Differential Responses and Controls of Soil CO2 and N2O Fluxes to Experimental Warming and Nitrogen Fertilization in a Subalpine Coniferous Spruce (Picea asperata Mast.) Plantation Forest


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Experimental Set-Up and Design
2.3. Microclimate Measurements
2.4. Soil CO2 and N2O Fluxes Measurements
2.5. Soil Samples and Analysis
2.6. Data Analysis
3. Results
3.1. Soil Carbon, Nitrogen and Microbial Properties
3.2. Soil CO2 and N2O Fluxes
3.3. Relationship between the Soil CO2 and N2O Fluxes and Environmental Factors
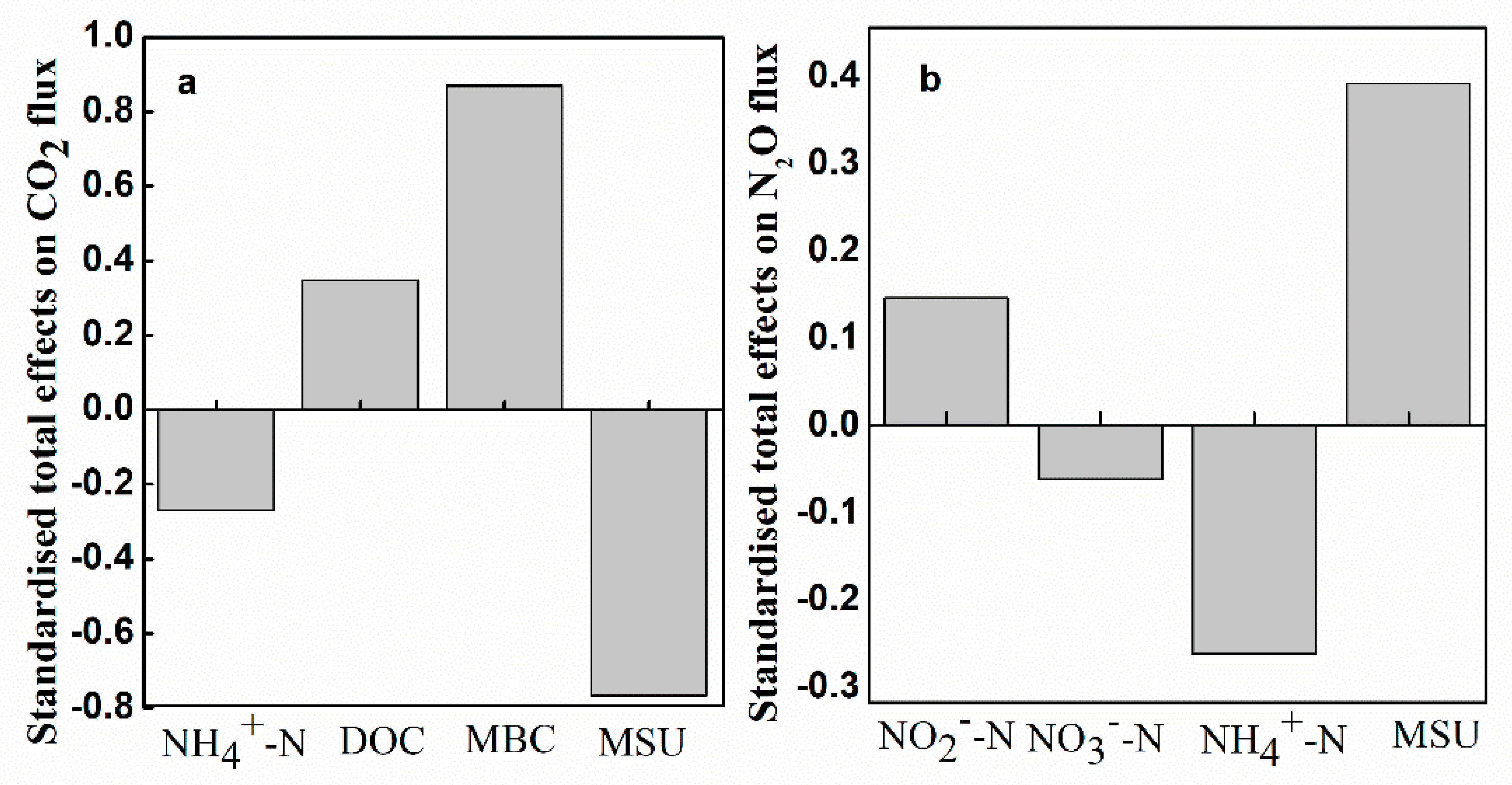
3.4. Contributions of Soil Variables to Soil CO2 and N2O Fluxes
4. Discussion
4.1. Effects of Warming and Nitrogen Fertilization on Soil CO2 Flux
4.2. Effects of Warming and Nitrogen Fertilization on Soil N2O Flux
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Trenberth, K.E. Stronger evidence of human influences on climate—The 2001 IPCC assessment. Environment 2001, 43, 8–19. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2013: The Physical Science Basis; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar]
- Chen, X.P.; Wang, G.X.; Zhang, T.; Mao, T.X.; Wei, D.; Hu, Z.Y.; Song, C.L. Effects of warming and nitrogen fertilization on GHG flux in the permafrost region of an alpine meadow. Atmos. Environ. 2017, 157, 111–124. [Google Scholar] [CrossRef]
- Galloway, J.N.; Cowling, E.B. Reactive nitrogen and the world: 200 years of change. Ambio 2002, 31, 64–71. [Google Scholar] [CrossRef]
- Zou, J.L.; Tobin, B.; Luo, Y.Q.; Osborne, B. Differential responses of soil CO2 and N2O fluxes to experimental warming. Agric. For. Meteorol. 2018, 259, 11–22. [Google Scholar] [CrossRef]
- Wu, Z.T.; Dijkstra, P.; Koch, G.W.; Penuelas, J.; Hungate, B.A. Responses of terrestrial ecosystems to temperature and precipitation change: A meta-analysis of experimental manipulation. Glob. Chang. Biol. 2011, 17, 927–942. [Google Scholar] [CrossRef]
- Yin, H.J.; Xiao, J.; Li, Y.F.; Chen, Z.; Cheng, X.Y.; Zhao, C.Z.; Liu, Q. Warming effects on root morphological and physiological traits: The potential consequences on soil C dynamics as altered root exudation. Agric. For. Meteorol. 2013, 180, 287–296. [Google Scholar] [CrossRef]
- Bijoor, N.S.; Czimczik, C.I.; Pataki, D.E.; Billings, S.A. Effects of temperature and fertilization on nitrogen cycling and community composition of an urban lawn. Glob. Chang. Biol. 2008, 14, 2119–2131. [Google Scholar] [CrossRef] [Green Version]
- Cantarel, A.A.M.; Bloor, J.M.G.; Pommier, T.; Guillaumaud, N.; Moirot, C.; Soussana, J.F.; Poly, F. Four years of experimental climate change modifies the microbial drivers of N2O fluxes in an upland grassland ecosystem. Glob. Chang. Biol. 2012, 18, 2520–2531. [Google Scholar] [CrossRef]
- Hu, Y.G.; Chang, X.F.; Lin, X.W.; Wang, Y.F.; Wang, S.P.; Duan, J.C.; Zhang, Z.H.; Yang, X.X.; Luo, C.Y.; Xu, G.P.; et al. Effects of warming and grazing on N2O fluxes in an alpine meadow ecosystem on the Tibetan plateau. Soil Biol. Biochem. 2010, 42, 944–952. [Google Scholar] [CrossRef]
- Jassal, R.S.; Black, T.A.; Trofymow, J.A.; Roy, R.; Nesic, Z. Soil CO2 and N2O flux dynamics in a nitrogen-fertilized Pacific Northwest Douglas-fir stand. Geoderma 2010, 157, 118–125. [Google Scholar] [CrossRef]
- Geng, J.; Cheng, S.L.; Fang, H.J.; Yu, G.R.; Li, X.Y.; Si, G.Y.; He, S.; Yu, G.X. Soil nitrate accumulation explains the nonlinear responses of soil CO2 and CH4 fluxes to nitrogen addition in a temperate needle-broadleaved mixed forest. Ecol. Indic. 2017, 79, 28–36. [Google Scholar] [CrossRef]
- Huang, R.; Wang, Y.; Liu, J.; Li, J.; Xu, G.; Luo, M.; Xu, C.; Ci, E.; Gao, M. Variation in N2O emission and N2O related microbial functional genes in straw- and biochar-amended and non-amended soils. Appl. Soil Ecol. 2019, 137, 57–68. [Google Scholar] [CrossRef]
- Zhang, J.J.; Peng, C.H.; Zhu, Q.A.; Xue, W.; Shen, Y.; Yang, Y.Z.; Shi, G.H.; Shi, S.W.; Wang, M. Temperature sensitivity of soil carbon dioxide and nitrous oxide emissions in mountain forest and meadow ecosystems in China. Atmos. Environ. 2016, 142, 340–350. [Google Scholar] [CrossRef]
- Shrestha, R.K.; Strahm, B.D.; Sucre, E.B. Greenhouse gas emissions in response to nitrogen fertilization in managed forest ecosystems. New For. 2015, 46, 167–193. [Google Scholar] [CrossRef]
- Seo, J.; Jang, I.; Jung, J.Y.; Lee, Y.K.; Kang, H. Warming and increased precipitation enhance phenol oxidase activity in soil while warming induces drought stress in vegetation of an Arctic ecosystem. Geoderma 2015, 259, 347–353. [Google Scholar] [CrossRef]
- Zhang, W.; Parker, K.M.; Luo, Y.; Wan, S.; Wallace, L.L.; Hu, S. Soil microbial responses to experimental warming and clipping in a tallgrass prairie. Glob. Chang. Biol. 2005, 11, 266–277. [Google Scholar] [CrossRef]
- Qin, H.L.; Xing, X.Y.; Tang, Y.F.; Hou, H.J.; Yang, J.; Shen, R.; Zhang, W.Z.; Liu, Y.; Wei, W.X. Linking soil N2O emissions with soil microbial community abundance and structure related to nitrogen cycle in two acid forest soils. Plant Soil 2019, 435, 95–109. [Google Scholar] [CrossRef]
- Hogberg, P.; Hogberg, M.N.; Gottlicher, S.G.; Betson, N.R.; Keel, S.G.; Metcalfe, D.B.; Campbell, C.; Schindlbacher, A.; Hurry, V.; Lundmark, T.; et al. High temporal resolution tracing of photosynthate carbon from the tree canopy to forest soil microorganisms. New Phytol. 2008, 177, 220–228. [Google Scholar] [CrossRef]
- Djukic, I.; Zehetner, F.; Watzinger, A.; Horacek, M.; Gerzabek, M.H. In situ carbon turnover dynamics and the role of soil microorganisms therein: A climate warming study in an Alpine ecosystem. FEMS Microbiol. Ecol. 2013, 83, 112–124. [Google Scholar] [CrossRef]
- Gholz, H.L.; Wedin, D.A.; Smitherman, S.M.; Harmon, M.E.; Parton, W.J. Long-term dynamics of pine and hardwood litter in contrasting environments: Toward a global model of decomposition. Glob. Chang. Biol. 2000, 6, 751–765. [Google Scholar] [CrossRef]
- Wang, Y.S.; Cheng, S.L.; Fang, H.J.; Yu, G.R.; Yang, X.M.; Xu, M.J.; Dang, X.S.; Li, L.S.; Wang, L. Relationships between ammonia-oxidizing communities, soil methane uptake and nitrous oxide fluxes in a subtropical plantation soil with nitrogen enrichment. Eur. J. Soil Biol. 2016, 73, 84–92. [Google Scholar] [CrossRef]
- Martins, C.S.C.; Macdonald, C.A.; Anderson, I.C.; Singh, B.K. Feedback responses of soil greenhouse gas emissions to climate change are modulated by soil characteristics in dryland ecosystems. Soil Biol. Biochem. 2016, 100, 21–32. [Google Scholar] [CrossRef]
- Xu, Z.F.; Yin, H.J.; Xiong, P.; Wan, C.; Liu, Q. Short-term responses of Picea asperata seedlings of different ages grown in two contrasting forest ecosystems to experimental warming. Environ. Exp. Bot. 2012, 77, 1–11. [Google Scholar] [CrossRef]
- Zhao, C.; Zhu, L.; Liang, J.; Yin, H.; Yin, C.; Li, D.; Zhang, N.; Liu, Q. Effects of experimental warming and nitrogen fertilization on soil microbial communities and processes of two subalpine coniferous species in Eastern Tibetan Plateau, China. Plant Soil 2014, 382, 189–201. [Google Scholar] [CrossRef]
- Yin, H.J.; Li, Y.F.; Xiao, J.; Xu, Z.F.; Cheng, X.Y.; Liu, Q. Enhanced root exudation stimulates soil nitrogen transformations in a subalpine coniferous forest under experimental warming. Glob. Chang. Biol. 2013, 19, 2158–2167. [Google Scholar] [CrossRef]
- Zhang, Z.L.; Qiao, M.F.; Li, D.D.; Yin, H.J.; Liu, Q. Do warming-induced changes in quantity and stoichiometry of root exudation promote soil N transformations via stimulation of soil nitrifiers, denitrifiers and ammonifiers? Eur. J. Soil Biol. 2016, 74, 60–68. [Google Scholar] [CrossRef]
- Yin, H.J.; Chen, Z.; Liu, Q. Effects of experimental warming on soil N transformations of two coniferous species, Eastern Tibetan Plateau, China. Soil Biol. Biochem. 2012, 50, 77–84. [Google Scholar] [CrossRef]
- Cai, Y.J.; Wang, X.D.; Tian, L.L.; Zhao, H.; Lu, X.Y.; Yan, Y. The impact of excretal returns from yak and Tibetan sheep dung on nitrous oxide emissions in an alpine steppe on the Qinghai-Tibetan Plateau. Soil Biol. Biochem. 2014, 76, 90–99. [Google Scholar] [CrossRef]
- Liu, H.; Zhao, P.; Lu, P.; Wang, Y.S.; Lin, Y.B.; Rao, X.Q. Greenhouse gas fluxes from soils of different land-use types in a hilly area of South China. Agric. Ecosyst. Environ. 2008, 124, 125–135. [Google Scholar] [CrossRef]
- Walkley, A.; Black, L.A. An examination of the Dgtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Li, Y.Q.; Qing, Y.X.; Lyu, M.K.; Chen, S.D.; Yang, Z.J.; Lin, C.F.; Yang, Y.S. Effects of artificial warming on different soil organic carbon and nitrogen pools in a subtropical plantation. Soil Biol. Biochem. 2018, 124, 161–167. [Google Scholar] [CrossRef]
- Cohen, J.B. Practical Organic Chemistry; Macmillan Collection Library: London, UK, 1910. [Google Scholar]
- Maynard, D.G.; Kalra, Y.P. Nitrate and exchangeable ammonium nitrogen. In Soil Sampling and Methods of Analysis; Carter, M.R., Ed.; Lewis: Edmonton, AB, Canada, 1993. [Google Scholar]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An Extraction Method for Measuring Soil Microbial Biomass-C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Dijkstra, F.A.; Morgan, J.A.; Follett, R.F.; Lecain, D.R. Climate change reduces the net sink of CH4 and N2O in a semiarid grassland. Glob. Chang. Biol. 2013, 19, 1816–1826. [Google Scholar] [CrossRef]
- Xu, Z.F.; Wan, C.A.; Xiong, P.; Tang, Z.; Hu, R.; Cao, G.; Liu, Q. Initial responses of soil CO2 efflux and C, N pools to experimental warming in two contrasting forest ecosystems, Eastern Tibetan Plateau, China. Plant Soil 2010, 336, 183–195. [Google Scholar] [CrossRef]
- Zhang, J.J.; Li, Y.F.; Chang, S.X.; Qin, H.; Fu, S.L.; Jiang, P.K. Understory management and fertilization affected soil greenhouse gas emissions and labile organic carbon pools in a Chinese chestnut plantation. For. Ecol. Manag. 2015, 337, 126–134. [Google Scholar] [CrossRef]
- Deng, Q.; Zhou, G.; Liu, J.; Liu, S.; Duan, H.; Zhang, D. Responses of soil respiration to elevated carbon dioxide and nitrogen addition in young subtropical forest ecosystems in China. Biogeosciences 2010, 7, 315–328. [Google Scholar] [CrossRef] [Green Version]
- Barrena, I.; Menéndez, S.; Duñabeitia, M.; Merino, P.; Stange, C.F.; Spott, O.; González-Murua, C.; Estavillo, J.M. Greenhouse gas fluxes (CO2, N2O and CH4) from forest soils in the Basque Country: Comparison of different tree species and growth stages. For. Ecol. Manag. 2013, 310, 600–611. [Google Scholar] [CrossRef]
- Luo, Y.Q.; Wan, S.Q.; Hui, D.F.; Wallace, L.L. Acclimatization of soil respiration to warming in a tall grass prairie. Nature 2001, 413, 622–625. [Google Scholar] [CrossRef]
- Oechel, W.C.; Vourlitis, G.L.; Hastings, S.J.; Zulueta, R.C.; Hinzman, L.; Kane, D. Acclimation of ecosystem CO2 exchange in the Alaskan Arctic in response to decadal climate warming. Nature 2000, 406, 978–981. [Google Scholar] [CrossRef]
- Walker, T.W.N.; Kaiser, C.; Strasser, F.; Herbold, C.W.; Leblans, N.I.W.; Woebken, D.; Janssens, I.A.; Sigurdsson, B.D.; Richter, A. Microbial temperature sensitivity and biomass change explain soil carbon loss with warming. Nat. Clim. Chang. 2018, 8. [Google Scholar] [CrossRef]
- Bossio, D.A.; Scow, K.M. Impact of Carbon and Flooding on the Metabolic Diversity of Microbial Communities in Soils. Appl. Environ. Microb. 1995, 61, 4043–4050. [Google Scholar]
- Paul, E.A.; Clark, F.E. Soil Microbiology and Biochemistry; Academic Press: San Diego, CA, USA, 1989. [Google Scholar]
- Sakamoto, K.; Oba, Y. Effect of Fungal to Bacterial Biomass Ratio on the Relationship between CO2 Evolution and Total Soil Microbial Biomass. Biol. Fert. Soils 1994, 17, 39–44. [Google Scholar] [CrossRef]
- Chen, R.R.; Senbayram, M.; Blagodatsky, S.; Myachina, O.; Dittert, K.; Lin, X.G.; Blagodatskaya, E.; Kuzyakov, Y. Soil C and N availability determine the priming effect: Microbial N mining and stoichiometric decomposition theories. Glob. Chang. Biol. 2014, 20, 2356–2367. [Google Scholar] [CrossRef]
- Fisk, M.; Santangelo, S.; Minick, K. Carbon mineralization is promoted by phosphorus and reduced by nitrogen addition in the organic horizon of northern hardwood forests. Soil Biol. Biochem. 2015, 81, 212–218. [Google Scholar] [CrossRef]
- Qiu, Q.Y.; Wu, L.F.; Ouyang, Z.; Li, B.B.; Xu, Y.Y.; Wu, S.S.; Gregorich, E.G. Priming effect of maize residue and urea N on soil organic matter changes with time. Appl. Soil Ecol. 2016, 100, 65–74. [Google Scholar] [CrossRef]
- Wang, Y.S.; Cheng, S.L.; Fang, H.J.; Yu, G.R.; Xu, X.F.; Xu, M.J.; Wang, L.; Li, X.Y.; Si, G.Y.; Geng, J.; et al. Contrasting effects of ammonium and nitrate inputs on soil CO2 emission in a subtropical coniferous plantation of southern China. Biol. Fert. Soils 2015, 51, 815–825. [Google Scholar] [CrossRef]
- TingTing, Z.; ZILiang, Z.; Na, L.; YuanShuang, Y.; DongHui, Z.; Qin, L.; HuaJun, Y. Differential uptakes of different forms of soil nitrogen among major tree species in subalpine coniferous forests of western Sichuan, China. Chin. J. Plant Ecol. 2017, 41, 1051–1059. [Google Scholar]
- Fang, H.J.; Cheng, S.L.; Yu, G.R.; Xu, M.J.; Wang, Y.S.; Li, L.S.; Dang, X.S.; Wang, L.; Li, Y.N. Experimental nitrogen deposition alters the quantity and quality of soil dissolved organic carbon in an alpine meadow on the Qinghai-Tibetan Plateau. Appl. Soil Ecol. 2014, 81, 1–11. [Google Scholar] [CrossRef]
- Schindlbacher, A.; Zechmeister-Boltenstern, S.; Butterbach-Bahl, K. Effects of soil moisture and temperature on NO, NO2, and N2O emissions from European forest soils. J. Geophys. Res.-Atmos. 2004, 109. [Google Scholar] [CrossRef]
- Wu, X.; Bruggemann, N.; Gasche, R.; Shen, Z.Y.; Wolf, B.; Butterbach-Bahl, K. Environmental controls over soil-atmosphere exchange of N2O, NO, and CO2 in a temperate Norway spruce forest. Glob. Biogeochem. Cycles 2010, 24, 45. [Google Scholar] [CrossRef]
- Kiese, R.; Butterbach-Bahl, K. N2O and CO2 emissions from three different tropical forest sites in the wet tropics of Queensland, Australia. Soil Biol. Biochem. 2002, 34, 975–987. [Google Scholar] [CrossRef]
- Werner, C.; Kiese, R.; Butterbach-Bahl, K. Soil-atmosphere exchange of N2O, CH4, and CO2 and controlling environmental factors for tropical rain forest sites in western Kenya. J. Geophys. Res.-Atmos. 2007, 112, 71. [Google Scholar] [CrossRef]
- Yan, J.H.; Zhang, W.; Wang, K.Y.; Qin, F.; Wang, W.T.; Dai, H.T.; Li, P.X. Responses of CO2, N2O and CH4 fluxes between atmosphere and forest soil to changes in multiple environmental conditions. Glob. Chang. Biol. 2014, 20, 300–312. [Google Scholar] [CrossRef]
- Venterea, R.T.; Groffman, P.M.; Verchot, L.V.; Magill, A.H.; Aber, J.D.; Steudler, P.A. Nitrogen oxide gas emissions from temperate forest soils receiving long-term nitrogen inputs. Glob. Chang. Biol. 2003, 9, 346–357. [Google Scholar] [CrossRef]
- Barnard, R.; Leadley, P.W.; Hungate, B.A. Global change, nitrification, and denitrification: A review. Glob. Biogeochem. Cycles 2005, 19, 152. [Google Scholar] [CrossRef]
- Signor, D.; Cerri, C.E.P. Nitrous oxide emissions in agricultural soils: A review. Pesq. Agropec. Trop. 2013, 43, 322–338. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | NO3−-N | NH4+-N | NO2−-N | TN | SOC | DOC | MBC | MBN | MBC/MBN | AWCD | H | U |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Season (S) | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.001 | <0.001 | 0.010 |
| Warming (W) | <0.001 | 0.756 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.009 | 0.461 | 0.006 |
| Nitrogen (N) | <0.001 | 0.103 | <0.001 | 0.009 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.237 | 0.953 | 0.276 |
| S × W | <0.001 | 0.322 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.032 | <0.001 |
| S × N | <0.001 | 0.008 | 0.040 | 0.365 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.002 | 0.013 | 0.006 |
| W ×N | 0.565 | 0.011 | 0.059 | 0.015 | <0.001 | 0.002 | 0.003 | <0.001 | <0.001 | 0.016 | 0.115 | 0.032 |
| S × W × N | <0.001 | 0.209 | <0.001 | 0.080 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.001 | 0.002 | 0.004 |
| Variables | Treatment | CO2 | N2O |
|---|---|---|---|
| Fluxes | W0N0 | 36.04 ± 3.77 ab | 0.51 ± 0.11 a |
| WN0 | 27.90 ± 3.14 a | 0.65 ± 0.27 a | |
| W0N | 46.08 ± 5.39 b | 4.68 ± 1.61 b | |
| WN | 29.07 ± 3.29 a | 2.02 ± 0.32 b | |
| GWP | W0N0 | 9984 ± 321 ab | 20.31 ± 3.02 a |
| WN0 | 7800 ± 844 a | 25.63 ± 10.33 a | |
| W0N | 12748 ± 2110 b | 208.8 ± 56.37 b | |
| WN | 8002 ± 282 a | 79.88 ± 8.90 b | |
| ANOVA (F values) | Warming | 8.97 * | 4.52 |
| N fertilization | 1.64 | 17.47 ** | |
| Warming ∗ N fertilization | 1.27 | 5.34 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, D.; Liu, Q.; Yin, H.; Luo, Y.; Hui, D. Differential Responses and Controls of Soil CO2 and N2O Fluxes to Experimental Warming and Nitrogen Fertilization in a Subalpine Coniferous Spruce (Picea asperata Mast.) Plantation Forest. Forests 2019, 10, 808. https://doi.org/10.3390/f10090808
Li D, Liu Q, Yin H, Luo Y, Hui D. Differential Responses and Controls of Soil CO2 and N2O Fluxes to Experimental Warming and Nitrogen Fertilization in a Subalpine Coniferous Spruce (Picea asperata Mast.) Plantation Forest. Forests. 2019; 10(9):808. https://doi.org/10.3390/f10090808
Chicago/Turabian StyleLi, Dandan, Qing Liu, Huajun Yin, Yiqi Luo, and Dafeng Hui. 2019. "Differential Responses and Controls of Soil CO2 and N2O Fluxes to Experimental Warming and Nitrogen Fertilization in a Subalpine Coniferous Spruce (Picea asperata Mast.) Plantation Forest" Forests 10, no. 9: 808. https://doi.org/10.3390/f10090808