Abstract
Research Highlights: The correlations between seed mass and the remaining physical properties were determined in selected fir species, and a method for seed sorting was proposed to promote uniform germination and emergence. Background and Objectives: Information about the variation and correlations between the physical properties of seeds is essential for designing and modeling seed processing operations such as seed separation. The aim of this study was to determine the range of variation in the physical properties of seeds in eleven fir species, and to analyze the correlations between these parameters to improve the seed sorting process. Materials and Methods: Terminal velocity, thickness, width, length, the angle of external friction and mass were determined in the seeds of 11 fir species. The measured parameters were used to calculate the geometric mean diameter, three aspect ratios, sphericity index and the specific mass of each seed. Results: The seeds of Sierra white fir where most similar, whereas the seeds of balsam fir differed most considerably from the seeds of the remaining fir species. In the majority of the analyzed fir species, seed mass was most strongly correlated with terminal velocity, followed by thickness. Conclusions: Fir seeds should be sorted primarily with the use of mesh sieves with longitudinal openings to obtain fractions with similar seed mass and to eliminate the need for dewinging.
1. Introduction
All fir species are indigenous to the northern hemisphere. They thrive in temperate and cool climates at altitudes not exceeding 4700 m above sea level. Firs are distributed mainly in North America (9 species), Central America (6 species), Siberia and Eastern Asia (17 species), the Mediterranean Region and the adjacent areas (8 species) [1,2,3,4,5,6,7].
Fir species reach maturity and begin to produce seeds at 12 to 50 years of age in low-density stands, and at a later stage in areas with high density of trees. Fir cones have a length of 7.5 to 25 cm, and their shape and size vary across species. Cones can be cylindrical or ovate-oblong, and they are often covered with resin. They mature in flowering years and break apart on trees, leaving the central stem on the branch. Fir seeds have wings for effective propagation, and wings are shed together with seed scales and bract scales [2,8,9]. The released seeds have irregular triangular shape, and seed wings are brittle and easily damaged. Despite the above, wings are strongly attached to seeds, and wing fragments are often retained on seeds. Fir seeds are classified as orthodox and have to be stored at low temperature (not higher than 4 °C) after their relative moisture content has to be reduced below 12%, preferably to around 8%. Long-term seed storage requires much lower temperatures, sometimes as low as −18 °C. Most fir species have dormant seeds, and dormancy is broken after cold-moist stratification [2,10,11,12].
Firs reproduce sexually via seeds. Consequently, their commercial production depends on seed viability and quality [13,14]. Previous research [15,16,17,18,19,20] shows that germination efficiency is significantly affected by seed mass. However, large seeds do not always germinate faster than small seeds. Practical experience indicates that plant uniformity can be improved by sorting seeds and sowing selected seed fractions to provide seedlings with equal opportunities for growth and to limit competition [21]. However, during large-scale production processes, it is difficult to divide seeds into fractions based only on their mass. Seeds can be effectively sorted using vibratory or vibratory-pneumatic separators [22] provided they differ in mass and have similar dimensions, or that they differ in dimensions and have similar mass. Fir seeds characterized by different mass and different dimensions may be difficult to sort. Therefore, other traits that are strongly correlated with seed mass should be found to facilitate seed separation.
The aim of this study was to determine the range of variation in the physical properties of seeds in eleven fir species, and to analyze the correlations between these parameters to improve the seed sorting process.
2. Materials and Methods
2.1. Sample Preparation
The physical properties of seeds of the following fir species were analyzed in this study (Figure 1): balsam fir (Abies balsamea (L.) P. Mill.), corkbark fir (Abies lasiocarpa var. arizonica (Hook.) Nutt.), Forrest’s fir (Abies forrestii Coltm.-Rog.), grand fir (Abies grandis (Doug. ex D. Don) Lindl.), Japanese fir (Abies firma Sieb. & Zucc.), Korean fir (Abies koreana E. H. Wilson), noble fir (Abies procera Rehd.), Sierra white fir (Abies concolor var. lowiana (Gord.) Lemm.), silver fir (Abies alba P. Mill.), subalpine fir (Abies lasiocarpa (Hook.) Nutt.) and white fir (Abies concolor (Gord.) Lindl. ex Hildebr.). Seeds were obtained in 2017 from the available resources of the Dendrona Company in Pęcice, a supplier of tree, shrub, perennial plant and herbaceous plant seeds.

Figure 1.
Seeds of: (a) balsam fir, (b) corkbark fir, (c) Forrest’s fir, (d) grand fir, (e) Japanese fir, (f) Korean fir, (g) noble fir, (h) Sierra white fir, (i) silver fir, (j) subalpine fir, (k) white fir.
Analytical specimens were obtained by halving from samples of approximately 100 g from each seed batch [16]. The initial sample of seeds was divided into two parts, and one part was randomly selected for successive halving. This procedure was repeated to obtain samples comprising approximately 100 seeds each.
2.2. Physical Properties
The physical properties of seeds were determined according to the method described by Kaliniewicz and Poznański [23] and Kaliniewicz et al. [24]. Terminal velocity (v) was measured using the Petkus K-293 pneumatic classifier (PETKUS Technologie GmbH, Wutha-Farnroda, Germany); seed length (L) and seed width (W) were measured under the MWM 2325 laboratory microscope; seed thickness (T) was determined with a clock thickness gauge; the angle of external friction (γ) was measured using a steel friction plate with surface roughness of Ra = 0.48 µm, placed on a horizontal plane; and seed mass (m) was determined on the WAA 100/C/2 laboratory scale (RADWAG, Radom, Polska). The angle of external friction was expressed as the mean value of two measurements performed for different positions of seeds which were placed on the steel friction plate with their longitudinal axis parallel and perpendicular to the direction of movement.
The results of the above measurements were used to calculate the geometric mean diameter D, aspect ratios (T/W, T/L and W/L) and the sphericity index Φ [25] of every seed:
Specific mass mD was determined for each seed [26]:
In nurseries, seeds are usually sorted using pneumatic separators and mesh sieves. Therefore, seeds of each fir species were divided into three potential fractions, roughly equal in size, based on their terminal velocity and thickness: fraction I—seeds with the lowest terminal velocity or thinnest seeds, fraction II—seeds with average terminal velocity or average thickness, fraction III—seeds with the highest terminal velocity or thickest seeds. Terminal velocity was measured to the nearest 5 m s−1, and seed thickness was measured to the nearest 0.1 mm.
2.3. Statistical Analysis
The measured values of the physical properties of seeds and the calculated indicators were analyzed statistically using Statistica PL v. 13.3 software (StatSoft Poland Ltd., Cracow, Poland) at a significance level of α = 0.05. The differences between the measured values and calculated indicators were determined by one-way analysis of variance (ANOVA). If the null hypothesis postulating that the mean values of a given parameter or indicator are equal was rejected, the significance of differences was estimated using Duncan’s test and homogenous groups were identified. The correlations between the physical properties of seeds were evaluated based on Pearson’s correlation coefficients [27].
3. Results and Discussion
3.1. Experimental Material
The accuracy of determining the mean value of a given physical parameter can be inferred from the standard error of the estimate based on the sample size, standard deviation of the analyzed trait and Student t-values at the adopted significance level. In this study, each sample comprised 101 to 118 seeds; therefore, the standard error of the estimate of the mean values of the physical properties of fir seeds did not exceed: 0.2 m s−1 for terminal velocity, 0.1 mm for seed thickness, 0.2 mm for seed width, 0.3 mm for seed length, 1° for the angle of external friction, and 0.5 mm (balsam fir) to 2.5 mm (Forrest’s fir) for seed mass.
The physical properties of the analyzed seeds are presented in Table 1. The average terminal velocity ranged from 4.8 m s−1 (balsam fir) to 7.1 m s−1 (silver fir). The following fir species formed homogeneous groups in terms of terminal velocity: (1) balsam fir and subalpine fir, (2) grand fir, Japanese fir and Sierra white fir, (3) corkbark fir and Sierra white fir, (4) corkbark fir, Forrest’s fir and Korean fir. The average terminal velocity of silver fir seeds was comparable to that presented by Kaliniewicz et al. [28], and somewhat higher than that determined by Tylek [29,30] in seeds from southern Poland.

Table 1.
Mean values (± standard deviation) of the physical properties of seeds of selected fir species. Superscript letters denote significant differences between properties at p < 0.05 (Duncan’s test).
The average seed thickness ranged from 1.76 mm (Korean fir) to 3.22 mm (silver fir). In turn, the average seed width ranged from 3.29 mm (balsam fir) to 5.57 mm (silver fir), and the average seed length—from 5.44 mm (balsam fir) to 11.06 mm (noble fir). Seeds characterized by similar values of the three basic dimensions were noted only in grand fir and Sierra white fir. The seeds of silver fir were somewhat larger than those analyzed by Tracz and Barzdajn [31], Tylek [30] and Kaliniewicz et al. [28], but their dimensions were within the range of average values noted in Poland [16]. The evaluated seeds were similar to selected batches of Normann fir seeds in terms of length [14,32], and they were similar to pindrow fir seeds in terms of length and width [33]. The analyzed Korean fir seeds were approximately 10% larger than those studied by Song et al. [34].
Relatively minor variation was noted in the values of the angle of external friction which ranged from 26° (corkbark fir) to 33° (balsam fir). Six fir species formed a homogeneous group in terms of the angle of external friction: Forrest’s fir, Korean fir, noble fir, Sierra white fir, silver fir and white fir. Balsam fir seeds differed most considerably from the remaining fir species in this respect.
In the studied fir species, seed mass (Figure 2, Table 1) ranged from 2.8 mg (balsam fir) to 78.6 mg (silver fir), and average seed mass ranged from 7.9 to 48.3 mg. The following fir species formed homogeneous pairs in terms of seed mass: (1) Korean fir and subalpine fir, (2) grand fir and Sierra white fir, (3) Forrest’s fir and noble fir. According to the literature [16,20,35,36,37], seed mass is largely influenced by environmental conditions, genetic traits, tree age and, above all, geographic location. If the above factors are not taken into account, the average mass of silver fir seeds is somewhat lower than that reported by Tylek [38], Balian [20] and Gradečki-Poštenjak and Ćelepirović [37], and similar to that noted by Skrzyszewska and Chłanda [13] and Kaliniewicz et al. [28]. The average mass of Sierra white fir and subalpine fir seeds was somewhat lower (by approx. 17% and 3%, respectively), whereas the average mass of grand fir seeds was higher (by approx. 32%) than that observed by Veech et al. [39] in the corresponding species.
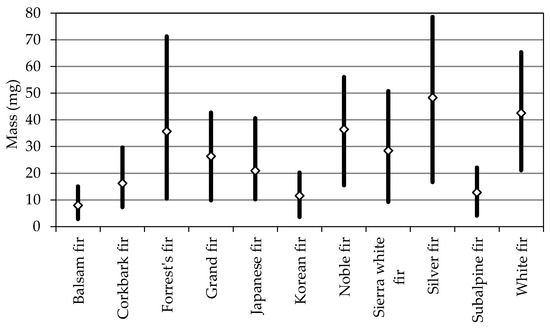
Figure 2.
Range of variation in seed mass in selected fir species.
Silver fir was characterized by the largest seeds (geometric mean diameter—5.77 mm), and balsam fir produced the smallest seeds (geometric mean diameter—3.16 mm) (Table 2). No significant differences in the values of the geometric mean diameter were noted between the following species: (1) balsam fir and Korean fir, (2) corkbark fir and subalpine fir, (3) grand fir and Sierra white fir, (4) Forrest’s fir and white fir.
Table 2.
Mean values (± standard deviation) of the calculated indicators of seeds of selected fir species. Superscript letters denote significant differences between properties at p < 0.05 (Duncan’s test).
The average aspect ratios were determined in the following range of values: T/W—from od 43.76% (Forrest’s fir) to 59.83% (noble fir); T/L—from 20.78% (Japanese fir) to 32.80% (balsam fir); W/L—from 40.11% (noble fir) to 60.58% (balsam fir). Aspect ratios were similar in the seeds of grand fir, Sierra white fir and white fir, i.e., species that belong to the section Grandis [2].
The highest values of the sphericity index were determined in balsam fir (58.20%), and the lowest values—in Japanese fir (43.86%). Eight homogeneous groups were identified in terms of the sphericity index, and the following species formed common groups: (1) grand fir, Sierra white fir and white fir, (2) silver fir and subalpine fir.
The average specific mass of fir seeds varied widely from 2.48 g m−1 (balsam fir) to 8.29 g m−1 (silver fir). The above indicates that balsam fir and silver fir seeds were characterized by the largest and smallest proportions of the seed coat, respectively, in seed mass, and that they contained empty spaces not filled with parenchymal tissue. The following species formed homogeneous pairs in terms of average seed mass: (1) Korean fir and subalpine fir, (2) silver fir and white fir.
Species pairs characterized by significant similarities in all physical parameters were not identified. The seeds of grand fir and Sierra white fir were most similar, and they did not differ significantly in the values of terminal velocity, basic dimensions, mass, geometric mean diameter, aspect ratios or sphericity index. In general, balsam fir seeds were most different, whereas Sierra white fir seeds were most similar to the seeds of the remaining fir species. Therefore, the seeds of Sierra white fir can be regarded as representative of the pine family and used to differentiate between the seeds of different fir species. The W/L aspect ratio (four homogeneous groups) was the least differentiating trait, whereas specific mass (nine homogeneous groups) was the most differentiating attribute in the analyzed fir species.
3.2. Correlations Between Seed Parameters
In further analysis, the measured parameters of fir seeds were pooled into a single experimental group. An analysis of the linear correlations between the basic physical properties of seeds (Table 3) revealed that seed mass, geometric mean diameter and specific mass were most correlated, whereas the angle of external friction was least correlated with the remaining parameters. The strongest correlation was noted between seed mass and specific mass (0.98), whereas the weakest correlation was observed between terminal velocity and the sphericity index (0.01). Fifty-five out of the 66 compared pairs of traits were significantly correlated at a significance level of 0.05. The correlation coefficient was practically significant (minimum 0.4) in 28 cases. The dimensions and mass of the analyzed fir seeds were more strongly correlated than in Turkish fir seeds [14], Cilician fir and Nordmann fir seeds [32] and silver fir seeds [28].
Table 3.
Coefficients of linear correlation between the physical properties (indicators) of fir seeds.
3.3. Seed Separation
According to many authors [2,13,33,37,40,41,42], fir seeds differ in germination capacity that can range from 0 to approximately 90% in freshly harvested seeds. The above can be attributed to the fact fir seeds are characterized by a high proportion of empty seeds [2,13,20,37,43,44], which can reach 70% in some cases. Empty seeds are difficult to separate because similarly to filled seeds, they contain resin globules whose specific gravity is similar to that of filled seeds. The ranges of seed mass values overlap in empty and filled seeds, but empty and filled seeds differ in average mass, and this trait can be potentially used in the separation of fir seeds [13,20,30]. The mass of fir seeds was most significantly influenced by seed thickness and seed length (Table 3). Therefore, the above parameters should be regarded as the primary distinguishing features in seed separation processes, and fir seeds should be sorted with the use of mesh sieves with longitudinal openings or with a seed grader. According to the authors of this study, a seed grader is less effective because indented pockets on the surface of the cylinder are more suitable for separating elliptical seeds [22] rather than triangular seeds. Seed thickness should be regarded as a distinguishing feature because this parameter was highly correlated with specific mass (coefficient of determination of 0.62). Sorting operations based on seed thickness will produce seed fractions with different content of parenchymal tissue. The resulting fractions should be sown separately to improve germination rates and germination efficiency.
A detailed analysis of the linear correlations between seed mass and the remaining physical attributes of seeds (Table 4) revealed that seeds of selected fir species can also be sorted with a pneumatic separator. The above applies particularly to the seeds of balsam fir, Forrest’s fir, grand fir, noble fir, Sierra white fir and subalpine fir. According to Załęski [16] and Tylek [29,30,38,45], terminal velocity should be the primary distinguishing trait in the process of separating silver fir seeds.
Table 4.
Coefficients of linear correlation between seed mass and the remaining physical properties of fir seeds.
The results of the analysis (Table 5) indicate that fir seeds can also be effectively separated based on their terminal velocity. In most cases, the resulting seed fractions were characterized by uniform seed mass. Before sorting, the coefficient of variation of seed mass ranged from around 22% (white fir) to around 37% (Forrest’s fir), and it decreased after sorting, particularly in fractions II and III. Fraction III seeds (8 out of 11 cases) and fraction II seeds (3 out of 11 cases) were least varied in terms of mass. The coefficient of variation of seed mass in each separated fraction differed across fir species from around 12% (fraction III, noble fir) to around 36% (fraction I, Sierra white fir). Seed mass was the most reliable separation trait in noble fir seeds in fraction III (change of 54.4%) and the least reliable trait in Sierra white fir seeds in fraction I (change of 36.2% relative to unsorted material). The average increase in the homogeneity of seeds separated into three fractions based on terminal velocity ranged from approximately 4.1% (Sierra white fir) to approximately 32.5% (noble fir).
Table 5.
Coefficient of variation of seed mass in three seed fractions separated based on terminal velocity.
In most cases (excluding three cases in fraction I), seed thickness was also a reliable parameter for sorting seeds into fractions with similar mass (Table 6). The coefficient of variation of seed thickness differed across fir species from around 16% (fraction III, silver fir) to around 35% (fraction I, Forrest’s fir), and silver fir seeds in fraction III were characterized by the most uniform thickness (change of 40% relative to unsorted material). The average increase in the homogeneity of seeds separated into three fractions based on seed thickness ranged from around 2.6% (grand fir) to around 24.7% (Japanese fir).
Table 6.
Coefficient of variation of seed mass in three seed fractions separated based on thickness.
The homogeneity of fir seeds separated into three fractions based on terminal velocity or seed thickness indicates that a pneumatic separator should be potentially used for sorting balsam fir, corkbark fir, Forrest’s fir, grand fir, noble fir and white fir seeds, whereas a mesh sieve with longitudinal openings is most suitable for sorting Japanese fir, Korean fir, Sierra white fir, silver fir and subalpine fir seeds. A mesh sieve appears to be the preferred solution for sorting fir seeds because the resulting fractions do not have to be dewinged before storage (seed wings do not disrupt the separation process), whereas dewinging operations increase the risk of damage to resin ducts and make seeds more susceptible to infection [38]. It should also be noted that the elimination of resin globules, for example during rapid dewinging, leads to a rapid deterioration in seed quality [2,9,46].
The obtained seed fractions should be analyzed for germination capacity and sown in the most appropriate locations. Seeds of fractions I and II, with the potentially lowest germination capacity, can be sown in rows or broadcast in conventional nurseries (by choosing the most appropriate sowing rate), whereas fraction III seeds can be sown individually in beds or containers.
4. Conclusions
Balsam fir produced the smallest seeds, and silver fir produced the largest seeds.
Sierra white fir seeds were most similar, and balsam fir seeds differed most considerably from the remaining fir species.
Specific mass (nine homogeneous groups) was the most differentiating attribute, whereas the W/L aspect ratio (four homogeneous groups) was the least differentiating trait in the analyzed fir species.
Fir seeds should be sorted primarily with the use of mesh sieves with longitudinal openings to obtain potential fractions with similar seed mass.
The seeds of balsam fir, corkbark fir, Forrest’s fir, grand fir, noble fir and white fir can also be sorted with pneumatic separators.
Author Contributions
Conceptualization, Z.K. (Zdzisław Kaliniewicz); methodology, Z.K. (Zdzisław Kaliniewicz) and Z.Ż.; formal analysis, Z.K. (Zdzisław Kaliniewicz) and K.J.; investigation, Z.K. (Zdzisław Kaliniewicz); resources, Z.K. (Zdzisław Kaliniewicz); data curation, Z.Ż.; writing—original draft preparation, Z.K. (Zdzisław Kaliniewicz), Z.K. (Zbigniew Krzysiak), P.M. and A.A.; writing—review and editing, Z.K. (Zdzisław Kaliniewicz); visualization, Z.K. (Zdzisław Kaliniewicz); supervision, Z.K. (Zdzisław Kaliniewicz); project administration, Z.K. (Zdzisław Kaliniewicz); funding acquisition, Z.K. (Zdzisław Kaliniewicz).
Funding
This research received no external funding.
Acknowledgments
The authors would like to thank Aleksandra Poprawska, MA for English-language editing and proofreading this manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Arista, M.; Herrera, J.; Talavera, S. Abies pinsapo Boiss.: A protected species in a protected area. Bocconea 1997, 7, 427–436. [Google Scholar]
- Edwards, D.G.W. Abies P. Mill. In Woody Plant Seed Manual—Part II; Agriculture Handbook 727; United States Department of Agriculture, Forest Service: Washington, DC, USA, 2008; pp. 149–198. [Google Scholar]
- Linares, J.C. Biogeography and evolution of Abies (Pinaceae) in the Mediterranean Basin: The roles of long-term climatic change and glacial refugia. J. Biogeogr. 2011, 38, 619–630. [Google Scholar] [CrossRef]
- Aguirre-Planter, É.; Jaramillo-Correa, J.P.; Gómez-Acevedo, S.; Khasa, D.P.; Bousquet, J.; Eguiarte, L.E. Phylogeny, diversification rates and species boundaries of Mesoamerican firs (Abies, Pinaceae) in a genus-wide context. Mol. Phylogenet. Evol. 2012, 62, 263–274. [Google Scholar] [CrossRef]
- Ghimire, B.; Lee, C.; Yang, J.; Heo, K. Comparative leaf anatomy of some species of Abies and Picea (Pinaceae). Acta Bot. Brasilica 2015, 29, 346–353. [Google Scholar] [CrossRef]
- Caudullo, G.; Tinner, W. Abies—Circum-Mediterranean firs in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publication Office of the European Union: Luxembourg, Luksembourg, 2016; p. e015be7+. [Google Scholar]
- Agrawal, T. Abies: A threatened genus. Br. J. Res. 2017, 4, 26. [Google Scholar] [CrossRef]
- Seneta, W.; Dolatowski, J. Dendrologia (Dendrology); Wydawnictwo Naukowe PWN: Warszawa, Poland, 2012; pp. 31–38. ISBN 9788301153694. (In Polish) [Google Scholar]
- Huth, F.; Wehnert, A.; Tiebel, K.; Wagner, S. Direct seeding of silver fir (Abies alba Mill.) to convert Norway spruce (Picea Abies L.) forests in Europe: A review. Forest Ecol. Manag. 2017, 403, 61–78. [Google Scholar] [CrossRef]
- Gosling, P. Raising Trees and Shrubs from Seed (Practice Guide (FCPS)); Forestry Commission: Edinburgh, UK, 2007; pp. 1–34. ISBN 9780855387365.
- Varsamis, G.K.; Takos, I.A.; Merou, T.P.; Galatsidas, S.A.; Panayiotis, D.D. Germination characteristics of Abies × borisii-regis. Seed Technol. 2014, 36, 51–59. [Google Scholar]
- Bhat, H.A.; Mughal, A.H.; Din Dar, M.U.; Mugloo, J.A. Cone, seed and germination characteristics in silver fir (Abies pindrow Spach) along the altitudinal gradient in western Himalayas. Int. J. Chem. Stud. 2018, 6, 2052–2055. [Google Scholar]
- Skrzyszewska, K.; Chłanda, J. A study on the variation of morphological characteristics of silver fir (Abies alba Mill.) seeds and their internal structure determined by X-ray radiography in the Beskid Sądecki and Beskid Niski mountain ranges of the Carpathians (Southern Poland). J. For. Sci. 2009, 55, 403–414. [Google Scholar] [CrossRef]
- Sevik, H.; Yahyaoglu, Z.; Turna, I. Determination of genetic variation between populations of Abies nordmanniana subsp. bornmulleriana Mattf according to some seed characteristics. In Genetic Diversity in Plants; Caliskan, M., Ed.; InTech: Rijeka, Croatia, 2012; pp. 231–248. ISBN 9789535101857. [Google Scholar]
- Sabor, J. Relation between the weight and the germination capacity of seed of silver fir. Sylwan 1984, 4, 59–69. (In Polish) [Google Scholar]
- Załęski, A. Nasiennictwo Leśnych Drzew i Krzewów Iglastych (Management of Coniferous Forest Trees and Shrubs for Seed Production); Oficyna Edytorska “Wydawnictwo Świat”: Warszawa, Poland, 1995; pp. 1–178. ISBN 8385597271. (In Polish) [Google Scholar]
- Khan, M.L. Effects of seed mass on seedling success in Artocarpus heterophyllus L., a tropical tree species of north-east India. Acta Oecol. 2004, 25, 103–110. [Google Scholar] [CrossRef]
- Upadhaya, K.; Pandey, H.N.; Law, P.S. The effect of seed mass on germination, seedling survival and growth in Prunus jenkinsii Hook.f. & Thoms. Turk. J. Bot. 2007, 31, 31–36. [Google Scholar] [CrossRef]
- Norden, N.; Daws, M.I.; Antoine, C.; Gonzalez, M.A.; Garwood, N.C.; Chave, J. The relationship between seed mass and mean time to germination for 1037 tree species across five tropical forests. Funct. Ecol. 2009, 23, 203–210. [Google Scholar] [CrossRef]
- Ballian, D. Genetic overload of silver fir (Abies alba Mill.) from five populations from central Bosnia and Herzegovina. Folia For. Pol. Ser. A For. 2013, 55, 49–57. [Google Scholar] [CrossRef]
- Chaisurisri, K.; Edwards, D.G.W.; El-Kassaby, Y.A. Effects of seed size on seedling attributes in Sitka spruce. New For. 1994, 8, 81–87. [Google Scholar]
- Grochowicz, J. Maszyny do Czyszczenia i Sortowania Nasion (Seed Cleaning and Sorting Machines); Wydawnictwo Akademii Rolniczej: Lublin, Poland, 1994; pp. 25–33. ISBN 839016129X. (In Polish) [Google Scholar]
- Kaliniewicz, Z.; Poznański, A. Variability and correlation of selected physical attributes of small-leaved lime (Tilia cordata Mill.) seeds. Sylwan 2013, 157, 39–46. (In Polish) [Google Scholar]
- Kaliniewicz, Z.; Markowski, P.; Anders, A.; Jadwisieńczak, B.; Rawa, T.; Szczechowicz, D. Basic physical properties of Norway spruce (Picea abies (L.) Karst.) seed. Tech. Sci. 2016, 19, 103–115. [Google Scholar]
- Mohsenin, N.N. Physical Properties of Plant and Animal Materials; Gordon and Breach Science Public: New York, NY, USA, 1986; pp. 1–891. ISBN 9780677213705. [Google Scholar]
- Kaliniewicz, Z. Analysis of frictional properties of cereal seed. Afr. J. Agricult. Res. 2013, 8, 5611–5621. [Google Scholar] [CrossRef]
- Rabiej, M. Statystyka z Programem Statistica (Statisctics in Statistica Software); Helion: Gliwice, Poland, 2012; pp. 1–344. ISBN 9788324641109. (In Polish) [Google Scholar]
- Kaliniewicz, Z.; Mańkowski, S.; Tylek, P.; Krzysiak, Z.; Peda, W. Correlations between the physical properties of silver fir seeds. Acta Agroph. 2018, 25, 197–212. (In Polish) [Google Scholar] [CrossRef]
- Tylek, P. Problems of pneumatic selection of forest tree seeds. Sylwan 1999, 12, 65–72. (In Polish) [Google Scholar]
- Tylek, P. Efficiency of some basic criteria of silver fir seeds separation. Acta Agroph. 2003, 2, 857–866. (In Polish) [Google Scholar]
- Tracz, M.; Barzdajn, W. The morphological traits of cones and seeds of Abies alba in the Middle Sudeten. Dendrobiology 2007, 58, 59–65. [Google Scholar]
- Velioğlu, E.; Tayanç, Y.; Çengel, B.; Kandemir, G. Genetic variability of seed characteristics of Abies populations from Turkey. J. Kast. Forf. 2012, 3, 27–35. [Google Scholar]
- Masoodi, H.U.R.; Thapliyal, M.; Singh, V.R.R. Studies on the variation in germination and seedling growth of Abies pindrow Spach. (Royle) in Garhwal region of Uttarakhand, India. J. Appl. Nat. Sci. 2014, 6, 711–715. [Google Scholar] [CrossRef]
- Song, J.-H.; Jang, K.-H.; Hur, S.-D. Variation of seed and germination characteristics of natural populations of Abies koreana Wilson, a Korean endemic species. J. Korean For. Soc. 2010, 99, 849–854. [Google Scholar]
- Oleksyn, J.; Reich, P.B.; Tjoelker, M.G.; Chalupka, W. Biogeographic differences in shoot elongation pattern among European Scots pine populations. For. Ecol. Manag. 2001, 148, 207–220. [Google Scholar] [CrossRef]
- Moles, A.T.; Westoby, M. Latitude, seed predation and seed mass. J. Biogeogr. 2003, 30, 105–128. [Google Scholar] [CrossRef]
- Gradečki-Poštenjak, M.; Ćelepirović, N. The influence of crown defoliation on the variability of some physiological and morphological properties of silver fir (Abies alba) seeds in the seed zone of Dinaric beech-fir forests in Croatia. Period. Biol. 2015, 117, 479–492. [Google Scholar] [CrossRef]
- Tylek, P. Agrophysical characteristics of silver fir seeds after pneumatic separation. Acta Sci. Pol. Silv. Colendar. Rat. Ind. Lignar. 2005, 4, 97–105. (In Polish) [Google Scholar]
- Veech, J.A.; Charlet, D.A.; Jenkins, S.H. Interspecific variation in seed mass and the co-existence of conifer species: A null model test. Evol. Ecol. Res. 2000, 2, 353–363. [Google Scholar]
- Arista, M.; Talavera, S. Density effect on the fruit-set, seed crop viability and seedling vigour of Abies pinsapo. Ann. Bot. 1996, 77, 187–192. [Google Scholar] [CrossRef]
- Andersen, U.S.; Cordova, J.P.; Nielsen, U.B.; Kollmann, J. Provenance variation in germination and seedling growth of Abies guatemalensis Rehder. Forest Ecol. Manag. 2008, 255, 1831–1840. [Google Scholar] [CrossRef]
- Kurt, Y.; Frampton, J.; Isik, F.; Landgren, C.; Chastagner, G. Variation in needle and cone characteristics and seed germination ability of Abies bornmuelleriana and Abies equi-trojani populations from Turkey. Turk. J. Agric. For. 2016, 40, 169–176. [Google Scholar] [CrossRef]
- Nielsen, U.B.; Hansen, O.K. Response to selfing in seed set, seedling establishment and nursery growth based on controlled crosses of Abies nordmanniana clones. Silvae Genet. 2009, 59, 90–98. [Google Scholar] [CrossRef]
- McCartan, S.A.; Jinks, R.L. Upgrading seed lots of European silver fir (Abies alba Mill.) using imbibition-drying-separation. Tree Planters’ Notes 2015, 58, 21–27. [Google Scholar]
- Tylek, P. Analysis of aerodynamic properties of common fir and common beech. Inżynieria Rolnicza 2011, 6, 247–253. (In Polish) [Google Scholar]
- Aniszewska, M.; Gendek, A.; Śliwińka, J. Variability of silver fir (Abies alba Mill.) cones—Variability structure of scale surface area. For. Res. Pap. 2017, 78, 5–13. [Google Scholar] [CrossRef]
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).