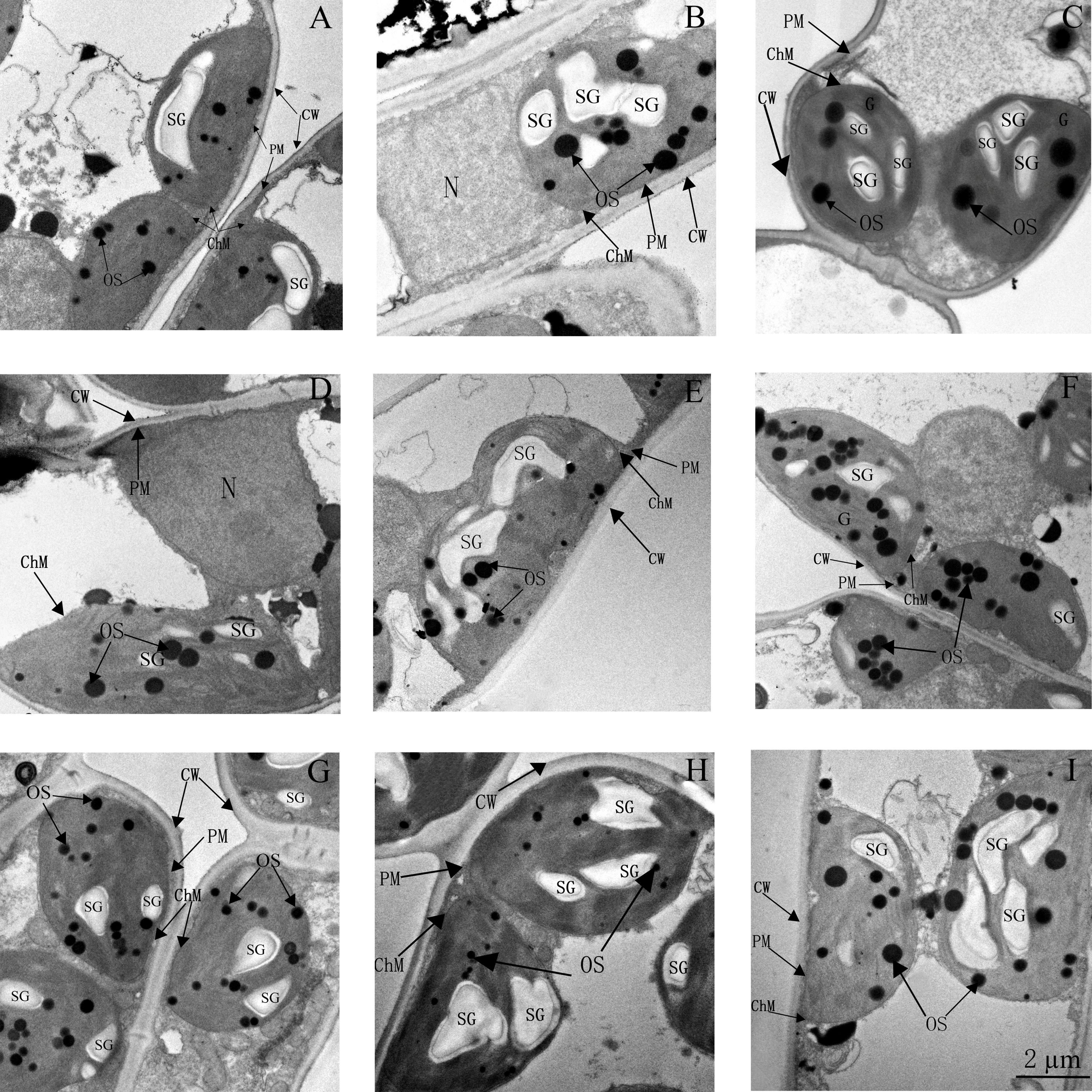
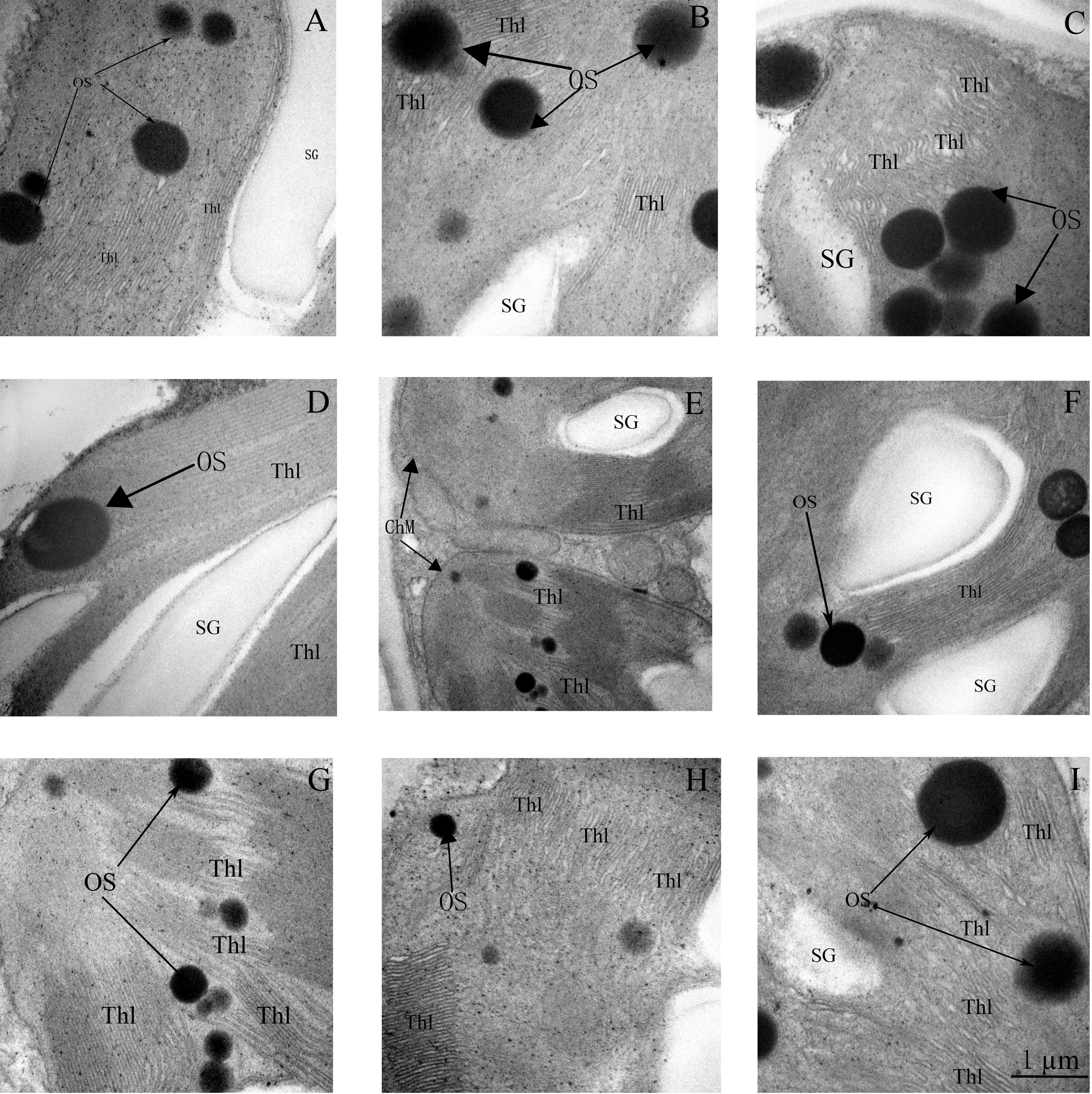
3.7. Ultrastructure of Chloroplasts and Thylakoids
Based on the physiological results detailed earlier, the optimal concentration of 24-epiBL for application was 1.04 µM. Therefore, ultrastructure analyses were only performed on plants grown using Sew, Spw, and Diw with 1.04 µM 24-epiBL. Alterations in the structure of chloroplasts and thylakoids induced by salt stress are shown in Figure 7 and Figure 8 relative to controls. The chloroplasts of salt-stressed plants were swollen and partly separated from the plasma membrane, with a more irregular lamellar structure to the thylakoids, particularly at 200 mM NaCl. Chloroplasts changed from being elliptical in shape to almost round, particularly at 200 mM NaCl. Plastoglobuli were swollen, but fewer in number compared with CK plants. Under higher salt stress (200mM NaCl), the cell wall was bent, and starch granules were smaller and less well defined (Figure 7 and Figure 8A–C).
These severe impacts on chloroplast and thylakoid structure were partly alleviated by 24-epiBL pretreatments. Application of 24-epiBL increased the number of plastoglobuli relative to stressed controls (Figure 7D–I). At 100 mM NaCl, all 24-epiBL pretreatments decreased the swelling of chloroplasts and resulted in more plastoglobuli, particularly in the Diw groups (compare panels D–F with B in Figure 7). The swelling of chloroplasts at 200 mM NaCl was significantly modified by Diw pretreatments (Figure 7G–I). Under 100 mM NaCl stress, the lamellae were more similar to CK plants in the Sew and Diw groups than in the Spw group. However, at higher salinity (200 mM NaCl stress), the loose lamellae were not modified significantly by 24-epiBL and Diw pretreatment resulted in less lamellar modification compared with Sew and Spw (compare panels D–F and G–I with B,C in Figure 8).
4. Discussion
Many studies have reported the benefits of 24-epiBL applications on plants, although studies have focused on herbaceous crops rather than woody plants and have used different doses and application methods. For example, foliar spraying of melon cultivars (Cucumis melo L.) with 2.1 µM 24-epiBL enabled the plants to adapt to higher temperatures [31], soaking radish seeds (Raphanus sativus L. cv. ‘Japanese White’) with 0.5–2 µM 24-epiBL enabled seedlings to adapt to zinc stress [32], and dipping the roots of wheat seedlings in 0.052–0.156 µM 24-epiBL improved their salt tolerance [33]. Thus, in the current study, we investigated the effects of 0.21–1.04 µM 24-epiBL pretreatments (seed soaking, foliar spraying, and root dipping) on the adaptation to different levels of salinity by R. pseudoacacia seedlings. Given that the number of potential treatment combinations was unwieldy, we focused on a key subset (Table 1) to examine the effects of 24-epiBL and to select the optimal pretreatment combination to inform the use of such treatments in the field.
4.1. Photosynthesis
NaCl inhibited photosynthesis (evidenced by a decrease in Pn, Gs, Ci/Ca, and WUEi compared with CK) combined with a reduction in RWC and pigment content (Chl a, Chl b, Chl). There was no significant difference between the impact of the two NaCl levels on the RWC and pigment content. Gs was less severely impacted than Pn by 100 mM NaCl, leading to a large decrease in WUEi; the reduction on Gs of 200 mM NaCl was much lower than that of 100 mM NaCl, Pn was similar at both salinity levels, whereas WUEi was obvious higher in 200 mM NaCl compared with SS-0S1.This suggests that mesophyll conductance (gm) at both levels of NaCl stress and stomatal conductance under 200 mM NaCl negatively impacted photosynthesis. This result is similar to other studies that showed that Gm was more seriously affected than Gs in poplars (Populus cathayana Rehder) under alkaline (75mM Na2CO3) stress compared with control plants [34]. The decrease in Fv/Fm observed in salt-stressed plants indicates a progressive reduction in the maximum quantum yield of photosynthesis [35]. The increases in NPQ and Hd were consistent with reduced rates of carbon fixation and greater energy dissipation as heat [36]. The above results were similar to the significant inhibition of Pn, Gs, and WUEi caused by 200 mM NaCl in Brassica napus leaves accompanied with the reduction in Chl a, Chl b, and total Chl after three days of salt stress [37]. 24-epiBL pretreatments improved the photosynthetic rate by regulating the limitation on Gm and Gs given the significant reduction in Ci/Ca and increase in WUEi in response to 100 mM NaCl and the change in Gs in response to 200 mM NaCl. The most positive effects on photosynthesis were seen in the Diw-1.04S1 and Diw-1.04S2 groups, also indicated by the reduction in Hd and changes in Fv/Fm and NPQ. 24-epiBL application also modified the changes in pigment contents (21.1%–51.1% increase in Cha, 12.5%–91.6% in Chb, and 15.5%–62.1% in Chl compared with SS-S1; and 23%–58.6% in Cha, 4.8%–25.8% in Chb, and 16.9%–49.4% in Chl compared with SS-S2). In grape seedlings (V. vinifera L.) with 10% PEG stress, the application of 0.105–0.42 µM 24-epiBL significantly increased the chlorophyll content (Chl a, Chl b, and Chl), maximal fluorescence, Fv/Fm, and NPQ, to improve drought tolerance [38]. Thussagunpanit and co-workers sprayed the foliage of rice (Oryza sativa L.) with 0.1 µM 24-epiBL and reported that 24-epiBL improved Pn, Gs, Fv/Fm, and Chl a, Chl b, and Chl contents with a reduction in Ci to protect the plants from heat stress (47 °C) [39].
4.2. Antioxidative Effect in Leaves
NaCl increased the MDA content (1.64~2.04-fold compared with CK) and the activity of POD and SOD (1.05~1.92-fold and 1.23~1.36-fold compared with CK, respectively), although POD activity was not significantly different under 200 mM NaCl stress, whereas that of SOD and MDA was significant at both levels of NaCl stress compared with controls. These results suggest that NaCl was harmful to the leaves and induced oxidative stress at both levels of NaCl. A previous study reported that 150 mM NaCl increased the MDA content and SOD activity in European larch needles (Larix decidua Mill.), also suggesting that salinity induces oxidative stress that negatively impacts on plant growth (i.e., reduced the increases in stem length, fresh weight, and leaf water content compared with controls) [40]. Application of 24-epiBL modified the MDA content (reduced by 6.3%–27.6% in SS-S1 and 5.6%–39.8% in SS-S2), antioxidant enzyme activity (increased the activities of SOD and POD by 0.8%–12% and 39.4%–90.4%, respectively, compared with SS-S1 and by 1.4%–41% and 43.1%–116.8% compared with SS-S2, respectively). The weaker changes recorded at 100 mM NaCl and stronger changes at 200 mM NaCl suggested the moderate tolerance of R. pseudoacacia seedlings to 100 mM NaCl stress and that 24-epiBL could increase the tolerance of the plant to both levels of salt stress, although this was more effective under the 200 mM NaCl stress. The optimal effects were seen with Diw pretreatments and 1.04 µmol L−1 24-epiBL. These results indicate that 24-epiBL is able to modify the oxidative stress induced by abiotic stressors. The antioxidant effects of BRs were also reported by Xia et al., who showed that overexpression of the BR biosynthetic gene, Dwarf, enhanced the ratio of reduced/oxidized 2-cysteine peroxiredoxin (2-Cys Prx) and activated antioxidant enzymes, including APX, MDAR, DHAR, and GR, in correlation with a reduction in H2O2 in tomatos (Solanum lycopersicum L.) under chilling stress [41]. Spraying 1µM 24-epiBL onto a single-cross maize hybrid PMH 3 (LM 17 × LM 14) improved its adaptation to heat stress by arresting protein degradation and improving the cell membrane stability, accompanied by modulation of the biochemical activities of antioxidant enzymes (CAT, SOD, and POD), highlighting BRs as anti-stress agents [42]. The presence of endogenous 28-norbrassinolide in a resistant (~4.8 pg mg−1 fresh mass) genotype (CE704) of maize (Zea mays L.) resulted in higher chlorophyll and proline contents and lower MDA content, and reduced membrane injury compared with a sensitive maize genotype (2023; ~1.5 pg mg−1 fresh mass) under drought stress [43], also confirming the ability of BRs to regulate antioxidative effects under different stressors, including salinity.
4.3. Ion Toxicity and Chloroplast Ultrastructure
NaCl induces an ion imbalance in leaves, with significant Na+ accumulation in plant organs in most cases accompanying decreases in K+ and Ca2+ and damage to chloroplasts. The improved salt tolerance of barley (H. vulgare) seedlings compared with wheat (T. turgidum L. ssp. durum Desf.) correlated with a higher photosynthetic capacity and higher K+ and lower Na+ in the cytoplasm of mesophyll cells [44]. In the current study, salinity caused 31.7%–54.1% and 9.2%–15.4% reductions in MSI and RWC, respectively, and a 40–140 mg∙ mg−1prot increase in Na+ accompanied by a significant decrease in K+ (2~5.5 mg∙mg−1prot) and Ca2+ (18-33 mg∙mg−1prot) in chloroplasts regardless of NaCl concentration (Figure 6). These results were similar to the increased Na+ content reported in chloroplasts of poplar leaves (P. euphratica and P. popularis) accompanied by reduced Pn under salt stress [45]. These results support the occurrence of ion toxicity in leaves and chloroplasts in response to saline stress. Na+ accumulation also induced changes to the chloroplasts, such as swollen chloroplasts partly separated from the plasma membrane and the more irregular lamellar structure of thylakoids (Figs 7 and 8). This was similar to the swollen chloroplasts and the looser thylakoid lamellar structures reported in diploid (2×) R. pseudoacacia L., as well as increases in Na+ and decreases in K+/Na+ in response to 500 mM NaCl stress for 15 days [14].
Three 24-epiBL pretreatment types reversed the changes to the chloroplasts (i.e., decreased swelling, maintained normal size of starch granules, increased the number of plastoglobuli, and kept a normal lamellar structure) and resulted in higher Pn, effects that were similar to putrescine (Put) in regulating the salt tolerance of C. sativus by maintaining regular thylakoid membrane structures with normal Pn and the photochemical efficiency of PSII, and reducing the ion toxicity (Na+ contents in chloroplasts) under 75 mM NaCl [46]. These results were also similar to the ability of silicon (Si) to maintain a normal chloroplast structure and normal thylakoid lamellae, accompanied by higher Chl a and Chl b contents in leaves and lower electrolyte leakage in response to drought in tomatos (S. lycopersicum) [47]. A study also reported the ability of exogenous K2SiO3 to maintain chloroplast lamellae and reduce foliar Na+ levels to increase Pn and Gs of L. japonica under 100 and 200 mM NaCl stress [12]. In the current study, application of 24-epiBL inhibited the increase in Na+ and the reduction of K+ and Ca2+ particularly in response to 200 mM NaCl. Ca2+ levels showed obvious changes in response to the different 24-epiBL treatments, whereas K+ only showed significant differences in response to 200 mM NaCl, and the Diw pretreatment group showed more significant responses overall than either the Sew or Spw groups. The most effective pretreatments were Diw-1.04 S1 (64.2% Na+ increase, 75.2% K+ decrease, and 37.8% Ca2+ decrease compared with SS-S1) and Diw-1.04 S2 (87.2% Na+ increase, 60.9% K+ decrease, and 39.7% Ca2+ decrease compared with SS-S2). Although the concentration of 24-epiBL in our pretreatment groups differed, the effects were similar to a study in which foliage was sprayed with 0.01–0.1 µM 24-epiBL to improve the salt tolerance (to 250 mM NaCl) of perennial ryegrass (Lolium perenne L.), resulting in higher concentrations of leaf proline, K+, Mg2+, Ca2+, and chlorophyll, increased Pn, and a weak reduction in Na+ [48]. In addition, soaking seeds in 0.0001~1µM 24-epiBL improved the tolerance of R. sativus L. seedlings to mercury (Hg) by increasing Na+ and K+ and activating the antioxidative ability of the plant (i.e., reduced MDA and increased activities of POD, SOD, glutathione reductase (GR), and dehydroascorbate reductase (DHAR)) [49]. Thus, we can conclude that 24-epiBL has the ability to improve the tolerance of plants to abiotic stress by maintaining the ion balance and overcoming any ion toxicity.
5. Conclusions
Our results showed that salt stress greatly induced photoinhibition in R. pseudoacacia seedlings as a result of oxidative stress and ion toxicity. Active oxygen can be generated from the interaction of O2 and an obstructed electron transport chain [50], damaging various photosynthetic components [51]. Ion toxicity also harms leaves and chloroplasts when combined with oxidative effects caused by stressors [45,52]. Exogenous 24-epiBL pretreatments alleviated this photoinhibition and stabilized the chloroplast structure, by reducing ion toxicity and increasing the activities of antioxidant enzymes. Application of 24-epiBL by either Sew, Spw, or Diw was effective, although the Diw approach resulted in the optimal overall performance, with the effects of the 24-epiBL concentration being in the order 1.04 > 0.62 > 0.21µM.
Author Contributions
J.Y. led to the submission, acquired data, wrote the draft and revised the manuscript; Z.F. did the part of experiment and got some data used in this paper, L.Z. did the part of experiment and got some data used in this paper, Z.Z. did the modification of manuscript and J.Z. conceived and designed the work and applied all the funding.
Funding
Financial support for this study was provided by the Jiangsu Agriculture Science and Technology Innovation Fund (Grant No. CX(17)1004), National Special Fund for Forestry Scientific Research in the Public Interest (Grant No. 201504406), Major Fund for Natural Science of Jiangsu Higher Education Institutions (Grant No. 15KJA220004), National Foundation of Forestry Science and Technology Popularization (Grant No. [2015]17), Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD), and the Doctorate Fellowship Foundation of Nanjing Forestry University.
Acknowledgments
Financial support for this study was provided by the Jiangsu Agriculture Science and Technology Innovation Fund (Grant No. CX(17)1004), National Special Fund for Forestry Scientific Research in the Public Interest (Grant No. 201504406), Major Fund for Natural Science of Jiangsu Higher Education Institutions (Grant No. 15KJA220004), National Foundation of Forestry Science and Technology Popularization (Grant No. [2015]17), Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD), and the Doctorate Fellowship Foundation of Nanjing Forestry University. We thank International Science Editing (http://www.internationalscienceediting.com) for editing this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Shahid, S.A. Developments in spoil salinity assessment, modeling, mapping, and monitoring from regional to submicroscopic scales. In Developments in Soil Salinity Assessment and Reclamation; Shahid, S.A., Abelfattah, M.A., Taha, F.K., Eds.; Springer: Dordrecht, The Netherland, 2013; pp. 3–44. [Google Scholar] [CrossRef]
- Wang, W.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef]
- Li, J.; Pu, L.; Han, M.; Zhu, M.; Zhang, R.; Xiang, Y. Soil salinization research in China: Advances and prospects. J. Geogra. Sci. 2014, 24, 943–960. [Google Scholar] [CrossRef]
- Chen, X.; Shen, Q.; Xu, Y. Hydraulic properties of typical salt-affected soils in Jiangsu Province, China. Front. Environ. Sci. Eng. China 2007, 1, 443–447. [Google Scholar] [CrossRef]
- DeGomez, T.; Wagner, M.R. Culture and use of black locust. HortTechnology 2001, 11, 279–288. [Google Scholar] [CrossRef]
- Liu, W.N.; Jiang, H.Y.; Yang, M.S. Selection of salt—tolerance on Robinia pseudoacacia seedings and impact of salt stress on seedlings’ physiological characteristics. J. Agric. Univ. Hebei 2010, 3, 62–66. [Google Scholar]
- Sitzia, T.; Campagnaro, T.; Dainese, M.; Cierjacks, A. Forest Ecology and Management Plant species diversity in alien black locust stands: A paired comparison with native stands across a north-Mediterranean range expansion. For. Ecol. Manag. 2012, 285, 85–91. [Google Scholar] [CrossRef]
- Tateno, R.; Tokuchi, N.; Yamanaka, N.; Du, S.; Otsuki, K.; Shimamura, T.; Xue, Z.; Wang, S.; Hou, Q. Comparison of litterfall production and leaf litter decomposition between an exotic black locust plantation and an indigenous oak forest near Yan’an on the Loess Plateau, China. For. Ecol. Manag. 2007, 241, 84–90. [Google Scholar] [CrossRef]
- Akatov, V.V.; Akatova, T.V.; Shadzhe, A.E. Robinia pseudoacacia L. in the Western Caucasus. Rus. J. Biol. Invasion 2016, 7, 105–118. [Google Scholar] [CrossRef]
- Holle, B.V.; Neill, C.; Largay, E.F.; Budreski, K.A.; Ozimec, B.; Clark, S.A.; Lee, K. Ecosystem legacy of the introduced N2-fixing tree Robinia pseudoacacia in a coastal forest. Oecologia 2013, 172, 915–924. [Google Scholar] [CrossRef]
- Mao, P.; Zhang, Y.; Cao, B.; Guo, L.; Shao, H.; Cao, Z.; Jiang, Q.; Wang, X. Effects of salt stress on eco-physiological characteristics in Robinia pseudoacacia based on salt-soil rhizosphere. Sci. Total Environ. 2016, 568, 118–123. [Google Scholar] [CrossRef]
- Zhao, G.; Li, S.; Sun, X.; Wang, Y.; Chang, Z. The role of silicon in physiology of the medicinal plant (Lonicera japonica L.) under salt stress. Sci. Rep. 2015, 5, 12696. [Google Scholar] [CrossRef]
- Manivannan, A.; Soundararajan, P.; Muneer, S.; Ko, C.H.; Jeong, B.R. Silicon mitigates salinity stress by regulating the physiology, antioxidant enzyme activities, and protein expression in Capsicum annuum “Bugwang”. BioMed Res. Int. 2016, 2016. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, M.; Liu, L.; Meng, F. Physiological and proteomic responses of diploid and tetraploid black locust (Robinia pseudoacacia L.) subjected to salt stress. Int. J. Mol. Sci. 2013, 14, 20299–20325. [Google Scholar] [CrossRef]
- Xi, Z.; Wang, Z.; Fang, Y.; Hu, Z.; Hu, Y.; Deng, M.; Zhang, Z. Effects of 24-epibrassinolide on antioxidation defense and osmoregulation systems of young grapevines (V. vinifera L.) under chilling stress. Plant Growth Regul. 2013, 71, 57–65. [Google Scholar] [CrossRef]
- An, Y.; Zhou, H.; Zhong, M.; Sun, J.; Shu, S. Root proteomics reveals cucumber 24-epibrassinolide responses under Ca (NO3)2 stress. Plant Cell Rep. 2016, 35, 1081–1101. [Google Scholar] [CrossRef]
- Cui, L.; Zou, Z.; Zhang, J.; Zhao, Y.; Yan, F. 24-Epibrassinoslide enhances plant tolerance to stress from low temperatures and poor light intensities in tomato (Lycopersicon esculentum Mill.). Funct. Integr. Genomic. 2016, 16, 29–35. [Google Scholar] [CrossRef]
- Talaat, N.B.; Shawky, B.T. 24-Epibrassinolide ameliorates the saline stress and improves the productivity of wheat (Triticum aestivum L.). Environ. Exp. Bot. 2012, 82, 80–88. [Google Scholar] [CrossRef]
- Azhar, N.; Su, N.; Shabala, L.; Shabala, S. Exogenously applied 24-epibrassinolide (EBL) ameliorates detrimental effects of salinity by reducing K+ efflux via depolarization-activated K+ channels. Plant Cell Physiol. 2017, 58, 802–810. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Circ. Calif. Agr. Ex. Stn. 1950, 347, 1–32. [Google Scholar]
- Yue, J.M.; You, Y.H.; Zhang, L.; Fu, Z.Y.; Wang, J.P.; Zhang, J.C.; Guy, R.D. Exogenous 24-epibrassinolide alleviates effects of salt stress on chloroplasts and photosynthesis in Robinia pseudoacacia L. seedlings. J. Plant Growth Reg. 2018, 1–14. [Google Scholar] [CrossRef]
- Sairam, R.K. Effects of homobrassinolide application on plant metabolism and grain yield under irrigated and moisture-stress conditions of two wheat varieties. Plant Growth Regul. 1994, 306, 173–181. [Google Scholar] [CrossRef]
- Hayat, S.; Ali, B.; Aiman Hasan, S.; Ahmad, A. Brassinosteroid enhanced the level of antioxidants under cadmium stress in Brassica juncea. Environ. Exp. Bot. 2007, 60, 33–41. [Google Scholar] [CrossRef]
- He, M.; Shi, D.; Wei, X.; Hu, Y.; Wang, T.; Xie, Y. Gender-related differences in adaptability to drought stress in the dioecious tree Ginkgo biloba. Acta Physiol. Plant. 2016, 38. [Google Scholar] [CrossRef]
- Fridovich, I. Superoxide Dismutases. Annu. Rev. Biochem. 1975, 44, 147–159. [Google Scholar] [CrossRef]
- Hammerschmidt, R.; Nuckles, E.M.; Kuć, J. Association of enhanced peroxidase activity with induced systemic resistance of cucumber to Colletotrichum lagenarium. Physiol. Plant Pathol. 1982, 20, 73–82. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Cerović, Z.G.; Plesnicar, M. An improved procedure for the isolation of intact chloroplasts of high photosynthetic capacity. Biochem. J. 1984, 223, 543–545. [Google Scholar] [CrossRef] [Green Version]
- Song, X.S.; Tiao, C.L.; Shi, K.; Mao, W.H.; Ogweno, J.O.; Zhou, Y.H.; Yu, J.Q. The response of antioxidant enzymes in cellular organelles in cucumber (Cucumis sativus L.) leaves to methyl viologen-induced photo-oxidative stress. Plant Growth Regul. 2006, 49, 85–93. [Google Scholar] [CrossRef]
- Abo-Ogiala, A.; Carsjens, C.; Diekmann, H.; Fayyaz, P.; Herrfurth, C.; Feussner, I.; Polle, A. Temperature-induced lipocalin (TIL) is translocated under salt stress and protects chloroplasts from ion toxicity. J. Plant Physiol. 2014, 171, 250–259. [Google Scholar] [CrossRef]
- Zhang, Y.P.; Zhu, X.H.; Ding, H.D.; Yang, S.J.; Chen, Y.Y. Foliar application of 24-epibrassinolide alleviates high-temperature-induced inhibition of photosynthesis in seedlings of two melon cultivars. Photosynthetica 2013, 51, 341–349. [Google Scholar] [CrossRef]
- Ramakrishna, B.; Rao, S.S.R. 24-Epibrassinolide alleviated zinc-induced oxidative stress in radish (Raphanus sativus L.) seedlings by enhancing antioxidative system. Plant Growth Regul. 2012, 68, 249–259. [Google Scholar] [CrossRef]
- Ali, Q.; Athar, H.U.R.; Ashraf, M. Modulation of growth, photosynthetic capacity and water relations in salt stressed wheat plants by exogenously applied 24-epibrassinolide. Plant Growth Regul. 2008, 56, 107–116. [Google Scholar] [CrossRef]
- Xu, G.; Huang, T.F.; Zhang, X.L.; Duan, B.L. Significance of mesophyll conductance for photosynthetic capacity and water-use efficiency in response to alkaline stress in Populus cathayana seedlings. Photosynthetica 2013, 51, 438–444. [Google Scholar] [CrossRef] [Green Version]
- Kalaji, H.M.; Govindjee; Bosa, K.; Koscielniak, J.; Zuk-Golaszewska, K. Effects of salt stress on photosystem II efficiency and CO2 assimilation of two Syrian barley landraces. Environ. Exp. Bot. 2011, 73, 64–72. [Google Scholar] [CrossRef]
- Oxborough, K. Imaging of chlorophyll a fluorescence: Theoretical and practical aspects of an emerging technique for the monitoring of photosynthetic performance. J. Exp. Bot. 2004, 55, 1195–1205. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Shao, M.; He, Y.; Guan, R.; Chu, P.; Jiang, H. Proteome dynamics and physiological responses to short-term salt stress in brassica napus leaves. PLoS ONE 2015, 10. [Google Scholar] [CrossRef]
- Wang, Z.; Zheng, P.; Meng, J.; Xi, Z. Effect of exogenous 24-epibrassinolide on chlorophyll fluorescence, leaf surface morphology and cellular ultrastructure of grape seedlings (Vitis vinifera L.) under water stress. Acta Physiol. Plant. 2015, 37, 1–12. [Google Scholar] [CrossRef]
- Thussagunpanit, J.; Jutamanee, K.; Kaveeta, L.; Chai-arree, W.; Pankean, P.; Homvisasevongsa, S.; Suksamrarn, A. Comparative effects of brassinosteroid and brassinosteroid mimic on improving photosynthesis, lipid peroxidation, and rice seed set under heat stress. J. Plant Growth Regul. 2015, 34, 320–331. [Google Scholar] [CrossRef]
- Plesa, I.; González-Orenga, S.; Al Hassan, M.; Sestras, A.; Vicente, O.; Prohens, J.; Sestras, R.; Boscaiu, M. Effects of Drought and Salinity on European Larch (Larix decidua Mill.) Seedlings. Forests 2018, 9, 320. [Google Scholar] [CrossRef]
- Xia, X.J.; Fang, P.P.; Guo, X.; Qian, X.J.; Zhou, J.; Shi, K.; Zhou, Y.H.; Yu, J.Q. Brassinosteroid-mediated apoplastic H2O2-glutaredoxin 12/14 cascade regulates antioxidant capacity in response to chilling in tomato. Plant Cell Environ. 2018, 41, 1052–1064. [Google Scholar] [CrossRef]
- Yadava, P.; Kaushal, J.; Gautam, A.; Parmar, H.; Singh, I. Physiological and biochemical effects of 24-epibrassinolide on heat-stress adaptation in Maize (Zea mays L.). Nat. Sci. 2016, 8, 171–179. [Google Scholar] [CrossRef]
- Tůmová, L.; Tarkowská, D.; Řřová, K.; Marková, H.; Kočová, M.; Rothová, O.; čečetka, P.; Holá, D. Drought-tolerant and drought-sensitive genotypes of maize (Zea mays L.) differ in contents of endogenous brassinosteroids and their drought-induced changes. PLoS ONE 2018, 13, 1–22. [Google Scholar] [CrossRef] [PubMed]
- James, R.A.; Munns, R.; Von Caemmerer, S.; Trejo, C.; Miller, C.; Condon, T. Photosynthetic capacity is related to the cellular and subcellular partitioning of Na+, K+ and Cl− in salt-affected barley and durum wheat. Plant Cell Environ. 2006, 29, 2185–2197. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Chen, S.; Deng, L.; Fritz, E.; Hüttermann, A.; Polle, A. Leaf photosynthesis, fluorescence response to salinity and the relevance to chloroplast salt compartmentation and anti-oxidative stress in two poplars. Trees Struct. Funct. 2007, 21, 581–591. [Google Scholar] [CrossRef]
- Shu, S.; Yuan, Y.; Chen, J.; Sun, J.; Zhang, W.; Tang, Y.; Zhong, M.; Guo, S. The role of putrescine in the regulation of proteins and fatty acids of thylakoid membranes under salt stress. Sci. Rep. 2015, 5, 1–16. [Google Scholar] [CrossRef]
- Cao, B.L.; Ma, Q.; Zhao, Q.; Wang, L.; Xu, K. Effects of silicon on absorbed light allocation, antioxidant enzymes and ultrastructure of chloroplasts in tomato leaves under simulated drought stress. Sci. Hortic. 2015, 194, 53–62. [Google Scholar] [CrossRef]
- Wu, W.; Zhang, Q.; Ervin, E.H.; Yang, Z.; Zhang, X. Physiological mechanism of enhancing salt stress tolerance of Perennial ryegrass by 24-epibrassinolide. Fron. Plant Sci. 2017, 8, 1–11. [Google Scholar] [CrossRef]
- Kapoor, D.; Rattan, A.; Gautam, V.; Bhardwaj, R. Mercury-induced changes in growth, metal & ions uptake, photosynthetic pigments, osmoprotectants and antioxidant defence system in Raphanus sativus L. seedlings and role of steroid hormone in stress amelioration. J. Pharmacogn. Phytochem. 2016, 5, 259–265. [Google Scholar]
- Fisher, A.B. Redox signaling across cell membranes. Antioxid. Redox Signal. 2009, 11, 1349–1356. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Wali, M.; Gunsè, B.; Llugany, M.; Corrales, I.; Abdelly, C.; Poschenrieder, C.; Ghnaya, T. High salinity helps the halophyte Sesuvium portulacastrum in defense against Cd toxicity by maintaining redox balance and photosynthesis. Planta 2016, 244, 333–346. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effects of exogenous 24-epibrassinolide (24-epiBL) on (A) net photosynthetic rate (Pn), (B) stomatal conductance (Gs), (C) ratio of intercellular to ambient CO2 concentration (Ci/Ca), and (D) intrinsic water use efficiency (WUEi) of leaves of black locust seedlings at 100 (SS-S1) or 200 (SS-S2) mM NaCl. Each panel shows data for the zero salt control (CK, grey bar) and salt-stressed control (S1 & S2, black bar) and salt-stressed plants pretreated with either 0.21, 0.62, or 1.04 mM 24-epiBL by seed soaking (Sew, yellow bars), foliar spraying (Spw, green bars), or roots dipping (Diw, red bars). Each bar represents the mean (± standard deviation (SE)) of three replicates. Different letters above the bars show significant differences between means within each panel (p < 0.05). The p values are for the salt concentration effect (Sc), the 24-epiBL application method effect (Am), the 24-epiBL application concentration effect (BRc), the interaction effect between the salt concentration and 24-epiBL application method (Sc*Am), the interaction effect between the salt concentration and 24-epiBL application concentration (Sc*BRc), the interaction effect between the 24-epiBL application method and 24-epiBL application concentration (Am*BRc), and the interaction effect among the salt concentration, 24-epiBL application method, and 24-epiBL application concentration (Sc*Am*BRc), respectively. The significant effect was p < 0.05. Control (CK); 100 mM NaCl treatment (S1); 200 mM NaCl treatment (S2); treatments with 24-epiBL (0.21, 0.62, 1.04 µM) under 100 mM NaCl stress (0.21S1, 0.62S1, 1.04S1); treatments with 24-epiBL (0.21, 0.62, 1.04 µM) under 200 mM NaCl stress (0.21S2, 0.62S2, 1.04S2).
Figure 1.
Effects of exogenous 24-epibrassinolide (24-epiBL) on (A) net photosynthetic rate (Pn), (B) stomatal conductance (Gs), (C) ratio of intercellular to ambient CO2 concentration (Ci/Ca), and (D) intrinsic water use efficiency (WUEi) of leaves of black locust seedlings at 100 (SS-S1) or 200 (SS-S2) mM NaCl. Each panel shows data for the zero salt control (CK, grey bar) and salt-stressed control (S1 & S2, black bar) and salt-stressed plants pretreated with either 0.21, 0.62, or 1.04 mM 24-epiBL by seed soaking (Sew, yellow bars), foliar spraying (Spw, green bars), or roots dipping (Diw, red bars). Each bar represents the mean (± standard deviation (SE)) of three replicates. Different letters above the bars show significant differences between means within each panel (p < 0.05). The p values are for the salt concentration effect (Sc), the 24-epiBL application method effect (Am), the 24-epiBL application concentration effect (BRc), the interaction effect between the salt concentration and 24-epiBL application method (Sc*Am), the interaction effect between the salt concentration and 24-epiBL application concentration (Sc*BRc), the interaction effect between the 24-epiBL application method and 24-epiBL application concentration (Am*BRc), and the interaction effect among the salt concentration, 24-epiBL application method, and 24-epiBL application concentration (Sc*Am*BRc), respectively. The significant effect was p < 0.05. Control (CK); 100 mM NaCl treatment (S1); 200 mM NaCl treatment (S2); treatments with 24-epiBL (0.21, 0.62, 1.04 µM) under 100 mM NaCl stress (0.21S1, 0.62S1, 1.04S1); treatments with 24-epiBL (0.21, 0.62, 1.04 µM) under 200 mM NaCl stress (0.21S2, 0.62S2, 1.04S2).

Figure 2.
Effects of exogenous 24-epiBL on (A) maximum yield of photosystem II (PSII) (Fv/Fm), (B) thermal dissipation (Hd), and (C) non-photochemical quenching (NPQ) of leaves of black locust seedlings at 100 (SS-S1) or 200 (SS-S2) mM NaCl. Other details as in Figure 1.
Figure 2.
Effects of exogenous 24-epiBL on (A) maximum yield of photosystem II (PSII) (Fv/Fm), (B) thermal dissipation (Hd), and (C) non-photochemical quenching (NPQ) of leaves of black locust seedlings at 100 (SS-S1) or 200 (SS-S2) mM NaCl. Other details as in Figure 1.

Figure 3.
Effects of exogenous 24-epiBL on (A) relative water content (RWC) and (B) membrane stability index (MSI) of leaves of black locust seedlings at 100 (SS-S1) or 200 (SS-S2) mM NaCl. Other details as in Figure 1.
Figure 3.
Effects of exogenous 24-epiBL on (A) relative water content (RWC) and (B) membrane stability index (MSI) of leaves of black locust seedlings at 100 (SS-S1) or 200 (SS-S2) mM NaCl. Other details as in Figure 1.

Figure 4.
Effects of exogenous 24-epiBL on (A) chlorophyll a (Chl a), (B) chlorophyll b (Chl b), and (C) total chlorophyll (Chl) of leaves of black locust seedlings at 100 (SS-S1) or 200 (SS-S2) mM NaCl. Other details as in Figure 1.
Figure 4.
Effects of exogenous 24-epiBL on (A) chlorophyll a (Chl a), (B) chlorophyll b (Chl b), and (C) total chlorophyll (Chl) of leaves of black locust seedlings at 100 (SS-S1) or 200 (SS-S2) mM NaCl. Other details as in Figure 1.

Figure 5.
Effects of exogenous 24-epiBL on (A) malondialdehyde (MDA), (B) peroxidase (POD), and (C) superoxide dismutase (SOD) of leaves of black locust seedlings at 100 (SS-S1) or 200 (SS-S2) mM NaCl. Other details as in Figure 1.
Figure 5.
Effects of exogenous 24-epiBL on (A) malondialdehyde (MDA), (B) peroxidase (POD), and (C) superoxide dismutase (SOD) of leaves of black locust seedlings at 100 (SS-S1) or 200 (SS-S2) mM NaCl. Other details as in Figure 1.

Figure 6.
Effects of exogenous 24-epiBL on (A) the increase of Na+, (B) the decrease of K+, and (C) decrease of Ca2+ in chloroplasts of leaves of black locust seedlings at 100 (SS-S1) or 200 (SS-S2) mM NaCl. Other details as in Figure 1.
Figure 6.
Effects of exogenous 24-epiBL on (A) the increase of Na+, (B) the decrease of K+, and (C) decrease of Ca2+ in chloroplasts of leaves of black locust seedlings at 100 (SS-S1) or 200 (SS-S2) mM NaCl. Other details as in Figure 1.

Figure 7.
Ultrastructure of chloroplasts of leaves of black locust seedlings at three levels of salt stress (0, 100, or 200 mM NaCl), or with salt stress imposed after pretreatment with 1.04 µM 24-epiBL by either seed soaking, foliar spraying, or roots dipping (see Table 1 for treatment designation). (A) CK; (B) SS-S1; (C) SS-S2; (D) Sew-1.04S1; (E) Spw-1.04S1; (F) Diw-1.04S1; (G) Sew-1.04S2; (H) Spw-1.04S2; (I) Diw-1.04S2. CW: cell wall, PM: plasma membrane, ChM: chloroplast membrane, SG: starch granule, OS: osmiophilic plastoglobuli. Scale bars for chloroplasts are 2µm. Other details as in Figure 1.
Figure 7.
Ultrastructure of chloroplasts of leaves of black locust seedlings at three levels of salt stress (0, 100, or 200 mM NaCl), or with salt stress imposed after pretreatment with 1.04 µM 24-epiBL by either seed soaking, foliar spraying, or roots dipping (see Table 1 for treatment designation). (A) CK; (B) SS-S1; (C) SS-S2; (D) Sew-1.04S1; (E) Spw-1.04S1; (F) Diw-1.04S1; (G) Sew-1.04S2; (H) Spw-1.04S2; (I) Diw-1.04S2. CW: cell wall, PM: plasma membrane, ChM: chloroplast membrane, SG: starch granule, OS: osmiophilic plastoglobuli. Scale bars for chloroplasts are 2µm. Other details as in Figure 1.

Figure 8.
Ultrastructure of thylakoid membranes of leaves of black locust seedlings at three levels of salt stress (0, 100, or 200 mM NaCl), or with salt stress imposed after pretreatment with 1.04 µM 24-epiBL by either seed soaking, foliar spraying, or roots dipping (see Table 1 for treatment designation). (A) CK; (B) SS-S1; (C) SS-S2; (D) Sew-1.04S1; (E) Spw-1.04S1; (F) Diw-1.04S1; (G) Sew-1.04S2; (H) Spw-1.04S2; (I) Diw-1.04S2. Thl: thylakoid lamella, SG: starch granule, OS: osmiophilic plastoglobuli. Scale bars for thylakoids are 1µm. Other details are the same as Figure 1.
Figure 8.
Ultrastructure of thylakoid membranes of leaves of black locust seedlings at three levels of salt stress (0, 100, or 200 mM NaCl), or with salt stress imposed after pretreatment with 1.04 µM 24-epiBL by either seed soaking, foliar spraying, or roots dipping (see Table 1 for treatment designation). (A) CK; (B) SS-S1; (C) SS-S2; (D) Sew-1.04S1; (E) Spw-1.04S1; (F) Diw-1.04S1; (G) Sew-1.04S2; (H) Spw-1.04S2; (I) Diw-1.04S2. Thl: thylakoid lamella, SG: starch granule, OS: osmiophilic plastoglobuli. Scale bars for thylakoids are 1µm. Other details are the same as Figure 1.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The different treatments’ details.
| Name | Seeds Pretreatment | Seedling Pretreatment | Salt Stress | |
|---|---|---|---|---|
| 24-epiBL(µM) by Seeds Soaking | 24-epiBL(µM) by Foliar Spraying | 24-epiBL(µM) by Roots Dipping | NaCl (mM) | |
| CK | — | — | — | |
| SS-S1 | — | — | 100 | |
| SS-S2 | — | — | 200 | |
| Sew-0.21S1 | 0.21 | — | 100 | |
| Sew-0.62S1 | 0.62 | — | 100 | |
| Sew-1.04S1 | 1.04 | — | 100 | |
| Spw-0.21S1 | 0.21 | 100 | ||
| Spw-0.62S1 | 0.62 | 100 | ||
| Spw-1.04S1 | 1.04 | 100 | ||
| Diw-0.21S1 | 0.21 | 100 | ||
| Diw-0.62S1 | 0.62 | 100 | ||
| Diw-1.04S1 | 1.04 | 100 | ||
| Sew-0.21S2 | 0.21 | — | 200 | |
| Sew-0.62S2 | 0.62 | — | 200 | |
| Sew-1.04S2 | 1.04 | — | 200 | |
| Spw-0.21S2 | 0.21 | 200 | ||
| Spw-0.62S2 | 0.62 | 200 | ||
| Spw-1.04S2 | 1.04 | 200 | ||
| Diw-0.21S2 | 0.21 | 200 | ||
| Diw-0.62S2 | 0.62 | 200 | ||
| Diw-1.04S2 | 1.04 | 200 | ||
Abbreviations: Control (CK); 100 mM NaCl treatment (SS-S1); 200 mM NaCl treatment (SS-S2); seeds soaking with 24-epiBL (0.21, 0.62, 1.04 µM) under 100 mM NaCl stress (Sew-0.21S1/0.62S1/1.04S1); foliar spraying with 24-epiBL (0.21, 0.62, 1.04 µM) under 100 mM NaCl stress (Spw-0.21S1/0.62S1/1.04S1); roots dipping with 24-epiBL (0.21, 0.62, 1.04 µM) under 100 mM NaCl stress (Diw-0.21S1/0.62S1/1.04S1); seeds soaking with 24-epiBL (0.21, 0.62, 1.04 µM) under 200 mM NaCl stress (Sew-0.21S2/0.62S2/1.04S2); foliar spraying with 24-epiBL (0.21, 0.62, 1.04 µM) under 200 mM NaCl stress (Spw-0.21S2/0.62S2/1.04S2); roots dipping with 24-epiBL (0.21, 0.62, 1.04 µM) under 200 mM NaCl stress (Diw-0.21S2/0.62S2/1.04S2).
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yue, J.; Fu, Z.; Zhang, L.; Zhang, Z.; Zhang, J. The Positive Effect of Different 24-epiBL Pretreatments on Salinity Tolerance in Robinia pseudoacacia L. Seedlings. Forests 2019, 10, 4. https://doi.org/10.3390/f10010004
AMA Style
Yue J, Fu Z, Zhang L, Zhang Z, Zhang J. The Positive Effect of Different 24-epiBL Pretreatments on Salinity Tolerance in Robinia pseudoacacia L. Seedlings. Forests. 2019; 10(1):4. https://doi.org/10.3390/f10010004
Chicago/Turabian StyleYue, Jianmin, Zhiyuan Fu, Liang Zhang, Zihan Zhang, and Jinchi Zhang. 2019. "The Positive Effect of Different 24-epiBL Pretreatments on Salinity Tolerance in Robinia pseudoacacia L. Seedlings" Forests 10, no. 1: 4. https://doi.org/10.3390/f10010004
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.