Combining a Quantum Cascade Laser Spectrometer with an Automated Closed-Chamber System for δ13C Measurements of Forest Soil, Tree Stem and Tree Root CO2 Fluxes

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Measurements of Soil, Root and Stem Respiration
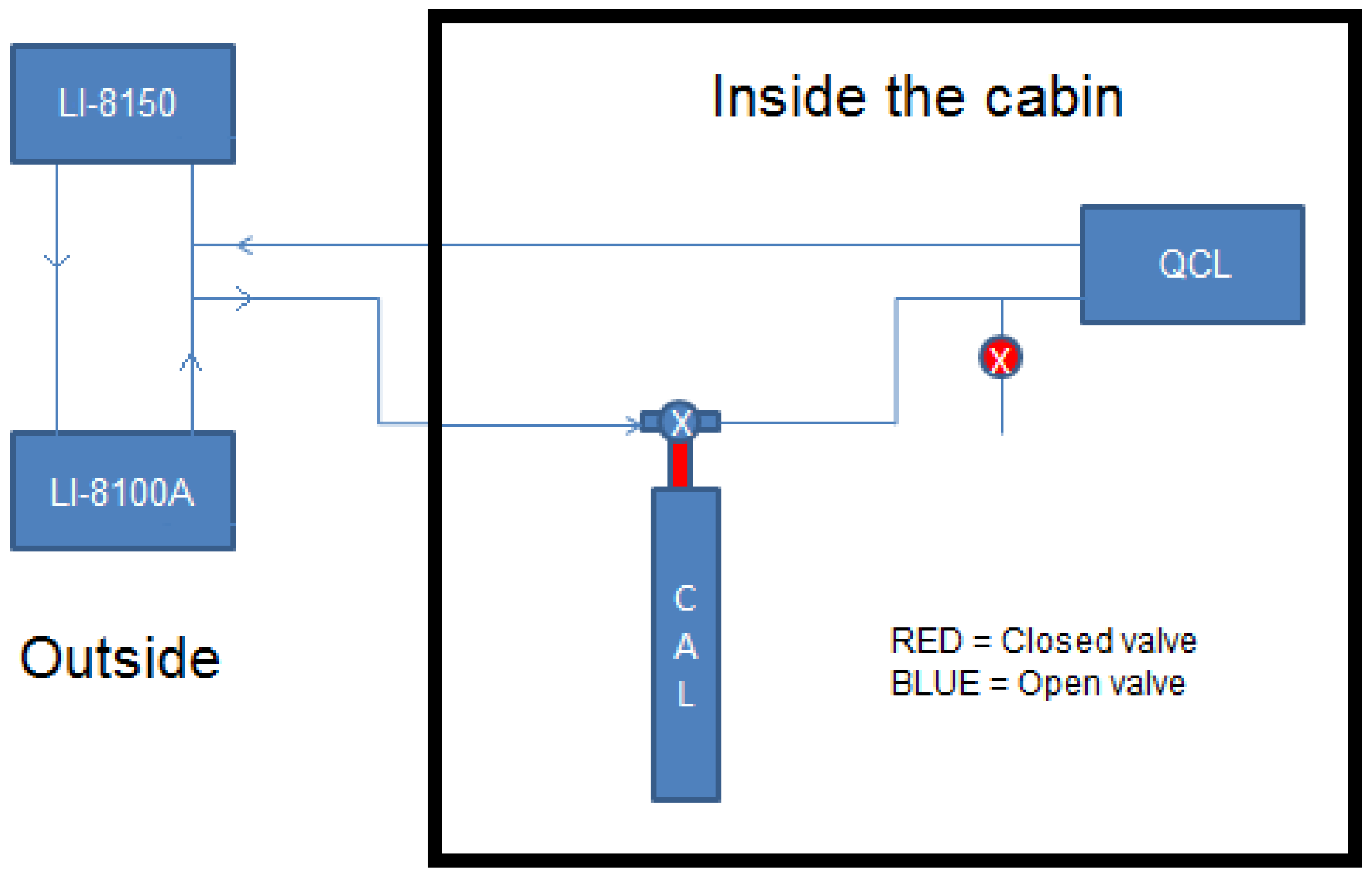
2.3. Combining the Aerodyne Laser Spectrometer with the Li-8100A/8150
2.4. Measurement Protocol
2.5. Test for Water Vapour and CO2 Concentration Dependence
2.6. Data Analysis
3. Results
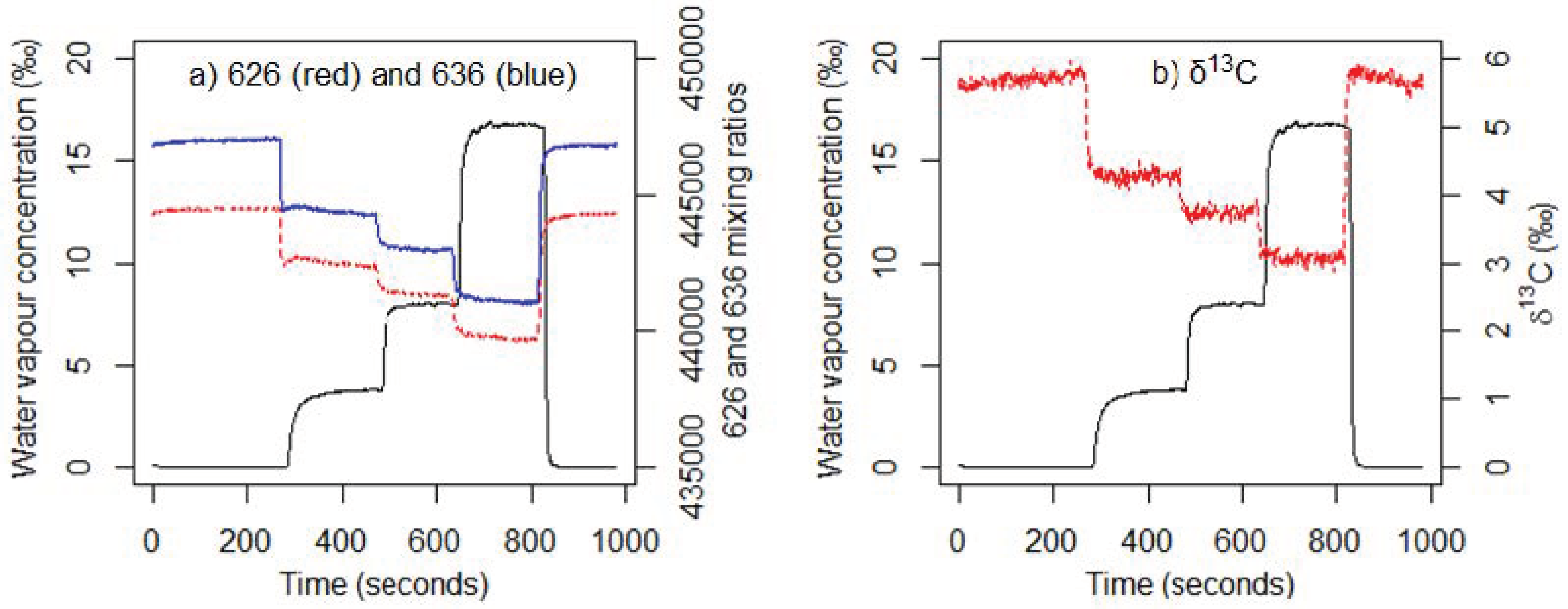
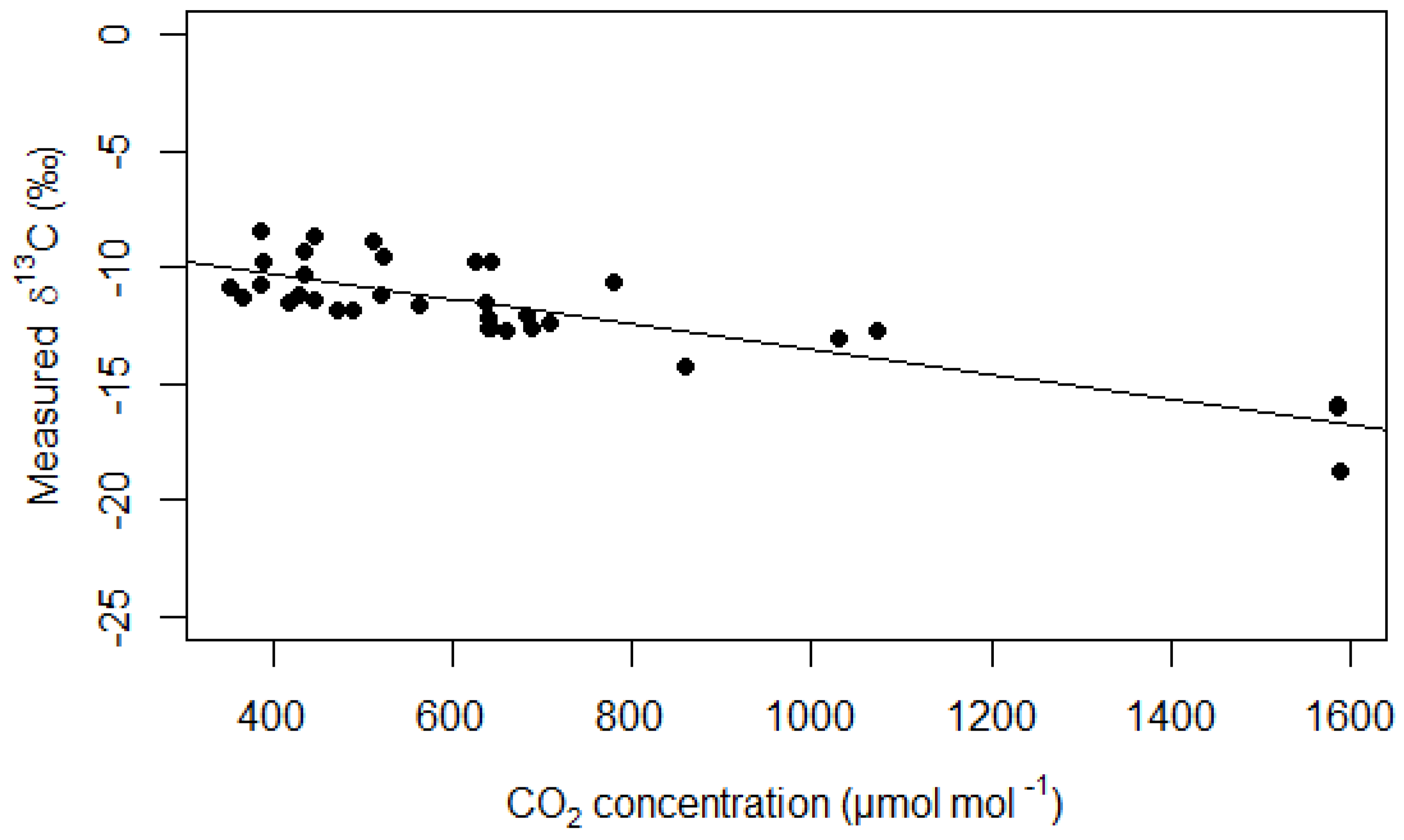
3.1. Water Vapour and CO2 Concentration Dependence
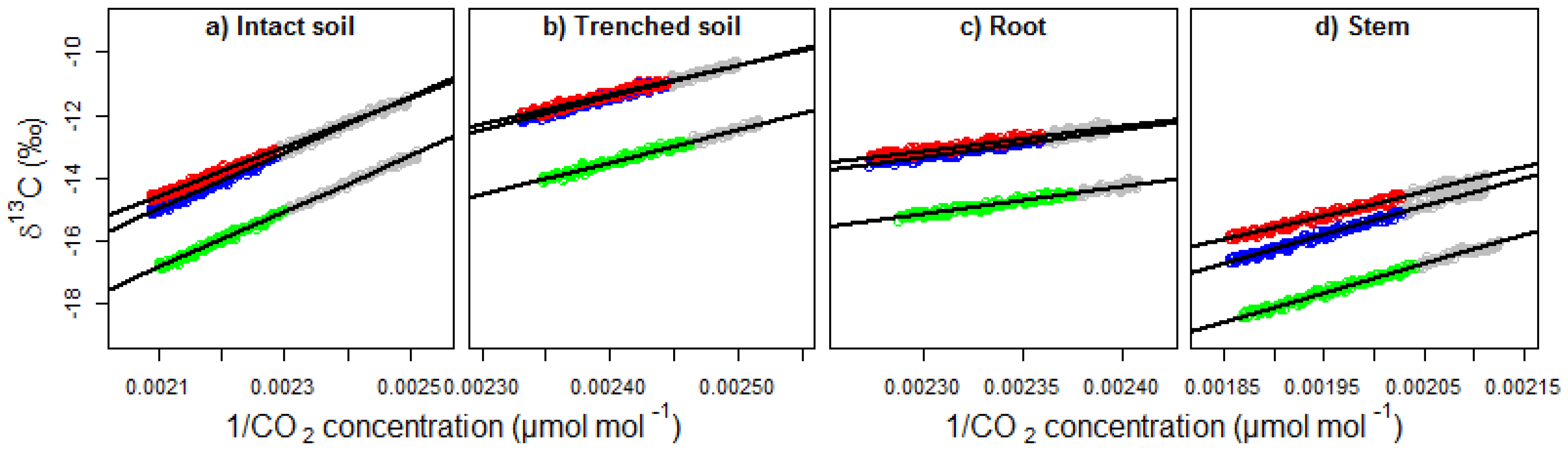
3.2. Effect of the δ13C Corrections on the Keeling Plots
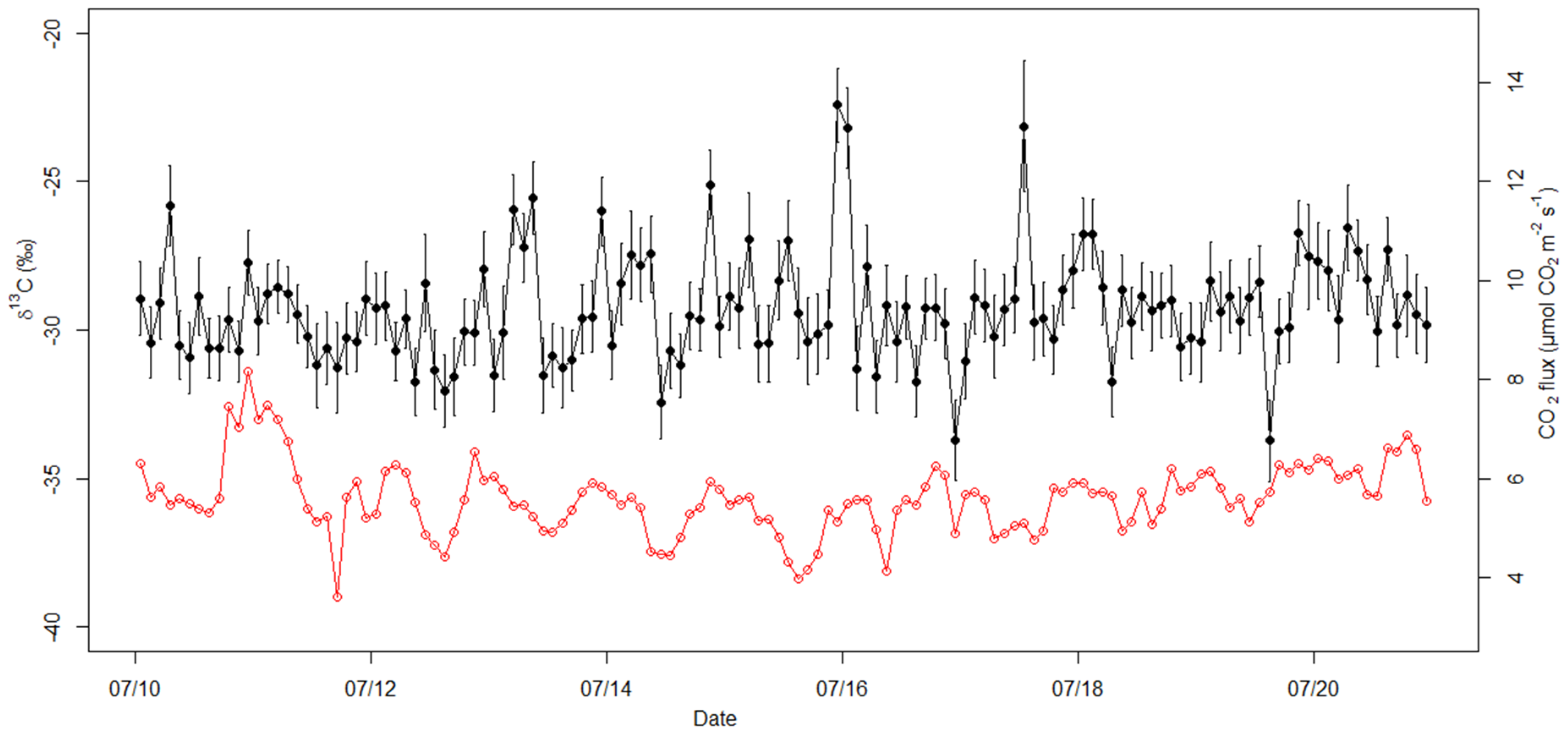
3.3. Automated Chamber Measurement Campaign
4. Discussion
4.1. Effect of the δ13C Corrections
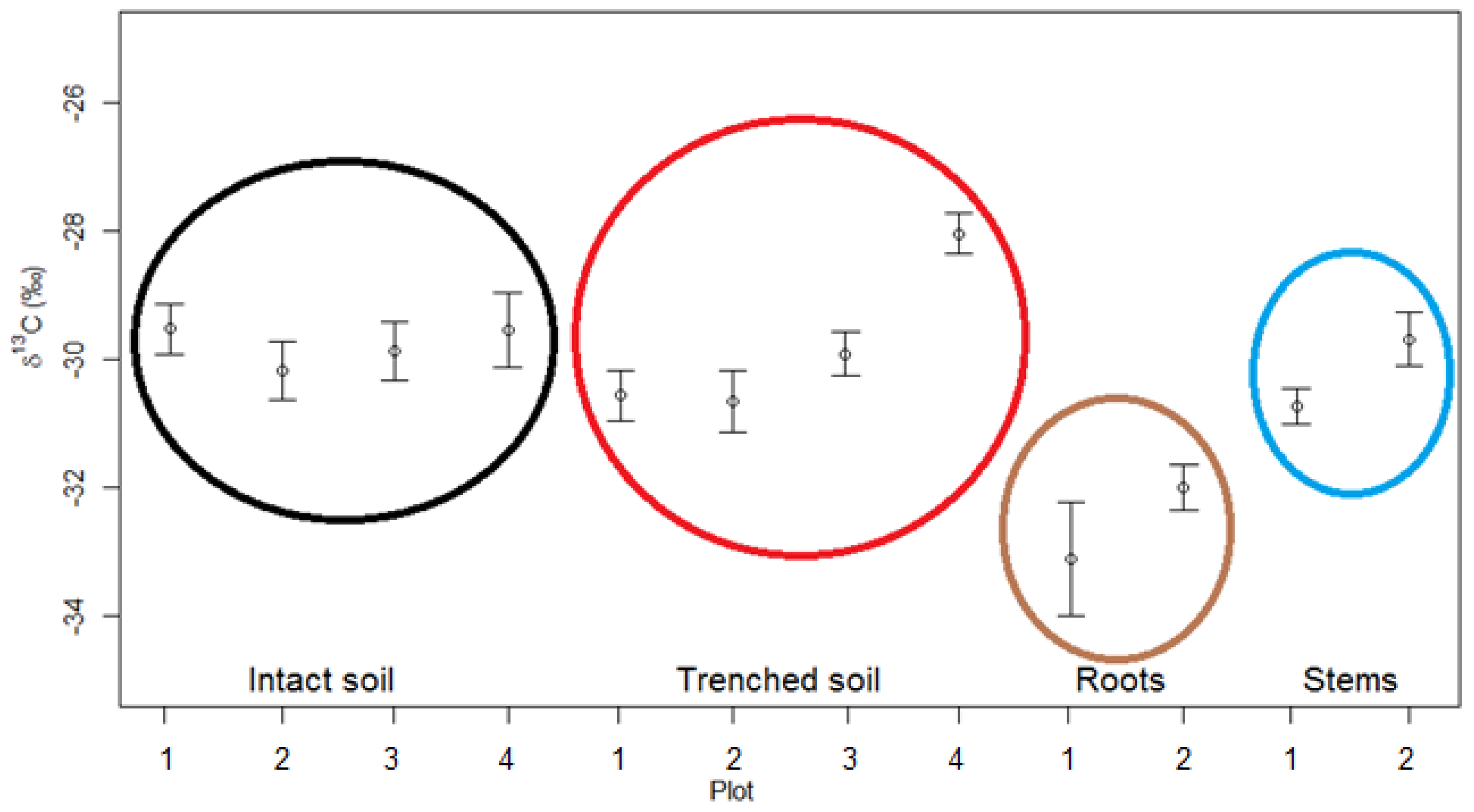
4.2. Overall δ13C Values of the CO2 Fluxes From Intact Soils, Trenched Soils, Roots and Stems
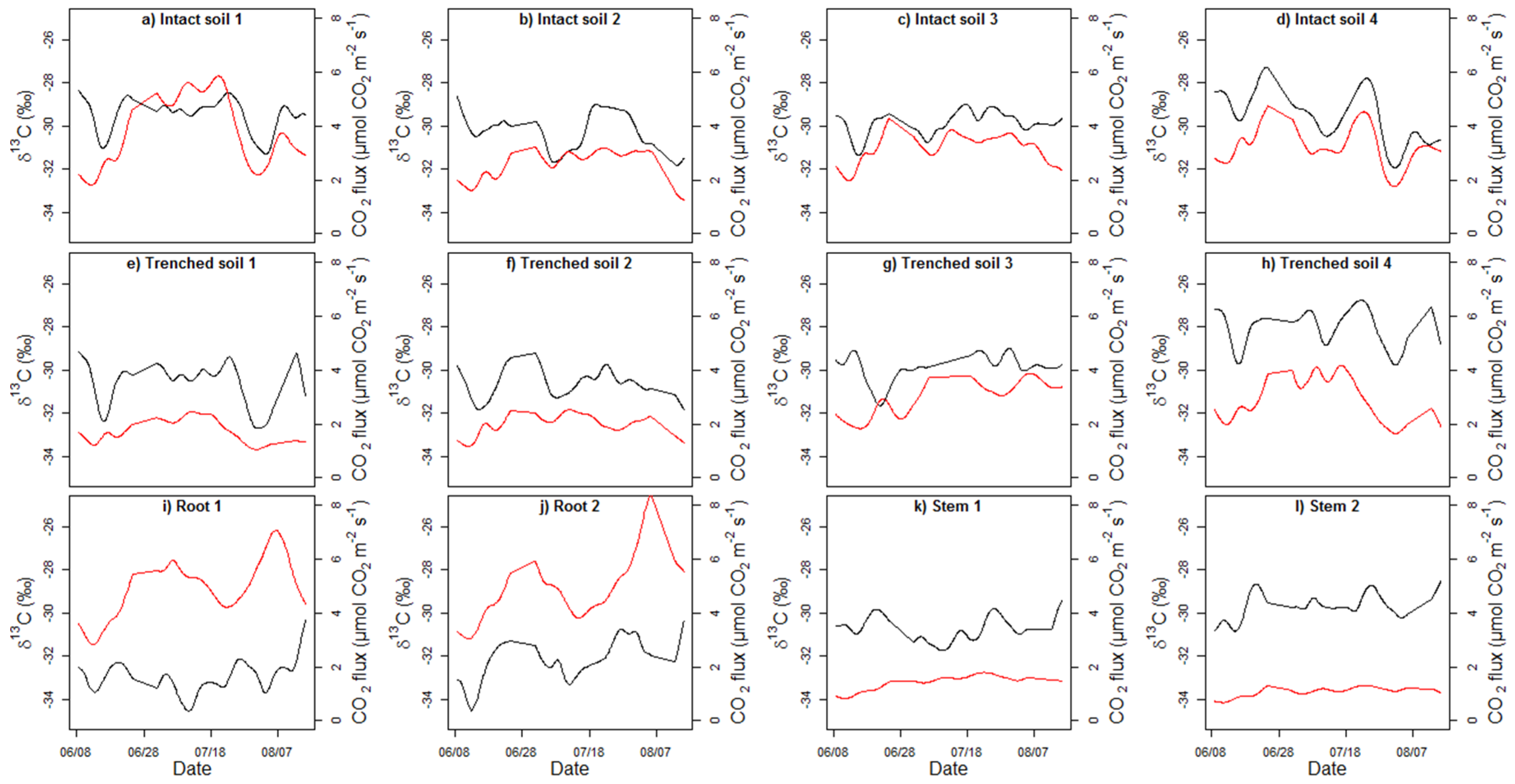
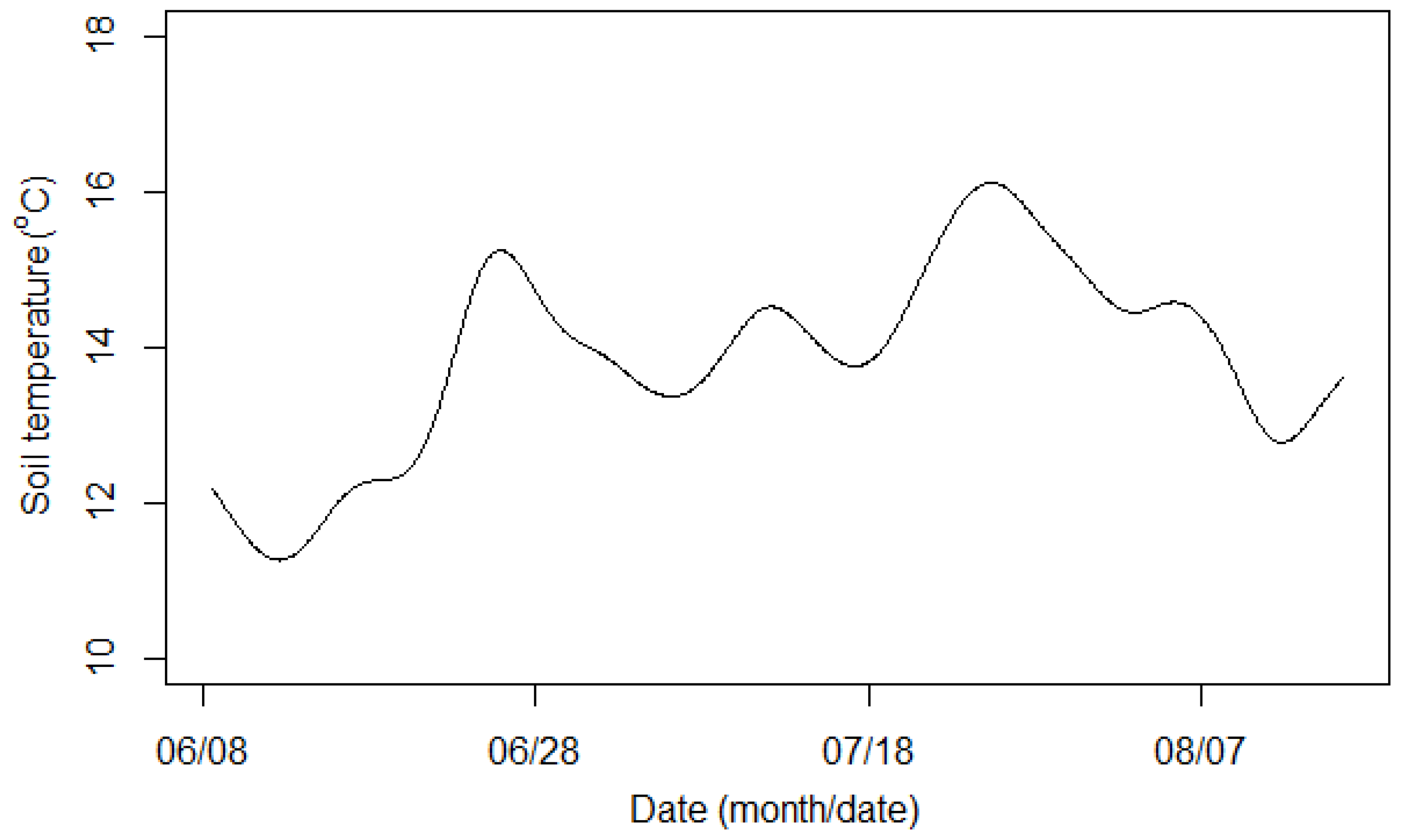
4.3. Seasonality of δ13C and Correlation with Flux and Temperature
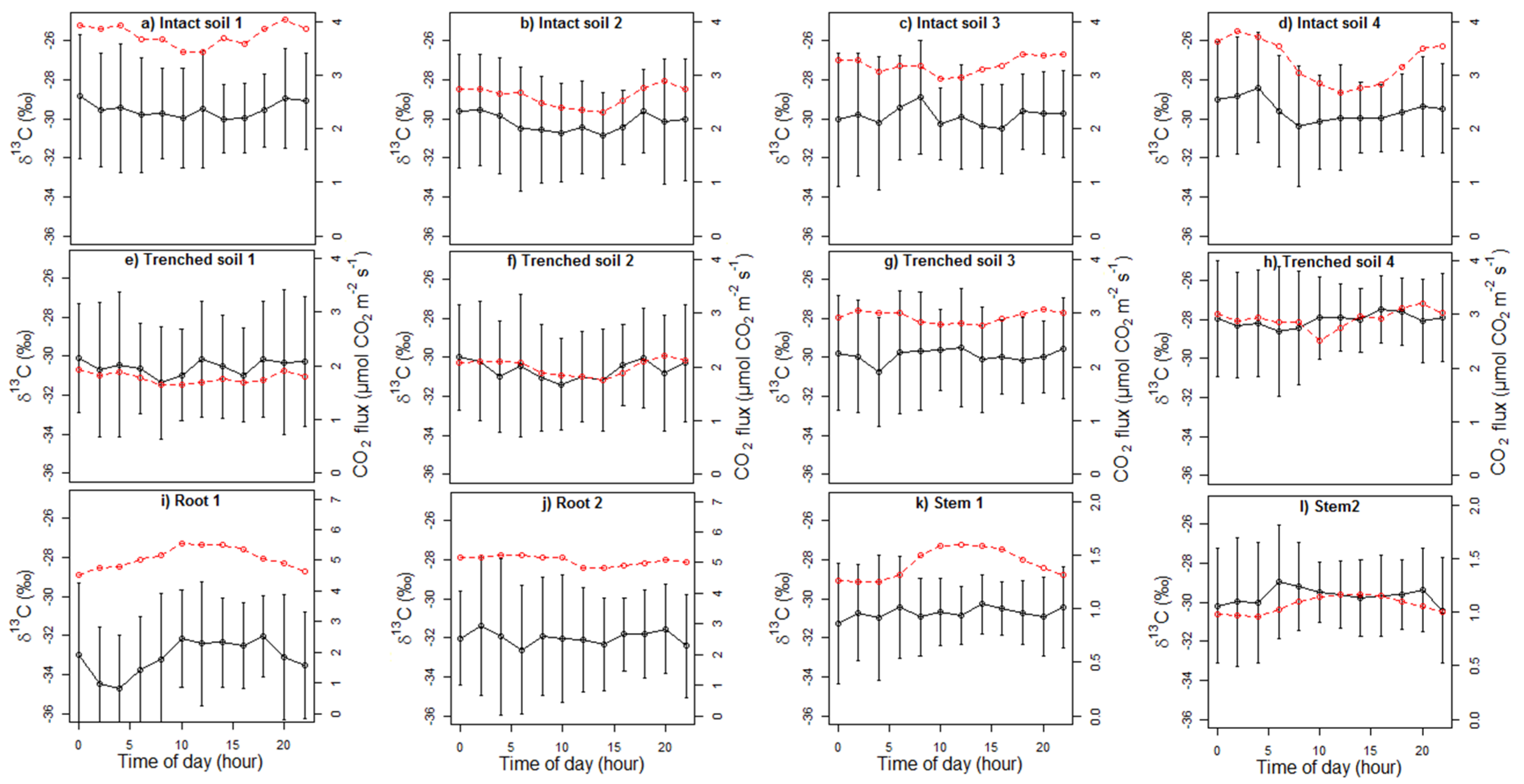
4.4. Diel Patterns of δ13C
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bowling, D.R.; Pataki, D.E.; Randerson, J.T. Carbon Isotopes in Terrestrial Ecosystem Pools and CO2 Fluxes. New Phytol. 2008, 178, 24–40. [Google Scholar] [CrossRef] [PubMed]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Park, R.; Epstein, S. Carbon isotope fractionation during photosynthesis. Geochim. Cosmochim. Acta. 1960, 21, 110–126. [Google Scholar] [CrossRef]
- Janssens, I.A.; Lankreijer, H.; Matteucci, G.; Kowalski, A.S.; Buchmann, N.; Epron, D.; Pilegaard, K.; Kutsch, W.; Longdoz, B.; Grünwald, T.; et al. Productivity overshadows temperature in determining soil and ecosystem respiration across European forests. Glob. Chang. Biol. 2000, 7, 269–278. [Google Scholar] [CrossRef]
- Rochette, P.; Hutchinson, G.L. Measurement of Soil Respiration in situ: Chamber Techniques. In Micrometeorology in Agricultural Systems; Agronomy Monograph 47; Hatfield, J.L., Baker, J.M., Eds.; American Society of Agronomy, Crop Science Society of America and Soil Science Society of America: Madison, WI, USA, 2005; pp. 247–286. [Google Scholar]
- Edwards, N.T.; Riggs, J.S. Automated monitoring of soil respiration: A moving chamber design. Soil Sci. Soc. Am. J. 2003, 67, 1266–1271. [Google Scholar] [CrossRef]
- Koskinen, M.; Minkkinen, K.; Ojanen, P.; Kämäräinen, M.; Laurila, T.; Lohila, A. Measurements of CO2 exchange with an automated chamber system throughout the year: Challenges in measuring night-time respiration on porous peat soil. Biogeosciences 2014, 11, 347–363. [Google Scholar] [CrossRef]
- Liang, N.; Inoue, G.; Fujinuma, Y. A multichannel automated chamber system for continuous measurement of forest soil CO2 efflux. Tree Physiol. 2003, 23, 825–832. [Google Scholar] [CrossRef] [PubMed]
- McGinn, S.; Akinremi, O.; McLean, H.; Ellert, B. An automated chamber system for measuring soil respiration. Can. J. Soil Sci. 1998, 78, 573–579. [Google Scholar] [CrossRef] [Green Version]
- Keeling, C.D. The concentration and isotopic abundances of atmospheric carbon dioxide in rural areas. Geochim. Cosmochim. Acta 1958, 13, 322–334. [Google Scholar] [CrossRef]
- Zeeman, M.J.; Werner, R.A.; Eugster, W.; Siegwolf, R.T.W.; Wehrle, G.; Mohn, J.; Buchmann, N. Optimization of automated gas sample collection and isotope ratio mass spectrometric analysis of δ13C of CO2 in air. Rapid Commun. Mass Spectrom. 2008, 22, 3883–3892. [Google Scholar] [CrossRef]
- Bowling, D.R.; Sargent, S.D.; Tanner, B.D.; Ehleringer, J.R. Tunable diode laser absorption spectroscopy for stable isotope studies of ecosystem–atmosphere CO2 exchange. Agric. For. Meteorol. 2003, 118, 1–19. [Google Scholar] [CrossRef]
- Guimbaud, C.; Noel, C.; Chartier, M.; Catoire, V.; Blessing, M.; Gourry, J.C.; Robert, C. A quantum cascade laser infrared spectrometer for CO2 stable isotope analysis: Field implementation at a hydrocarbon contaminated site under bio-remediation. J. Environ. Sci. 2016, 40, 60–74. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.D.; McManus, J.B.; Herndon, S.C.; Zahniser, M.S.; Tuzson, B.; Emmenegger, L. New method for isotopic ratio measurements of atmospheric carbon dioxide using a 4.3 μm pulsed quantum cascade laser. Appl. Phys. B 2008, 90, 301–309. [Google Scholar] [CrossRef] [Green Version]
- Tuzson, B.; Zeeman, M.J.; Zahniser, M.S.; Emmenegger, L. Quantum cascade laser based spectrometer for in situ stable carbon dioxide isotope measurements. Infrared Phys. Technol. 2008, 51, 198–206. [Google Scholar] [CrossRef]
- Wahl, E.H.; Fidric, B.; Rella, C.W.; Koulikov, S.; Tan, S.; Kachanov, A.A.; Richman, B.A.; Crosson, E.R.; Paldus, B.A.; Kalaskar, S.; et al. Applications of cavity ring-down spectroscopy to high precision isotope ratio measurement of 13C/12C in carbon dioxide. Isotopes Environ. Health Stud. 2006, 42, 20–35. [Google Scholar] [CrossRef] [PubMed]
- McManus, J.B.; Nelson, D.D.; Shorter, J.H.; Jimenez, R.; Herndon, S.; Saleska, S.; Zahniser, M. A high precision pulsed quantum cascade laser spectrometer for measurements of stable isotopes of carbon dioxide. J. Mod. Opt. 2005, 52, 2309–2321. [Google Scholar] [CrossRef]
- Sturm, P.; Eugster, W.; Knohl, A. Eddy covariance measurements of CO2 isotopologues with a quantum cascade laser absorption spectrometer. Agric. For. Meteorol. 2012, 152, 73–82. [Google Scholar] [CrossRef]
- Wehr, R.; Munger, J.W.; Nelson, D.D.; Mcmanus, J.B.; Zahniser, M.S.; Wofsy, S.C.; Saleska, S.R. Long-term eddy covariance measurements of the isotopic composition of the ecosystem–atmosphere exchange of CO2 in a temperate forest. Agric. For. Meteorol. 2013, 181, 69–84. [Google Scholar] [CrossRef]
- Gentsch, L.; Sturm, P.; Hammerle, A.; Siegwolf, R.; Wingate, L.; Ogée, J.; Baur, T.; Plüss, P.; Barthel, M.; Buchmann, N.; et al. Carbon isotope discrimination during branch photosynthesis of Fagus sylvatica: Field measurements using laser spectrometry. J. Exp. Bot. 2014, 65, 1481–1496. [Google Scholar] [CrossRef]
- Kammer, A.; Tuzson, B.; Emmenegger, L.; Knohl, A.; Mohn, J.; Hagedorn, F. Application of a quantum cascade laser-based spectrometer in a closed chamber system for real-time δ13C and δ18O measurements of soil-respired CO2. Agric. For. Meteorol. 2011, 151, 39–48. [Google Scholar] [CrossRef]
- Guillon, S.; Pili, E.; Agrinier, P. Using a laser-based CO2 carbon isotope analyser to investigate gas transfer in geological media. Appl. Phys. B 2012, 107, 449–457. [Google Scholar] [CrossRef]
- Pitt, J.R.; Le Breton, M.; Allen, G.; Percival, C.J.; Gallagher, M.W.; Bauguitte, S.J.-B.; O’Shea, S.J.; Muller, J.B.A.; Zahniser, M.S.; Pyle, J.; et al. The development and evaluation of airborne in situ N2O and CH4 sampling using a quantum cascade laser absorption spectrometer (QCLAS). Atmos. Meas. Tech. 2016, 9, 63–77. [Google Scholar] [CrossRef]
- Wen, X.F.; Meng, Y.; Zhang, X.Y.; Sun, X.M.; Lee, X. Evaluating calibration strategies for isotope ratio infrared spectroscopy for atmospheric 13CO2/12CO2 measurement. Atmos. Meas. Tech. 2013, 6, 1491–1501. [Google Scholar] [CrossRef]
- Griffith, D.W.T. Calibration of isotopologue-specific optical trace gas analysers: A practical guide. Atmos. Meas. Tech. 2018, 11, 6189–6201. [Google Scholar] [CrossRef]
- Sturm, P.; Tuzson, B.; Henne, S.; Emmenegger, L. Tracking isotopic signatures of CO2 at the high altitude site Jungfraujoch with laser spectroscopy: Analytical improvements and representative results. Atmos. Meas. Tech. 2013, 6, 1659–1671. [Google Scholar] [CrossRef]
- Pilegaard, K.; Hummelshøj, P.; Jensen, N.O.; Chen, Z. Two years of continuous CO2 eddy-flux measurements over a Danish beech forest. Agric. For. Meteorol. 2001, 107, 29–41. [Google Scholar] [CrossRef]
- Pilegaard, K.; Ibrom, A.; Courtney, M.S.; Hummelshøj, P.; Jensen, N.O. Increasing net CO2 uptake by a Danish beech forest during the period from 1996 to 2009. Agric. For. Meteorol. 2011, 151, 934–946. [Google Scholar] [CrossRef]
- Brændholt, A.; Larsen, K.S.; Ibrom, A.; Pilegaard, K. Partitioning of ecosystem respiration in a beech forest. Agric. For. Meteorol. 2018, 252, 88–98. [Google Scholar] [CrossRef]
- LI-COR Biosciences. Capturing and Processing Soil GHG Fluxes Using the LI-8100A. 2014. Available online: https://www.licor.com/env/pdf/soil_flux/8100A_AppNote_GHG_Fluxes_15121.pdf (accessed on 15 April 2019).
- Rothman, L.S.; Gordon, I.E.; Babikov, Y.; Barbe, A.; Benner, D.C.; Bernath, P.F.; Birk, M.; Bizzocchi, L.; Boudon, V.; Brown, L.R.; et al. The HITRAN2012 molecular spectroscopic database. J. Quant. Spectrosc. Radiat. Transf. 2013, 130, 4–50. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Brændholt, A.; Larsen, K.S.; Ibrom, A.; Pilegaard, K. Overestimation of closed-chamber soil CO2 effluxes at low atmospheric turbulence. Biogeosciences 2017, 14, 1603–1616. [Google Scholar] [CrossRef]
- Braden-Behrens, J.; Yan, Y.; Knohl, A. A new instrument for stable isotope measurements of 13C and 18O in CO2–instrument performance and ecological application of the Delta Ray IRIS analyzer. Atmos. Meas. Tech. 2017, 10, 4537–4560. [Google Scholar] [CrossRef]
- Pang, J.; Wen, X.; Sun, X.; Huang, K. Intercomparison of two cavity ring-down spectroscopy analyzers for atmospheric 13CO2/12CO2 measurement. Atmos. Meas. Tech. 2016, 9, 3879–3891. [Google Scholar] [CrossRef]
- Vogel, F.R.; Huang, L.; Ernst, D.; Giroux, L.; Racki, S.; Worthy, D.E.J. Evaluation of a cavity ring-down spectrometer for in situ observations of 13CO2. Atmos. Meas. Tech. 2013, 6, 301–308. [Google Scholar] [CrossRef]
- McManus, J.B.; Nelson, D.D.; Zahniser, M.S. Design and performance of a dual-laser instrument for multiple isotopologues of carbon dioxide and water. Opt. Express. 2015, 23, 6518–6569. [Google Scholar] [CrossRef]
- Formánek, P.; Ambus, P. Assessing the use of δ13C natural abundance in separation of root and microbial respiration in a Danish beech (Fagus sylvatica L.) forest. Rapid Commun. Mass Spectrom. 2004, 18, 897–902. [Google Scholar] [CrossRef]
- Wu, J.; Larsen, K.S.; van der Linden, L.; Beier, C.; Pilegaard, K.; Ibrom, A. Synthesis on the carbon budget and cycling in a Danish, temperate deciduous forest. Agric. For. Meteorol. 2013, 181, 94–107. [Google Scholar] [CrossRef]
- Millard, P.; Midwood, A.J.; Hunt, J.E.; Barbour, M.M.; Whitehead, D. Quantifying the contribution of soil organic matter turnover to forest soil respiration, using natural abundance δ13C. Soil Biol. Biochem. 2010, 42, 935–943. [Google Scholar] [CrossRef]
- Sakata, T.; Ishizuka, S.; Takahashi, M. Separation of soil respiration into CO2 emission sources using 13C natural abundance in a deciduous broad-leaved forest in Japan. Soil Sci. Plant Nutr. 2007, 53, 328–336. [Google Scholar] [CrossRef]
- Andersen, C.P.; Nikolov, I.; Nikolova, P.; Matyssek, R.; Häberle, K.H. Estimating “autotrophic” belowground respiration in spruce and beech forests: Decreases following girdling. Eur. J. For. Res. 2005, 124, 155–163. [Google Scholar] [CrossRef]
- Brumme, R. Mechanisms of carbon and nutrient release and retention in beech forest gaps. Plant Soil 1995, 168, 593–600. [Google Scholar] [CrossRef]
- Epron, D.; Le Dantec, V.; Dufrene, E.; Granier, A. Seasonal dynamics of soil carbon dioxide efflux and simulated rhizosphere respiration in a beech forest. Tree Physiol. 2001, 21, 145–152. [Google Scholar] [CrossRef] [Green Version]
- Epron, D.; Farque, L.; Lucot, E.; Badot, P.-M. Soil CO2 efflux in a Beech Forest: The Contribution of Root Respiration. Ann. For. Sci. 1999, 56, 289–295. [Google Scholar] [CrossRef]
- Silver, W.L.; Thompson, A.W.; McGroddy, M.E.; Varner, R.K.; Dias, J.D.; Silva, H.; Crill, P.M.; Keller, M. Fine root dynamics and trace gas fluxes in two lowland tropical forest soils. Glob. Chang. Biol. 2005, 11, 290–306. [Google Scholar] [CrossRef]
- Subke, J.A.; Inglima, I.; Cotrufo, M.F. Trends and methodological impacts in soil CO2 efflux partitioning: A metaanalytical review. Glob. Chang. Biol. 2006, 12, 921–943. [Google Scholar] [CrossRef]
- Salomón, R.; Valbuena-Carabana, M.; Rodriguez-Calcerrada, J.; Aubrey, D.; McGuire, M.A.; Teskey, R.; Gil, L.; Gonzalez-Doncel, I. Xylem and soil CO2 fluxes in a Quercus pyrenaica Willd. coppice: Root respiration increases with clonal size. Ann. For. Sci. 2015, 72, 1065–1078. [Google Scholar] [CrossRef]
- Bekele, A.; Kellman, L.; Beltrami, H. Soil Profile CO2 concentrations in forested and clear cut sites in Nova Scotia, Canada. For. Ecol. Manag. 2007, 242, 587–597. [Google Scholar] [CrossRef]
- Kindler, R.; Siemens, J.; Kaiser, K.; Walmsley, D.C.; Bernhofer, C.; Buchmann, N.; Cellier, P.; Eugster, W.; Gleixner, G.; Grunwald, T.; et al. Dissolved carbon leaching from soil is a crucial component of the net ecosystem carbon balance. Glob. Chang. Biol. 2011, 17, 1167–1185. [Google Scholar] [CrossRef] [Green Version]
- Yavitt, J.B.; Fahey, T.J.; Simmons, J.A. Methane and carbon-dioxide dynamics in a northern hardwood ecosystem. Soil Sci. Soc. Am. J. 1995, 59, 796–804. [Google Scholar] [CrossRef]
- Nickerson, N.; Risk, D. Keeling plots are non-linear in non-steady state diffusive environments. Geophys. Res. Lett. 2009, 36, 6–9. [Google Scholar] [CrossRef]
- Pataki, D.E.; Ehleringer, J.R.; Flanagan, L.B.; Yakir, D.; Bowling, D.R.; Still, C.J.; Buchmann, N.; Kaplan, J.O.; Berry, J.A. The Application and Interpretation of Keeling Plots in Terrestrial Carbon Cycle Research. Glob. Biogeochem. Cycles 2003, 17, 1022. [Google Scholar] [CrossRef]
- Albanito, F.; Mcallister, J.L.; Cescatti, A.; Smith, P.; Robinson, D. Dual-chamber measurements of δ13C of soil-respired CO2 partitioned using a field-based three end-member model. Soil Biol. Biochem. 2012, 47, 106–115. [Google Scholar] [CrossRef]
- Boone, R.D.; Nadelhoffer, K.J.; Canary, J.D.; Kaye, J.P. Roots exert a strong influence on the temperature sensitivity of soil respiration. Nature 1998, 396, 570–572. [Google Scholar] [CrossRef]
- Graham, S.L.; Millard, P.; Hunt, J.E.; Rogers, G.N.D.; Whitehead, D. Roots affect the response of heterotrophic soil respiration to temperature in tussock grass microcosms. Ann. Bot. 2012, 110, 253–258. [Google Scholar] [CrossRef] [Green Version]
- Schnyder, H.; Schäufele, R.; Lötscher, M.; Gebbing, T. Disentangling CO2 fluxes: Direct measurements of mesocosm-scale natural abundance 13CO2/12CO2 gas exchange, 13C discrimination and labelling of CO2 exchange flux components in controlled environments. Plant Cell Environ. 2003, 26, 1863–1874. [Google Scholar] [CrossRef]
- Sun, W.; Resco, V.; Williams, D.G. Environmental and physiological controls on the carbon isotope composition of CO2 respired by leaves and roots of a C3 woody legume (Prosopis velutina) and a C4 perennial grass (Sporobolus wrightii). Plant Cell Environ. 2012, 35, 567–577. [Google Scholar] [CrossRef]
- Maunoury, F.; Berveiller, D.; Lelarge, C.; Pontailler, J.Y.; Vanbostal, L.; Damesin, C. Seasonal, daily and diurnal variations in the stable carbon isotope composition of carbon dioxide respired by tree trunks in a deciduous oak forest. Oecologia 2007, 151, 268–279. [Google Scholar] [CrossRef]
- Teskey, R.O.; Mcguire, M.A. Measurement of stem respiration of sycamore (Platanus occidentalis L.) trees involves internal and external fluxes of CO2 and possible transport of CO2 from roots. Plant Cell Environ. 2007, 30, 570–579. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot Type | Keeling Plot Parameter | Using Measured δ13C | Using δ13C Corrected for Water Vapour | Using δ13C Corrected for Water and CO2 |
|---|---|---|---|---|
| All | Intercept | −35.7 ± 2.9 | −33.6 ± 2.8 | −30.2 ± 2.9 |
| Slope | 9577 ± 1304 | 9588 ± 1325 | 8164 ± 1329 | |
| Intact soil | Intercept | −35.3 ± 2.6 | −33.1 ± 2.6 | −29.8 ± 2.7 |
| Slope | 9412 ± 1176 | 9370 ± 1172 | 7958 ± 1212 | |
| Trenched soil | Intercept | −35.1 ± 2.9 | −32.8 ± 2.8 | −29.7 ± 2.9 |
| Slope | 9272 ± 1277 | 9251 ± 1265 | 7954 ± 1288 | |
| Roots | Intercept | −37.4 ± 3.0 | −35.5 ± 3.0 | −32.6 ± 2.9 |
| Slope | 10246 ± 1415 | 10415 ± 1433 | 9214 ± 1380 | |
| Stems | Intercept | −36.2 ± 2.5 | −34.3 ± 2.5 | −30.2 ± 2.4 |
| Slope | 9901 ± 1261 | 9959 ± 1279 | 8134 ± 1206 |
| Plot | CO2 Flux and δ13C | Soil Temperature and δ13C | ||||
|---|---|---|---|---|---|---|
| Adjusted R2 | Slope ± SE | p-Value | Adjusted R2 | Slope ± SE | p-Value | |
| Intact soil 1 | 0.051 | 0.41 ± 0.071 | <0.001 | −0.00071 | 0.059 ± 0.077 | 0.45 |
| Intact soil 2 | 0.04 | 0.81 ± 0.16 | <0.001 | 0.0071 | 0.18 ± 0.080 | <0.05 |
| Intact soil 3 | 0.043 | 0.66 ± 0.13 | <0.001 | 0.0071 | 0.18 ± 0.075 | <0.05 |
| Intact soil 4 | 0.15 | 0.93 ± 0.085 | <0.001 | 0.0003 | −0.081 ± 0.074 | 0.276 |
| Trenched soil 1 | 0.054 | 1.4 ± 0.26 | <0.001 | −0.002 | −0.010 ± 0.098 | 0.92 |
| Trenched soil 2 | 0.033 | 0.98 ± 0.27 | <0.001 | 0.01 | 0.22 ± 0.089 | <0.05 |
| Trenched soil 3 | −0.0019 | 0.11 ± 0.19 | 0.56 | −0.0012 | 0.096 ± 0.13 | 0.45 |
| Trenched soil 4 | 0.048 | 0.44 ± 0.080 | <0.001 | −0.0013 | 0.038 ± 0.071 | 0.59 |
| Root 1 | 0.04 | 0.42 ± 0.090 | <0.001 | −0.0018 | 0.035 ± 0.11 | 0.74 |
| Root 2 | 0.052 | 0.50 ± 0.10 | <0.001 | 0.023 | 0.37 ± 0.11 | <0.01 |
| Stem 1 | −0.0016 | 0.018 ± 0.27 | 0.95 | −0.0014 | 0.025 ± 0.062 | 0.68 |
| Stem 2 | 0.0056 | 1.0 ± 0.49 | <0.05 | 0.0072 | 0.16 ± 0.071 | <0.05 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brændholt, A.; Ibrom, A.; Ambus, P.; Larsen, K.S.; Pilegaard, K. Combining a Quantum Cascade Laser Spectrometer with an Automated Closed-Chamber System for δ13C Measurements of Forest Soil, Tree Stem and Tree Root CO2 Fluxes. Forests 2019, 10, 432. https://doi.org/10.3390/f10050432
Brændholt A, Ibrom A, Ambus P, Larsen KS, Pilegaard K. Combining a Quantum Cascade Laser Spectrometer with an Automated Closed-Chamber System for δ13C Measurements of Forest Soil, Tree Stem and Tree Root CO2 Fluxes. Forests. 2019; 10(5):432. https://doi.org/10.3390/f10050432
Chicago/Turabian StyleBrændholt, Andreas, Andreas Ibrom, Per Ambus, Klaus Steenberg Larsen, and Kim Pilegaard. 2019. "Combining a Quantum Cascade Laser Spectrometer with an Automated Closed-Chamber System for δ13C Measurements of Forest Soil, Tree Stem and Tree Root CO2 Fluxes" Forests 10, no. 5: 432. https://doi.org/10.3390/f10050432