Sand Dune Height Increases Water Use Efficiency at the Expense of Growth and Leaf Area in Mongolian Pine Growing in Hulunbeier Steppe, Inner Mongolia, China

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
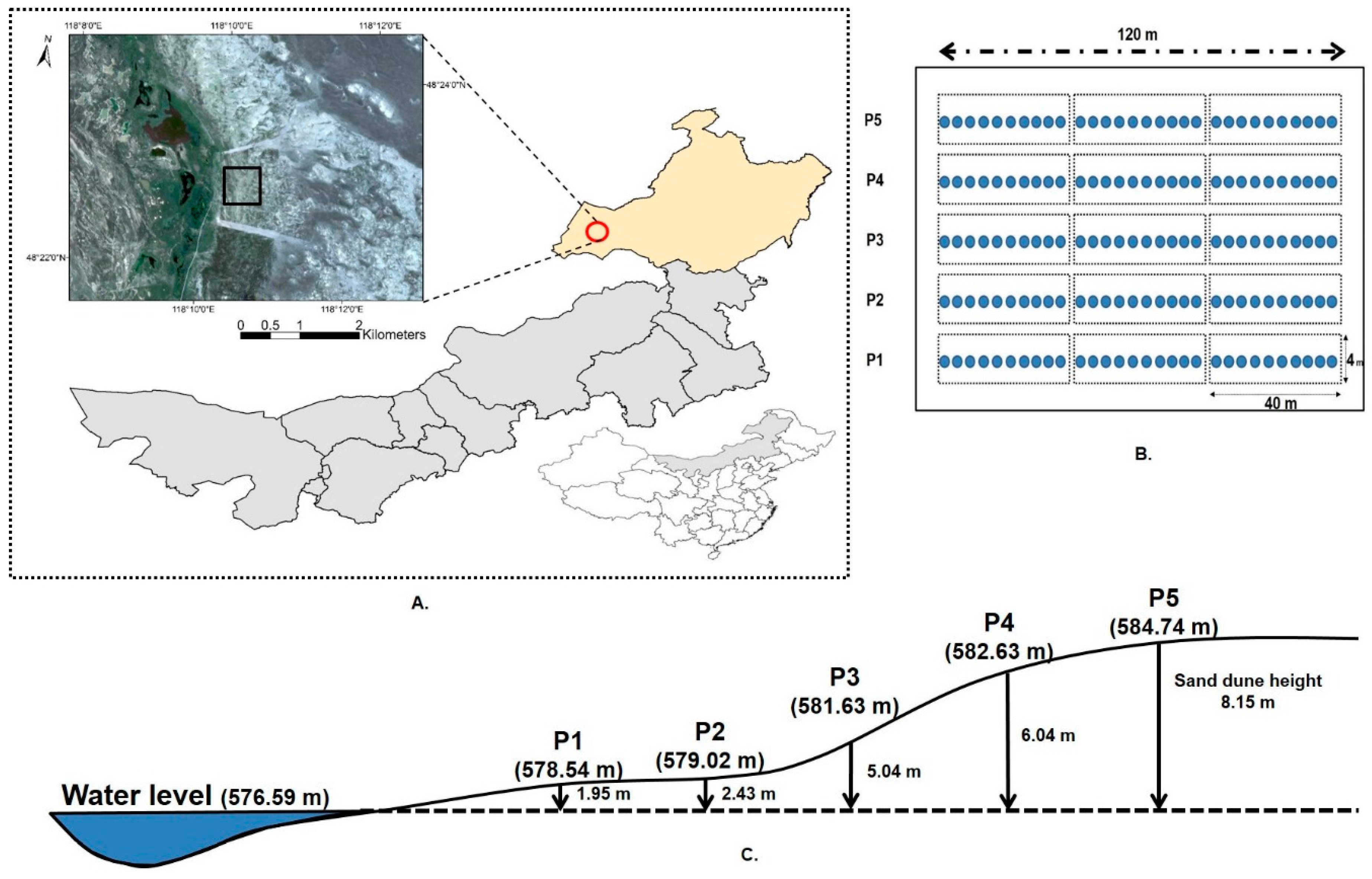
2.1. Study Site and Experimental Design
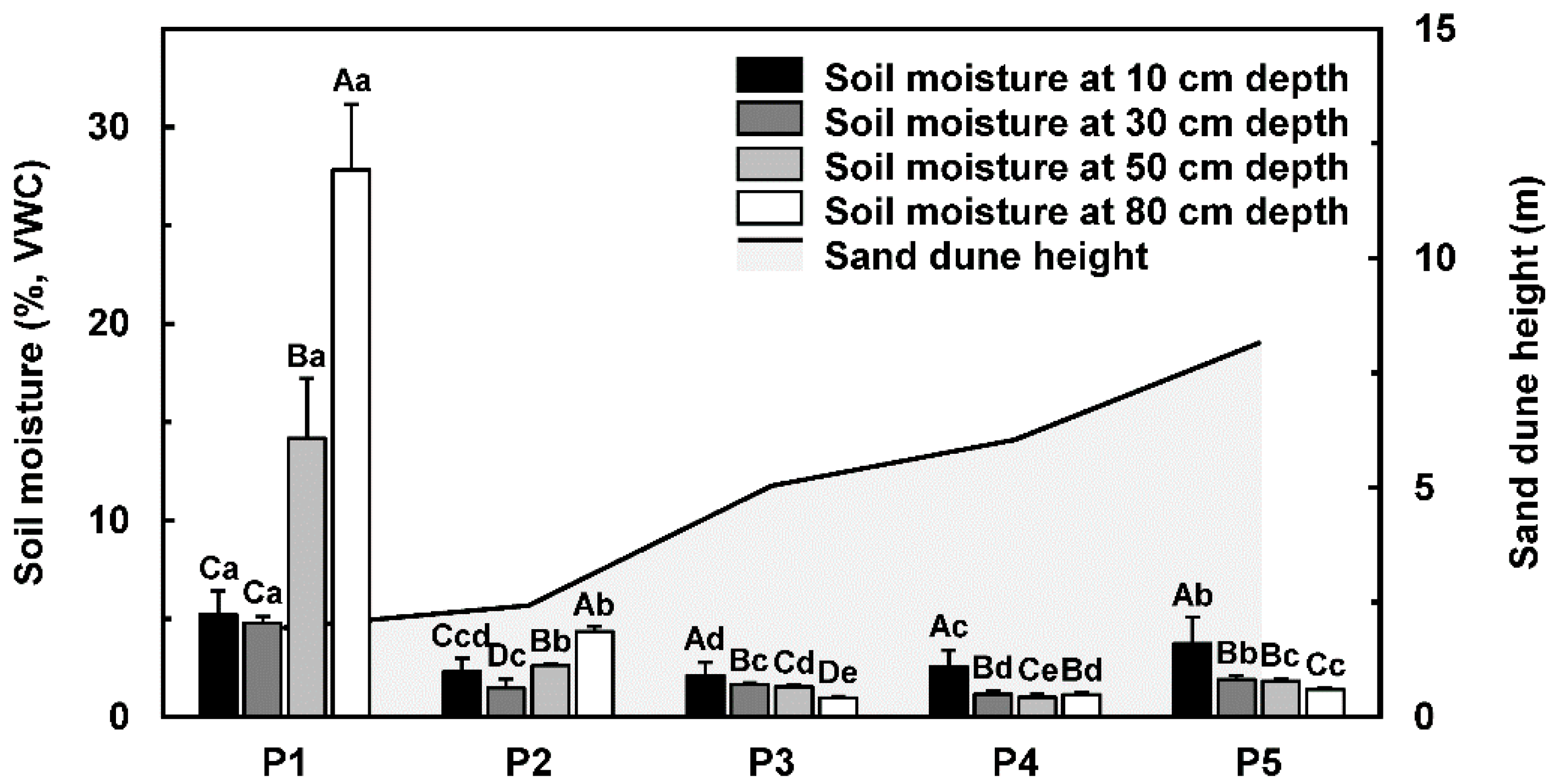
2.2. Soil Moisture
2.3. Survival Rate, Tree Growth, Pine Needle Features, and LMA
2.4. Carbon Isotope Analysis and iWUE
2.5. Statistical Analyses
3. Results
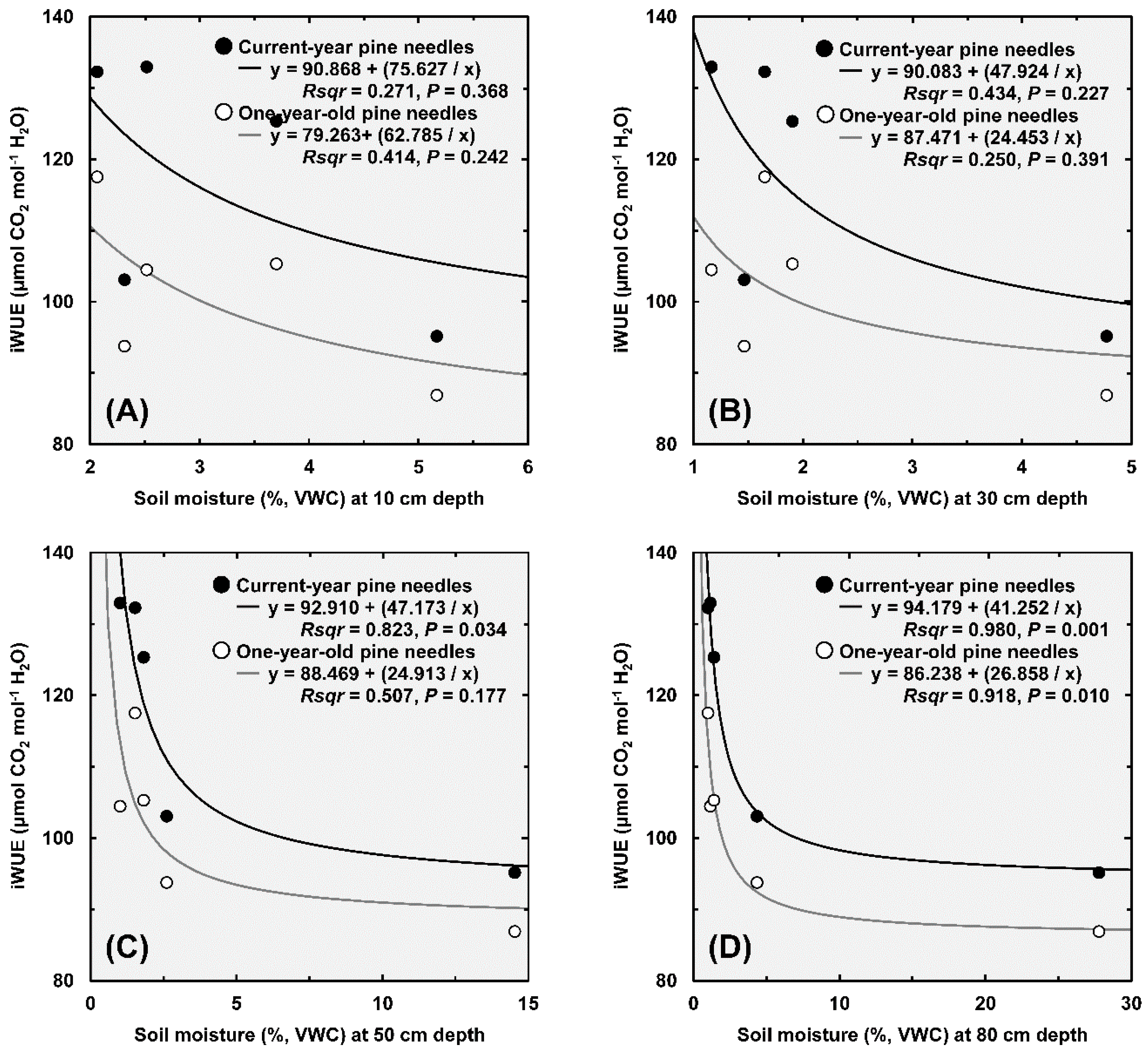
3.1. Soil Moisture
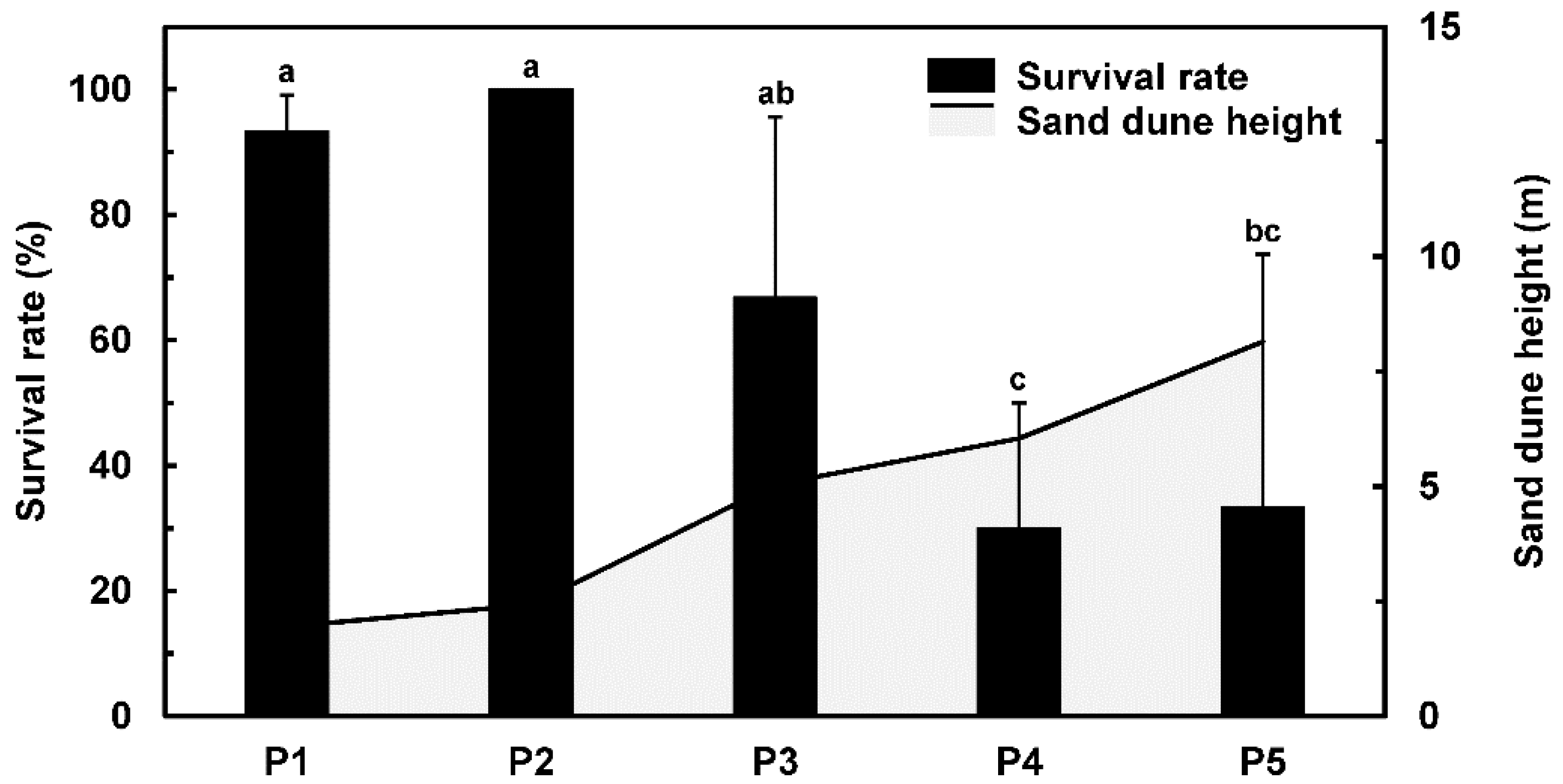
3.2. Survival Rate
3.3. Tree Growth Characteristics
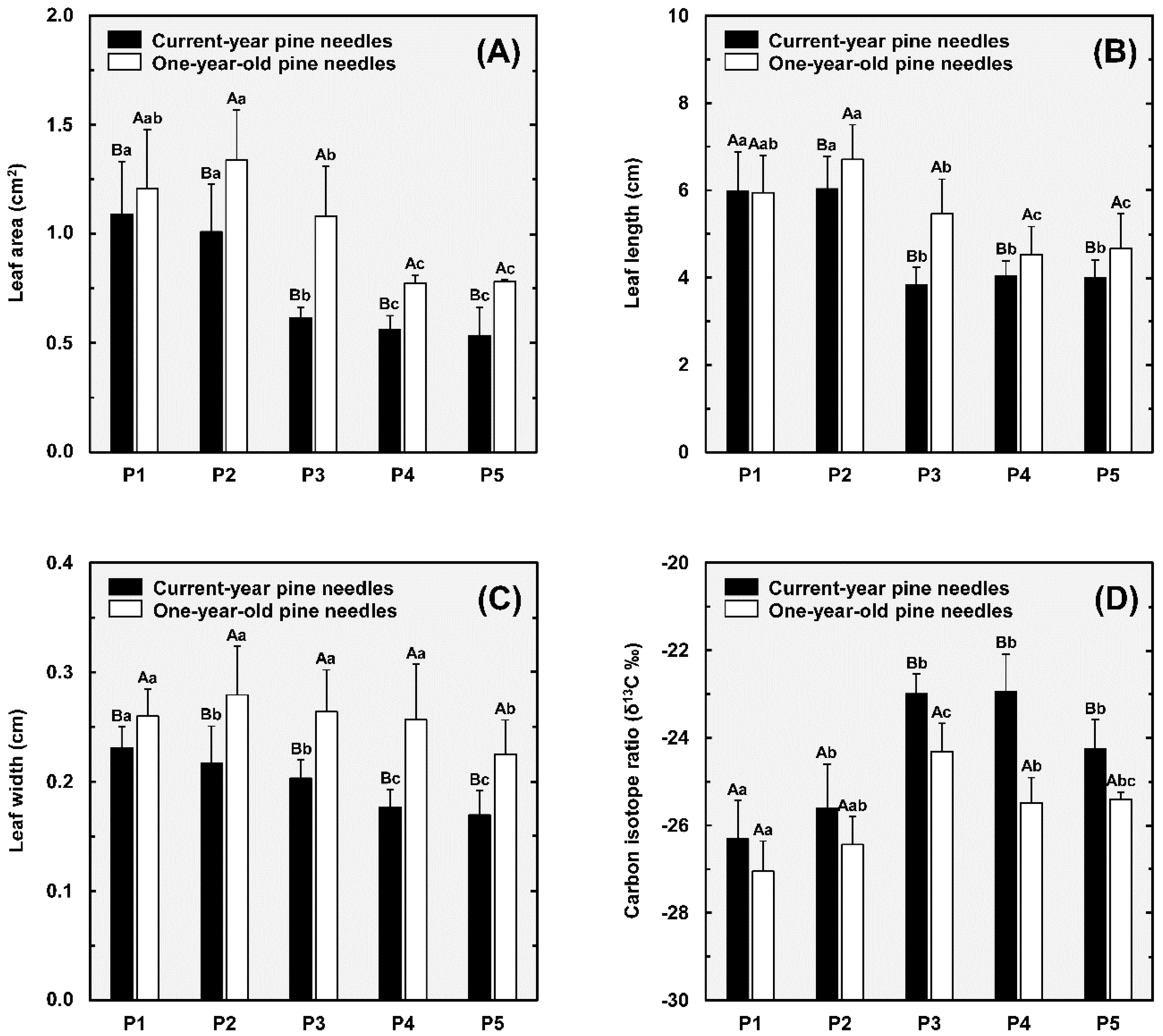
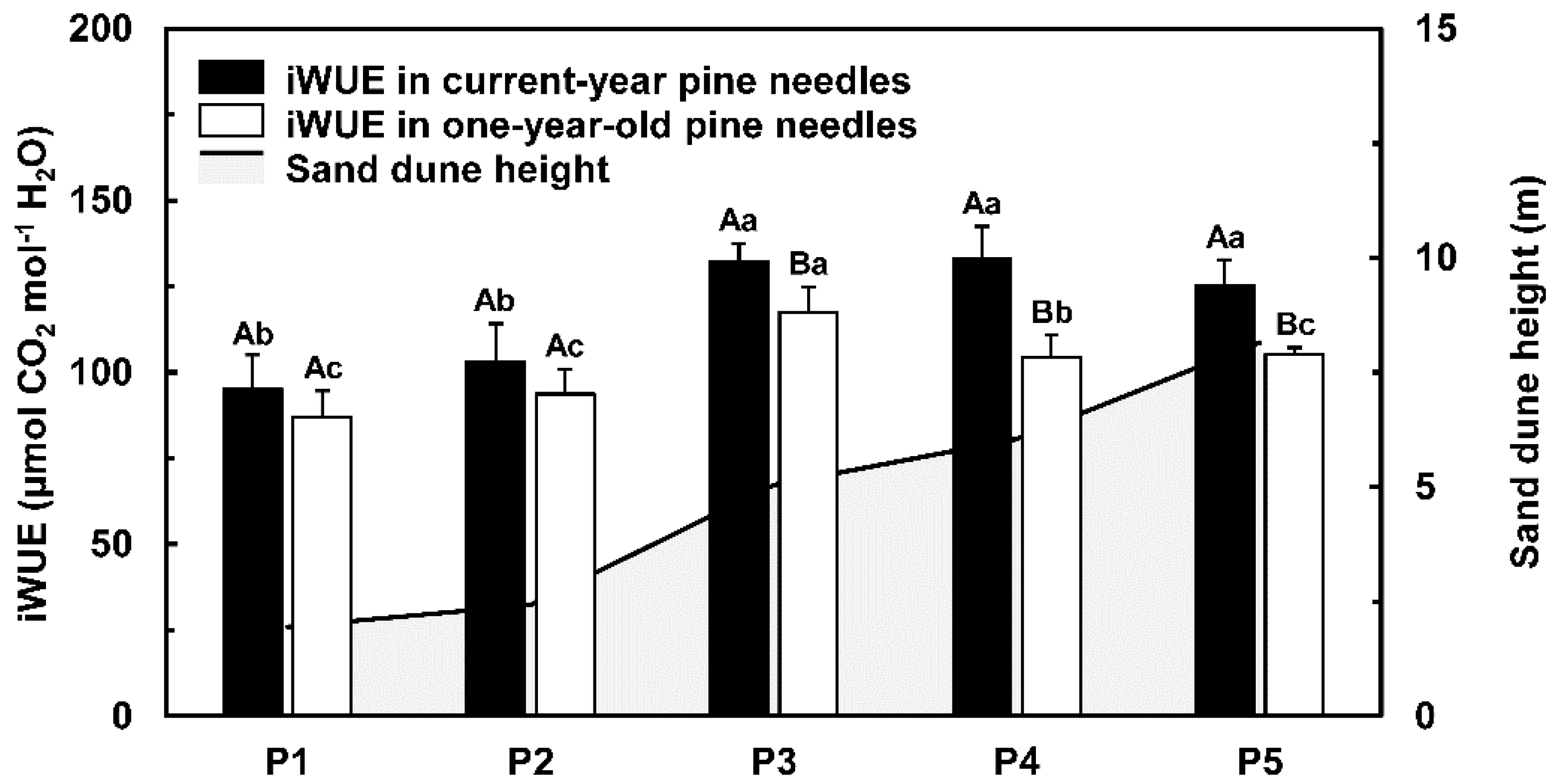
3.4. Pine Needle Characteristics, Carbon Isotope Composition, and iWUE
4. Discussion
4.1. Increased iWUE and Reduced Leaf Area Correlates with Sand Dune Height
4.2. Reduced Survival and Growth of Mongolian Pine Correlates with Sand Dune Height
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- United Nations Convention to Combat Desertification (UNCCD). Report of the Conference of the Parties on Its Thirteenth Session, Held in Ordos, China from 6 to 16 September 2017 Part One: Proceedings; United Nation Convention to Combat Desertification: Ordos, China, 2017; pp. 1–36. [Google Scholar]
- Adeel, Z.; Safriel, U.; Niemeijer, D.; White, R. Millennium Ecosystem Assessment: Ecosystems and Human Well-Being: Desertification Synthesis; World Resource Institute: Washington, DC, USA, 2005; pp. 1–36. [Google Scholar]
- Wang, T.; Wu, W.; Xue, X.; Han, Z.; Zhang, W.; Sun, Q. Spatial-temporal changes of sandy desertified land during last 5 decades in Northern China. Acta. Geogr. Sin. 2004, 59, 203–212. (In Chinese) [Google Scholar]
- Xu, D.; Song, A.; Li, D.; Ding, X.; Wang, Z. Assessing the relative role of climate change and human activities in desertification of North China from 1981 to 2010. Front. Earth Sci. 2019, 13, 43–54. [Google Scholar] [CrossRef]
- Park, K.H.; Qu, Z.Q.; Wan, Q.Q.; Ding, G.D.; Wu, B. Effects of enclosures on vegetation recovery and succession in Hulunbeier steppe, China. For. Sci Technol. 2013, 9, 25–32. [Google Scholar] [CrossRef]
- Lv, S.; Lu, X.; Jin, W. Studies on wind erosion desertification and methods of reversion in Hulunbeier Steppe. J. Arid Land Resour. Environ. 2005, 19, 59–63. (In Chinese) [Google Scholar]
- Bo, T.L.; Fu, L.T.; Zheng, X.J. Modeling the impact of overgrazing on evolution process of grassland desertification. Aeolian Res. 2013, 9, 183–189. [Google Scholar] [CrossRef]
- Zhao, H.; Zhao, Y.; Gao, G.; Ho, P. China’s desertification deconstructed: Towards scientists’ model based on long-term perceptions of grassland restoration (1995–2011). Ecol. Restor. 2018, 26, 26–36. [Google Scholar] [CrossRef]
- Ellenberg, H. Vegetation Ecology of Central Europe; Cambridge University Press: Cambridge, UK, 1988; pp. 193–236. [Google Scholar]
- Zhu, J.J.; Zeng, D.H.; Kang, H.Z.; Wu, X.G.; Fan, Z.P.; Jiang, F.Q.; Li, F.Q.; Wang, H.X.; Xu, H.; Tan, H.; et al. Decline of Pinus Sylvestris Var. Mongolica Plantations on Sandy Land; China Forestry Publisher: Beijing, China, 2005; pp. 1–264. [Google Scholar]
- Zhu, J.; Kang, H.; Tan, H.; Xu, M.; Wang, J. Regeneration characteristics of natural Pinus sylvestris var. mongolica forests on sandy land in Honghuaerji. J. For. Res. 2005, 16, 253–259. [Google Scholar]
- Zhang, X.L.; Guan, T.Y.; Zhou, J.H.; Cai, W.T.; Gao, N.N.; Du, H.; Jiang, L.H.; Lai, L.M.; Zheng, Y.R. Groundwater depth and soil properties are associated with variation in vegetation of a desert riparian ecosystem in an arid area of China. Forests 2018, 9, 34. [Google Scholar] [CrossRef]
- Deng, L.; Wang, K.B.; Li, J.; Shangguan, Z. Effect of soil moisture and atmospheric humidity on both plant productivity and diversity of native grasslands across the loess plateau, China. Ecol. Eng. 2016, 94, 525–531. [Google Scholar] [CrossRef]
- Yu, T.; Feng, Q.; Si, J.; Xi, H.; Li, Z.; Chen, A. Hydraulic redistribution of soil water by roots of two desert riparian phreatophytes in northwest China’s extremely arid region. Plant Soil. 2013, 372, 297–308. [Google Scholar] [CrossRef]
- Lu, Z.; Wei, Y.; Xiao, H.; Zou, S.; Ren, J.; Lyle, C. Trade-offs between midstream agricultural production and downstream ecological sustainability in the Heihe River basin in the past half century. Agric. Water Manag. 2015, 152, 233–242. [Google Scholar] [CrossRef]
- Chen, Y.N.; Pang, Z.; Chen, Y.P.; Li, W.H.; Xu, C.C.; Hao, X.M.; Huang, X.; Huang, T.M.; Ye, Z.X. Response of riparian vegetation to water-table changes in the lower reaches of Tarim River, Xinjiang Uygur, China. Hydrogeol. J. 2008, 16, 1371–1379. [Google Scholar] [CrossRef]
- Hao, X.M.; Li, W.H.; Huang, X.; Zhu, C.G.; Ma, J.X. Assessment of the groundwater threshold of desert riparian forest vegetation along the middle and lower reaches of the Tarim River, China. Hydrol. Process. 2010, 24, 178–186. [Google Scholar] [CrossRef]
- Peng, X.M.; Xiao, S.C.; Xiao, H.L. Preliminary dendrochronological studies on Populus euphratica in the lower reaches of Heihe River basin in northwest China. Dendrochronologia 2013, 31, 242–249. [Google Scholar] [CrossRef]
- Zhu, Y.H.; Chen, Y.N.; Ren, L.L.; Lu, H.S.; Zhao, W.Z.; Yuan, F.; Xu, M. Ecosystem restoration and conservation in the arid inland river basins of Northwest China: Problems and strategies. Ecol. Eng. 2016, 94, 629–637. [Google Scholar] [CrossRef]
- Gries, D.; Zeng, F.; Foetzki, A.; Arndt, S.K.; Bruelheide, H.; Thomas, F.M.; Zhang, X.; Runge, M. Growth and water relations of Tamarix ramosissima and Populus euphratica on Taklamakan desert dunes in relation to depth to a permanent water table. Plant Cell Environ. 2003, 26, 725–736. [Google Scholar] [CrossRef]
- Timofeeva, G.; Treydte, K.; Bugmann, H.; Rigling, A.; Schaub, M.; Siegwolf, R.; Saurer, M. Long-term effects of drought on tree-ring growth and carbon isotope variability in Scots pine in dry environment. Tree Physiol. 2017, 37, 1028–1041. [Google Scholar] [CrossRef] [PubMed]
- Park, G.E.; Lee, D.K.; Kim, K.W.; Batkhuu, N.O.; Tsogtbaatar, J.S.; Zhu, J.J.; Jin, Y.H.; Park, P.S.; Hyun, J.O.; Kim, H.S. Morphological characteristics and water-use efficiency of Siberian elm tree (Ulmus pumila L.) within arid regions of Northeast Asia. Forests 2016, 7, 280. [Google Scholar] [CrossRef]
- Sun, S.J.; Qiu, L.F.; He, C.X.; Li, C.Y.; Zhang, J.S.; Meng, P. Drought-affected Populus simonii Carr. Show lower growth and long-term increases in intrinsic water-use-efficiency prior to tree mortality. Forests 2018, 9, 564. [Google Scholar] [CrossRef]
- Ryan, M.G.; Yoder, B.J. Hydraulic limits to tree height and tree growth. Bioscience 1997, 47, 235–242. [Google Scholar] [CrossRef]
- Mencuccini, M. The ecological significance of long-distance water transport: Short-term regulation, long-term acclimation and the hydraulic costs of stature across plant life forms. Plant Cell Environ. 2003, 26, 163–182. [Google Scholar] [CrossRef]
- Addington, R.N.; Donovan, L.A.; Mitchell, R.J.; Vose, J.M.; Pecot, S.D.; Jack, S.B.; Hacke, U.G.; Sperry, J.S.; Oren, R. Adjustments in hydraulic architecture of Pinus palustris maintain similar stomatal conductance in xeric and mesic habitats. Plant Cell Environ. 2006, 29, 535–545. [Google Scholar] [CrossRef] [PubMed]
- Tanaka-Oda, A.; Kenzo, T.; Koretsune, S.; Sasaki, H.; Fukuda, K. Ontogenetic changes in water-use efficiency (δ13C) and leaf traits differ among tree species growing in a semiarid region of the Loess Plateau, China. For. Ecol. Manag. 2010, 259, 953–957. [Google Scholar] [CrossRef]
- Poorter, H.; Niinemets, U.; Pooter, L.; Wright, I.J.; Villar, R. Causes and consequences of variation in leaf mass per area (LMA): A meta-analysis. New Phytol. 2009, 182, 565–588. [Google Scholar] [CrossRef]
- Farquhar, G.D.; O′Leary, M.H.; Berry, J.A. On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves. Funct. Plant Boil. 1982, 9, 121–137. [Google Scholar] [CrossRef]
- Korol, R.L.; Kirschbaum, M.U.F.; Farquar, G.D.; Jeffreys, M. Effects of water status and soil fertility on the C-isotope signature in Pinus radiate. Tree Physiol. 1999, 19, 551–562. [Google Scholar] [CrossRef]
- Saurer, M.; Siegwolf, R.W.; Schweinfruber, F.H. Carbon isotope discrimination indicates improving water-use efficiency of trees in northern Eurasia over the last 100 years. Glob. Chang. Biol. 2004, 10, 2109–2120. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Richards, R. Isotopic composition of plant carbon correlates with water-use-efficiency of wheat genotypes. Funct. Plant Biol. 1984, 11, 539–552. [Google Scholar] [CrossRef]
- McDowell, N.; Allen, C.D.; Marshall, L. Growth, carbon-isotope discrimination, and drought-associated mortality across a Pinus ponderosa elevational transect. Glob. Chang. Biol. 2010, 16, 399–415. [Google Scholar] [CrossRef]
- Nock, C.A.; Baker, P.J.; Wanek, W.; Leis, A.; Grabner, M.; Bunyavejchewin, S.; Hietz, P. Long-term increases in intrinsic water-use efficiency do not lead to increased stem growth in a tropical monsoon forest in western Thailand. Glob. Chang. Biol. 2011, 17, 1049–1063. [Google Scholar] [CrossRef]
- Levesque, M.; Siegwolf, R.; Saurer, M.; Eilmann, B.; Rigling, A. Increased water-use efficiency does not lead to enhanced tree growth under xeric cinditions. New Phytol. 2014, 203, 94–109. [Google Scholar] [CrossRef] [PubMed]
- Beerling, D.J. Carbon isotope discrimination and stomatal responses of mature Pinus sylvestris L. trees exposed in situ for three years to elevated CO2 and temperature. Acta Oecol. 1997, 18, 697–712. [Google Scholar] [CrossRef]
- Fu, A.H.; Chen, Y.N.; Li, W.H. Water use strategies of the desert riparian forest plant community in the lower reaches of Heihe River Basin, China. Sci. China Earth Sci. 2014, 57, 1293–1305. [Google Scholar] [CrossRef]
- Brodribb, T. Dynamics of changing intercellular CO2 concentration during drought and determination of minimum functional Ci. Plant Physiol. 2006, 111, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Delucia, E.H.; Maherali, H.; Carey, E.V. Climate-driven changes in biomass allocation in pines. Glob. Chang. Biol. 2000, 6, 587–593. [Google Scholar] [CrossRef]
- Greenwood, S.; Ruiz-Benito, P.; Martínez-Vilalta, J.; Lloret, F.; Kitzberger, T.; Allen, C.D.; Fensham, R.; Laughlin, D.C.; Kattge, J.; Bönisch, G.; et al. Tree mortality across biomes is promoted by drought intensity, lower wood density and higher specific leaf area. Ecol. Lett. 2017, 20, 539–553. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Cerrillo, R.; Rodriguez-Vallejo, C.; Silveiro, E.; Hortal, A.; Palacios-Rodríguez, G.; Duque-Lazo, J.; Camarero, J. Cumulative drought stress leads to a loss of growth resilience and explains higher mortality in planted than in naturally regenerated Pinus pinaster stands. Forests 2018, 9, 358. [Google Scholar] [CrossRef]
- Giuggiola, A.; Zweifel, R.; Feichtinger, L.M.; Vollenwider, P.; Bugmann, H.; Haeni, M.; Rigling, A. Competition for water in a xeric forest ecosystem—Effects of understory removal on soil micro-climate, growth and physiology of dominant Scots pine trees. For. Ecol. Manag. 2018, 409, 241–249. [Google Scholar] [CrossRef]
- Oberhuber, W.; Stumbock, M.; Kofler, W. Climate-tree-growth relationships of Scots pine stands (Pinus sylvestris L.) exposed to soil dryness. Trees 1998, 13, 19–27. [Google Scholar] [CrossRef]
- Levanic, T.; Cater, M.; McDowell, N.G. Associations between growth, wood anatomy, carbon isotope discrimination and mortality in Quercus robur forest. Tree Physiol. 2011, 31, 298–308. [Google Scholar] [CrossRef]
- Moran-Lopez, T.; Poyatos, R.; Llorens, P.; Sabate, S. Effects of past growth trends and current water use strategies on Scots pine and pubescent oak drought sensitivity. Eur. J. For. Res. 2014, 133, 369–382. [Google Scholar] [CrossRef]
- Hentschel, R.; Rosner, S.; Kayler, Z.E.; Andreassen, K.; Børja, I.; Solberg, S.; Tveito, O.E.; Priesack, E.; Gessler, A. Norway spruce physiological and anatomical predisposition to dieback. For. Ecol. Manag. 2014, 322, 27–36. [Google Scholar] [CrossRef]
- Watt, M.S.; Whitehead, D.; Mason, E.G.; Richardson, B.; Kimberley, M.O. The influence of weed competition for light and water on growth and dry matter partitioning of young Pinus radiate at a dryland site. For. Ecol. Manag. 2003, 183, 363–376. [Google Scholar] [CrossRef]
- Newton, M.; Preest, D.S. Growth and water relations of Douglas fir (Pesudotsuga menziesii) seedlings under different weed control regimes. Weed Sci. 1988, 36, 653–662. [Google Scholar] [CrossRef]
- Powers, R.; Reynolds, P.E. Ten-year responses of ponderosa pine plantations to repeated vegetation and nutrient control along an environmental gradient. Can. J. For. Res. 1999, 29, 1027–1038. [Google Scholar] [CrossRef]
- Mason, E.G.; Rose, R.; Rosner, L.S. Time vs. light: A potentially useable light sum hybrid model to represent the juvenile growth of Douglas-far subject to varying levels of competition. Can. J. For. Res. 2007, 37, 795–805. [Google Scholar] [CrossRef]
- Dinger, E.J.; Rose, R. The integration of soil moisture, xylem water potential, and fall-spring herbicide treatments to achieve the maximum growth response in newly planted Douglas-fir seedlings. Can. J. For. Res. 2009, 39, 1401–1414. [Google Scholar] [CrossRef]
- Dinger, E.J.; Rose, R. Initial autumn-spring vegetation management regimes improve moisture conditions and maximize third-year Douglas-fir seedling growth in a Pacific Northwest plantation. N. Z. J. For. Sci. 2010, 40, 93–108. [Google Scholar]
- Maguire, D.A.; Mainwaring, D.B.; Rose, R.; Garber, S.M.; Dinger, E.J. Response of coastal Douglas-fir and competing vegetation to repeated and delayed weed control treatments during early plantation development. Can. J. For. Res. 2009, 39, 1208–1219. [Google Scholar] [CrossRef] [Green Version]
- Pinto, J.R.; Marshall, J.D.; Dumroese, R.K.; Davis, A.S.; Cobos, D.R. Photosynthetic response, carbon isotopic composition, survival, and growth of three stock types under water stress enhanced by vegetative competition. Can. J. For. Res. 2012, 42, 333–344. [Google Scholar] [CrossRef]
- Carlos, A.G.B.; Eric, J.D. Use of water stress integral to evaluate relationships between soil moisture, plant water stress and stand productivity in young Douglas-fir trees. New For. 2018, 49, 775–789. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Point | 1 | 2 | 3 | 4 | 5 |
|---|---|---|---|---|---|
| Tree height (m) | 3.45 ± 0.27 a | 3.77 ± 0.12 a | 2.17 ± 1.26 b | 1.73 ± 0.38 b | 1.82 ± 0.47 b |
| DRC (cm) | 8.81 ± 0.66 a | 9.48 ± 0.18 a | 6.52 ± 2.78 b | 4.75 ± 0.93 c | 5.24 ± 1.54 bc |
| DBH (cm) | 6.14 ± 0.35 ab | 6.79 ± 0.41 a | 3.64 ± 2.72 b | 2.09 ± 0.75 c | 1.98 ± 0.94 c |
| Shoot length (cm) | 18.91 ± 3.48 a | 19.63 ± 3.69 a | 14.12 ± 2.93 a | 7.87 ± 4.13 b | 6.21 ± 2.32 b |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, C.-B.; Kim, Y.S.; Choi, H.T.; Kim, J.; Kim, S.; Cha, S.; Gao, G.-L.; Bao, Y.-F.; Son, Y.; Kwon, J.; et al. Sand Dune Height Increases Water Use Efficiency at the Expense of Growth and Leaf Area in Mongolian Pine Growing in Hulunbeier Steppe, Inner Mongolia, China. Forests 2019, 10, 558. https://doi.org/10.3390/f10070558
Kim C-B, Kim YS, Choi HT, Kim J, Kim S, Cha S, Gao G-L, Bao Y-F, Son Y, Kwon J, et al. Sand Dune Height Increases Water Use Efficiency at the Expense of Growth and Leaf Area in Mongolian Pine Growing in Hulunbeier Steppe, Inner Mongolia, China. Forests. 2019; 10(7):558. https://doi.org/10.3390/f10070558
Chicago/Turabian StyleKim, Chan-Beom, Yong Suk Kim, Hyung Tae Choi, Jeonghwan Kim, Seongjun Kim, Sangsub Cha, Guang-Lei Gao, Yan-Feng Bao, Yowhan Son, Jino Kwon, and et al. 2019. "Sand Dune Height Increases Water Use Efficiency at the Expense of Growth and Leaf Area in Mongolian Pine Growing in Hulunbeier Steppe, Inner Mongolia, China" Forests 10, no. 7: 558. https://doi.org/10.3390/f10070558