Retention Forestry Supports Bird Diversity in Managed, Temperate Hardwood Floodplain Forests
by
 , , ,
, , ,
Ivo Machar
1,* ,
,
Martin Schlossarek
1,
Vilem Pechanec
2,
Lubos Uradnicek
3,
Ludek Praus
4 and
Ahmet Sıvacıoğlu
5 1
Department of Development and Environmental Studies, Faculty of Science, Palacky University Olomouc, 771 47 Olomouc, Czech Republic
2
Department of Geoinformatics, Faculty of Science, Palacky University Olomouc, 771 47 Olomouc, Czech Republic
3
Dendrology and Geobiocoenology, Department of Forest Botany, Faculty of Forestry and Wood Technology, Mendel University in Brno, 613 00 Brno, Czech Republic
4
Department of Wood Science, Faculty of Forestry and Wood Technology, Mendel University in Brno, 613 00 Brno, Czech Republic
5
Faculty of Forestry, Kastamonu University, 37000 Kastamonu, Turkey
*
Author to whom correspondence should be addressed.
Forests 2019, 10(4), 300; https://doi.org/10.3390/f10040300
Submission received: 11 February 2019
/
Revised: 17 March 2019
/
Accepted: 28 March 2019
/
Published: 1 April 2019
(This article belongs to the Special Issue Effects of Forest Management Practices on Forest Biodiversity)
Abstract
:The retention forestry approach is considered as one of the potentially effective tools for sustainable forest management for conservation of biodiversity in managed temperate and boreal forests. Retention of old-growth forest structures (e.g., very large old living trees) in forest stands during clear-cutting provides maintenance of key habitats for many old-growth forest interior-species. Most of ecological studies on green tree retention (GTR) consequences for biodiversity have been focused on birds. However, the long-term studies of GTR impacts on forest birds are very poor. In this paper, we focused on assessment of the long-term consequences of leaving legacy oak trees on the cut areas for bird diversity 18–22 years after clear-cutting in managed temperate European hardwood floodplain forests. Results based on bird counting using mapping of bird nesting territories revealed a key importance of legacy oak trees for maintaining bird diversity in the study area. These results are widely applicable for managed temperate hardwood forests with serious dominance of oak (Quercus sp.) in forest stands. Legacy oak trees in this habitat type are keystone structures for bird diversity. Retention approach focused on these trees is potentially an important conservation tool for preserving forest bird diversity and other associated species in temperate hardwood forests managed by clear-cutting.
1. Introduction
Forest management in temperate hardwood forests in North America and Europe uses clear-cutting as one of its main logging techniques [1]. Old-growth forest stands, important for biodiversity, are very rare in commercial hardwood forests that are managed for timber by clear-cutting. Efforts towards the maintenance of biodiversity in these forests are based on a general knowledge of the consequences of natural forest disturbances. The main difference between clear-cutting and natural disturbances in forests is that the “biological legacies” left by previous forest stands (such as the amount of coarse woody debris, dead standing trees, and living trees of various ages and sizes) still remain after disturbance. Clear-cutting usually removes almost all live and dead wood from logged areas and no important biological legacy remains. The biological structural complexity that results from natural disturbances is very important for the functioning of the forest ecosystem and the maintenance of forest biodiversity [2]. Clear-cutting could more closely resemble natural disturbances with the deliberate retention of the structural features of old-growth forests in logged areas [3]. In Northern America [4] and Europe [5] these retaining forestry practices aimed at maintaining biodiversity have spread over the last three decades. Various benefits for biodiversity to be gained from retention forestry have been studied, such as the production of the ectomycorrhizal fungus sporocarp [6], forest ground vegetation [7], invertebrates [8], and small terrestrial mammals [9].
One expanding form of retention forestry is green-tree retention (GTR), which is based on not cutting down live old trees in logged areas. This preserves forest biodiversity while retaining economic viability [10]. GTR reduces the negative impact of clear-cutting management on mature forest-associated species. Nearly 29% of the ecological studies on the consequences of GTR for biodiversity have focused on birds, and the short-term benefits of GTR for bird diversity are very clear [11]. The study [12], based on research into songbird responses to GTR in aspen stands, highlighted the importance of considering a forest stand’s age when assessing the impact of GTR on songbirds in harvest units. Forest bird communities in a landscape mosaic of variable forest stands, including old residual trees, could be very indicative for forest management [13,14]. The main role of GTR for forest birds could be the creation of suitable stand structures—old-growth features in young forests, such as individual residual trees in post-cut forest areas [15]. When managed on a rotational basis [16], these residual green trees would extent the duration of suitable phases for birds in forest stands. In general, GTR supports the maintenance of bird species associated with old-growth forests in clear-cut areas [17].
Individual old-growth trees that have not been cut down during harvests (or have survived stand-replacing natural disturbances) are referred to as “legacy trees” [18]. In order to distinguish very large legacy trees from other residual trees, the authors of study [19] defined legacy trees as having achieved near-maximum size and age, being significantly larger and older than the average trees in the landscape.
In this paper we use this definition of legacy trees because the individual trees retained during clear-cutting in our study are significantly larger and older than the average old trees in mature forest stands. This study focuses on the consequences for bird diversity of retaining legacy oak trees 18–22 years after clear-cutting and has taken place in managed, temperate, European hardwood floodplain forests. We tested the hypothesis that “individual legacy trees in managed hardwood floodplain forests can be a feature with important habitat value for forest bird diversity”. The hypothesis was tested by comparing nesting bird diversity in study plots in clear-cut areas; those with and those without retained individual legacy trees. The results are discussed in the frame of sustainable forest management using the retention forestry approach.
2. Materials and Methods
2.1. Study Area, Forest Management Practice, and Study Plots
The study area is covered by hardwood floodplain forest ecosystems of the European temperate zone along the Morava River in the Czech Republic [20]. The study area is protected as a buffer conservation zone, known as Litovelske Pomoravi Protected Landscape Area (LPPLA), and is included in the Natura 2000 European network [21]. Within the framework of the Natura 2000 habitat classification these forest ecosystems are considered as habitat “91F0 Hardwood forests of lowland rivers” [22]. The dominant tree species in the LPPLA habitat is Pedunculate Oak (Quercus robur L.), with an admixture of European Ash (Fraxinus excelsior L.), Small-Leaved Lime (Tilia cordata Mill.), Field Maple (Acer campestre L.), Sycamore Maple (Acer pseudoplatanus L.), Norway Maple (Acer platanoides L.), Hornbeam (Carpinus betulus L.), European White Elm (Ulmus laevis Pallas), and Bird Cherry (Prunus padus L.).
Since 1992 [23], following the Natura 2000 recommendations, forest management practice in the buffer zone LPPLA has been aimed at maintaining Pedunculate Oak as the main tree species, as in other hardwood floodplain forest habitats. Pedunculate Oak is a light-demanding tree species and thus there is clear-cutting forest management in progress. Forest stands are managed by clear-cutting with the retention of very large individual Pedunculate Oak trees in the clear-cut areas. The Management Plan for the buffer zone LPPLA prescribes retaining a minimum of 5 very large oak trees per hectare in cut areas, in order for biodiversity conservation. In some forest stands more very large oaks are retained than the prescribed minimum—see our study plots nos. 4–6 (Table 1). In fact, these very large remaining oak trees are considered to be typical individual legacy trees because they are very old (180–250 years)—considerably larger and older than the average trees in mature forest stands (110–120 years). The retention of very large oak trees has been applied in the study area since the establishment of planning forest management at the end of the 18th century as a basic principle of standard-with-coppice management [24]. In the past the reason for the retention of old oak standards in cut areas was economic (timber production) and currently these former oak standards are considered as individual legacy oak trees (ILOT) protected for biodiversity conservation [25].
In the study area, we established six study plots: three study plots without the presence of ILOT (study plots no. 1, 2, and 3) and three study plots with the presence of ILOT (study plots no. 4, 5, and 6). The size of study plots varies from 2.8 ha to 4.8 ha (see Table 1 in details). All these study plots are covered by young forest stands dominated by Pedunculate Oak which was established by artificial regeneration after clear-cutting. The age of young oak forest stands on all study plots varies between 15–22 years and the average stand height is 4 m above the ground. In the future another mixture of forest species typical for hardwood floodplain forest habitats (see above) will gradually appear via natural regeneration from surrounding mature forest stands. Clear-cut areas are fenced in order to prohibit ungulate browsing on young oak plants.
2.2. Bird Census in Study Plots
Nesting birds were counted in the breeding season from the last week in March to the first week of July in study plots no 1, 2, and 5 in 2006 and 2008, and in study plots no 3, 4, and 6 in 2007 and 2009. In each study plot, nesting birds were counted 10 times per breeding season in the early morning and twice in the evening (focused especially on Locustella fluviatilis and Luscinia megarhynchos) using the territory mapping method [26]. Birds were only included when they were spotted within 50 metres of the surveying researcher, in order to avoid mistakes based on different detectability of birds in hardwood floodplain forests [27]. We did not include in the field mapping those bird species that obviously have no nesting-relationship with forest habitats, and were only observed randomly in study plots. The field mapping of nesting territories was complemented with serious repeated visual inspections of ILOTs in all study plots, in order to determine birds nesting in holes. This was necessary for the precise identification of Leiopicus medius nests, as it is a very difficult species to count using territory mapping [28]. Additionally, in January/March 2006 and 2009 we surveyed the nests of Strix aluco in large holes in ILOTs. We found their location based on the calls of the owls in the study area on the previous night.
2.3. Data Analysis
The bird nesting community in the study area was subdivided into four guilds based on their nest-site locations in forest stands (ground-nesters, bush-nesters, canopy-nesters, and hole-nesters). Guild classification was carried out a priori [29] based on our previous ornithological experience in the study area and following relevant Czech ornithological literature [30]. We used the field data from the bird census (related to each of detected species) to calculate mean bird density in each of study plots for all years of field research (nesting pairs/10 ha). In order to analyse the predominant bird species, we calculated dominance values according to [31]. In order to compare the similarity and diversity of nesting bird communities, we calculated with the commonly used Jaccard similarity index [32].
The statistical analysis of data was based on the assumption that the density of recorded bird species in study plot areas with ILOT (i.e., Hrube les, Brezova, and Odchovna) would be different from the density in each study plot area without ILOT (i.e., Pnovice, Stepanov, and Horni les). To verify this assumption, the null hypothesis “The density of total bird species at study plot area with ILOT is the same as density of total bird species at study area without ILOT” was tested on 5% significance level against the alternative two-tailed hypothesis. The Mann-Whitney U test, also called Wilcoxon rank–sum test [33], was used for each pair of area with ILOT and area without ILOT. The test can detect differences in shape and spread as well as just differences in medians. As we did multiple (9) testing, the Holm–Bonferroni method of correction was used to adjust respective critical levels of p-values. The analysis was carried out using the Stata 12 program [34].
3. Results
A total of 39 bird species were identified as nesting bird species in all study plots (Table 1) during the research. The differences in the alpha-diversity of nesting bird communities in the study plots are evident between plots with and without the presence of ILOT. The largest difference (54%) has been found between plot 1 and 2 (16 nesting species, without ILOT) and plot 4 (35 nesting species with ILOT). The faunistic similarity of species composition among all study plots measured by the Jaccard index (Table 1) varies from 39% (between plots 1 and 6) to 67% (plots 5 and 6).
The density values calculated for the study plots indicate a similar pattern to alpha-diversity according to the presence/absence of ILOT. The lowest density of nesting birds was recorded in study plot 2 (54.1 pairs/10 ha), and the highest was in study plot 6 (85.4 pairs/10 ha)—see Table 1.
The predominant species (˃5%) in all study plots (Table 1) are common forest bush-nesters and canopy-nesters (Erithacus rubecula, Fringilla coelebs, Phylloscopus collybita, Prunella modularis, Turdus merula, and Sylvia atricapilla).
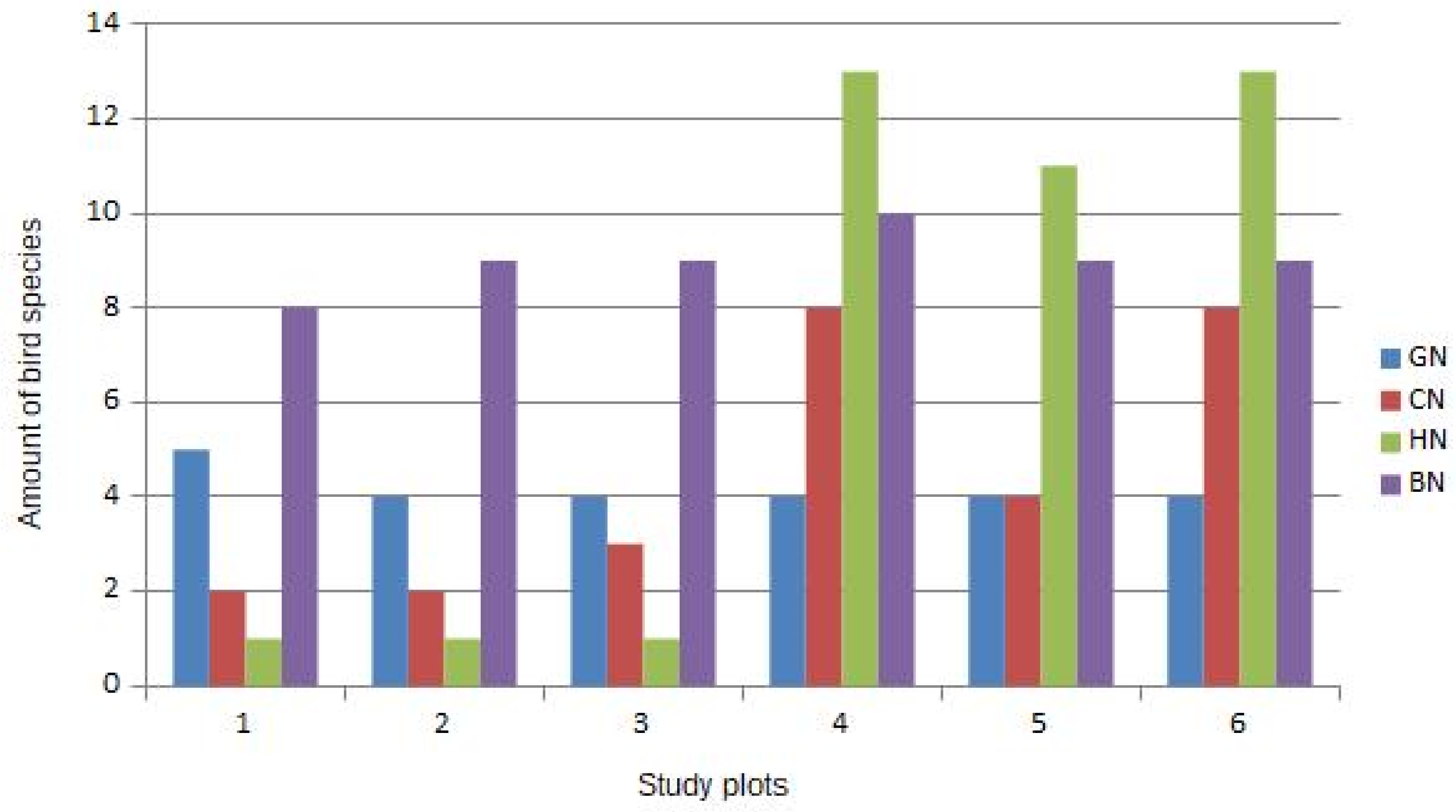
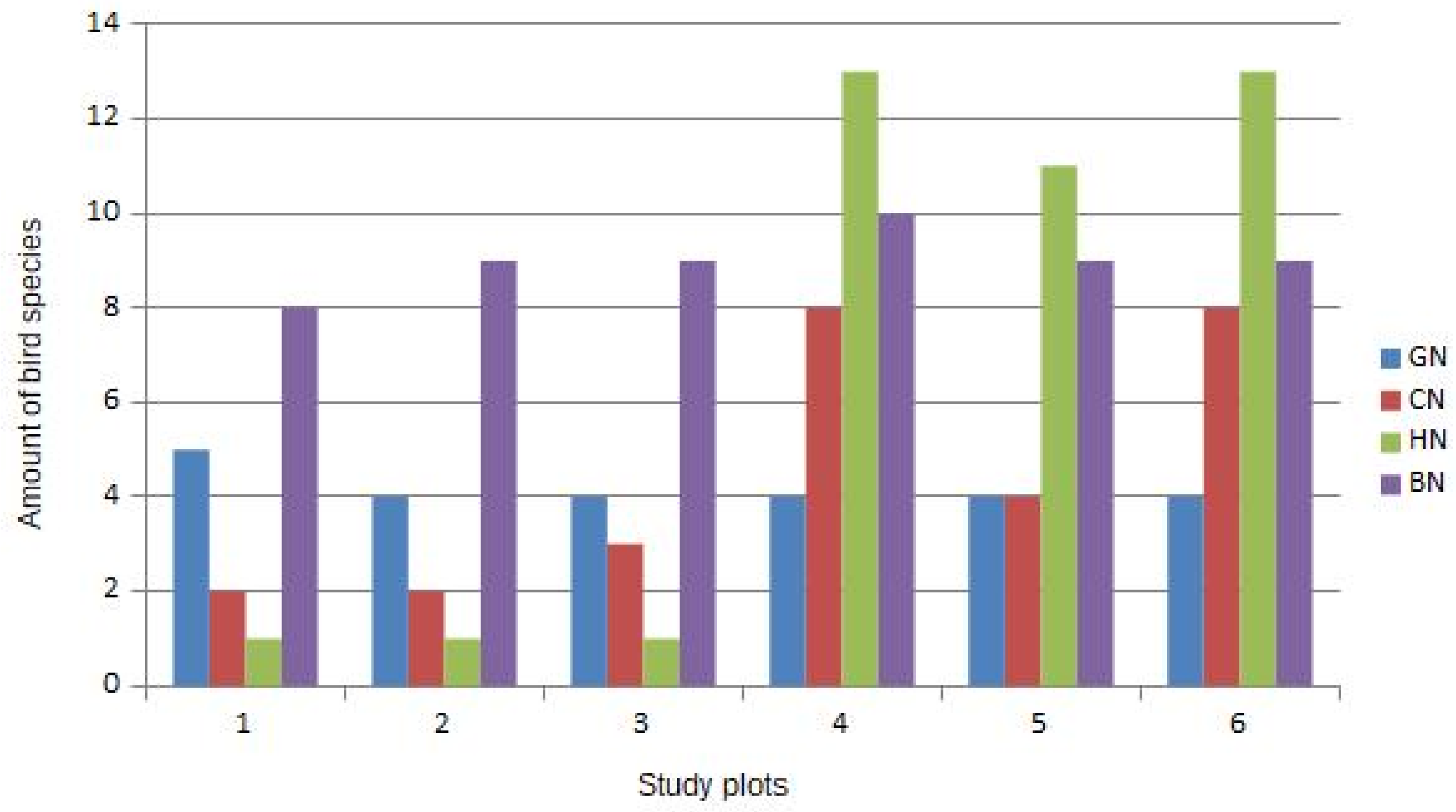
The guild structure in all study plots (Figure 1) revealed important information: In study plots with the presence of ILOT (nos. 4, 5, and 6) we can clearly see a high presence of hole-nesters (11–13 species). In study plots without ILOT (nos. 1, 2, and 3) the hole-nesters guild is almost totally absent, with the exception of Cyanistes caeruleus and Parus major. All species from the hole-nesters guild in the study plots are typical inhabitants of natural holes and small cavities in old forest trees. Thus, we can deduct from Figure 1 that the main reason for the high alfa-diversity in plots 4–6 is a guild of hole-nesters connected with the presence of ILOT.
The presence of ILOT has increased the average density as well as median of density of those 39 recorded bird species in individual study plot areas. The differences in measures of central tendency are related to lower proportions of species spotted in areas with and without ILOT. A total of 35 of 39 species (i.e., 89.7%) were spotted in Hrube les, 34 of 39 in Odchovna (87.2%) and 28 of 39 (71.8%) in Brezova. Proportions were much lower in study plot areas without ILOT: 17 of 39 (43.6%) in Horni les and 16 of 39 (41.0%) in Stepanov as well as in Pnovice. While standard deviations seem to be roughly same for both groups of study plot areas, adjusted Fisher–Pearson coefficients of skewness are higher for study plot areas without ILOT compared to study plot areas with ILOT. Basic descriptive statistics are summarized in Table 2 (N = 39 for each study plot).
We ran nine Mann-Whitney U tests for the difference in distributions of bird species densities. When interpreting results, we used the Holm–Bonferroni method of correction as a protection against type I errors. We found that the difference is statistically significant for 6 of 9 tests. We interpret these results as “another soft” evidence supporting our expectation that individual legacy oak trees have importance for maintaining bird diversity. Table 3 presents detailed results obtained by hypothesis testing.
4. Discussion
Our results from the study area Litovelske Pomoravi indicate that the ILOTs in managed temperate floodplain hardwood forests can be considered as keystone structures for the maintenance of forest bird diversity. The hypotheses tested, regarding the important habitat value of individual legacy trees for nesting birds, have been validated. In our study, the densities of nesting birds were significantly higher in study plots with ILOTs as retaining structures of the former old-growth stands. Not surprisingly, population density and numbers of bird species increased markedly with the stand age of temperate hardwood forests [35]. Findings from Litovelske Pomoravi are in accordance with the close relationship between temperate forest vegetation structure and bird diversity [36]. Biodiversity conservation in managed temperate forests requires a variety of strategies implemented at different spatial scales [37]. ILOTs, as old-growth forest structures, support the biodiversity of bird communities on a local scale. This statement is supported by study [38], which found that the high diversity among birds on a local scale was primarily connected with an increase in the proportion of old-growth hardwood forests. ILOTs retained on cuts (study plots 4–6 in this study) ensure that managed hardwood forests contribute to biodiversity conservation [39]. The importance of individual legacy oak trees for bird diversity in the hardwood forests of Litovelske Pomoravi can be probably explained by considering the extreme size and age of its ILOTs in accordance with study [40].
Generally, the intermediate-disturbance hypothesis (suggesting that species richness reaches a maximum at some intermediate level of disturbances) was confirmed for temperate hardwood floodplain forests (THFF) [41]. The clear-cutting system drastically disturbs forest ecosystems. Retention forestry practices based on GTR can be considered as possible means of achieving an intermediate level of disturbance from the effects of clear-cutting in floodplain forests, within the framework of biodiversity maintenance [42]—this is in accordance with results of our study.
We believe that GTR aimed at legacy trees can be a very important conservation tool for the sustainable management of hardwood floodplain forests [43]. We need further evaluation of the effectiveness of GTR in differently managed forest types/biomes in order to address the wider application of biodiversity conservation in sustainable forest management practice. We should be aware that the GTR benefits are specifically for different species—birds, ectomycorrhizal fungi, epiphytes, and wood-dependent beetles may be suitable bio-indicators for measuring the success of GTR in conservation effectiveness. Lifeboating of some species through GTR can cause ecological cascade effects, such as changes in invertebrate biomass due to GTR influencing the breeding success of passerine birds [44].
The ILOT in study plots 4-6 possess many tree cavities and basal hollows. Our results clearly indicated the importance of ILOT, especially for guild hole-nesters. The presence or absence of hole-nesters in study plots depends on the presence/absence of ILOT. This is not surprising as only ILOT are trees with cavities and holes in our study plots, and tree cavities are keystone habitat structures for forest biodiversity [45]. This finding is widely supported in studies of different types of temperate hardwood forests. Study [46] in young temperate hardwood forest stands with a predominance of Quercus petraea revealed a bird nesting community with 37% of hole-nesters in study plots with seven ILOTs and only 8% of hole-nesters in study plots without ILOTs. Hollows in legacy trees appear to be important nesting sites for Vaux´s swifts in California [47]. The absence of available hollows as nesting sites limits bird density [48].
In study plots 4 and 6 we found nests of Strix aluco, which is the biggest bird nesting species in our study. Similarly, old legacy trees are used by Strix occidentalis for nesting in Northern Americas’ managed forests [49]. The general consensus in literature is that old residual trees benefit songbirds, but there are more case studies from boreal forests [50] then from temperate ones. Study [51] from hemi-boreal forests in Sweden found the greater abundance of late-successional birds in cuts with higher retention levels, especially of deciduous and dead trees. That is very similar to our results from Litovelske Pomoravi, where we found a significantly higher abundance of birds in plots with residual ILOTs.
In our study canopy-dwelling birds also benefited from the presence of ILOTs (see Figure 1: study plots 4–6 in comparison with plots 1–3). ILOTs offer big tree crowns as nesting sites for canopy-nesters. This is reflected in many studies, showing the well-documented relationship between percentage of canopy cover and bird diversity [52,53]. Despite that, a study by Pierson et al. [54] found an inconsistent relationship between overstory vegetation cover and bird species diversity among temperate woodlands in Australia. This suggests that overstory cover will need to be thoroughly evaluated as a substitute for bird diversity in distinct regions.
The diversity of nesting bird communities in the study plots without ILOT (plots 1, 2, and 3) basically consists of open and semi-open habitat bird species (Table 1). The prevailing habitat in all of study plots was young (approximately 20 years old) deciduous forest stands in the form of bush. This habitat characteristic probably determined the diversity of bird species nesting in these plots. This supported the structure of bird nesting guilds analysis—the majority of bird species in all of the plots without ILOT are bush-nesters (Figure 1).
There is still a gap between current conservation practices and the actual habitat requirements of forest bird species which are of conservation concern [55]. There are many issues related to GTR that need future research. If old legacy trees are rare in some regions, those which do exist may be easily located by predators and this could cause their bird nesting populations to decline. These trees can function as ecological traps [56]. Titler and Hannon [57] did not find increased predation in this respect, but this issue appear to be an interesting future research topic related to the importance of individual legacy trees for biodiversity.
The ILOTs in our study plots in Litovelske Pomoravi are dispersed in cuts. Study [58] evaluated aggregate GTR in boreal forest which were more beneficial for forest birds than dispersed retention harvest. Although aggregate retention patches are not equivalent to larger areas of intact forests, aggregate GTR are very important breeding habitats for forest birds. Lindenmayer et al [59] studied whether dispersed or aggregated retention is more effective for bird conservation in mountain ash forests in South-Eastern Australia. They identified a significant gradient in bird alfa-diversity across treatments with the lowest values in clear-felled areas and the highest values in the unlogged controls. This suggests a need for future research to focus on the evaluation of aggregated retaining old-growth patches for the biodiversity of temperate hardwood forests as potential conservation tools in managed forest stands.
5. Conclusions
This study highlighted the importance of retain forestry in managed temperate hardwood floodplain forests. Individual legacy trees of pedunculate oak retaining as old-growth structures in clear-cutting areas seriously support bird diversity. Retaining and conservation of these legacy trees in managed forests is a very valuable part of sustainable forest management practice. Results of this study can be applied widely in temperate hardwood forests managed by timber production: Legacy trees need to be mapped in the field and retain during clear-cutting as keystone forest structure for biodiversity conservation.
Author Contributions
I.M. and A.S. surveyed birds in the field; M.S. was responsible for statistical tests of hypotheses; L.U., L.P., V.P., and A.S. prepared a research design and validated data; I.M. wrote an original draft and served as a corresponding author; V.P. was responsible for project administration and funding acquisition.
Funding
This study was supported by the grant: Significant Trees-Living Symbols of National and Cultural Identity, No. DG18P020VV027, funded by the Ministry of Culture of the Czech Republic from NAKI II (Programme to Support Applied Research and Experimental Development of National and Cultural Identity).
Acknowledgments
The authors thank David M. Richardson for correction of the English language. The authors are also grateful to the Academic Editor and Reviewers for their valuable comments on the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Keenan, R.J.; Kimmins, J.P. The ecological effects of clear-cutting. Environ. Rev. 1993, 1, 121–144. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Franklin, J.F. Conserving Forest Biodiversity: A Comprehensive Multiscaled Approach; Island Press: Washington, DC, USA, 2002. [Google Scholar]
- Beese, W.J.; Dunsworth, B.G.; Zielke, K.; Bancroft, B. Maintaining attributes of old-growth forests in coastal B.C. through variable retention. For. Chron. 2003, 79, 570–578. [Google Scholar] [CrossRef] [Green Version]
- Aubry, K.B.; Halpern, C.B.; Maguire, D.A. Ecological effects of variable-retention harvests in the northwestern United States: The DEMO study. For. Snow Landsc. Res. 2004, 78, 119–137. [Google Scholar]
- Vanha-Majamaa, I.; Jalonen, J. Green tree retention in Fennoscandian forestry. Scand. J. For. Res. 2001, 3, 79–90. [Google Scholar] [CrossRef]
- Luoma, D.L.; Eberhart, J.L.; Molina, R.; Amaranthus, M.P. Response of ectomycorrhizal fungus sporocarp production to varying levels and patterns of green-tree retention. For. Ecol. Manag. 2004, 202, 337–354. [Google Scholar] [CrossRef]
- Nelson, C.R.; Halpern, C.B. Edge-related responses of understory plants to aggregated retention harvest in the Pacific Northwest. Ecol. Appl. 2005, 15, 196–209. [Google Scholar] [CrossRef]
- Halaj, J.; Halpern, C.B.; Yi, H. Response of litter-dwelling spiders and carabid beetles to varying levels and patterns of green-tree retention. For. Ecol. Manag. 2008, 255, 887–900. [Google Scholar] [CrossRef]
- Sullivan, T.P.; Sullivan, D.S. Green-tree retention and recovery of an old-forest specialist, the southern red-backed vole (Myodes gapperi), 20 years after harvest. Wildl. Res. 2018, 44, 669–680. [Google Scholar] [CrossRef]
- Franklin, J.F.; Berg, D.R.; Thornburgh, D.A.; Tappeiner, J.C. Alternative silvicultural approaches to timber harvesting: Variable retention harvest systems. In Creating Forestry for the 21st Century: The Science of Ecosystem Management; Kohm, K.A., Franklin, J.F., Eds.; Island Press: Washington, DC, USA, 1997; pp. 111–139. [Google Scholar]
- Rosenvald, R.; Lõhmus, A. For what, when, and where is green-tree retention better than clear-cutting? A review of the biodiversity aspects. For. Ecol. Manag. 2008, 255, 1–15. [Google Scholar] [CrossRef]
- Otto, C.R.V.; Roloff, G.J. Songbird response to green-tree retention prescriptions in clearcut forests. Forest For. Ecol. Manag. 2012, 284, 241–250. [Google Scholar] [CrossRef]
- Machar, I. Attempt to summarize the problems: Is a sustainable management of floodplain forest geobiocenoses possible? In Biodiversity and Target Management of Floodplain Forests in the Morava River Basin (Czech Republic); Machar, I., Ed.; Palacky University: Olomouc, Czech Republic, 2010; pp. 189–226. [Google Scholar]
- Fuller, J.R. Avian responses to transitional habitats in temperate cultural landscapes. Woodland edges and young-growth. In Birds and Habitat. Relationships in Changing Landscapes; Fuller, R.J., Ed.; Cambridge Univ. Press: Cambridge, UK, 2012; pp. 125–149. [Google Scholar]
- Lance, A.; Phinney, M. Bird responses to partial retention timber harvesting in central interior British Columbia. For. Ecol. Manag. 2001, 142, 267–280. [Google Scholar] [CrossRef]
- Schieck, J.; Hobson, K.A. Bird communities associated with live residual tree patches within cut blocks and burned habitat in mixedwood boreal forests. Can. J. For. Res. 2000, 30, 1281–1295. [Google Scholar] [CrossRef]
- Poulin, J.F.; Villard, M.A.; Edman, M.; Goulet, P.J.; Eriksson, A.M. Thresholds in nesting habitat requirements of an old forest specialist, the Brown Creeper (Certhia americana), as conservation target. Biol. Conserv. 2008, 141, 1129–1137. [Google Scholar] [CrossRef]
- Franklin, J.F. Biological legacies: A critical management concept from Mount St. Helens. Trans. N. A. Wildlands Nat. Resour. Conf. 1990, 55, 216–219. [Google Scholar]
- Mazurek, M.J.; Zielinski, W.J. Individual legacy trees influence vertebrate wildlife diversity in commercial forests. For. Ecol. Manag. 2004, 193, 321–334. [Google Scholar] [CrossRef] [Green Version]
- Kilianova, H.; Pechanec, V.; Brus, J.; Kirchner, K.; Machar, I. Analysis of the development of land use in the Morava River floodplain, with special emphasis on the landscape matrix. Morav. Geogr. Rec. 2017, 25, 46–59. [Google Scholar] [CrossRef] [Green Version]
- Machar, I. Conservation and Management of Floodplain Forests in the Protected Landscape Area Litovelske Pomoravi (Czech Republic) Introduction. In Conservation and Management of Floodplain Forests in the Protected Landscape Area Litovelske Pomoravi (Czech Republic); Machar, I., Ed.; Palacky University: Olomouc, Czech Republic, 2009; pp. 7–108. [Google Scholar]
- Miko, L. Nature and landscape protection in the European context. In Ochrana prirody a krajiny v Ceske republice, vols I and II; Machar, I., Drobilova, L., Eds.; Palacky University: Olomouc, Czech Republic, 2012; pp. 43–49. [Google Scholar]
- Machar, I.; Simon, J.; Rejsek, K.; Pechanec, V.; Brus, J.; Kilianova, H. Assessment of Forest Management in Protected Areas Based on Multidisciplinary Research. Forests 2016, 7, 285. [Google Scholar] [CrossRef]
- Salekl, L.; Sivacioglu, A.; Topacoglu, O.; Zahradnile, D.; Jerabkoval, L.; Machar, I. Crowns of old remnant oak standards. Fresenius Environ. Bull. 2017, 26, 4023–4032. [Google Scholar]
- Simon, J.; Machar, I.; Bucek, A. Linking the historical research with the growth simulation model of hardwood floodplain forests. Polish J. Ecol. 2014, 62, 273–288. [Google Scholar] [CrossRef]
- Bibby, C.J.; Burges, N.D.; Hill, D.A.; Mustoe, S. Bird Census Techniques; Academic Press: London, UK, 2007; pp. 42–64. [Google Scholar]
- Alldredge, M.W.; Simons, T.R.; Pollock, K.H. A field evaluation of distance measurement error in auditory avian point count surveys. J. Wildl. Manag. 2007, 71, 2759–2766. [Google Scholar] [CrossRef]
- Kosinski, Z.; Kempa, M.; Hybsz, R. Accuracy and efficiency of different techniques for censusing territorial Middle Spotted Woodpeckers Dendrocopos medius. Acta Ornithol. 2004, 39, 29–34. [Google Scholar] [CrossRef]
- Wiens, J.A. The Ecology of Bird Communities, vol. 1, Foundation and Patterns; Cambridge University Press: Cambridge, UK, 1989; pp. 159–162. [Google Scholar]
- Stastny, K.; Hudec, K. Fauna of the Czech Republic. Birds 2 and 3, 2nd ed.; Academia: Prague, Czech Republic, 2011; pp. 1–1178. [Google Scholar]
- Sutherland, W.J. Ecological Census Techniques: A Handbook, 2nd ed.; Cambridge University Press: Cambridge, UK, 2006; pp. 33–44. [Google Scholar]
- Jaccard, P. Étude comparative de la distribution florale dans une portion des Alpes et des Jura. Bull. Soc. Vaudoise Sci. Nat. 1901, 37, 547–579. [Google Scholar]
- Rosner, B. Fundamentals of Biostatistics; Brooks/Cole, Cengage Learning: Boston, MA, USA, 2010; pp. 1–360. [Google Scholar]
- StataCorp. Stata Statistical Software: Release 15; StataCorp LLC: College Station, TX, USA, 2017. [Google Scholar]
- Machar, I.; Kulhavy, A.; Sejak, J.; Pechanec, V. Conservation effectiveness and monetary value of floodplain forests habitats in the Czech Republic. Rep. For. Res.-Zpr. Lesn. Vyzk. 2018, 63, 206–213. [Google Scholar]
- James, F.C.; Warner, N.O. Relationships between temperate forest bird communities and vegetation structure. Ecology 1982, 63, 159–171. [Google Scholar] [CrossRef]
- Perry, R.W.; Jenkins, J.M.A.; Thill, R.E.; Thompson, F.R. Long-term effects of different forest regeneration methods on mature forest birds. For. Ecol. Manag. 2018, 408, 183–194. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Franklin, J.F. Managing Stand Structure as Part of Ecologically Sustainable Forest Management in Australian Mountain Ash Forests. Conserv. Biol. 1997, 11, 1053–1068. [Google Scholar] [CrossRef]
- Skorupski, J.; Jankowiak, L.; Kiriaka, B.; Rek, T.; Wysocki, D. Beech forest structure and territory size of four songbird species in Puszcza Bukowa, NW Poland: Implications for bird-friendly silvicultural practices in a temperate forest. Ethol. Ecol. Evolut. 2017. [Google Scholar] [CrossRef]
- Uradnicek, L.; Sramek, M.; Dreslerova, J. Checklist of champion trees in the Czech Republic. J. Landsc. Ecol. 2017, 10, 109–120. [Google Scholar] [CrossRef]
- Kilianova, H.; Pechanec, V.; Svobodova, J.; Machar, I. Analysis of the evolution of the floodplain forests in the aluvium of the Morava river. In 12th International Multidisciplinary Scientific Geoconference, SGEM 2012, Albena, Bulgaria, 17–23 June 2012; SGEM: Albena, Bulgaria, 2012; Volume IV, pp. 1–8. [Google Scholar]
- Knutson, M.G.; McColl, L.E.; Suarez, S.A. Breeding bird assemblages associated with stages of forest succession in large river floodplains. Nat. Areas J. 2005, 25, 55–70. [Google Scholar]
- Miklin, J.; Hauck, D.; Konvicka, O.; Cizek, L. Veteran trees and saproxylic insects in the floodplains of Lower Morava and Dyje rivers, Czech Republic. J. Maps 2017, 13, 291–299. [Google Scholar] [CrossRef]
- Lindenmayer, D.B. The importance of managing and conserving large old trees: A case study from Victorian mountain ash forests. Royal Soc. Vic. 2016, 128, 64–70. [Google Scholar] [CrossRef]
- Remm, J.; Lõhmus, A. Tree cavities in forests—the broad distribution pattern of a keystone structure for biodiversity. For. Ecol. Manag. 2011, 262, 579–585. [Google Scholar] [CrossRef]
- Leso, P. Breeding bird communities of two succession stages of young oak forests. Sylvia 2003, 39, 67–78. [Google Scholar]
- Hunter, J.E.; Mazurek, M.J. Characteristics of trees used by nesting and roosting Vaux´s swifts nest in northwestern California. West. Birds 2004, 34, 225–229. [Google Scholar]
- Brawn, J.D.; Balda, R.P. Population biology of cavity-nesters in northern Arizona: Do nest sites limit breeding densities? Condor 1988, 90, 61–71. [Google Scholar] [CrossRef]
- Irwin, L.L.; Rock, D.F.; Miller, G.P. Stand structures used by northern spotted owls in managed forests. J. Raptor Res. 2000, 34, 175–186. [Google Scholar]
- Rodewald, A.D.; Yahner, R.H. Bird communities associated with harvested hardwood stands containing residual trees. J. Wildlife Manag. 2000, 64, 924–932. [Google Scholar] [CrossRef]
- Söderström, B. Effects of different levels of green- and dead-tree retention on hemi-boreal forest bird communities in Sweden. For. Ecol. Manag. 2009, 257, 215–222. [Google Scholar] [CrossRef]
- Müller, J.; Stadler, J.; Brandl, R. Composition versus physiognomy of vegetation as predictors of bird assemblages: The role of lidar. Remote Sens. Environ. 2010, 114, 490–495. [Google Scholar] [CrossRef]
- Radford, J.Q.; Bennett, A.F.; Cheers, G.J. Landscape-level tresholds of habitat cover for woodland-dependent birds. Biol. Conserv. 2005, 124, 317–337. [Google Scholar] [CrossRef]
- Pierson, J.C.; Mortelliti, A.; Barton, P.S.; Lane, P.W.; Lindenmayer, D.B. Evaluating the effectiveness of overstory cover as a surrogate for bird community diversity and population trends. Ecol. Indic. 2016, 61, 790–798. [Google Scholar] [CrossRef]
- Montague-Drake, R.M.; Lindenmayer, D.B.; Cunningham, R.B. Factors affecting site occupancy by woodland bird species of conservation concern. Biol. Conserv. 2009, 142, 2896–2903. [Google Scholar] [CrossRef]
- Machar, I.; Poprach, K. Tanks and cisterns for fodder molasses on farms as ecological traps. Listy Cukrov. Repar. 2012, 128, 347–349. [Google Scholar]
- Tittler, R.; Hannon, S.J.; Norton, M.R. Residual tree retention ameliorates short-term effects of clear-cutting on some boreal songbirds. Ecol. Appl. 2001, 11, 1656–1666. [Google Scholar] [CrossRef]
- Venier, L.A.; Dalley, K.; Goulet, P.; Mills, S.; Pitt, D.; Cowcill, K. Benefits of aggregate green tree retention to boreal forest birds. For. Ecol. Manag. 2015, 343, 80–87. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Wood, J.; McBurney, L.; Blair, D.; Banks, S.C. Single large versus several small: The SLOSS debate in the context of bird responses to a variable retention logging experiment. For. Ecol. Manag. 2015, 339, 1–10. [Google Scholar] [CrossRef]
Figure 1.
Bird nesting guilds in study plots 1–5 (Nesting guilds: BN = bush-nesters, CN = canopy-nesters, HN = hole-nesters, GN = ground-nesters).
Figure 1.
Bird nesting guilds in study plots 1–5 (Nesting guilds: BN = bush-nesters, CN = canopy-nesters, HN = hole-nesters, GN = ground-nesters).

{kind=link}
Table 1.
Bird density and dominance in study plots.
| Study Plots (Study Plot Number; Local Name and Coordinates; Size of the Study Plot Area; Total Amount of Individual Legacy Oak Trees (ILOT) in the Study Plot) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bird species | Nesting guilds 1 | No. 1 Pnovice 49.7100131N, 17.1330297E 2.8 ha ILOT | No. 2 Stepanov 49.6667017N, 17.1945703E 4.4 ha ILOT | No. 3 Horni les 49.7011319N, 17.0944058E 4.8 ha ILOT | No. 4 Hrube les 49.6821428N, 17.1744858E 3.4 ha 21 ILOT | No. 5 Brezova 49.6881956N, 17.1257342E 2.9 ha 19 ILOT | No. 6 Odchovna 49.6858078N, 17.1300256E 3.9 ha 26 ILOT | ||||||
| DE 2 | DO 3 | DE | DO | DE | DO | DE | DO | DE | DO | DE | DO | ||
| Aegithalos caudatus | BN | 1.7 | 3.0 | - | - | 1.9 | 3.2 | 0.4 | 0.5 | - | - | 0.6 | 0.7 |
| Anthus trivialis | GN | 0.8 | 1.4 | 2.2 | 4.1 | 0.5 | 0.9 | 0.4 | 0.5 | 0.2 | 0.2 | 0.3 | 0.4 |
| Carduelis carduelis | CN | 1.2 | 2.2 | - | - | - | - | 0.3 | 0.4 | - | - | 0.3 | 0.4 |
| Carduelis chloris | BN | 0.8 | 1.4 | 1.5 | 2.8 | 2.0 | 3.4 | 0.5 | 0.6 | 0.3 | 0.4 | - | - |
| Coccothraustes coccothraustes | BN | - | - | 0.3 | 0.6 | - | - | 0.3 | 0.4 | 0.3 | 0.4 | 0.4 | 0.5 |
| Certhia familiaris | HN | - | - | - | - | - | - | 0.3 | 0.4 | 0.6 | 0.8 | 0.2 | 0.2 |
| Columba palumbus | CN | - | - | - | - | - | - | 0.9 | 1.1 | 0.6 | 0.8 | 0.3 | 0.4 |
| Cyanistes caeruleus | HN | - | - | 0.5 | 0.9 | - | - | 4.5 | 5.4 | 3.9 | 5.1 | 5.2 | 6.1 |
| Dendrocopos major | HN | - | - | - | - | - | - | 4.2 | 5.1 | 4.2 | 5.4 | 5.0 | 5.8 |
| Dryobates minor | HN | - | - | - | - | - | - | 0.3 | 0.4 | 0.6 | 0.8 | 0.3 | 0.4 |
| Emberiza citrinella | BN | 4.4 | 7.9 | 2.2 | 4.1 | 3.9 | 6.6 | 2.2 | 2.6 | 3.0 | 3.9 | 2.1 | 2.5 |
| Erithacus rubecula | GN | 9.1 | 16.4 | 11.8 | 21.8 | 11.1 | 18.9 | 9.4 | 11.3 | 8.9 | 11.5 | 7.7 | 9.0 |
| Ficedula albicollis | HN | - | - | - | - | - | - | 4.5 | 5.4 | 3.8 | 4.9 | 5.6 | 6.5 |
| Fringilla coelebs | CN | 4.2 | 7.6 | 3.5 | 6.5 | 3.8 | 6.5 | 3.1 | 3.7 | 2.0 | 2.6 | 2.6 | 3.0 |
| Garrulus glandarius | CN | - | - | - | - | 0.5 | 0.9 | - | - | 0.3 | 0.4 | 1.0 | 1.2 |
| Hipolais icterina | BN | - | - | - | - | 0.5 | 0.9 | 0.3 | 0.4 | - | - | - | - |
| Lanius collurio | BN | - | - | 0.5 | 0.9 | - | - | - | - | 0.2 | 0.3 | - | - |
| Luscinia megarhynchos | BN | 0.8 | 1.4 | - | - | - | - | - | - | - | - | 0.3 | 0.4 |
| Leiopicus medius | HN | - | - | - | - | - | - | 3.0 | 3.6 | 3.1 | 4.0 | 3.1 | 3.5 |
| Locustella fluviatilis | GN | 1.6 | 2.9 | - | - | - | - | - | - | 0.3 | 0.4 | - | - |
| Muscicapa striata | HN | - | - | - | - | - | - | 0.6 | 0.7 | - | - | 0.3 | 0.4 |
| Oriolus oriolus | CN | - | - | - | - | - | - | 0.3 | 0.4 | 0.3 | 0.4 | 0.6 | 0.7 |
| Parus major | HN | 0.8 | 1.4 | - | - | 0.6 | 1.0 | 9.1 | 10.9 | 9.0 | 11.6 | 6.4 | 7.3 |
| Phasianus colchicus | GN | - | - | 1.0 | 1.8 | 0.9 | 1.5 | 0.6 | 0.7 | - | - | 0.3 | 0.4 |
| Phylloscopus collybita | GN | 7.2 | 13 | 10.9 | 20.1 | 9.9 | 16.9 | 4.1 | 4.9 | 5.2 | 6.5 | 7.8 | 9.1 |
| Picus viridis | HN | - | - | - | - | - | - | 0.4 | 0.5 | 0.3 | 0.4 | 0.3 | 0.4 |
| Poecile palustris | HN | - | - | - | - | - | - | 3.5 | 4.2 | 3.9 | 5.0 | 0.5 | 0.6 |
| Prunella modularis | BN | 3.5 | 6.3 | 2.5 | 4.6 | 2.4 | 4.2 | 2.1 | 2.5 | 1.9 | 2.5 | 0.7 | 0.8 |
| Sitta europaea | HN | - | - | - | - | - | - | 5.0 | 6.0 | 4.2 | 5.4 | 5.6 | 6.5 |
| Steptopelia turtur | CN | - | - | - | - | - | - | 0.2 | 0.2 | - | - | 1.3 | 1.5 |
| Strix aluco | HN | - | - | - | - | - | - | 0.3 | 0.4 | - | - | 0.3 | 0.4 |
| Sturnus vulgaris | HN | - | - | - | - | - | - | 9.6 | 11.6 | 7.1 | 9.2 | 8.7 | 10.2 |
| Turdus merula | BN | 7.2 | 13.0 | 6.2 | 11.5 | 5.9 | 10.1 | 3.9 | 4.7 | 5.5 | 7.1 | 5.3 | 6.2 |
| Turdus philomelos | CN | - | - | 1.5 | 2.8 | 0.5 | 0.9 | 0.3 | 0.4 | - | - | 0.6 | 0.7 |
| Turdus pilaris | CN | - | - | - | - | - | - | 0.6 | 0.7 | - | - | 0.9 | 1.1 |
| Turdus viscivorus | CN | - | - | - | - | - | - | 0.3 | 0.4 | - | - | - | - |
| Troglodytes troglodytes | BN | 1.7 | 3.0 | 2.1 | 3.9 | 2.3 | 3.9 | 0.3 | 0.4 | 0.6 | 0.8 | 0.9 | 1.1 |
| Sylvia atricapilla | BN | 9.1 | 16.4 | 6.9 | 12.7 | 10.1 | 17.3 | 6.6 | 7.8 | 6.7 | 8.7 | 8.7 | 10.2 |
| Sylvia curruca | BN | 1.5 | 2.7 | 0.5 | 0.9 | 1.7 | 2.9 | 0.7 | 0.8 | 0.3 | 0.4 | 1.2 | 1.4 |
| Total of DE/study plot | 55.6 | 54.1 | 58.5 | 83.1 | 77.3 | 85.4 | |||||||
| Total of species/study plot | 16 | 16 | 17 | 35 | 28 | 34 | |||||||
| Jaccard index (%) | 52 | 59 | 41 | 62 | 67 | 39 | |||||||
1 Nesting guilds: BN = bush-nesters, CN = canopy-nesters, HN = hole-nesters, GN = ground-nesters; 2 DE = density of nesting birds (nesting pairs/10 ha); 3 DO = dominance of nesting birds (%).
Table 2.
Descriptive statistics for study plot areas.
| Study Plot Number (and Local Name) | Presence of ILOT | Mean Density | Median | Standard Deviation | Skewness |
|---|---|---|---|---|---|
| 1 (Pnovice) | No | 1.43 | 0 | 2.58 | 1.97 |
| 2 (Stepanov) | No | 1.39 | 0 | 2.85 | 2.56 |
| 3 (Horni les) | No | 1.50 | 0 | 2.92 | 2.30 |
| 4 (Hrube les) | Yes | 2.13 | 0.6 | 2.76 | 1.48 |
| 6 (Odchovna) | Yes | 1.98 | 0.3 | 2.66 | 1.27 |
| 5 (Brezova) | Yes | 2.19 | 0.6 | 2.80 | 1.19 |
Table 3.
Results of Mann-Whitney U tests. (statistically significant differences are highlighted by asterisk).
Table 3.
Results of Mann-Whitney U tests. (statistically significant differences are highlighted by asterisk).
| Study Plot Number (and Local Name) | 3 (Horni les) | 2 (Stepanov) | 1 (Pnovice) |
|---|---|---|---|
| 4 (Hrube les) | U = 496.5 0.007 * | U = 462.5 0.002 * | U = 495 0.007 * |
| 6 (Odchovna) | U = 502.5 0.008 * | U = 476.5 0.004 * | U = 505 0.009 * |
| 5 (Brezova) | U = 583.5 0.066 | U = 556.0 0.033 | U = 587.5 0.071 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Machar, I.; Schlossarek, M.; Pechanec, V.; Uradnicek, L.; Praus, L.; Sıvacıoğlu, A. Retention Forestry Supports Bird Diversity in Managed, Temperate Hardwood Floodplain Forests. Forests 2019, 10, 300. https://doi.org/10.3390/f10040300
AMA Style
Machar I, Schlossarek M, Pechanec V, Uradnicek L, Praus L, Sıvacıoğlu A. Retention Forestry Supports Bird Diversity in Managed, Temperate Hardwood Floodplain Forests. Forests. 2019; 10(4):300. https://doi.org/10.3390/f10040300
Chicago/Turabian StyleMachar, Ivo, Martin Schlossarek, Vilem Pechanec, Lubos Uradnicek, Ludek Praus, and Ahmet Sıvacıoğlu. 2019. "Retention Forestry Supports Bird Diversity in Managed, Temperate Hardwood Floodplain Forests" Forests 10, no. 4: 300. https://doi.org/10.3390/f10040300
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.