N Isotope Fractionation in Tree Tissues During N Reabsorption and Remobilization in Fagus crenata Blume

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Sample Collection and Treatments
2.3. Soil Analysis
2.3.1. Total Nitrogen and δ15N Isotope Analysis
2.3.2. Amino Acid Analysis
2.3.3. Whole-Tree N Calculations
2.3.4. Statistical Analysis
3. Results
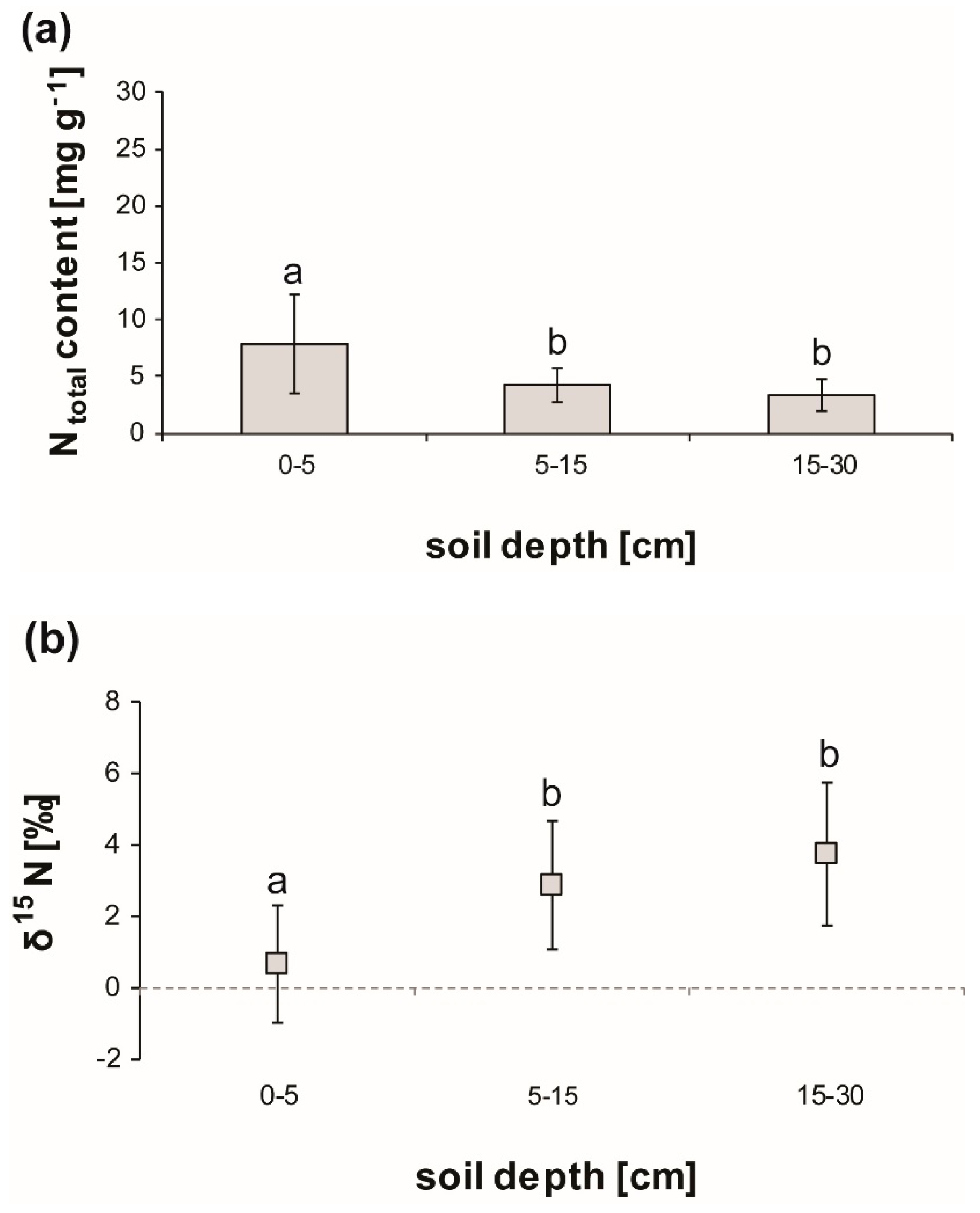
3.1. Soil Characteristics
3.2. Total Nitrogen Content in Soil and Tissues
3.3. δ15N Isotope Analysis
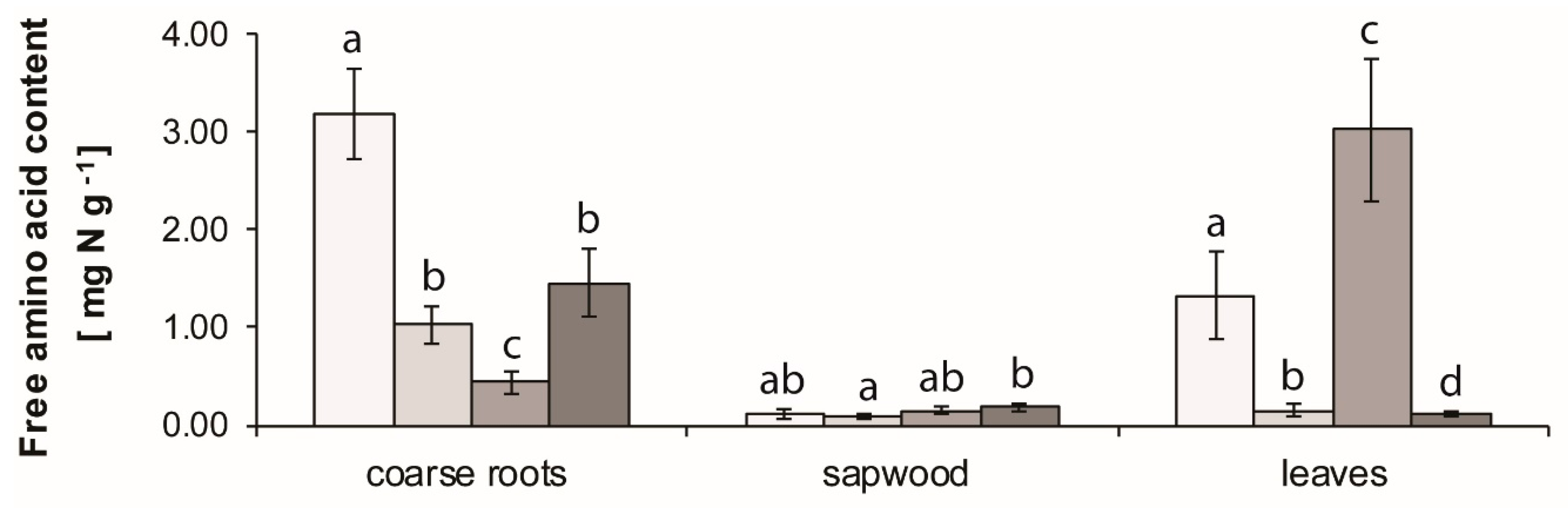
3.4. Amino Acid Analysis in Plant Tissues
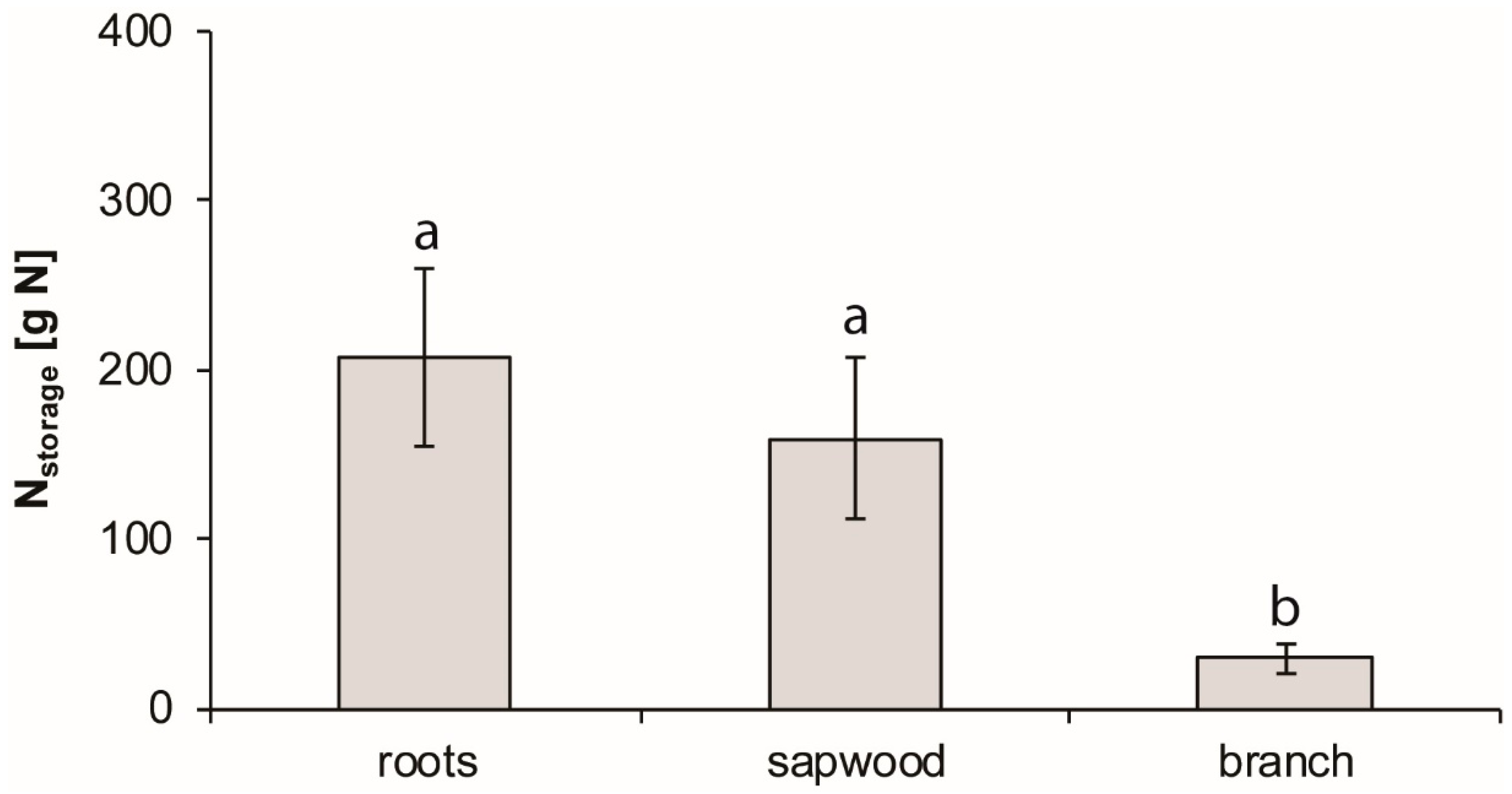
3.5. Whole-Tree Level
4. Discussion
4.1. Reabsorption, Storage, and Remobilization of N in Plant Tissues
4.2. Effect of Soil N Uptake on Plant δ15N
4.3. Effect of Intra-Plant N Movement on δ15N
4.4. Plant Tissues Free Amino Acids
4.5. Correlation between Free Amino Acid Content and Plant Tissues δ15N
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Liang, N.; Nagayama, M.; Nakata, M.; Maruyama, K. Growth, photosynthesis and nitrogen content in Japanese beech (Fagus crenata Bl.) seedlings grown under five irradiances. Photosynthetica 1995, 31, 257–268. [Google Scholar]
- Watanabe, M.; Yamaguchi, M.; Matsumura, H.; Kohno, Y.; Izuta, T. Risk assessment of ozone impact on Fagus crenata in Japan: Consideration of atmospheric nitrogen deposition. Eur. J. For. Res. 2012, 131, 475–484. [Google Scholar] [CrossRef]
- Fukushima, T.; Takasuna, H.; Matsui, T.; Nishio, T.; Kyan, T.; Tsunetomi, Y. New phytosociological classification of beech forests in Japan. Jpn. J. Ecol. 1995, 45, 79–98, (In Japanese with English Synopsis). [Google Scholar]
- Shimano, K.; Masuzawa, T. Effects of snow accumulation on survival of beech (Fagus crenata) seed. Plant Ecol. 1998, 134, 235–241. [Google Scholar]
- Yasumura, Y.; Hikosaka, K.; Hirose, T. Resource allocation to vegetative and reproductive growth in relation to mast seeding in Fagus crenata. For. Ecol. Manag. 2006, 229, 228–233. [Google Scholar] [CrossRef]
- Han, Q.; Kabeya, D.; Iio, A.; Kakubari, Y. Masting in Fagus crenata and its influence on the nitrogen content and dry mass of winter buds. Tree Physiol. 2008, 28, 1269–1276. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, Y.; Maruyama, Y.; Chiba, Y.; Kobayashi, M.J.; Joseph, B.; Shimizu, K.K.; Mochida, K.; Hiura, T.; Kon, H.; Satake, A. Nitrogen as a key regulator of flowering in Fagus crenata: Understanding the physiological mechanism of masting by gene expression analysis. Ecol. Lett. 2014, 17, 1299–1309. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.; Kabeya, D.; Iio, A.; Inagaki, Y.; Kakubari, Y. Nitrogen storage dynamics are affected by masting events in Fagus crenata. Oecologia 2014, 174, 679–687. [Google Scholar] [CrossRef] [PubMed]
- Aerts, R.; Chapin, F.S. The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns. Adv. Ecol. Res. 2000, 30, 1–67. [Google Scholar]
- Kolb, K.J.; Evans, R.D. Implication of leaf nitrogen recycling on the nitrogen isotope composition of deciduous plant tissues. New Phytol. 2002, 156, 57–64. [Google Scholar] [CrossRef]
- Yasumura, Y.; Hikosaka, K.; Matsui, K.; Hirose, T. Leaf-level nitrogen-use efficiency of canopy and understorey species in a beech forest. Funct. Ecol. 2002, 16, 826–834. [Google Scholar] [CrossRef] [Green Version]
- Yasumura, Y.; Onoda, Y.; Hikosaka, K.; Hirose, T. Nitrogen resorption from leaves under different growth irradiance in three deciduous woody species. Plant Ecol. 2005, 178, 29–37. [Google Scholar] [CrossRef]
- Lopez, M.L.; Mizota, C.; Nobori, Y.; Sasaki, T.; Yamanaka, T. Temporal changes in nitrogen acquisition of Japanese black pine (Pinus thunbergii) associated with black locust (Robinia Pseudoacacia). J. For. Res. 2014, 25, 585–589. [Google Scholar]
- Millard, P.; Grelet, G.-A. Nitrogen storage and remobilization by trees: Ecophysiological relevance in a changing world. Tree Physiol. 2010, 30, 1083–1095. [Google Scholar] [CrossRef] [PubMed]
- Dyckmans, J.; Flessa, H. Influence of tree internal N status on uptake and translocation of C and N in beech: A dual 13C and 15N labeling approach. Tree Physiol. 2001, 21, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Millard, P.; Hester, A.; Wendler, R.; Baillier, G. Interspecific defoliation responses of trees depend on sites of winter nitrogen storage. Funct. Ecol. 2001, 15, 535–543. [Google Scholar] [CrossRef] [Green Version]
- Kayama, M.; Makoto, K.; Nomura, M.; Satoh, F.; Koike, T. Nutrient dynamics and carbon partitioning in larch seedlings (Larix kaempferi) regenerated on a serpentine soil in northern Japan. Landsc. Ecol. Eng. 2009, 5, 125–135. [Google Scholar] [CrossRef]
- Leberecht, M.; Dannenmann, M.; Tejedor, J.; Simon, J.; Rennenberg, H.; Polle, A. Segregation of nitrogen use between ammonium and nitrate of ectomycorrhizas and beech trees. Plant Cell Environ. 2016, 39, 2691–2700. [Google Scholar] [CrossRef]
- Tateno, R.; Osada, N.; Terai, M.; Tokuchi, N.; Takeda, Y. Inorganic nitrogen source utilization by Fagus crenata on different soil types. Trees 2005, 19, 477–481. [Google Scholar] [CrossRef]
- Hobbie, E.A.; Högberg, P. Nitrogen isotopes link mycorrhizal fungi and plants to nitrogen dynamics. New Phytol. 2012, 196, 367–382. [Google Scholar] [Green Version]
- Gebauer, G.; Dietrich, P. Nitrogen Isotope Ratios in Different Compartments of a Mixed Stand of Spruce, Larch and Beech Trees and of Understorey Vegetation Including Fungi. Isot. Environ. Health Stud. 1993, 29, 35–44. [Google Scholar] [CrossRef]
- Fotelli, M.N.; Rennenberg, H.; Geßler, A. Effects of Drought on the Competitive Interference of an Early Succsessional Species (Rubus fruticosus) on Fagus sylvatica L. Seedlings: 15N Uptake and Partitioning, Responses of Amino Acids and other N Compounds. Plant Boil 2002, 4, 311–320. [Google Scholar] [CrossRef]
- Chikaraishi, Y.; Ogawa, N.O.; Kashiyama, Y.; Takano, Y.; Suga, H.; Tomitani, A.; Miyashita, M.; Kitazato, H.; Ohkouchi, N. Determination of aquatic food-web structure based on compound-specific nitrogen isotopic composition of amino acids. Limnol. Oceanogr. Methods 2009, 7, 740–750. [Google Scholar] [CrossRef]
- Chikaraishi, Y.; Ogawa, N.O.; Doi, H.; Ohkouchi, N. 15N/14N ratios of amino acids as a tool for studying terrestrial foodwebs: A case study of terrestrial insects (bees, wasps, and hornets). Ecol. Res. 2011, 26, 835–844. [Google Scholar] [CrossRef]
- Food and Agriculture Organization (FAO). Guidelines for Soil Description, 4th ed.; FAO: Rome, Italy, 2006. [Google Scholar]
- Tomlinson, G.; Siegwolf, R.T.W.; Buchmann, N.; Schleppi, P.; Waldner, P.; Weber, P. The mobility of nitrogen across tree-rings of Norway spruce (Picea abies L.) and the effect of extraction method on tree-ring δ 15N and δ 13C values. Rapid Commun. Mass Sprectrometry 2014, 28, 1258–1264. [Google Scholar] [CrossRef]
- Osada, N.; Yasumura, Y.; Ishida, A. Leaf nitrogen distribution in relation to crown architecture in the tall canopy species, Fagus crenata. Oecologia 2014, 175, 1093–1106. [Google Scholar] [CrossRef]
- Crooke, W.M.; Simpson, W.E. Determination of ammonium in Kjedahl digest of crops by an automated procedure. J. Sci. Food Agric. 1971, 22, 9–10. [Google Scholar] [CrossRef]
- Mulvaney, R.L. Nitrogen—Inorganic Forms. In Methods of Soil Analysis. Part 3, Chemical Methods; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Soltanpoor, P.N., Tabatabai, M.A., Johnston, C.T., Sumner, M.E., Eds.; SSSA Book Series No. 5; SSSA: Madison, WI, USA, 1996; pp. 125–1184. [Google Scholar]
- Miranda, K.M.; Espey, M.G.; Wink, D.A. A rapid, simple spectrophotometric method for simultaneous detection of nitrate and nitrite. Nitric Oxide 2001, 5, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Oikawa, A.; Ostuka, T.; Nakabayashi, R.; Jikumaru, Y.; Isuzugawa, K.; Murayama, H.; Saito, K.; Shiratake, K. Metabolic Profiling of Developing Pear Fruits Reveals Dynamic Variation in Primary and Secondary Metabolites, Including Plant Hormones. PLoS ONE 2015, 10, e0131408. [Google Scholar] [CrossRef] [PubMed]
- Ono, K.; Yasuda, Y.; Matsuo, T.; Hoshino, D.; Chiba, Y.; Mori, S. Estimating forest biomass using allometric model in a cool-temperate Fagus crenata forest in the Appi highlands, Iwate, Japan. For. Res. Inst. Res. Rep. 2013, 12, 125–141. [Google Scholar]
- Tateishi, M.; Kumagai, T.; Suyama, Y.; Hiura, T. Differences in transpiration characteristics of Japanese beech trees, Fagus crenata, in Japan. Tree Physiol. 2010, 30, 748–760. [Google Scholar] [CrossRef]
- Gebauer, T.; Horna, V.; Leuschner, C. Variability in radial sap flux density patterns and sapwood area among seven co-occurring temperate broad-leaved tree species. Tree Physiol. 2008, 28, 1821–1830. [Google Scholar] [CrossRef] [Green Version]
- Enta, A.; Hayashi, M.; Lopez, C.M.L.; Fujiyoshi, L.; Yamanaka, T.; Oikawa, A.; Seidel, F. Nitrogen resorption and fractionation during leaf senescence in typical tree species in Japan. J. For. Res. 2019, in press. [Google Scholar]
- Ueda, M.U.; Mizumachi, E.; Tokuchi, N. Foliage nitrogen turnover: Differences among nitrogen absorbed at different times by Quercus serrata saplings. Ann. Bot. 2011, 108, 169–175. [Google Scholar] [CrossRef]
- Warren, C.R.; Dreyer, E.; Adams, M.A. Photosynthesis-Rubisco relationships in foliage of Pinus sylvestris in response to nitrogen supply and the proposed role of Rubisco and amino acids as nitrogen stores. Trees 2003, 17, 359–366. [Google Scholar]
- Gessler, A.; Schneider, S.; von Sengbusch, D.; Weber, P. Field and laboratory experiments on net uptake of nitrate and ammonium by the roots of spruce (Picea abies) and beech (Fagus sylvatica) trees. New Phytol. 1998, 138, 275–285. [Google Scholar] [CrossRef]
- Cooke, J.E.K.; Weih, M. Nitrogen storage and seasonal nitrogen cycling in Populus: Bridging molecular physiology and ecophysiology. New Phytol. 2005, 167, 19–30. [Google Scholar] [CrossRef]
- Wyka, T.P.; Zytkowiak, R.; Oleksyn, J. Seasonal dynamics of nitrogen level and gas exchange in different cohorts of Scots pine needles: A conflict between nitrogen mobilization and photosynthesis? Eur. J. For. Res. 2016, 135, 483–493. [Google Scholar] [CrossRef]
- Koba, K.; Tokuchi, N.; Wada, E.; Nakajima, T.; Iwatsubo, G. Intermittent denitrification: The application of a 15N natural abundance method to a forested ecosystem. Gerchimica Cosmochim. Acta 1997, 61, 5043–5050. [Google Scholar] [CrossRef]
- Koba, K.; Tokuchi, N.; Yoshioka, T.; Hobbie, E.A.; Iwatsubo, G. Natural Abundance of Nitrogen-15 in a Forest Soil. Soil Sci. Soc. Am. J. 1998, 62, 778–781. [Google Scholar] [CrossRef]
- Shi, J.; Ohte, N.; Tokuchi, N.; Imamura, N.; Nagayama, M.; Oda, T.; Suzuki, M. Nitrate isotopic composition reveals nitrogen deposition and transformation dynamics along the canopy-soil continuum of a suburban forest in Japan. Rapid Commun. Mass Spectrom. 2014, 28, 2539–2549. [Google Scholar] [CrossRef]
- Zhou, W.-J.; Sha, L.-Q.; Schaefer, D.A.; Zhang, Y.-P.; Song, Q.-H.; Tan, Z.-H.; Deng, Y.; Deng, X.-B.; Guan, H.-L. Direct effects of litter decomposition on soil dissolved organic carbon and nitrogen in a tropical rainforest. Soil Biol. Biochem. 2014, 81, 255–258. [Google Scholar] [CrossRef]
- Rennenberg, H.; Dannenmann, M. Nitrogen nutrition of trees and temperate forests – the significance of nitrogen availability in pedosphere and atmosphere. Forests 2015, 6, 2820–2835. [Google Scholar] [CrossRef]
- Dannenmann, M.; Bimüller, C.; Gschwendtner, S.; Leberecht, M.; Tejedor, J.; Bilela, S.; Gasche, R.; Haewinkel, M.; Baltensweiler, A.; Kögel-Knabner, I.; et al. Climate change impairs nitrogen cycling in European beech forests. PLoS ONE 2016, 11, e0158823. [Google Scholar] [CrossRef]
- Högberg, P.; Johannisson, C.; Yarwood, S.; Callesen, I.; Näsholm, T.; Myrold, D.D.; Högberg, M.N. Recovery of ectomycorrhizal after ‘nitrogen saturation’ of a conifer forest. New Phytol. 2011, 189, 515–525. [Google Scholar] [CrossRef]
- Clemmensen, K.E.; Bahr, A.; Ovaskainen, O.; Dahlberg, A.; Ekblad, A.; Wallander, H.; Stenlid, J.; Finlay, R.D.; Wardle, D.A.; Lindahl, B.D. Roots and Associated Fungi Drive Long-Term Carbon Sequestration in Boreal Forests. Science 2013, 339, 1615–1618. [Google Scholar] [CrossRef]
- Finzi, A.C.; Berthrong, S.T. The uptake of amino acids by microbes and trees in three cold temperate forests. Ecology 2005, 86, 3345–3353. [Google Scholar] [CrossRef]
- Jones, D.L.; Shannon, D.; Murphy, D.V.; Farrar, J. Role of dissolved organic nitrogen (DON) in soil N cycling in grassland soils. Soil Biol. Biochem. 2004, 36, 749–756. [Google Scholar] [CrossRef]
- Roberts, P.; Jones, D.L. Microbial and plant uptake of free amino sugars in grassland soils. Soil Biol. Biochem. 2012, 49, 139–149. [Google Scholar] [CrossRef]
- Malaguti, D.; Millard, P.; Wendler, R.; Hepburn, A.; Tagliavini, M. Translocation of amino acids in the xylem of apple (Malus domestica Borkh.) trees in spring as a consequence of both N remobilization and root uptake. J. Exp. Bot. 2001, 52, 1665–1671. [Google Scholar]
- Miyashita, Y.; Dolferus, R.; Ismond, K.P.; Goog, A.G. Alanine aminotransferase catalyses the breakdown of alanine after hypoxia in Arabidopsis thaliana. Plant J. 2007, 49, 1108–1121. [Google Scholar] [CrossRef]
- Verbruggen, N.; Hermans, C. Proline accumulation in plants: A review. Amino Acids 2008, 35, 753–759. [Google Scholar] [CrossRef]
- Näsholm, T. Removal of nitrogen during needle senescence in Scots pine (Pinus sylvestris L.). Oecologia 1994, 99, 290–296. [Google Scholar] [CrossRef]
- Näsholm, T.; Ericsson, A. Seasonal changes in amino acids, protein and total nitrogen in needles of fertilized Scots pine trees. Tree Physiol. 1990, 6, 267–281. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Coordinates | m.a.s.l. | Tree | DBH | Height | Age |
|---|---|---|---|---|---|---|
| B 1 | N38° 33.390’ E139° 52.630’ | 670 | A | 30.7 | 25.4 | 71 |
| B | 30.2 | 24.4 | 71 | |||
| C | 17.0 | 27.5 | 73 | |||
| B 2 | N38° 33.260’ E139° 52.570’ | 730 | A | 25.8 | 35.2 | 75 |
| B | 31.4 | 29.1 | 78 | |||
| C | 29.9 | 43.2 | 73 | |||
| B 3 | N38° 33.310’ E139° 52.600’ | 740 | A | 17.0 | 28.1 | 80 |
| B | 25.8 | 23.0 | 78 | |||
| C | 26.0 | 31.8 | 76 |
| Depth [cm] | pH [-] | Total N [g N kg−1] | TNb [g N kg−1] | Ammonium [g N kg−1] | Nitrate [g N kg−1] | DON [g N kg−1] | C : N |
|---|---|---|---|---|---|---|---|
| 0–5 | 3.6 ± 0.1 | 7.8 ± 2.1 | 0.29 ± 0.08 | 0.25 ± 0.05 | 0.004 ± 0.006 | 0.04 ± 0.03 | 20 ± 2 |
| 5–15 | 3.7 ± 0.3 | 3.6 ± 0.9 | 0.17 ± 0.03 | 0.10 ± 0.07 | 0.003 ± 0.005 | 0.07 ± 0.07 | 19 ± 1 |
| 15–30 | 3.8 ± 0.2 | 1.0 ± 0.2 | 0.14 ± 0.05 | 0.10 ± 0.08 | 0.003 ± 0.004 | 0.03 ± 0.03 | 15 ± 1 |
| Tissue | Period | Alanine [g N kg−1] | Arginine [g N kg−1] | Asparagine [g N kg−1] |
|---|---|---|---|---|
| Perennial roots | shoot growth | 0.34 ± 0.06 | 0.15 ± 0.10 | 2.2 ± 0.03 |
| green leaf | 0.06 ± 0.02 | 0.06 ± 0.03 | 0.76 ± 0.09 | |
| pre-abscission | 0.05 ± < 0.01 | 0.03 ± 0.01 | 0.24 ± 0.10 | |
| post abscission | 0.18 ± 0.05 | 0.13 ± 0.08 | 0.88 ± 0.021 | |
| Sapwood | shoot growth | 0.03 ± 0.10 | 0.00 ± 0.00 | 0.02 ± 0.01 |
| green leaf | 0.02 ± 0.01 | 0.00 ± 0.00 | 0.03 ± 0.01 | |
| pre-abscission | 0.06 ± 0.01 | 0.00 ± 0.00 | 0.04 ± 0.02 | |
| post abscission | 0.06 ± 0.01 | 0.01 ± <0.01 | 0.07 ± 0.02 | |
| leaf | shoot growth | 0.63 ± 0.28 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| green leaf | 0.05 ± 0.02 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| pre-abscission | 1.02 ± <0.01 | 0.01 ± <0.01 | 0.14 ± 0.01 | |
| Litter | post abscission | 0.02 ± <0.01 | 0.01 ± <0.01 | 0.09 ± 0.05 |
| Tissue | Amino Acid | Estimate | R2 | Adjusted R2 | p |
|---|---|---|---|---|---|
| roots | Asparagine | 1.52 | 0.67 | 0.60 | 0.002** |
| Other 1 | −2.90 | 0.003** | |||
| sapwood | Alanine | −14.53 | 0.53 | 0.36 | 0.026* |
| Asparagine | 37.18 | 0.349 | |||
| Other 1 | 0.38 | 0.976 | |||
| leaves | Alanine | 1.56 | 0.76 | 0.73 | 0.013* |
| Other 1 | 0.87 | 0.001** |
| Tissue | Biomass [kg] | N [g N] | ||
|---|---|---|---|---|
| Green Leaf Stage | Pre-abscission Stage | Post-abscission Stage | ||
| roots | 117 ± 9 a | 679 ± 53 | 862 ± 119* | 893 ± 106* |
| sapwood | 362 ± 20 b | 245 ± 70 | 208 ± 84 | 339 ± 126* |
| branches | 68 ± 5 a | 54 ± 8 | 52 ± 5 | 83 ± 10* |
| leaves | 8.7 ± 0.1 c | 193 ± 20 | 144 ± 26* | 64 ± 6* |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seidel, F.; Lopez C., M.L.; Celi, L.; Bonifacio, E.; Oikawa, A.; Yamanaka, T. N Isotope Fractionation in Tree Tissues During N Reabsorption and Remobilization in Fagus crenata Blume. Forests 2019, 10, 330. https://doi.org/10.3390/f10040330
Seidel F, Lopez C. ML, Celi L, Bonifacio E, Oikawa A, Yamanaka T. N Isotope Fractionation in Tree Tissues During N Reabsorption and Remobilization in Fagus crenata Blume. Forests. 2019; 10(4):330. https://doi.org/10.3390/f10040330
Chicago/Turabian StyleSeidel, Felix, M. Larry Lopez C., Luisella Celi, Eleonora Bonifacio, Akira Oikawa, and Toshiro Yamanaka. 2019. "N Isotope Fractionation in Tree Tissues During N Reabsorption and Remobilization in Fagus crenata Blume" Forests 10, no. 4: 330. https://doi.org/10.3390/f10040330