The Role of Traditional Livelihood Practices and Local Ethnobotanical Knowledge in Mitigating Chestnut Disease and Pest Severity in Turkey

 , ,
, ,
Abstract
:1. Introduction
Human Niche Construction, Traditional Ecological Maintenance, and Forest Policy
2. Materials and Methods
2.1. Study Context
2.2. Fieldwork
2.3. Data Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lewontin, R. Organism and Environment. In Learning, Development and Culture; Plotkin, E.C., Ed.; Wiley: Hoboken, NJ, USA, 1982; pp. 151–170. [Google Scholar]
- Lewontin, R. Gene, organism and environment. In Evolution from Molecules to Men; Bendall, D.S., Ed.; Cambridge University Press: Cambridge, UK, 1983; pp. 273–285. [Google Scholar]
- Lewontin, R. The Triple Helix: Gene, Organism and Environment; Harvard University Press: Cambridge, MA, USA, 2001. [Google Scholar]
- Smith, B.D.; Zeder, M.A. The onset of the Anthropocene. Anthropocene 2013, 4, 8–13. [Google Scholar] [CrossRef]
- Villani, F.; Sansotta, A.; Cherubini, M.; Cesaroni, D.; Sbordoni, V. Genetic structure of natural populations of Castanea sativa in Turkey: Evidence of a hybrid zone. J. Evol. Biol. 2000, 12, 233–244. [Google Scholar] [CrossRef]
- Mattioni, C.; Martin, M.A.; Cherubini, M.; Taurchini, D.; Villani, F. Genetic Diversity in European Chestnut Populations. Actahort 2010, 866, 163–168. [Google Scholar] [CrossRef]
- Erdem, R. Causes of Die-Back of Chestnut Trees in Turkey and Control of Disease. Minist. Agric. For. Gen. Dir. 1951, 102, 82. [Google Scholar]
- Akdogan, S.; Erkman, E. Dikkat kestane kasseri görüldü. Tomurcuk 1968, 1, 4–5. [Google Scholar]
- Cetin, G.; Orman, E.; Polat, Z. First record of the Oriental gall wasp, Dryocosmus kuriphilus Yasumatsu (Hymenoptera: Cynipidae) in Turkey. Bitki Koruma Bul. 2014, 54, 303–309. [Google Scholar]
- TNGDF. Kestane Eylem Plani 2013-2017. Available online: http://www.cevresehirkutuphanesi.com/basili-yayinlar/detay/kestane-eylem-plani-2013-2017 (accessed on 25 June 2019).
- Odling-smee, J.; Laland, K.N.; Feldman, W. Niche Construction: Monographs in Population Biology, no. 37; Princeton University Press: Princeton, NJ, USA, 2003. [Google Scholar]
- Crutzen, P. The “Anthropocene.” In Earth System Science in the Anthropocene; Springer: Berlin/Heidelberg, Germany, 2006; pp. 13–18. [Google Scholar]
- Soule, M.; Wilcox, B. Conservation Biology: An Evolutionary-Ecological Perspective; Sinaur: Sunderland, MA, USA, 1980. [Google Scholar]
- Dasmann, R. A Different Kind of Country; Collier Books: New York, NY, USA, 1968. [Google Scholar]
- Brush, S.B.S. In Situ Conservation of Landraces in Centers of Crop Diversity. Crop Sci. 1995, 35, 346–354. [Google Scholar] [CrossRef]
- Food and Agricultural Organization. Survey of Crop Genetic Resources in Their Centres of Diversity; Food and Agricultural Organization: Rome, Italy, 1973. [Google Scholar]
- Harlan, J. Crops and Man; American Society of Agronomy: Madison, WI, USA, 1992. [Google Scholar]
- Jarvis, D.I.; Brown, A.H.D.; Cuong, P.H.; Collado-Panduro, L.; Latournerie-Moreno, L.; Gyawali, S.; Tanto, T.; Sawadogo, M.; Mar, I.; Sadiki, M.; et al. A global perspective of the richness and evenness of traditional crop-variety diversity maintained by farming communities. Proc. Natl. Acad. Sci. USA 2008, 105, 5326–5331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vavilov, N.I. Studies on the Origin of Cultivated Plants. Bull. Appl. Bot. Plant Breed. 1926, 1–248. [Google Scholar]
- Allard, R.W. Relationship Between Genetic Diversity and Consistency of Performance in Different Environments. Crop Sci. 1961, 1, 127–133. [Google Scholar] [CrossRef]
- Smithson, J.; Lenne, J. Varietal mixtures: A viable strategy for sustainable productivity in subsistence agriculture. Ann. Appl. Biol. 1996, 128, 127–158. [Google Scholar] [CrossRef]
- Clawson, D.L. Harvest Security and Intraspecific Diversity in Traditional Tropical Agriculture. Econ. Bot. 1985, 39, 56–67. [Google Scholar] [CrossRef]
- Tuxill, J.; Reyes, L.A.; Moreno, L.L.; Uicab, V.C.; Jarvis, D.I. All Maize Is Not Equal: Maize Variety Choices and Mayan Foodways in Rural Yucatan, Mexico. In Pre-Columbian Foodways: Interdiscipilanary Approaches to Food, Culture, and Markets in Ancient Mesoamerica; Staller, J., Carrasco, M., Eds.; Springer New York: New York, NY, USA, 2010; pp. 467–486. ISBN 978-1-4419-0470-6. [Google Scholar]
- Ehlers, J.; Hall, A. Cowpea (Vigna unguiculata L. Walp.). F. Crop. Res. 1997, 53, 187–204. [Google Scholar] [CrossRef]
- Asfaw, Z. The barleys of Ethiopia. In Genes in the Field: On-Farm Conservation of Crop Diversity; Brush, S.B., Ed.; International Plant Genetic Resources Institute: Ottawa, ON, Canada, 2000; pp. 77–108. [Google Scholar]
- Ferguson, A.E.; Mkandawire, R.M. Common Beans and Farmer Managed Diversity: Regional Variations in Malawi. Cult. Agric. 1993, 13, 14–17. [Google Scholar] [CrossRef]
- Smith, B.D. Resource Resilience, Human Niche Construction, and the Long- Term Sustainability of Pre-Columbian Subsistence Economies in the Mississippi River Valley Corridor Your use of this PDF, the BioOne Web site, and all posted and associated content PRE-COLUM. J. Ethnobiol. 2009, 29, 167–183. [Google Scholar] [CrossRef]
- Rostlund, E. The Myth of a Natural Prairie Belt in Alabama: An Interpretation of Historical Records. Ann. Assoc. Am. Geogr. 1957, 47, 392–411. [Google Scholar] [CrossRef]
- Anderson, K.; Blackburn, T.C. Before the Wilderness: Environmental Management by Native Californians; Ballena Press: Los Altos, CA, USA, 1993. [Google Scholar]
- Anderson, K. Tending the Wild: Native American Knowledge and the Management of California’s Natural Resources; University of California Press: Berkeley, CA, USA, 2006. [Google Scholar]
- Blondel, J.; Aronson, J.; Bodiou, J.-Y.; Boeuf, G. The Meditteranean Region: Biological Diversity in Space and Time, 2nd ed.; Oxford University Press: Oxford, UK, 2010. [Google Scholar]
- Behr, K.-E. Man’s Role in the Shaping of the Eastern Mediterranean Landscape; Bottema, S., Entjes-Nieborg, G., Van Zeist, W., Eds.; A.A. Balkema: Rotterdam, The Netherlands, 1990; pp. 219–230. [Google Scholar]
- Johnson-Gottesfeld, L. Conservation, Territory, and Traditional Beliefs: An Analysis of Gitksan and Wet’suwet’en Subsistence, Northwest British Columbia, Canada. J. Hum. Ecol. 2016, 22, 443–465. [Google Scholar] [CrossRef]
- Pimbert, M.P.; Pretty, J.N. People, Parks and Professionals: Putting “participation” into protected area management. UNRISD 1997, 16, 297–330. [Google Scholar]
- Hill, M.A.; Press, A.J. Kakadu National Park—A history, Biodiversity—Broadening the Debate. Canberra Aust. Nat. Conserv. Agency Publ. 1994, 2, 4–23. [Google Scholar]
- Roberts, R.G.; Jones, R.; Smith, A. Thermoluminescence dating of a 50,000 year old human occupation site in northern Australia, Nature. Nature 1990, 345, 153–156. [Google Scholar] [CrossRef]
- Mckechnie, I.M.; Thomsen, C.J.M.; Sargent, R.D. Agriculture, Ecosystems and Environment Forested fi eld edges support a greater diversity of wild pollinators in lowbush blueberry (Vaccinium angustifolium). Agric. Ecosyst. Environ. 2017, 237, 154–161. [Google Scholar] [CrossRef]
- Lewis, H.T.; Ferguson, T.A. Yards, Corridors, and Mosaics: How to Burn a Boreal Forest. Hum. Ecol. 1988, 16, 57–77. [Google Scholar] [CrossRef]
- Smith, E.A.; Wishnie, M. Conservation and Subsistence in Small-Scale Societies. Annu. Rev. Anthropol. 2000, 29, 493–524. [Google Scholar] [CrossRef] [Green Version]
- Johnson, P.T.J.; Preston, D.L.; Hoverman, J.T.; Richgels, K.L.D. Biodiversity decreases disease through predictable changes in host community competence. Nature 2013, 494, 230–233. [Google Scholar] [CrossRef] [PubMed]
- Spielman, D.; Brook, B.W.; Briscoe, D.; Frankham, R. Does inbreeding and loss of genetic diversity reduce disease resistance? Conserv. Genet. 2004, 5, 439–448. [Google Scholar] [CrossRef]
- Pearman, P.B.; Garner, T.W.J. Susceptibility of Italian agile frog populations to an emerging strain of Ranavirus parallels population genetic diversity. Ecol. Lett. 2005, 8, 401–408. [Google Scholar] [CrossRef]
- Altermatt, F.; Ebert, D. Genetic diversity of daphnia magna populations enhances resistance to parasites. Ecol. Lett. 2008, 11, 918–928. [Google Scholar] [CrossRef]
- Letourneau, D.K.; Armbrecht, I.; Rivera, B.S.; Lerma, J.M.; Carmona, E.J.; Daza, M.C.; Escobar, S.; Galindo, V.; Gutiérrez, C.; López, S.D.; et al. Does plant diversity benefit agroecosystems? A synthetic review. Ecol. Appl. 2011, 21, 9–21. [Google Scholar] [CrossRef]
- Powers, A. Plant Community Diversity, Herbivore Movement, and an Insect-Transmitted Disease of Maize. Ecol. Soc. Am. 1987, 68, 1658–1669. [Google Scholar] [CrossRef]
- Upadhyaya, H.D.; Ortiz, R.; Bramel, P.J.; Singh, S. Phenotypic diversity for morphological and agronomic characteristics in chickpea core collection. Euphytica 2002, 123, 333–342. [Google Scholar] [CrossRef]
- Gibson, R.W. A Review of Perceptual Distinctiveness in Landraces Including an Analysis of How Its Roles Have Been Overlooked in Plant Breeding for Low-Input Farming Systems. Econ. Bot. 2009, 63, 242–255. [Google Scholar] [CrossRef]
- Youyong, Z.; Chen, H.; Fan, J.; Wang, Y.; Li, Y.; Chen, J.; Fan, J.; Yang, S.; Hu, L.; Leung, H.; et al. Genetic diversity and disease control in rice. Nature 2000, 406, 718–722. [Google Scholar]
- Ssekandi, W.; Mulumba, J.W.; Colangelo, P.; Nankya, R.; Fadda, C.; Karungi, J.; Otim, M.; De Santis, P.; Jarvis, D.I. The use of common bean (Phaseolus vulgaris) traditional varieties and their mixtures with commercial varieties to manage bean fly (Ophiomyia spp.) infestations in Uganda. J. Pest Sci. 2016, 89, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Van Leur, J.A.G.; Ceccarelli, S.; Grando, S. Diversity for disease resistance in barley landraces from Syria and Jordan. Plant Breed. 1989, 103, 324–335. [Google Scholar] [CrossRef]
- Naegele, R.P.; Boyle, S.; Quesada-Ocampo, L.M.; Hausbeck, M.K. Genetic diversity, population structure, and resistance to Phytophthora capsici of a worldwide collection of eggplant germplasm. PLoS ONE 2014, 9, 31–32. [Google Scholar] [CrossRef] [PubMed]
- Singh, B. Breeding Cowpea Varieties for resistance to Striga gesneriodes and Alectra vogelii. In Challenges and Opportunities for Enhancing Sustainable Cowpea Production; Fatokun, C.A., Tarawali, S., Singh, B.B., Kormawa, P.M., Tamo, M., Eds.; International Institute of Tropical Agriculture: Ibadan, Nigeria, 2002. [Google Scholar]
- Balée, W. Cultural Forests of the Amazon:A Historical Ecology of People and Their Landscapes; University of Alabama Press: Tuscaloosa, AL, USA, 2013. [Google Scholar]
- Earnshaw, J.T.K. Cultural Forests of the Southern Nuu-chah-nulth: Historical Ecology and Salvage Archaeology on Vancouver Island’s West Coast. Ph.D. Thesis, University of Victoria, Victoria, BC, Canada, 2016. [Google Scholar]
- Balooni, K.; Inoue, M. Decentralized Forest Management in South and Southeast Asia. J. For. 2007, 105, 414–420. [Google Scholar]
- Nguyen, Q.; Hoang, M.H.; Oborn, I.; van Noordwijk, M. Multipurpose agroforestry as a climate change resiliency option for farmers: An example of local adaptation in Vietnam Multipurpose agroforestry as a climate change resiliency option for farmers: An example of local adaptation in Vietnam. Clim. Chang. 2013, 117, 241–257. [Google Scholar] [CrossRef]
- Pokharel, B.K.; Branney, P.; Nurse, M.; Malla, Y.B. Community Forestry: Conserving Forests, Sustaining Livelihoods and Strengthening Democracy. J. For. Livelihood 2007, 6, 8–19. [Google Scholar]
- Kaya, Z.; Raynal, D.J. Biodiversity and conservation of Turkish forests. Biol. Conserv. 2001, 97, 131–141. [Google Scholar] [CrossRef]
- Okan, T.; Köse, C.; Wall, J. Socioeconomic Perspectives on Household Chestnut Fruit Utilization and Chestnut Blight Prevention Efforts in Turkey. Sustainability 2017, 9, 831. [Google Scholar] [CrossRef]
- Wall, J.; Aghayeva, D.N. The Practice and Importance of Chestnut Cultivation in Azerbaijan in the Face of Blight, Cryphonectria parasitica ( Murrill ) Barr. Ethnobot. Res. Appl. 2014, 12, 165–174. [Google Scholar] [CrossRef]
- Pezzi, G.; Maresi, G.; Conedera, M.; Ferrari, C. Woody species composition of chestnut stands in the Northern Apennines: The result of 200 years of changes in land use. Landsc. Ecol. 2011, 26, 1463–1476. [Google Scholar] [CrossRef]
- Mattioni, C.; Cherubini, M.; Micheli, E.; Villani, F.; Bucci, G. Role of domestication in shaping Castanea sativa genetic variation in Europe. Tree Genet. Genomes 2008, 4, 563–574. [Google Scholar] [CrossRef]
- Miles, M.; Huberman, A. Qualitative Data Analysis: A Sourcebook of New Methods; Sage Publications: Beverly Hills, CA, USA, 1984. [Google Scholar]
- Tizado, E.J.; Terrón, A.; Núñez-Pérez, E. A methodology to evaluate disease severity: A case study of chestnut blight in El Bierzo region (northwestern Spain). Ann. Appl. Biol. 2012, 161, 81–90. [Google Scholar] [CrossRef]
- Evans, J. Quantitative Methods in Spatial Ecology. Available online: http://evansmurphy.wixsite.com/evansspatial/arcgis-gradient-metrics-toolbox (accessed on 25 June 2019).
- Le Roux, B.; Rouanet, H. Multiple Correspondence Analysis; Sage Publications: Beverly Hills, CA, USA, 2010. [Google Scholar]
- Husson, F.; Josse, J.; Le, S.; Mazet, J. FactoMineR: Factor Analysis and Data Mining with R. Available online: http://CRAN.R-project.org/package=FactoMineR (accessed on 25 June 2019).
- Pagès, J. Multiple factor analysis: Main features and application to sensory data. Rev. Colomb. Estad. 2004, 27, 1–26. [Google Scholar]
- Abdi, H.; Williams, L.J.; Valentin, D. Multiple factor analysis: Principal component analysis for multitable and multiblock data sets. Wiley Interdiscip. Rev. Comput. Stat. 2013, 5, 149–179. [Google Scholar] [CrossRef]
- Anderson, C.J.; Verkuilen, J.; Johnson, T.R. Applied Generalized Linear Mixed Models: Continuous and Discrete Data; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A protocol for data exploration to avoid common statistical problems. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Conedera, M.; Krebs, P.; Tinner, W.; Pradella, M.; Torriani, D. The cultivation of Castanea sativa (Mill.) in Europe, from its origin to its diffusion on a continental scale. Veg. Hist. Archaeobot. 2004, 13, 161–179. [Google Scholar] [CrossRef] [Green Version]
- Flower, M. (Ed.) Xenophon Anabasis; Oxford University Press: Oxford, UK, 2012. [Google Scholar]
- Krebs, P.; Conedera, M.; Pradella, M.; Torriani, D.; Felber, M.; Tinner, W. Quaternary refugia of the sweet chestnut (Castanea sativa Mill.): an extended palynological approach. Veg. Hist. Archaeobot. 2004, 13, 145–160. [Google Scholar] [CrossRef] [Green Version]
- Prospero, S.; Lutz, A.; Tavadze, B.; Supatashvili, A.; Rigling, D. Discovery of a new gene pool and a high genetic diversity of the chestnut blight fungus cryphonectria parasitica in caucasian georgia. Infect. Genet. Evol. 2013, 20, 131–139. [Google Scholar] [CrossRef]
- FAO (Food and Agriculture Organization). Report on Chestnut Blight and Its Management in Turkey. Available online: http://www.gonder.org.tr/wp-content/uploads/2015/12/FINAL_REPORT_of_ TCPTUR3201_Management_of_Chestnut_blight.doc (accessed on 25 June 2019).
- Johnson, L.; Hunn, E. Introduction. In Landscape Ethnoecology: Concepts of Biotic and Physical Space; Johnson, L., Hunn, E., Eds.; Berghahn Books: New York, NY, USA, 2010. [Google Scholar]
- Smith, I.M.; Dunez, J.; Phillips, D.; Lelliott, R.A.; Archer, S. European Handbook of Plant Diseases; John Wiley & Sons: Hoboken, NJ, USA, 2009. [Google Scholar]
- Mattioni, C.; Martin, M.A.; Pollegioni, P.; Cherubini, M.; Villani, F. Microsatellite markers reveal a strong geographical structure in European populations of Castanea sativa (Fagaceae): Evidence for multiple glacial refugia. Am. J. Bot. 2013, 100, 951–961. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, D.; Crabtree, J.; Wiesinger, G.; Dax, T.; Stamou, N.; Fleury, P.; Gutierrez Lazpita, J.; Gibon, A. Agricultural abandonment in mountain areas of Europe: Environmental consequences and policy response. J. Environ. Manag. 2000, 59, 47–69. [Google Scholar] [CrossRef]
- Lasanta-Martínez, T.; Vicente-Serrano, S.M.; Cuadrat-Prats, J.M. Mountain Mediterranean landscape evolution caused by the abandonment of traditional primary activities: A study of the Spanish Central Pyrenees. Appl. Geogr. 2005, 25, 47–65. [Google Scholar] [CrossRef]
- Mottet, A.; Ladet, S.; Coque, N.; Gibon, A. Agricultural land-use change and its drivers in mountain landscapes: A case study in the Pyrenees. Agric. Ecosyst. Environ. 2006, 114, 296–310. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Statistical Measures | Diameter (cm) | Elevation (m) | Slope (m) | Wetness (NDVI) | Coppiced (%) | Grafted (%) |
|---|---|---|---|---|---|---|---|
| 3 | Average | 13.8 | 138 | 7.5 | 6016 | 0 | |
| Standard deviation | 6.5 | 36.0 | 6.6 | 1541 | 53 | ||
| Range | 5–33 | 102–222 | 1–19 | 1321–7082 | |||
| 2 | Average | 40.4 | 592 | 18.4 | 6959 | 2.6 | |
| Standard deviation | 37.6 | 253 | 9.2 | 600 | 9 | ||
| Range | 5–320 | 134–1178 | 1–44 | 5392–7950 | |||
| 1 | Average | 37.5 | 551 | 18.8 | 6328 | 58 | |
| Standard deviation | 36 | 436 | 6.6 | 966 | 3 | ||
| Range | 5–241 | 40–1356 | 3–32 | 4497–7503 |
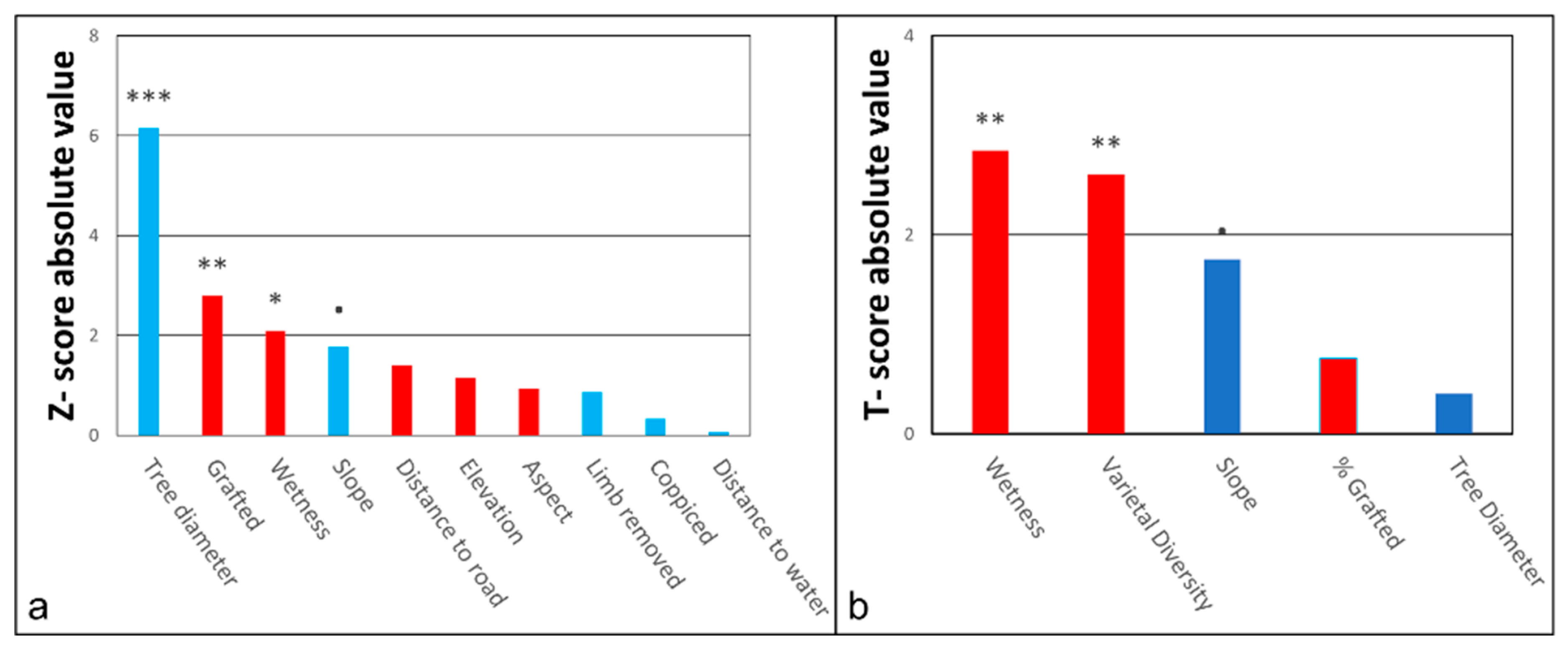
| Factor | DF | Chisq | Z-Value | Pr(>|z|) |
|---|---|---|---|---|
| Tree diameter | 1 | 37.8359 | 6.151 | 7.7e−10 *** |
| Grafted | 1 | 7.7861 | −2.790 | 0.00526 ** |
| Wetness | 1 | 4.3792 | −2.093 | 0.03638 * |
| Slope | 1 | 3.1451 | 1.773 | 0.07615 • |
| Distance to Road | 1 | 1.9748 | −1.405 | 0.15994 |
| Elevation | 1 | 1.3206 | −1.149 | 0.25049 |
| Aspect | 1 | 0.8769 | −0.936 | 0.34905 |
| Limb Removed | 1 | 0.7524 | 0.867 | 0.38572 |
| Coppiced | 1 | 0.1122 | 0.335 | 0.73766 |
| Distance to Water | 1 | 0.0053 | −0.072 | 0.94223 |
| Factor | DF | Chisq | T-Value | Pr(>Chisq) |
|---|---|---|---|---|
| Wetness | 1 | 8.0928 | −2.845 | 0.00444 ** |
| Varietal Diversity | 1 | 6.7944 | −2.607 | 0.00914 ** |
| Slope | 1 | 3.0764 | 1.754 | 0.79438 • |
| Percent Grafted | 1 | 0.5760 | −0.759 | 0.447872 |
| Tree Diameter | 1 | 0.1631 | 0.404 | 0.686357 |
| Land Use or Access Code | Number of Reports |
|---|---|
| Near house on titled land | 33 |
| Near house on untitled land | 9 |
| Away from the house on titled land | 9 |
| On state land under traditional claim | 93 |
| On state land without traditional claim | 2 |
| Treasury, local municipality land | 2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wall, J.; Köse, C.; Köse, N.; Okan, T.; Aksoy, E.B.; Jarvis, D.; Allred, S. The Role of Traditional Livelihood Practices and Local Ethnobotanical Knowledge in Mitigating Chestnut Disease and Pest Severity in Turkey. Forests 2019, 10, 571. https://doi.org/10.3390/f10070571
Wall J, Köse C, Köse N, Okan T, Aksoy EB, Jarvis D, Allred S. The Role of Traditional Livelihood Practices and Local Ethnobotanical Knowledge in Mitigating Chestnut Disease and Pest Severity in Turkey. Forests. 2019; 10(7):571. https://doi.org/10.3390/f10070571
Chicago/Turabian StyleWall, Jeffrey, Coşkun Köse, Nesibe Köse, Taner Okan, Elif Başak Aksoy, Devra Jarvis, and Shorna Allred. 2019. "The Role of Traditional Livelihood Practices and Local Ethnobotanical Knowledge in Mitigating Chestnut Disease and Pest Severity in Turkey" Forests 10, no. 7: 571. https://doi.org/10.3390/f10070571