Carbon Isotopes of Riparian Forests Trees in the Savannas of the Volta Sub-Basin of Ghana Reveal Contrasting Responses to Climatic and Environmental Variations


Abstract
:1. Introduction
2. Materials and Methods
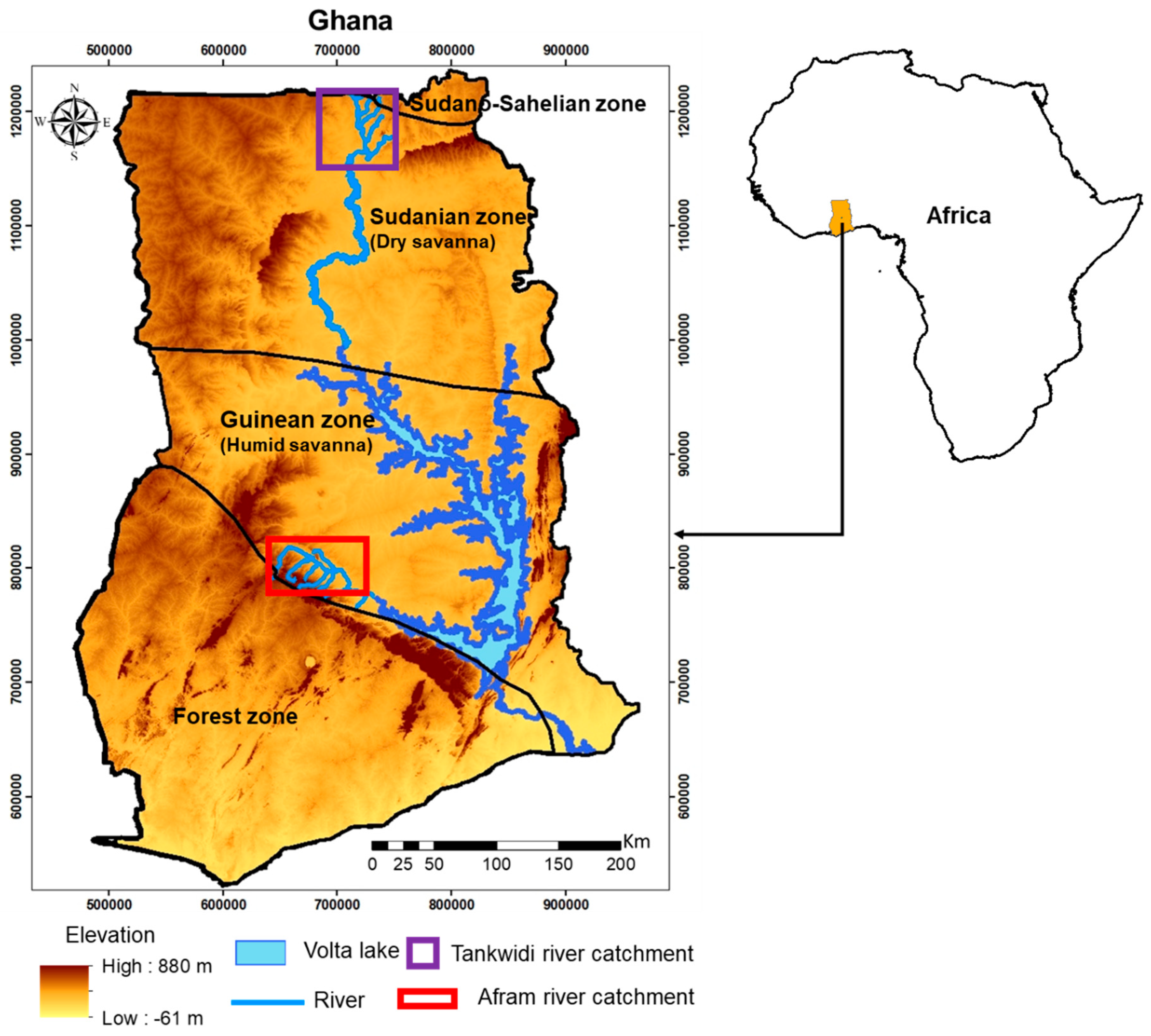
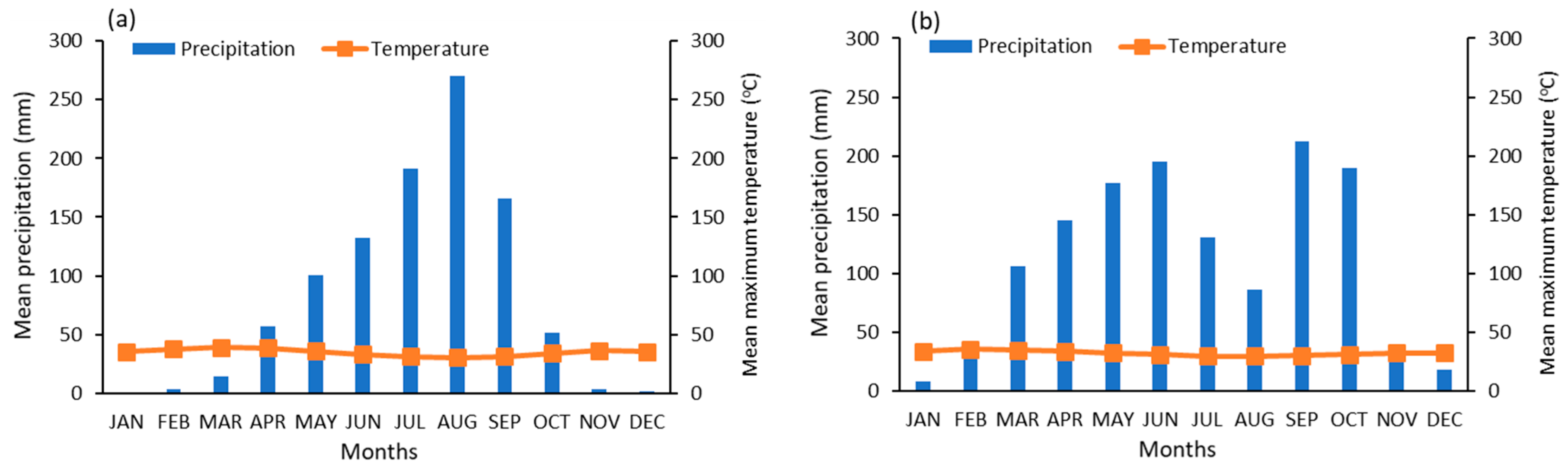
2.1. Study Area
2.2. Tree-Ring Stable Isotope Analyses
2.3. Statistical Analysis
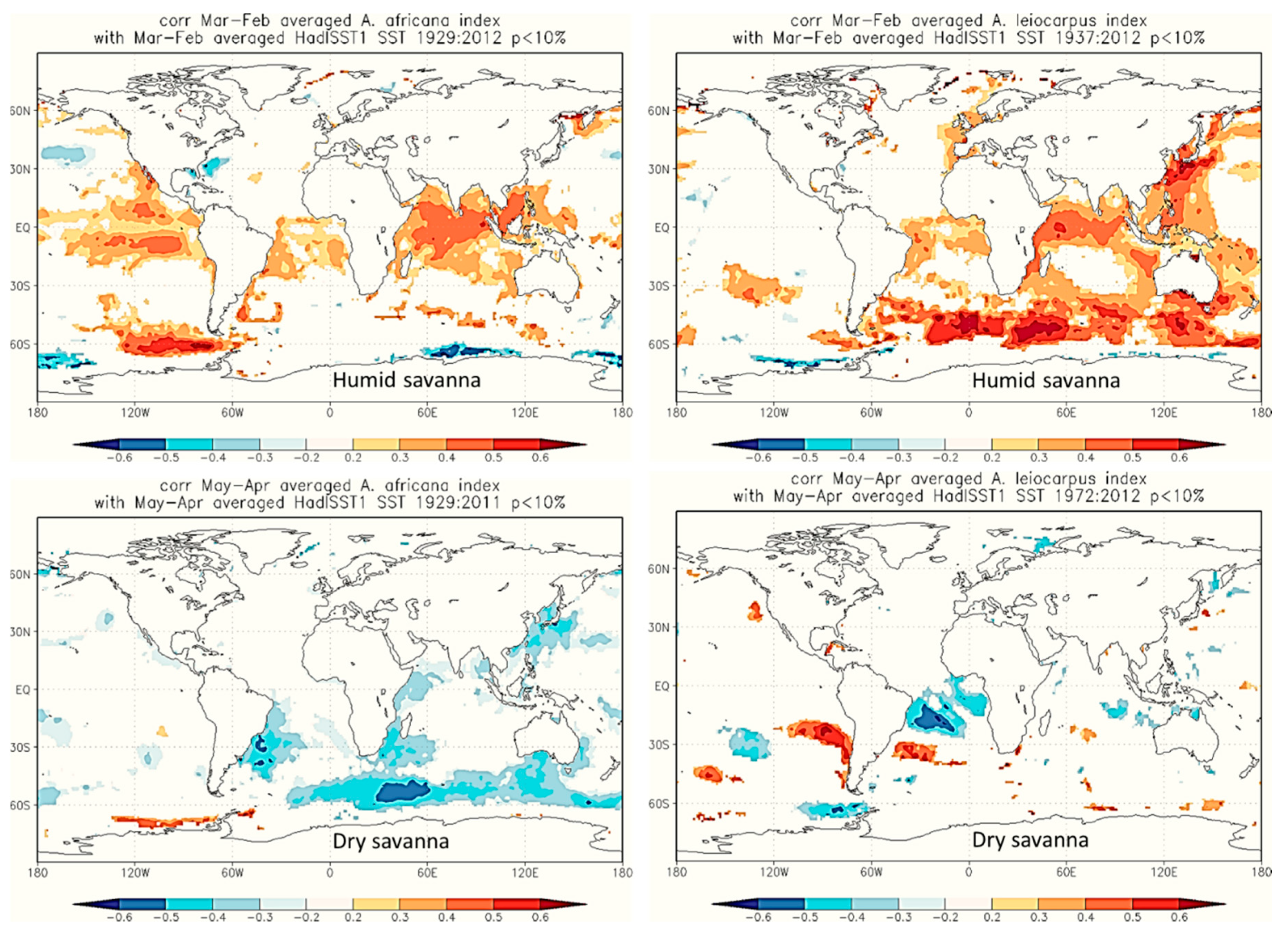
3. Results
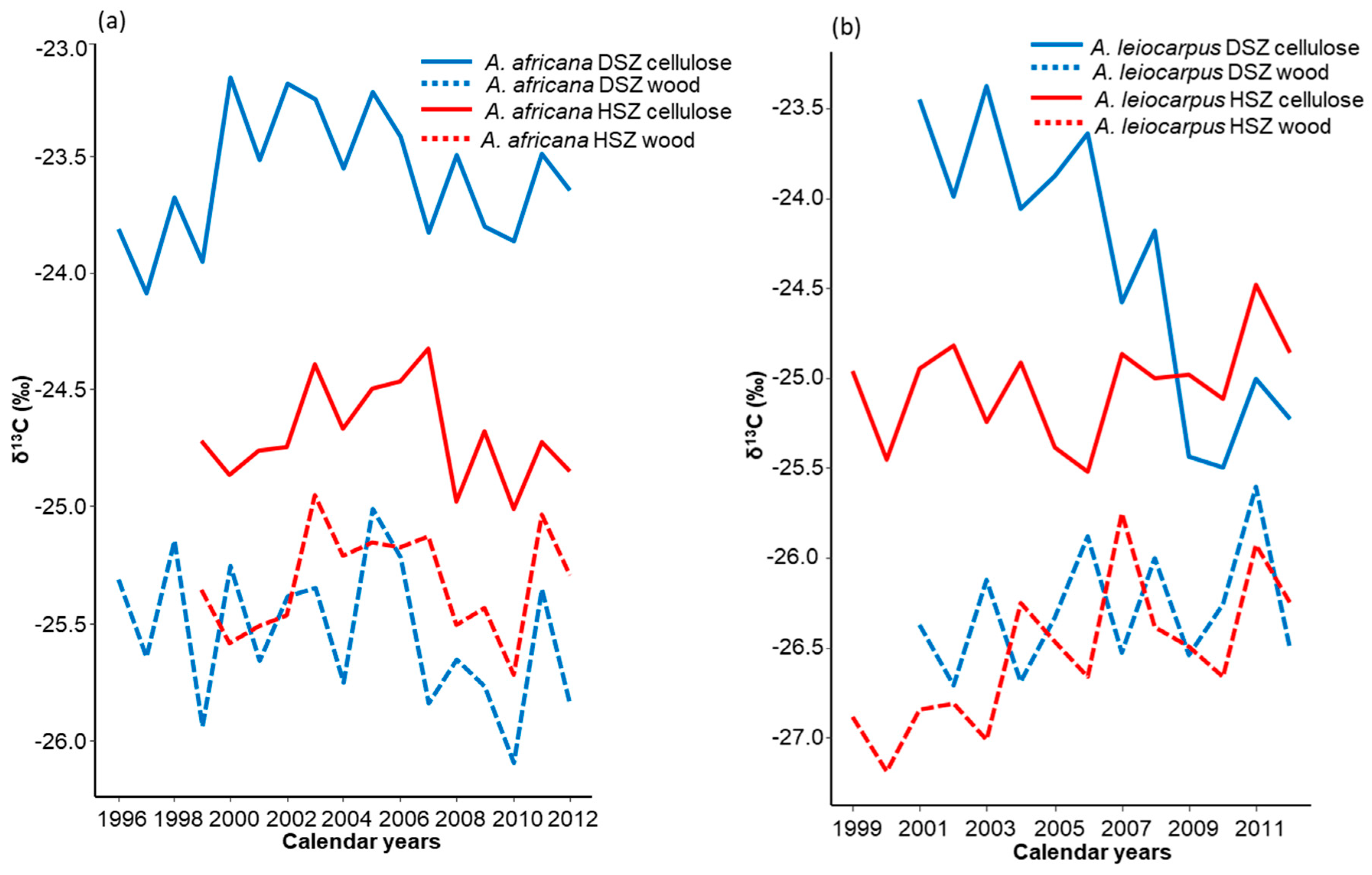
3.1. Relationship between 13C Chronologies of Whole Wood and Cellulose
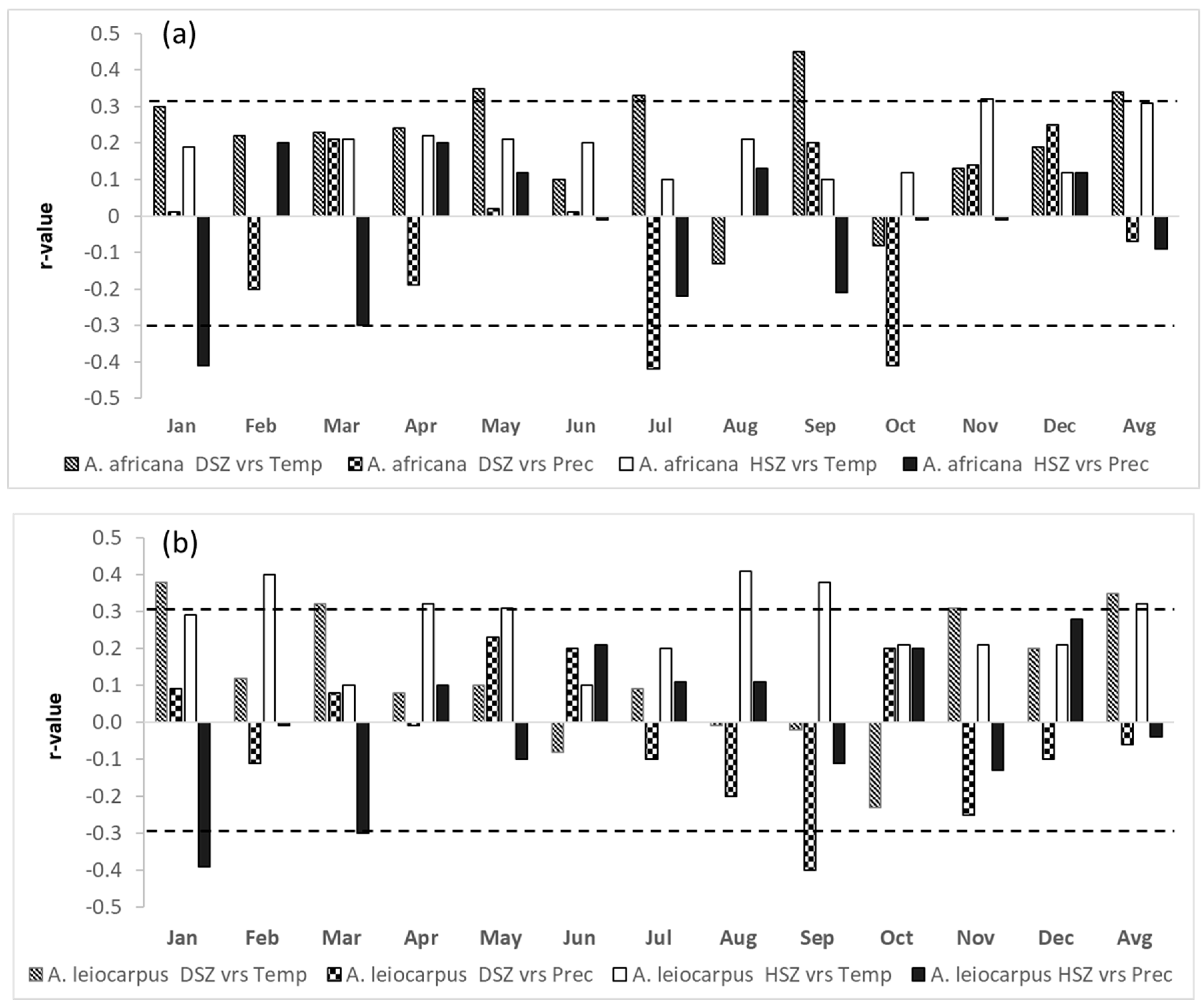
3.2. Relationships between Tree-Ring δ13C Values and Climatic Parameters
4. Discussion
4.1. Variation of δ13C Values of Bulk Wood and Cellulose in Tree Rings
4.2. Patterns and Variations in Tree Rings δ13C Values in Different Tree Species
4.3. Relationship Between Tree-Ring δ13C and Climatic Parameters
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sambare, O.; Bognounou, F.; Wittig, R.; Thiombiano, A. Woody species composition, diversity and structure of riparian forests of four watercourses types in Burkina Faso. J. Res. 2011, 22, 145–158. [Google Scholar] [CrossRef]
- Azihou, F.K.; Glele, K.R.; Ronald, B.; Brice, S. Distribution of tree species along a gallery forest–savanna gradient: Patterns, overlaps and ecological thresholds. J. Trop. Ecol. 2013, 29, 25–37. [Google Scholar] [CrossRef]
- Naiman, R.J.; Decamps, H. The Ecology of Interfaces: Riparian Zones. Annu. Rev. Ecol. Evol. Syst. 1997, 28, 621–658. [Google Scholar] [CrossRef]
- Gray, C.L.; Slade, E.M.; Mann, D.J.; Lewis, O.T. Do riparian reserves support dung beetle biodiversity and ecosystem services in oil palm-dominated tropical landscapes? Ecol. Evol. 2014, 4, 1049–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Intergovernmental Panel on Climate Change (IPCC). IPCC Fourth Assessment Report: Climate Change (AR5); IPCC: Geneva, Switzerland, 2013. [Google Scholar]
- Gebrekirstos, A.; Teketay, D.; Fetene, M.; Worbes, M.; Mitlohner, R. Stable carbon isotope ratios in tree rings of co-occurring species from semi-arid tropics in Africa: Patterns and climatic signals. Glob. Planet. Chang. 2009, 66, 253–260. [Google Scholar] [CrossRef]
- Schöngart, J.; Orthmann, B.; Hennenberg, K.J.; Porembski, S.; Worbes, M. Climate-growth relationships of tropical tree species in West Africa and their potential for climate reconstruction. Glob. Chang. Biol. 2006, 12, 1130–1150. [Google Scholar] [CrossRef]
- Schöngart, J.; Bräuning, A.; Barbosa, A.C.M.C.; Lisi, C.S.; de Oliveira, J.M. Dendroecological Studies in the Neotropics: History, Status and Future Challenges. In Dendroecology. Ecological Studies (Analysis and Synthesis); Amoroso, M., Daniels, L., Baker, P., Camarero, J., Eds.; Springer: Cham, Germany, 2017; Volume 231. [Google Scholar]
- Gebrekirstos, A.; Mitlohner, R.; Teketay, D.; Worbes, M. Climate–growth relationships of the dominant tree species from semi-arid savanna woodland in Ethiopia. Trees 2008, 22, 631–641. [Google Scholar] [CrossRef] [Green Version]
- Colombaroli, D.; Cherubini, P.; De Ridder, M.; Saurer, M.; Toirambe, B.; Zweifel, N.; Beeckman, H. Stable carbon and oxygen isotopes in tree rings show physiological responses of Pericopsis elata to precipitation in the Congo Basin. J. Trop. Ecol. 2016, 32, 213–225. [Google Scholar] [CrossRef]
- Boakye, E.A.; Gebrekirstos, A.; Hyppolite, D.N.; Barnes, V.R.; Kouamé, F.N.; Porembski, S.; Bräuning, A. Influence of climatic variation factors on the tree growth of the trees Afzelia africana and Anogeissus leiocarpus in riparian forests in the humid and dry savannas of the Volta basin, in Ghana. Trees 2016, 30, 1695–1709. [Google Scholar] [CrossRef]
- Mokria, M.; Gebrekirstos, A.; Abiyu, A.; Van Noordwijk, M.; Bräuning, A. Multi-century tree-ring precipitation record reveals increasing frequency of extreme dry events in the upper Blue Nile River catchment. Glob. Chang. Biol. 2017, 23, 5436–5454. [Google Scholar] [CrossRef]
- Brienen, R.J.W.; Wanek, W.; Hietz, P. Stable carbon isotopes in tree rings indicate improved water use efficiency and drought responses of a tropical dry forest tree species. Trees 2011, 25, 103–113. [Google Scholar] [CrossRef]
- Fichtler, E.; Helle, G.; Worbes, M. Stable-carbon isotope time series from tropical tree rings indicate a precipitation signal. Trees 2010, 66, 35–50. [Google Scholar] [CrossRef]
- Nakai, W.; Okada, N.; Sano, M.; Nakatsuka, T. Sample preparation of ring-less tropical trees for δ18O measurement in isotope dendrochronology. Tropics 2018, 27, 49–58. [Google Scholar] [CrossRef]
- Gebrekirstos, A.; Mitlohner, R.; van Noordwijk, M.; Bräuning, A. Tracing responses to climate variability from stable isotopes in tree rings of Anogeissus leiocarpus and Sclerocarya birrea from the Sahel zone, Burkina Faso. Trace 2012, 10, 6–12. [Google Scholar]
- Gebrekirstos, A.; Bräuning, A.; Van Noordwijk, M.; Mitlohner, R. Understanding past, present, and future climate changes from East to West Africa. Agric. Innov. Sustain. Dev. 2011, 3, 77–86. [Google Scholar]
- Huang, R.; Zhu, H.; Liu, X.; Liang, E.; Grießinger, J.; Braeuning, A.; Li, X.; Wu, G. Does increasing intrinsic water use efficiency (iWUE) stimulate tree growth at natural alpine timberline on the southeastern Tibetan Plateau? Glob. Planet. Chang. 2017, 148, 217–226. [Google Scholar] [CrossRef]
- Saurer, M.; Siegwolf, R.T.W. Human impacts on tree-ring growth reconstructed from stable isotopes. In Stable Isotopes as Indicators of Ecological Change; Dawson, T.E., Siegwolf, R.T.W., Eds.; Terrestrial Ecology; Elsevier: Amsterdam, The Netherlands, 2007; pp. 49–62. [Google Scholar]
- Tognetti, R.; Lombardi, F.; Lasserre, B.; Cherubini, P.; Marchetti, M. Tree-ring stable isotopes reveal twentieth-century increases in water-use efficiency of Fagus sylvatica and Nothofagus spp. in Italian and Chilean mountains. PLoS ONE 2014, 9, e113136. [Google Scholar] [CrossRef]
- Gebrekirstos, A.; Bräuning, A.; Sass-Klassen, U.; Mbow, C. Opportunities and applications of dendrochronology in Africa. Curr. Opin. Environ. Sustain. 2014, 6, 48–53. [Google Scholar] [CrossRef] [Green Version]
- Gebrekirstos, A.; Teketay, D.; Mitlöhner, R. Responses of Dobera glabra and Eight Co-occurring Species to Drought and Salinity Stress at a Savanna-Scrub Ecotone: Implications in the Face of Climate Change. Open J. For. 2014, 4, 327–337. [Google Scholar] [CrossRef]
- Hartl-Meier, C.; Zang, C.; Büntgen, U.; Esper, J.; Rothe, A.; Göttlein, A.; Dirnböck, T.; Treydte, K. Uniform climate sensitivity in tree-ring stable isotopes across species and sites in a mid-latitude temperate forest. Tree Physiol. 2015, 35, 4–15. [Google Scholar] [CrossRef]
- O’Leary, M.H. Carbon Isotopes in Photosynthesis: Fractionation techniques may reveal new aspects of carbon dynamics in plants. BioScience 1988, 38, 328–336. [Google Scholar] [CrossRef]
- Brugnoli, E.; Farquhar, G.D. Photosynthetic Fractionation of Carbon Isotopes. In Photosynthesis: Physiology and Metabolism; Leegood, R.C., Sharkey, T.D., Von Caemmerer, S., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; pp. 352–434. [Google Scholar]
- Ehleringer, J.R.; Dawson, T.E. Water-Uptake by Plants: Perspectives from Stable Isotope Composition. Plant Cell Environ. 1992, 15, 1073–1082. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Lloyd, J. Carbon and Oxygen Isotope Effects in the Exchange of Carbon Dioxide between Terrestrial Plants and the Atmosphere. In Stable Isotopes and Plant Carbon-water Relations; Ehleringer, J.R., Hall, A.E., Farquhar, G.D., Eds.; Academic Press: San Diego, CA, USA, 1993; pp. 47–70. [Google Scholar]
- McCarroll, D.; Loader, N.J. Stable isotopes in tree rings. Quat. Sci. Rev. 2004, 23, 771–801. [Google Scholar] [CrossRef] [Green Version]
- Poorter, L.; Markesteijn, L. Seedling traits determine drought tolerance of tropical tree species. Biotropica 2008, 40, 321–331. [Google Scholar] [CrossRef]
- Mensah, S.; Houehanou, T.D.; Sogbohossou, E.A.; Assogbadjo, A.E.; Glèlè Kakaï, R. Effect of human disturbance and climatic variability on the population structure of Afzelia africana Sm. ex pers. (Fabaceae-Caesalpinioideae) at country broad-scale (Bénin, West Africa). S. Afr. J. Bot. 2014, 95, 165–173. [Google Scholar] [CrossRef]
- UNEP-GEF. Volta River Basin, West Africa. Available online: http://www.iwmi.cgiar.org/assessment/files_new/research_projects/River_Basin_Development_and_Management/VoltaRiverBasin_Boubacar.pdf (accessed on 6 September 2015).
- De Condappa, D.; Lemoalle, J. Volta River Basin Atlas; CGIAR: Montpellier, France, 2009; p. 96. [Google Scholar]
- Callo-Concha, D.; Gaiser, T.; Ewert, F. Farming and Cropping Systems in the West African Sudanian Savanna. WASCAL Research Area: Northern Ghana, Southwest Burkina Faso and Northern Benin; ZEF Working Paper 100; University of Bonn: Bonn, Germany, 2012. [Google Scholar]
- Konate, S.; Kampmann, D. Biodiversity Atlas of West Africa; Goethe University: Frankfurt, German, 2010; Volume III. [Google Scholar]
- Boakye, E.A.; Hyppolite, D.N.; Barnes, V.R.; Porembski, S.; Thiel, M.; Kouame’, F.N.; Kone, D. Threat of agricultural production on woody plant diversity in Tankwidi riparian buffer in the Sudanian Savanna of Ghana. Int. J. Biodivers. Conserv. 2015, 7, 354–363. [Google Scholar]
- Boakye, E.A.; Hyppolite, D.N.; Barnes, V.R.; Porembski, S.; Thiel, M.; Kouamé, F.N.; Kone, D. Comparative analysis of woody composition of farmlands and forest reserve along Afram river in a tropical humid savanna of Ghana: Implications to climate change adaptation. In Innovation in Climate Change Adaptation, Climate Change; Filho, W.L., Ed.; Springer: Basel, Switzerland, 2016; pp. 195–209. [Google Scholar]
- Odekunle, T.O.; Eludoyin, A.O. Sea surface temperature patterns in the Gulf of Guinea: Their implications for the spatio-temporal variability of precipitation in West Africa. Int. J. Clim. 2008, 28, 1507–1517. [Google Scholar] [CrossRef]
- van der Sleen, P.; Zuidema, P.A.; Pons, T.L. Stable isotopes in tropical tree rings: Theory, methods and applications. Funct. Ecol. 2017, 31, 1674–1689. [Google Scholar] [CrossRef]
- D’Alessandro, C.M.; Guerrieri, M.R.; Saracino, A. Comparing carbon isotope composition of bulk wood and holocellulose from Quercus cerris, Fraxinus ornus and Pinus radiata tree rings. Forest 2004, 1, 51–57. [Google Scholar] [CrossRef]
- Sidorova, O.V.; Siegwolf, R.T.W.; Saurer, M.; Naurzbaev, M.M.; Vaganov, E.A. Isotopic composition (d13C, d18O) in wood and cellulose of Siberian larch trees for early Medieval and recent periods. J. Geophys. Res. 2008, 113, 148–227. [Google Scholar] [CrossRef]
- Krepkowski, J.; Gebrekirstos, A.; Shibistova, O.; Brauning, A. Stable carbon isotope labeling reveals different carry-over effects between functional types of tropical trees in an Ethiopian mountain forest. New Phytol. 2013, 199, 441–451. [Google Scholar] [CrossRef] [PubMed]
- Borella, S.; Leuenberger, M.; Saurer, M.; Siegwolf, R.T.W. Reducing uncertainties in δ13C analysis of tree rings: Pooling, milling and cellulose extraction. J. Geophys. Res. 1998, 103, 19519–19526. [Google Scholar] [CrossRef]
- Loader, N.J.; Robertson, I.; McCarroll, D. Comparison of stable carbon isotope ratios in the whole wood, cellulose and lignin of oak tree-rings. Palaeogeogr. Palaeoclim. Palaeoecol. 2003, 196, 395–407. [Google Scholar] [CrossRef]
- Schleser, G.H.; Anhuf, D.; Helle, G.; Vos, H. A remarkable relationship of the stable carbon isotopic compositions of wood and cellulose in tree-rings of the tropical species Cariniana micrantha (Ducke) from Brazil. Chem. Geol. 2015, 401, 59–66. [Google Scholar] [CrossRef]
- Wieloch, T.; Helle, G.; Heinrich, I.; Voigt, M.; Schyma, P. A novel device for batchwise isolation of a-cellulose from small-amount wholewood samples. Dendrochronologia 2011, 29, 115e–117e. [Google Scholar] [CrossRef]
- Laumer, W.; Andreu, L.; Helle, G.; Schleser, G.H.; Wieloch, T.; Wissel, H. A novel approach for the homogenization of cellulose to use micro-amounts for stable isotope analyses. Rapid Commun. Mass Spectrom. 2009, 23, 1934–1940. [Google Scholar] [CrossRef] [PubMed]
- Battipaglia, G.; Jäggi, M.; Saurer, M.; Siegwolf, R.T.W.; Cotrufo, M.F. Climatic sensitivity of δ18O in the wood and cellulose of tree rings: Results from a mixed stand of Acer pseudoplatanus L. and Fagus sylvatica L. Palaeogeogr. Palaeoclim. Palaeoecol. 2008, 261, 193–202. [Google Scholar] [CrossRef]
- Ferrio, J.P.; Voltas, J. Carbon and oxygen isotope ratios in wood constituents of Pinus halepensis as indicators of precipitation, temperature and vapour pressure deficit. Tellus B 2005, 57, 164–173. [Google Scholar] [CrossRef]
- Roden, J.S.; Farquhar, G.D. A Controlled Test of the Dual-Isotope Approach for the Interpretation of Stable Carbon and Oxygen Isotope Ratio Variation in Tree Rings. Tree Physiol. 2012, 32, 490–503. [Google Scholar] [CrossRef]
- Gebrekirstos, A.; Teketay, D.; Fetene, M.; Mitlohner, R. Adaptation of five co-occurring tree and shrub species to water stress and its implication in restoration of degraded lands. For. Ecol. Manag. 2006, 229, 259–267. [Google Scholar] [CrossRef]
- Hasselquist, N.J.; Allen, M.F.; Santiago, L.S. Water relations of evergreen and drought-deciduous trees along a seasonally dry tropical forest chronosequence. Oecologia 2010, 164, 881–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brodribb, T.J.; McAdam, S.A.M.; Jordan, G.J.; Martins, S.C.V. Conifer species adapt to low-rainfall climates by following one of two divergent pathways. Proc. Natl. Acad. Sci. USA 2014, 111, 14489–14493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Bioi. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Tomlinson, K.W.; Poorter, L.; Bongers, F.; Borghetti, F.; Jacobs, L.; van Langevelde, F. Relative growth rate variation of evergreen and deciduous savanna tree species is driven by different traits. Ann. Bot. 2014, 114, 315–324. [Google Scholar] [CrossRef] [Green Version]
- Dodd, I.C.; Ryan, A.C. Whole-Plant Physiological Responses to Water-Deficit Stress; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2016. [Google Scholar]
- Balas, N.; Nicholson, S.E.; Klotter, D. The relationship of rainfall variability in West Central Africa to sea-surface temperature fluctuations. Int. J. Clim. 2007, 27, 1335–1349. [Google Scholar] [CrossRef] [Green Version]
- Fontaine, B.; Janicot, S. Sea surface temperature fields associated with West African rainfall anomaly types. J. Clim. 1996, 9, 2935–2940. [Google Scholar] [CrossRef]
- Schifman, L.A.; Stella, J.C.; Teece, M.; Volk, T.A. Plant growth and water stress response of hybrid willow (Salix spp.) among sites and years in central New York. Biomass Bioenergy 2012, 36, 316–326. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dry Savanna Zone | Humid Savanna Zone | |||
|---|---|---|---|---|
| A. africana | A. leiocarpus | A. africana | A. leiocarpus | |
| Whole wood (δ13C) | ||||
| Number of rings measured | 101 | 41 | 91 | 76 |
| Mean (‰) | −25.54 | −26.29 | −25.32 | −26.54 |
| Std. Dev. (‰) | 0.31 | 0.34 | 0.22 | 0.41 |
| Maximum (‰) | −25.01 | −25.60 | −24.95 | −25.75 |
| Minimum (‰) | −26.09 | −26.71 | −25.72 | −27.19 |
| Correlation (ring width and δ13C) | −0.06 | 0.14 | −0.15 | −0.09 |
| p-value of correlation | 0.52 | 0.39 | 0.18 | 0.44 |
| Cellulose (δ13C) | ||||
| Mean (‰) | −23.59 | −24.36 | −24.69 | −25.04 |
| Std. Dev. (‰) | 0.28 | 0.77 | 0.21 | 0.28 |
| Number of rings measured | 17 | 12 | 14 | 14 |
| Offset (Whole wood-Cellulose, ‰) | 1.95 | 1.93 | 0.63 | 1.50 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boakye, E.A.; Gebrekirstos, A.; Hyppolite, D.N.; Barnes, V.R.; Porembski, S.; Bräuning, A. Carbon Isotopes of Riparian Forests Trees in the Savannas of the Volta Sub-Basin of Ghana Reveal Contrasting Responses to Climatic and Environmental Variations. Forests 2019, 10, 251. https://doi.org/10.3390/f10030251
Boakye EA, Gebrekirstos A, Hyppolite DN, Barnes VR, Porembski S, Bräuning A. Carbon Isotopes of Riparian Forests Trees in the Savannas of the Volta Sub-Basin of Ghana Reveal Contrasting Responses to Climatic and Environmental Variations. Forests. 2019; 10(3):251. https://doi.org/10.3390/f10030251
Chicago/Turabian StyleBoakye, Emmanuel Amoah, Aster Gebrekirstos, Dibi N’da Hyppolite, Victor Rex Barnes, Stefan Porembski, and Achim Bräuning. 2019. "Carbon Isotopes of Riparian Forests Trees in the Savannas of the Volta Sub-Basin of Ghana Reveal Contrasting Responses to Climatic and Environmental Variations" Forests 10, no. 3: 251. https://doi.org/10.3390/f10030251
APA StyleBoakye, E. A., Gebrekirstos, A., Hyppolite, D. N., Barnes, V. R., Porembski, S., & Bräuning, A. (2019). Carbon Isotopes of Riparian Forests Trees in the Savannas of the Volta Sub-Basin of Ghana Reveal Contrasting Responses to Climatic and Environmental Variations. Forests, 10(3), 251. https://doi.org/10.3390/f10030251