To Plant or Not to Plant, That Is the Question: Reforestation vs. Natural Regeneration of Hurricane-Disturbed Mangrove Forests in Guanaja (Honduras)


Faculty of Arts and Humanities, University of Passau, Innstraße 40, D-94032 Passau, Germany
Forests 2020, 11(10), 1068; https://doi.org/10.3390/f11101068
Submission received: 24 August 2020
/
Revised: 2 October 2020
/
Accepted: 2 October 2020
/
Published: 6 October 2020
(This article belongs to the Special Issue Natural Plant Regeneration Ecology in Forest Ecosystems)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Background and Objectives: Mangrove forests offer many essential ecosystem services, including the protection of (sub)tropical coastlines, their inhabitants, and the infrastructure from severe storms and tsunamis. However, mangroves themselves suffer severely from such phenomena. After such events, reforestation efforts are widely undertaken to facilitate the recovery of the mangroves. Many of these laborious activities, however, fail for a number of reasons. Material and Methods: In October 1998, the Honduran Island of Guanaja was severely hit by Hurricane Mitch, and, after the storm, almost all of the mangrove forests (97%) were rated as dead. Seven years after Mitch, a longterm survey on the regeneration of the mangroves started. Field samplings in six variably disturbed mangrove zones was conducted in 2005, 2009, and 2016. Along permanent line-transects, all living plant species were recorded for statistical analyses and for quantifying progress. In this paper, the focus is on the three most severely hit areas. In two of them, planting efforts were carried out while the third one was left to regenerate naturally. This setting allowed a direct comparison between natural and human-assisted regeneration processes under otherwise similar conditions and equally severe previous disturbance. Results: Reforestation measures were characterized by high mortality rates of Rhizophora mangle L. propagules planted predominantly. Some, however, surely survived and might have contributed to regeneration after the disturbance. In 2016, roughly two decades after Hurricane Mitch, low-growing Rhizophora mangle forests, with high ground cover, were found. Surprisingly, the area without any planting also witnessed similar mangrove rejuvenation in the same period. Conclusion: Findings on the recovering mangroves in Guanaja confirm the lessons learned from other mangrove rehabilitation measures: planting mostly fails and commonly does not accelerate the revegetation of disturbed forests. In naturally regenerating forests, recolonization may occur after a certain time-lag only if few diaspore sources survived in the surrounding; as soon as established mangrove plants bear propagules, a quick return to viable forests takes place, proving the high resilience of these coastal ecosystems.
1. Introduction
Mangroves offer many fundamental ecosystem services to humans [1,2,3,4,5,6,7,8,9]. Besides playing an important role in the earth’s carbon cycle, mangroves mitigate land degradation and erosion, act as a nursing ground for fish and invertebrates, and provide both timber and nontimber produce. Mangrove forests protect coastal human and nonhuman populations from tropical storms and tsunamis. Coastal protection is one of the most undervalued ecosystem services, despite having the highest net present value [10,11,12]. However, while they limit damage to infrastructure and physical harm to humans and nonhumans alike (e.g., [12,13,14,15,16,17,18,19]), mangroves can experience severe disturbances when such natural events occur and, consequently, lose their ecosystem functions and services [15,20,21]. Mangrove restoration and rehabilitation measures are widespread after any severe disturbance to help accelerate the rejuvenation of the mangroves for their ecosystem services [22,23,24,25,26,27]. Planting of propagules (mainly Rhizophora species) or seedlings raised in nurseries is a common approach, combined, sometimes (and, if so, generally more successful), with hydrologic manipulation to restore proper physiochemical conditions for mangrove establishment and regrowth. Many of these laborious activities, however, fail for a number of reasons (e.g., planting in unsuitable topographic location, insufficient postplanting care, modified site conditions, predation by crabs and beetles [28,29,30,31,32]). On the other hand, there is a growing body of evidence about the resilience of mangroves, showing that even after severe disturbances, mangroves grow back by natural regeneration within a reasonable period of time [20,33,34,35]. So, adapting a Shakespearian saying as the title, this paper questions whether the labor-intensive and sometimes expensive reforestation of mangroves [36] is indeed the most practical, “default” way of helping severely disturbed mangrove forests to recover.
The observations presented here are derived from a longterm survey on the recovery of hurricane-disturbed mangroves in the Honduran Island of Guanaja across six variably disturbed mangrove forests [37]. This paper focuses on the three most severely affected mangrove areas located on the windward side of the island and, thus, particularly exposed to the forces of Hurricane Mitch, which was one of the most powerful storms of the last century within the Caribbean [38]. All three areas showed basically no signs of regeneration after the storm, and, as chances for natural regrowth were assumed to be low [39], reforestation measures were carried out at some locations. In two out of the three mangrove areas, planting efforts were conducted, namely, Mangrove Bight (MB) and Savannah Bight (SB), while the third, West End South (WES), was left to natural regeneration (personal communication Mariela Ochoa), providing an opportunity to directly compare the outcomes of natural and human-assisted regeneration under otherwise comparable site and storm conditions.
2. Materials and Methods
2.1. Study Area
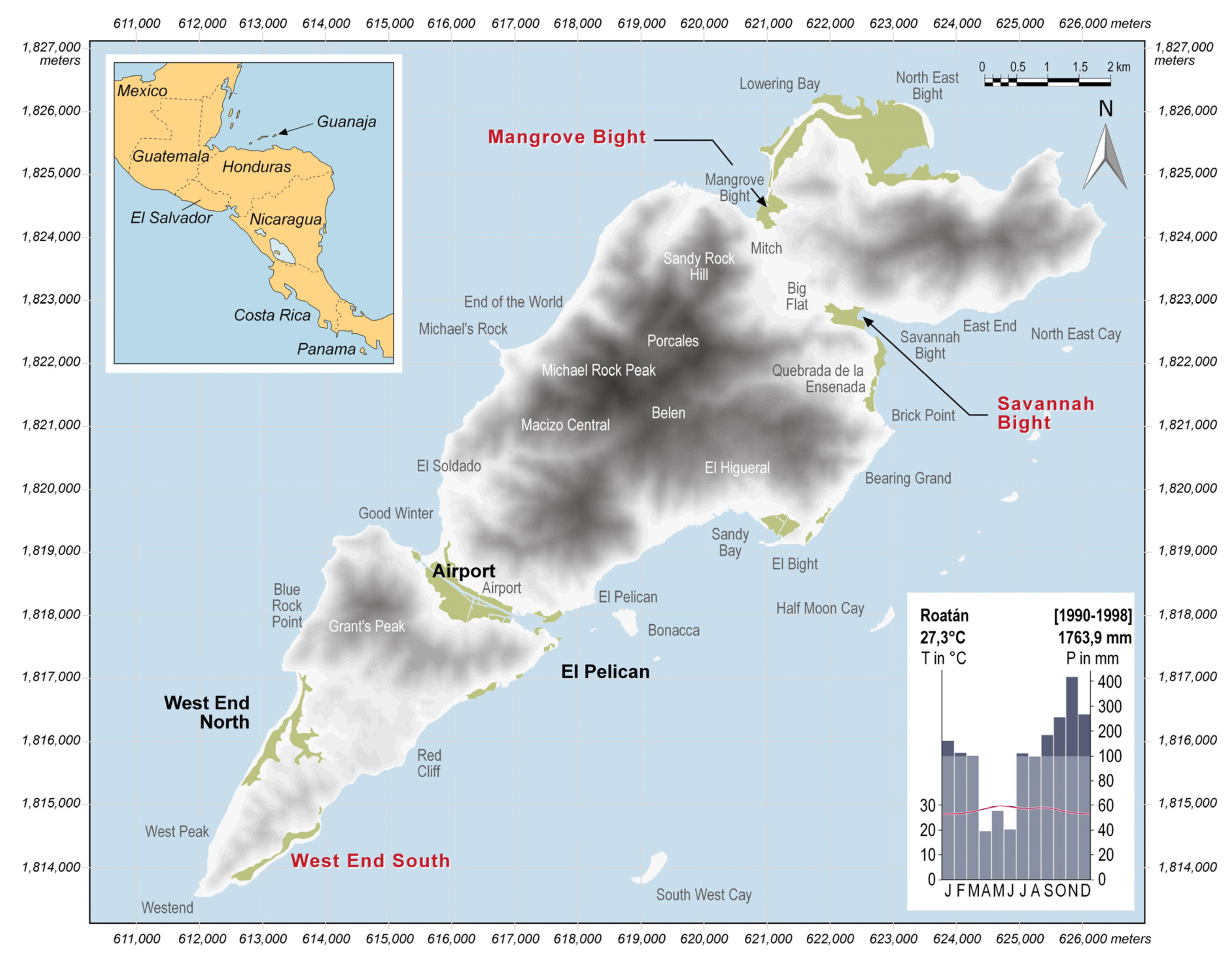
Guanaja is the second largest of the Bay Islands, located within the Gulf of Honduras (Figure 1). The relief of the island is hilly, with Michael Rock Peak (415 m a.s.l) being the highest point. Located at a latitude of 16° 28′ N, Guanaja has a tropical climate, affected by trade winds from East to Northeast year-round. Precipitation maximum is not caused by zenithal rains during passage of the Innertropical Convergence Zone but during the winter months, when continental-polar cold fronts occasionally move south into the Caribbean and the Gulf of Honduras. Annual mean temperature is 27.3 °C, and seasonal variation is low (Figure 1). Between May and October, a potential “hurricane season” arises when low-pressure systems move into the Caribbean from the Atlantic Ocean. One of the most powerful Atlantic hurricanes of the last century was Mitch [38], which hit Guanaja in October 1998. While making landfall on Guanaja on 27 October as a category SS4 storm, track speed slowed considerably (<8 km/h) and the eye of the hurricane persisted close to Guanaja between 27 and 29 October, thus striking the island with heavy winds for about 70 hours. During the night of 29 October, Mitch reached mainland Honduras and slowed down to a tropical storm only, but it still caused severe destruction and a high number of casualties in mainland Central America due to the landslides induced by the heavy rain [39,40].
Together with the coral reefs and the pine forests (Pinus caribaea var. hondurensis (Sénéclauze) W.H. Barrett & Golfari), the mangroves were among the most severely disturbed ecosystems on Guanaja [41]. Prior to the storm, the mangroves had a patchy distribution to leveled and poorly drained coastal areas with low wave action (Figure 1). The most common species in the mangroves on Guanaja was Rhizophora mangle, in many cases being monodominant. On slightly higher ground, with reduced inundation, Laguncularia racemosa (L.) C.F. Gaertn. and/or Avicennia germinans (L.) L. are commonly associated in the mangrove forests of the Bay Islands [37,42,43]. Transitional to terrestrial ecosystems, a belt of Conocarpus erectus L. and/or the mangrove fern Acrostichum aureum L. commonly occurs. Prior to the storm, the mangrove forests on Guanaja reached canopy heights of up to 20 m [43,44].
As chances for natural regrowth after Hurricane Mitch were regarded as low [39], the Honduran forestry agency ESCNACIFOR (Escuela Nacional de Ciencias Forestales) initiated a replanting program, carried out in 2003 and 2004 across locations in the seriously struck northeast of the island and along the canal at the Airport. Five years later, an American voluntary initiative commenced to plant tens of thousands of mangrove propagules over the next half-decade, again, mainly in the northeast of the island. Both planting campaigns almost exclusively used Rhizophora mangle propagules collected from the neighboring Island of Santa Elena (close to Roatan) or Guanaja itself. Initially the propagules were planted directly in both the initiatives. As mortality rates were found to be high, the second initiative started planting seedlings raised in nurseries (again primarily Rhizophora mangle, to a lesser degree also Laguncularia racemosa and Avicennia germinans) to increase chances of their survival. In the second reforestation campaign, local people as well as volunteers from abroad were involved, the most prominent one being Jimmy Carter, the former President of the United States of America. Total cost of the activities, covered by Canadian government in the first and by grants and donations in the second case, could not be exactly ascertained. In the year 2013, an annual budget of about USD10,000 was available to cover the expenses of the boat transport offered to collect propagules and to reach planting sites, as well as to instruct local people and volunteers on how to perform the planting (personal communication Mariela Ochoa). Similar figures can be assumed for other years. The exact number of propagules planted is also not available; however, estimates range between approximately 20,000 in 2010 and up to 100,000 in 2012 (personal communication Scott Duncan and Mariela Ochoa).
2.2. Field Sampling
Vegetation sampling was carried out using line-transects (line-intercept method, see [45]; Figure S1). The transects were set up, GPS-tracked, and marked by colored tapes in 2005 for allowing an exact match during resurveys, which so far took place in 2009 and 2016. The transects run from the seaward fringe to the landward margin, perpendicular to potential ecological gradients, as recommended by [46]. Depending on area size and outline of the mangrove areas, the length and number of the transects per site vary (SB: one transect of 550 m; MB: two transects of 250 and 270 m; WES: two transects of 110 and 150 m). Along the transects, the ground cover of all living plant species, mangrove seedling numbers, and the amount of woody debris were quantitatively recorded with a resolution of 0.1 m using a measuring tape. Shrub and tree species were recorded in different height classes (<2, 2–5, 5–10, 10–20 m). The linear measurements were translated into ground cover values (in %) per 10 m-segments for each species afterward for statistical analyses (see Figure S1). The taxonomy of plant species and life-form classification follows the Checklist of the Vascular Plants of Belize [47].
During the field campaign in March 2009, some important physiochemical soil parameters (temperature, pH, salinity, and redox potential) were measured along the transects with a multiparameter measuring device (Eijkelkamp 18.28, Agrisearch Equipment). Measurements were carried out at consistent spacings of 50 m, accounting for a total of 21 readings (SB: 10, MB: 7, WES: 4). Redox potential, which indicates whether soils are in aerobic or anaerobic conditions, were measured in-situ at a soil depth of 10 cm with a platinum Ag/AgCl 3 mol/L KCL electrode after equilibration for up to an hour, as soon as values remained constant. During this reading, the temperature was also recorded. Measurements of pH values and conductivity were conducted in a suspension of soil material from a 10 cm depth and the interstitial water filling the hole, mixed in a vessel at a ratio of 1:5. The pH values were obtained by a standard KCl pH electrode (T = 25°C) after calibration in 4.0 and 7.0 standards (NIST (National Institute of Standards and Technology) scale). The measured conductivity values were automatically translated into salinity figures in g/l by the measuring device.
2.3. Data Analysis
Data analysis included uni- and multivariate statistical procedures. Changes in the ground cover of species or functional groups are visualized by bar graphs showing mean ground cover per 10 m transect segment and for the study areas as a whole. A Principal Component Analysis (PCA), i.e., an unconstrained ordination analysis calculated from species data only [48], was employed to identify similarities, or rather, dissimilarities concerning species composition and/or structural attributes between transect segments of different sites and sampling dates. The data set includes both true species (common mangrove taxa such as Rhizophora mangle, Laguncularia racemosa, Avicennia germinans, Conocarpus erectus, and Acrostichum aureum) as well as functional groups integrating mainly less-common taxa (such as climbers, graminoids, herbs, epiphytes, and freshwater swamp trees/palms). The PCA results are displayed by an ordination scatter plot, arranging samples along meaningful gradients. Samples are shown as symbols and species/functional groups as arrows, pointing from the origin of the coordinates into the direction where samples with above-average values of the respective species/functional group are located. The arrangement of transect segments within the ordination space enables a spatiotemporal evaluation of the floristic and/or structural (dis)similarity between segments sampled atdifferent dates and/or sites. To depict the course of change, data are displayed separately for the three surveys in 2005, 2009, and 2016 because a scatterplot encompassing all sampling sites and dates would be very crowded. The PCA analysis was performed with the software Canoco 4.5 (Biometrics, Wageningen & České Budĕjovice). Species data were log-transformed (x’ = log (x+1)) prior to PCA calculation to moderate extreme values within the dataset [48,49].
3. Results
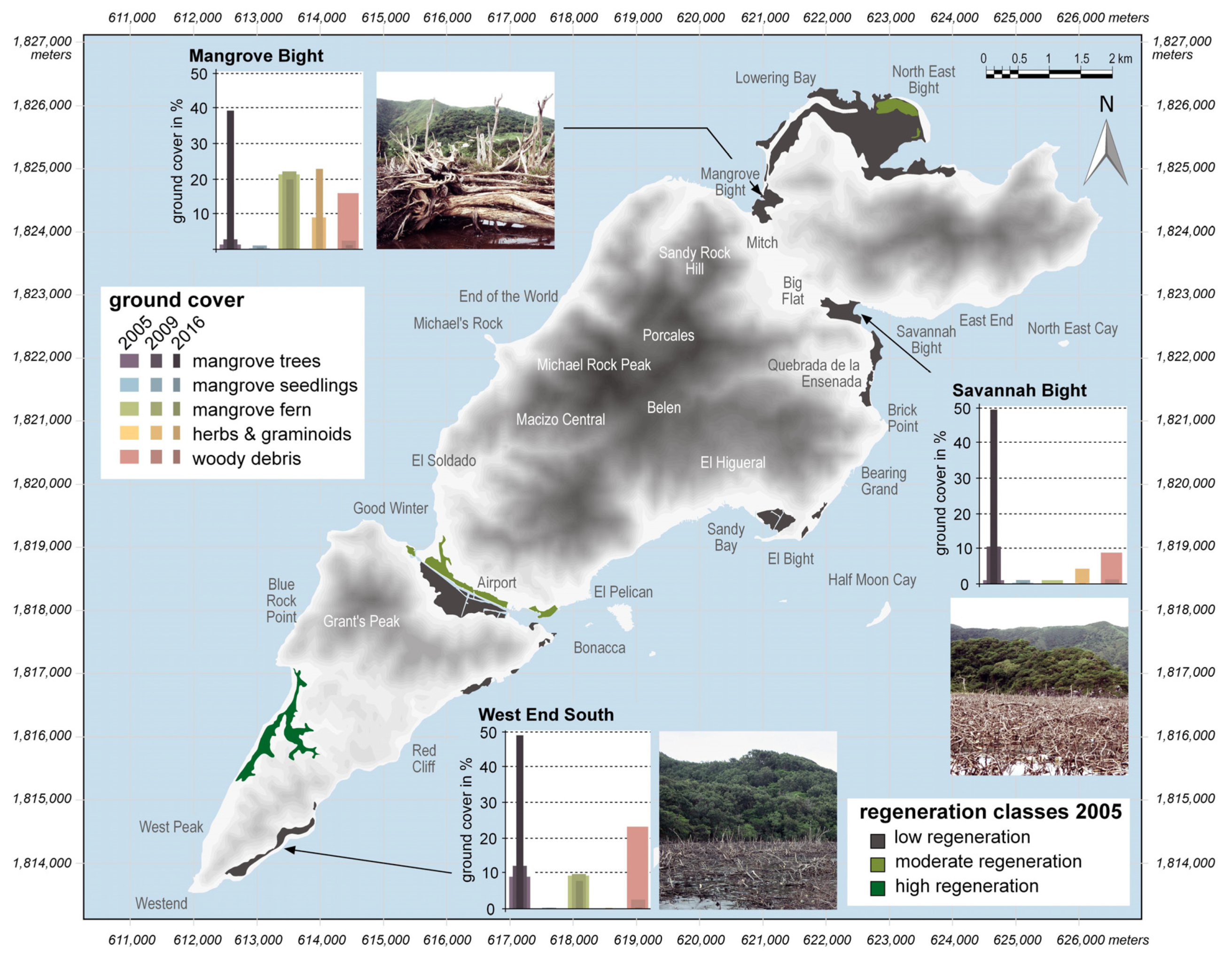
At the time of the initial survey in 2005, the three mangrove areas, MB, SB, and WES, had low groundcover of living mangrove tree species (<10%) and impacts of the hurricane forces were still prominent seven years after the storm (Figure 2; see also Table S1). Consequently, these sites were classified as low regeneration sites. Other less severely disturbed areas, in more sheltered locations on the island, were in a more advanced stage of regeneration and were accordingly classified as moderate to high regeneration sites (Figure 2) [37].
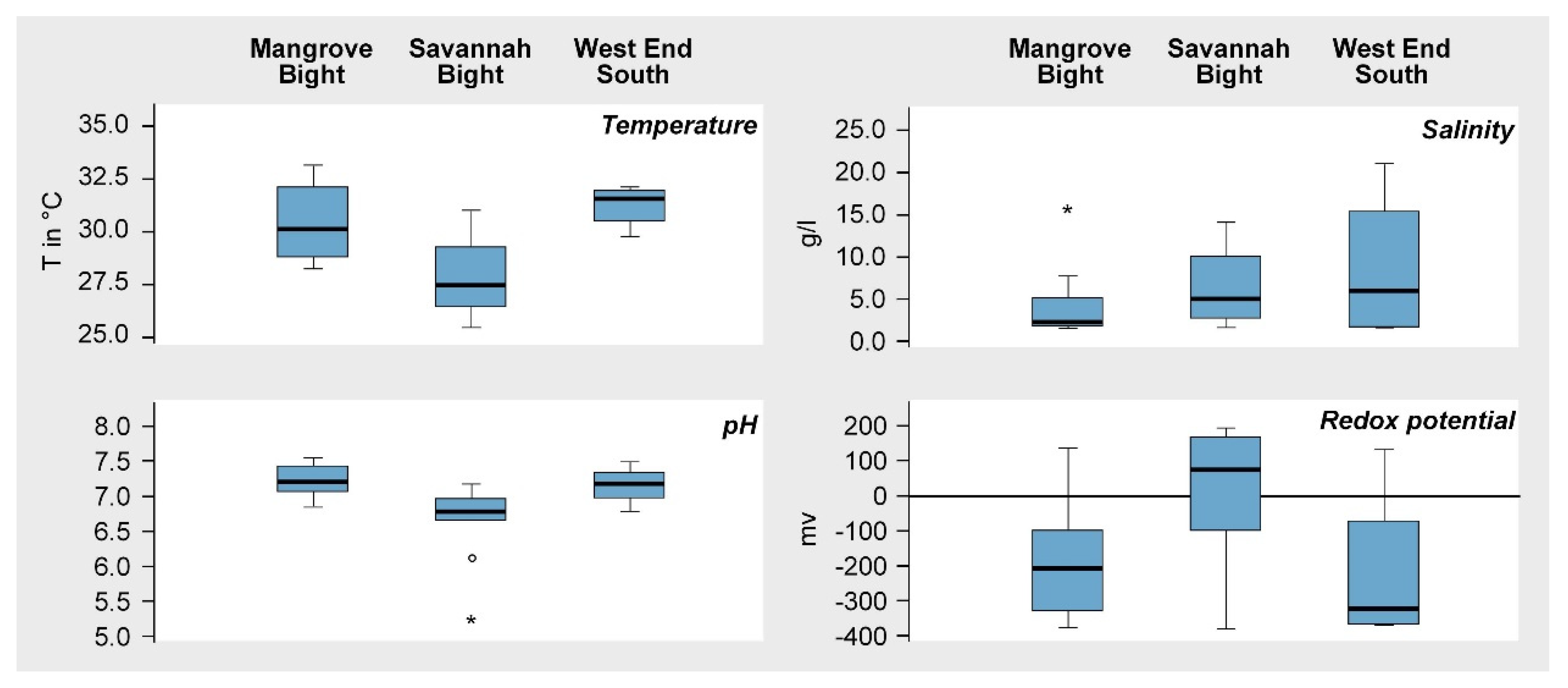
According to the measured values, soil conditions differed to some degree, both within and between areas (Figure 3). Soil temperatures varied between a minimum of 25.5 °C at SB and a maximum of 33.1 °C at MB. Median values were lowest at SB and highest at WES. The pH values measured indicate primarily near-neutral conditions, with only slight deviations from pH 7 (Figure 3) in all three areas. Salinity showed a wider range of variation between 1.6‰ (= 1.6 g/L), indicating almost nonsaline conditions at the transition to terrestrial formations in all three areas, and 21‰ (21 g/L), under more regular marine inundation. The recorded values of redox potential indicated predominantly anaerobic conditions (<100 mV), although, in all three areas, better aerated soils (>100 mV) also occur. Minimum and maximum values are fairly similar; however, the median and the interquartile range of the boxplots in Figure 3 differ between areas.
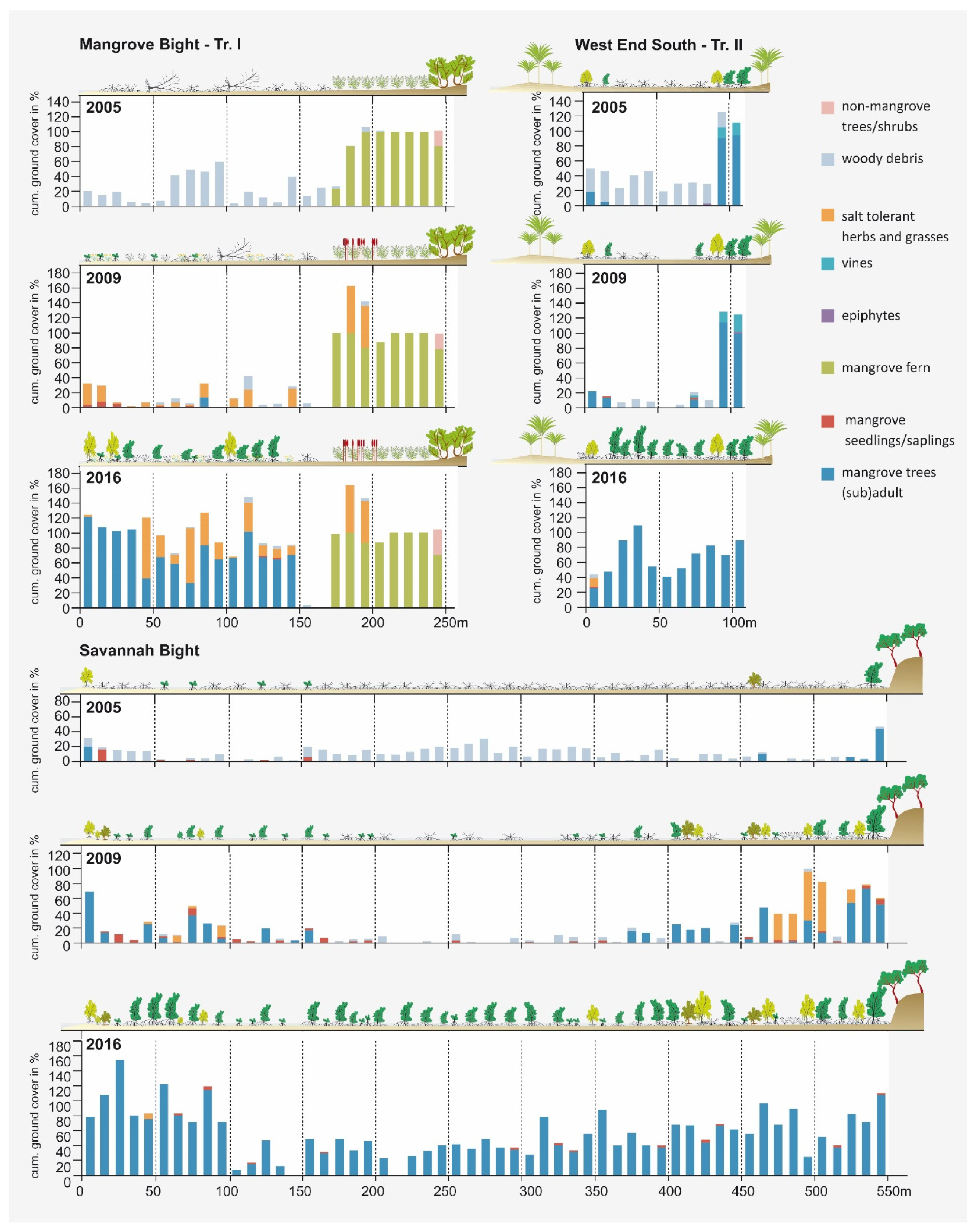
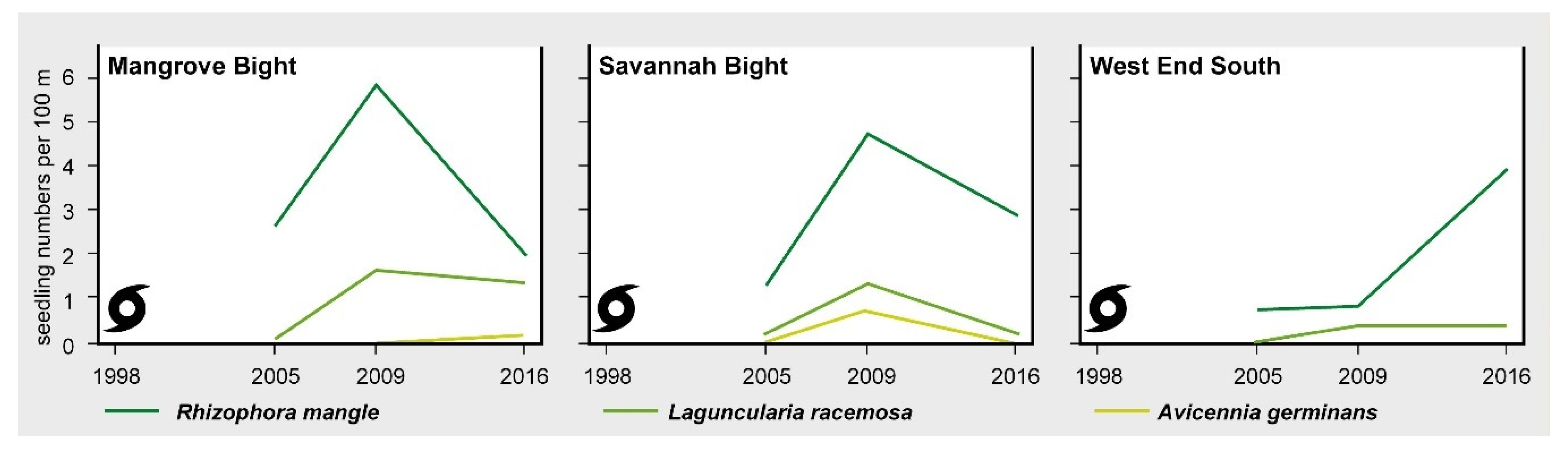
Figure 4 visualizes the vegetation development along three transects, one each in the three study areas. In 2005, a common feature of all transects was a high degree of woody debris covering the ground. Living plants were rare; only a broad belt of Acrostichum aureum was present on the landward side in Mangrove Bight and a few adult individuals of Rhizopohora mangle and Laguncularia racemosa at the edge of the mangrove areas in SB and WES, which survived the storm at protected locations. Protection was either provided by rocky escarpments (SB) or the adjoining terrestrial forests (WES) (Figure 4). Occasionally, seedlings of mangrove tree species were recorded, primarily Rhizophora mangle (14 at MB, 7 at SB, and 2 at WES), but also of Laguncularia racemosa (one each at SB and MB; Figure 5). Some of the Rhizophora mangle seedlings recorded in MB and SB might have originated from the ESNACIFOR planting campaigns in 2003 and 2004.
The survey of 2009 revealed an almost complete decay of the woody debris in all three sites (Figure 4). Other than that, the three transects showed remarkable differences in vegetation development during the preceding four-year period. WES, without any planting activities, persisted in a low regeneration state with only marginally higher ground cover of mangrove tree species and very few seedlings (2 Rhizophora mangle and 1 Laguncularia racemose; Figure 5). At MB, primarily heliophytic and salt-tolerant herbs (Sesuvium portulacastrum (L.) L., Blutaparon vermiculare (L.) Maers, Eclipta prostrata (L.) L., Cakile lanceolata (Willd.) O.E. Schulz) and grasses (Distichlis spicata (L.) Greene, Spartina spartinae (Trin.) Merr. Ex Hitchc.) invaded the area, and, within the Acrostichum aureum belt, Typha domingensis Pers. established in large numbers, while a substantial increase in the mangrove cover could not be observed (Table S1). However, along the two transects, establishment of mangrove propagules continued (30 Rhizophora mangle and 9 Laguncularia racemose; Figure 5). Most of the Rhizophora mangle propagules (a total of 22) were encountered at the seaward margin of the bay, which obviously were recently planted by the US planting initiative. In contrast, at SB, mangrove cover increased in some transect segments (see Figure 4) by the young mangrove trees, which might have originated from the ESNACIFOR planting campaign at the seaward margin of the transect, but also from natural regeneration at the landward side where propagules most likely were provided by the few surviving mangrove trees in sheltered locations. Due to greater distance to potential propagule sources at both ends of the transect and without planting, the center of the transect lacks young mangrove trees. In some segments, however, mangrove seedlings are present coming from propagule-bearing individuals at both ends of the transect (a total of 26 Rhizophora mangle, 7 Laguncularia racemose, and 4 Avicennia germinans; Figure 5). On the landward side of the transect at SB, a short section with salt-tolerant herbs (Sesuvium portulacastrum, Blutaparon vermiculare) and grasses (Distichlis spicata) was present (Figure 4).
A very distinct pattern was revealed in the survey of 2016. In all three areas, most transect segments were now covered by mangrove trees, with local ground-cover values well-above 100% due to the cumulative coverage of the different height classes. Herbaceous and grassy species predominantly disappeared at Savannah Bight but are still present to a very high degree (in particular, Blutaparon vermiculare) at Mangrove Bight. Seedling numbers decreased in MB (10 Rhizophora mangle, 7 Laguncularia racemose, and 1 Avicennia germinans) and SB (16 Rhizophora mangle, 1 Laguncularia racemose; Figure 5), probably due to shadowing effects preventing seedling establishment [50]. At WES, however, seedling numbers increased (10 Rhizophora mangle) or remained stable (1 Laguncularia racemose) (Figure 5), indicating a slightly delayed development if seedling development is dependent on a natural influx of propagules.
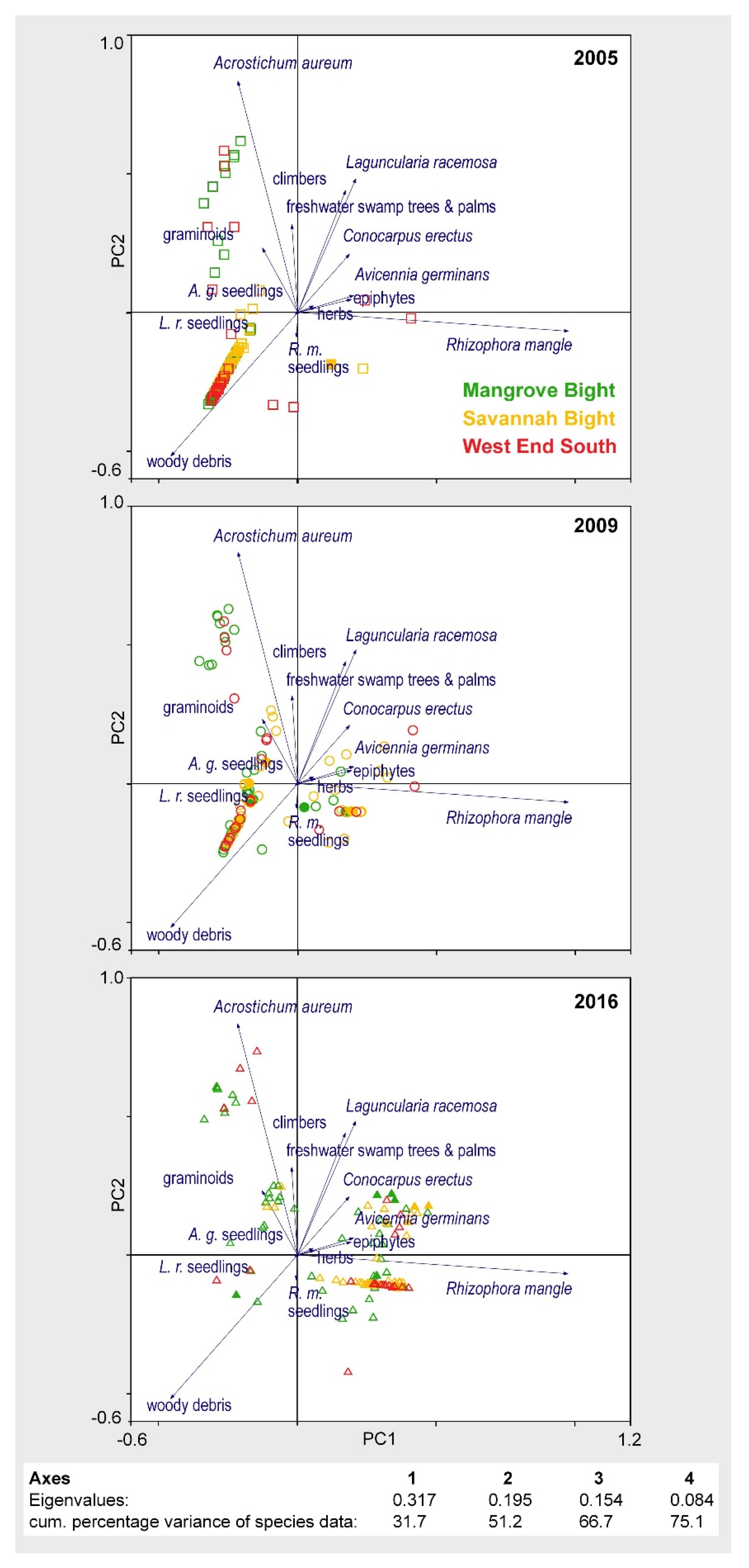
The transect resurveys revealed a high spatiotemporal heterogeneity within the dataset, i.e., vegetation development varies widely both within and between areas, as well as between sampling dates [37]. Together, Axis 1 and Axis 2 of the PCA in Figure 6 explain 51.2% of the variance in species data. The ordination space is defined by the presence of living mangrove species on the right side of the ordination plot, primarily Rhizophora mangle (correlation coefficient with Axis 1: r = 0.922) and, to a lesser extent, also Laguncularia racemosa (r = 0.446) and Avicennia germinans (r = 0.370), and by very low cover or even total absence of living mangrove species on the left side. Along the second axis, the two-dimensional space is defined by the presence of the heliophilous mangrove fern Acrostichum aureum (r = 0.855) in the upper part and by woody debris (r = −0.655) in the lower part.
In simple terms, transect segments without vital mangrove vegetation are pooled on the left side of the two PCA axes in Figure 6, while those with living mangrove tree species and/or exhibiting evidence of progressing regeneration are situated on the right side. In 2005, very few transect segments of the three study areas were to be found on the “living” side of the ordination plot, demonstrating the still very high destruction and low regeneration of the mangrove forests at MB, SB, and WES, seven years after Hurricane Mitch. MB and WES segments, with high Acrostichum aureum coverage, are arranged in the upper left quadrant; otherwise most of the transect segments are concentrated in the lower left quadrant, defined by a high amount of woody debris. By 2009, the number of transect segment located on the right side slightly increased for all the three areas, indicating a slowly ongoing regeneration—naturally and/or human-assisted—that caused some segments to shift from the left to the right side of the ordination space. A very different scenario emerged in 2016, with most segments now being located on the right side, indicating an initially delayed but continually progressing regeneration of the mangrove forests across all three study sites in Guanaja after Hurricane Mitch, regardless of reforestation measures.
4. Discussion
4.1. Mangrove Regeneration and Restoration on Guanaja after Hurricane Mitch
Regeneration of severely hurricane-disturbed mangrove areas in Guanaja shows different trajectories during the first two decades but a remarkably uniform outcome. Mangrove tree species, primarily Rhizophora mangle, recaptured the formerly disturbed areas, an effect visible in the time series analysis of NDVI (Normalized Difference Vegetation Index)-transformed satellite images as well [51]. In all of the three severely disturbed areas considered here, the mangrove cover increased only marginally during the first decade after the storm (i.e., between 2005 and 2009) but strongly in the second decade (between 2009 and 2016; see Figure 2 and Figure 4 and Table S1; repeated photographs of WES, SB, and MB for 2009 and 2016 can be found in [37]), irrespective of
- (1)
- whether no reforestation measures were undertaken, as in WES,
- (2)
- whether some planting was successful as in parts of SB and MB, or
- (3)
- whether an interim herbaceous phase was involved, as in MB.
In particular, for the latter area (MB), it was hypothesized during the 2009 resurvey that the low success of Rhizophora mangle propagule establishment (planted and natural) and the concurrent invasion of salt-tolerant herbs and grasses resulted from a permanent conversion of mangroves to treeless salt marshes. This was the dominant assumption for the severely disturbed mangroves in Guanaja earlier [52], and it was something that had been reported for other severely disturbed mangrove areas globally, too (e.g., [53,54,55,56,57]). It is attributed to a loss of “ecological memory” (seed banks, saplings, refugia, favorable soil conditions) caused by a severe disturbance that transforms a system into a different state [57].
The absence of natural recolonization, as well as the low success of the ESNACIFOR replanting campaign observed during the first survey in 2005, suggested edaphic constraints in the aftermath of Hurricane Mitch were preventing recovery [39,58]. The recorded soil characteristics, however, were well within the range reported from other mangroves globally [50,59,60]. For explaining differences in soil temperature between the three mangrove areas, reasons discussed for Jamaican mangroves might be taken into account [61]. The authors explained thermal variations there by the degree of shading and freshwater input. As shading in the “ghost forests” on Guanaja can be ignored as being responsible, the amount of freshwater input is most likely the reason for thermal differences within and between areas (Figure 3). While MB and WES do not have a substantial freshwater input, the Quebradas Big Gully and Flat Gully, draining the high central part of the island, flow into the bay of SB. The variation of pH values within and between sites was low (Figure 3), deviating only slightly from near-neutral conditions. Mangrove soils, in general, tend to be neutral or only weakly acidic due to a high sulfur content, which is, to some extent, bound with iron to form pyrites [50]. In addition, the buffering capacity of dissolved salts such as Na+ and Mg2+ often prevents soil acidification [62]. Soil desiccation, however, allows the oxidation of sulfides like pyrite, which can lead to soil acidification [50]. This might be the reason for the two outlier values at SB in Figure 3. During the field exploration in March 2009, large stretches of SB lacked inundation, being responsible for the generally higher redox values here, too. In contrast, the transects at MB and WES, for the most part, were inundated and measurements showed more typical redox conditions for mangroves due to waterlogged soils [60]. Soil salinity values, despite some variation within and between areas (Figure 3), stayed well below the salinity of seawater (35‰) due to leaching of salts during the preceding rainy season.
Edaphic conditions should not have been accountable for the low success of mangrove propagule establishment during the first decade after Hurricane Mitch, both planted ones and buoyant ones that arrived by floating. This assumption is supported by the rapid vegetation development in the severely disturbed mangroves areas during the following seven-year period after the measurements were conducted. The low progress in vegetation development during the first decade after Mitch is most likely related to the combined effects of a low number of propagule sources in the close surroundings of the disturbed areas, which would have been able to push forward natural regeneration, and the high mortality rate of the planted propagules due to planting in unsuitable locations and predation by crabs. Locally, more than 90% of the planted mangrove propagules did not survive (personal communication Scott Duncan). Only later, attempts with Rhizophora mangle seedlings raised in nurseries slightly reduced the mortality rates. The high mortality rates of planted propagules on Guanaja are well in line with findings of other mangrove restoration projects [28,29,30,31,32]. In spite of low success of planting in Guanaja, some of the planted propagules and/or seedlings made it into a propagule-bearing stage during subsequent years after planting, as did a few of the naturally arrived, waterborne propagules. Once propagule-bearing individuals were present, a state commonly achieved within a half-decade [63], the number of available propagules and, thus, the chance for successful establishment sharply increases, paving the way for a swift rejuvenation of the mangrove forest.
The fact that mangroves are able to regenerate without human intervention proves the high resilience of this ecosystem, even if the tree species (in particular, Rhizophora mangle, with its delicate stilt roots) are not very resistant to periodically occurring storm events. The process of regeneration might be accelerated by reforestation measures, which happened at MB and SB on the seaward fringe, but the difference from the naturally regenerating mangroves in WES was minimal. In addition, the coexistence of salt-tolerant herbs and grasses and small mangrove trees in MB proves that herbaceous and graminoid salt marsh species do not prevent the establishment of mangrove propagules; rather, they enable mutualistic effects and facilitate mangrove recovery. Similar patterns are reported from other disturbed mangrove areas within the Caribbean and beyond [64,65,66,67].
4.2. Mangrove Regeneration and Restoration—A More Global Perspective
Mangroves are one of the most threatened ecosystems in the world, primarily from direct anthropogenic impacts (e.g., conversion for aquaculture, urbanization, pollution) but, more recently (and increasingly), also by the effects of a changing climate (e.g., sea-level rise, drought, more high-intensity storms) [68,69,70,71]. As the negative effects of a continuously shrinking mangrove area in terms of the services, that these ecosystems provide to the local and global human population, are increasingly recognized, many initiatives have been started in the recent past to rehabilitate or restore disturbed and/or degraded mangroves [72,73]. While there are some positive examples of successful rehabilitation or restoration projects (e.g., [31,74,75,76]), most of these labor-intensive and sometimes costly efforts fail [27,28,29,30,31,32,77,78].
Replanting has the potential to accelerate the recovery of vital mangroves in the aftermath of severe disturbances [76]. However, replanted mangroves remain mere plantations rather than ecologically integrated ecosystems, in particular, if the focus is on the tree species only [28]. Compared to naturally regenerating forests, planted forests generally have a lower structural complexity. Structural complexity, however, is key for the long-term stability of mangrove forests [79]. Often, only very few (or even one) fast-growing species are considered for replanting—in the Neotropics, primarily Rhizophora mangle—representing the afforestation of monocultures rather than true ecosystem restoration [28]. In the long run, this will lead to an impaired secondary succession [80]. In addition, restoration is commonly done by the planting of locally (or nearby) collected propagules, which threatens genetic diversity [81]. Despite these shortcomings, there is nevertheless one tremendous benefit of reforestation campaigns if local people are involved: the chance to raise awareness about the value of mangroves. Participation of the general public in mangrove restoration projects not only helps to restore disturbed mangrove forests and reinstate important ecosystem services but also bolsters the social capital of communities [82].
However, not all degraded or disturbed mangrove areas need reforestation measures. Depending on the grade of deterioration, they might be able to recover naturally, which commonly leads to structurally and ecologically more diverse forests [76], which are more robust, can withstand natural disturbances better, and provide more diverse ecological functions and services. Based on the observations on the Island of Guanaja, where human-assisted (MB, SB) and natural regeneration (WES) have had the same outcome, labor-intensive and sometimes costly replantation of disturbed mangroves can be critically questioned. Due to the high mortality rates of planted propagules—which holds true for Guanaja too—the positive effects for enhancing the grow-back of mangrove trees are limited. Naturally regenerating mangroves, in contrast, might exhibit a certain time lag (Figure 5) if only a few diaspore sources survived in the surrounding. However, as soon as established individuals have grown up into a propagule-bearing stage, a quick return to viable forests takes place, proving the high resilience of mangrove forests. Once individuals have successfully established and, depending on the species, aerial or stilt-roots are formed, the trapping of floating propagules is supported, and the succession and further development of mangrove stands are enhanced. In contrast, if no adult propagule-bearing trees are left, the chance of seedlings establishment by an inflow of buoyant propagules from outside is low, and it might take a long time until the first seedlings are established. In those cases, planting operations might be helpful to avoid negative developments caused by a long-lasting absence of mangroves (e.g., coastal erosion, sedimentation of coral reefs and seagrass meadows, and peat collapse). Even more important than active planting is to ensure that the hydroecological conditions for a natural recolonization by mangrove species are maintained or restored. In particular, the impeding effects of infrastructure such as roads, dikes, and bridges have to be taken into account and dismantled if necessary [83].
5. Conclusions
This paper suggests that replanting should not be considered the primary means to restore mangrove forests after a severe disturbance. A comparison between the natural and the human-assisted regeneration of mangroves after Hurricane Mitch in Guanaja reveals no substantial differences concerning the success of the rejuvenation of the mangroves. On the contrary, despite a severe disturbance such as Hurricane Mitch, the mangroves proved to be highly resilient, even if not being particularly able to withstand the impacts of the storm [37]. Mangroves regenerate via different pathways, which might take longer in severely disturbed sites than in less disturbed sites. Planting, if successful, might slightly enhance the pace of regeneration, but, without human assistance, the regrowth of mangrove trees in severely disturbed mangrove areas in Guanaja during the first two decades after the storm remained the same. The high mortality of planted mangrove seedlings and the drawbacks of restoration measures for forest structure and ecosystem functioning make the labor-intensive and sometimes costly reforestation measures in disturbed mangroves questionable, and the need for human intervention should be carefully assessed in each case. In many cases, these programs neither fulfill their promise concerning ecological integrity and biodiversity nor are they sustainable if not supported by local people. Natural regeneration, in contrast, commonly leads to structurally and ecologically more diverse and robust forests, providing ecosystem services. In this respect, mangrove forests do not behave differently from other tropical forests, where natural regeneration has been found to be ecologically superior to active restoration from both economic (low cost vs. high cost) and ecological perspectives (ecosystem functioning and services, structural heterogeneity, species richness, growth rates, litterfall) [84]. In addition, major challenges for mangrove restoration can emanate from the poor consensus of the local people concerning the restoration goals and a lack of confidence between the actors involved, potentially leading to social opposition even before restoration measures are actually implemented [82,85,86]. This can be easily avoided by allowing disturbed sites to regenerate naturally. Human beings are notoriously impatient creatures who tend to activism. In the case of mangrove regeneration after natural disturbances, it could be better and easier to be patient, to ensure appropriate conditions for natural colonization by waterborne propagules instead of planting and let nature do its work [83].
Supplementary Materials
The following are available online at https://www.mdpi.com/1999-4907/11/10/1068/s1, Table S1: Annotated species list (including the family and life-form the species belongs to) of plant species recorded at MB, SB, and WES at different sampling dates. Figures are mean ground cover values. Figure S1: Schematic sketch of the line-transects used for vegetation sampling; sampled extents with a resolution of 10 cm (= units U) of each species (here, exemplarily for three species, A, B, and C) were translated to ground cover values in % per 10 m-transect segments afterward (taken from [37]).
Funding
Field data collection for this study was supported by the German Research Foundation (Deutsche Forschungsgemeinschaft (DFG)) under grants # FI 1254/2-1 and # FI 1254/4-1.
Acknowledgments
Field surveys were accompanied by Friederike Grüninger (University of Passau); Marielle Ochoa of BICA (Bay Islands Conservation Association), and Scott Duncan of the Guanaja Mangrove Restoration Project provided valuable insight on mangrove planting activities and their success in Guanaja; Aditya Ghosh (South Asia Institute, University of Heidelberg, Germany, and Jindal Global University, India) gave valuable linguistic advice. The author would also like to acknowledge valuable recommendations made by two anonymous reviewers, who greatly improved the outcome of the manuscript.
Conflicts of Interest
The author declares no conflict of interest.
References
- Ewel, K.C.; Twilley, R.R.; Ong, J.E. Different kinds of mangrove forests provide different goods and services. Glob. Ecol. Biogeogr. Lett. 1998, 7, 83–94. [Google Scholar] [CrossRef]
- Walters, B.B.; Rönnbäck, P.; Kovacs, J.M.; Crona, B.; Hussain, S.A.; Badola, R.; Primavera, J.H.; Barbier, E.; Dahdouh-Guebas, F. Ethnobiology, socioeconomics and management of mangrove forests: A review. Aquat. Bot. 2008, 89, 220–236. [Google Scholar] [CrossRef] [Green Version]
- Spalding, M.; Kainuma, M.; Collins, L. World Atlas of Mangroves; Earthscan: London, UK; Washington, DC, USA, 2010. [Google Scholar]
- Barbier, E.B.; Hacker, S.D.; Kennedy, C.; Koch, E.W.; Stier, A.C.; Silliman, B.R. The value of estuarine and coastal ecosystem services. Ecol. Monogr. 2011, 81, 169–193. [Google Scholar] [CrossRef]
- Salem, M.E.; Mercer, D.E. The economic value of mangroves: A meta-analysis. Sustainability 2012, 4, 359–383. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Primavera, J.H.; Dahdouh-Guebas, F.; McKee, K.; Bosire, J.O.; Cannicci, S.; Diele, K.; Fromard, F.; Koedam, N.; Marchand, C.L.; et al. Ecological role and services of tropical mangrove ecosystems: A reassessment. Glob. Ecol. Biogeogr. 2014, 23, 726–743. [Google Scholar] [CrossRef]
- UNEP. The Importance of Mangroves to People: A Call to Action; van Bochove, J., Sullivan, E., Nakamura, T., Eds.; United Nations Environment Programme World Conservation Monitoring Centre: Cambridge, UK, 2014. [Google Scholar]
- Cummings, A.R.; Shah, M. Mangroves in the global climate and environmental mix. Geogr. Compass 2017, e12353. [Google Scholar] [CrossRef]
- Huxham, M.; Dencer-Brown, A.; Diele, K.; Kathiresan, K.; Nagelkerken, I.; Wanjiru, C. Mangroves and people: Local ecosystem Services in a changing climate. In Mangrove Ecosystems: A Global Biogeographic Perspective—Structure, Function, and Services; Rivera-Monroy, V.H., Lee, S.Y., Kristensen, E., Twilley, R.R., Eds.; Springer: Cham, Switzerland, 2017; pp. 245–274. [Google Scholar]
- Barbier, E.B.; Koch, E.M.; Silliman, B.R.; Hacker, S.D.; Wolanski, E.; Primavera, J.; Granek, E.F.; Polasky, S.; Aswani, S.; Cramer, L.A.; et al. Coastal Ecosystem-Based Management with Nonlinear Ecological Functions and Values. Science 2008, 319, 321–323. [Google Scholar] [CrossRef]
- Costanza, R.; de Groot, R.; Sutton, P.; van der Ploeg, S.; Anderson, J.; Kubiszewski, I.; Farber, S.; Turner, R.K. Changes in the global value of ecosystem services. Glob. Environ. Chang. 2014, 26, 152–158. [Google Scholar] [CrossRef]
- Marois, D.E.; Mitsch, W.J. Coastal protection from tsunamis and cyclones provided by mangrove wetlands—A review. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2015, 11, 71–83. [Google Scholar] [CrossRef]
- Danielsen, F.; Sørensen, M.K.; Olwig, M.F.; Selvam, V.; Parish, F.; Burgess, N.D.; Hiraishi, T.; Karunagaran, V.M.; Rasmussen, M.S.; Hansen, L.B.; et al. The Asian Tsunami: A protective role for coastal vegetation. Science 2005, 310, 643. [Google Scholar] [CrossRef] [Green Version]
- Kathiresan, K.; Rajendran, N. Coastal mangrove forests mitigated tsunami. Estuar. Coast. Shelf Sci. 2005, 65, 601–606. [Google Scholar] [CrossRef]
- Barbier, E.B. Natural barriers to natural disasters: Replanting mangroves after the tsunami. Front. Ecol. Environ. 2006, 4, 124–131. [Google Scholar] [CrossRef]
- Barbier, E.B. The protective service of mangrove ecosystems: A review of valuation methods. Mar. Pollut. Bull. 2016, 109, 676–681. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Vincent, J.R. Mangroves protected villages and reduced death toll during Indian super cyclone. PNAS 2009, 106, 7357–7360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandilyan, S.; Kathiresan, K. Mangroves as bioshield: An undisputable fact. Ocean Coast. Manag. 2005, 103, 94–96. [Google Scholar] [CrossRef]
- Hochard, J.P.; Hamilton, S.; Barbier, E.B. Mangroves shelter coastal economic activity from cyclones. PNAS 2019, 116, 12232–12237. [Google Scholar] [CrossRef] [Green Version]
- Spalding, M.; McIvor, A.; Tonneijck, F.H.; Tol, S.; van Eijk, P. Mangroves for coastal defence. In Guidelines for Coastal Managers & Policy Makers; Wetlands International, Wageningen, Neherlands, and The Nature Conservancy: Arlington, TX, USA, 2014; p. 42. [Google Scholar]
- Krauss, K.W.; Osland, M.J. Tropical cyclones and the organization of mangrove forests: A review. Ann. Bot. 2020, 125, 213–234. [Google Scholar] [CrossRef]
- Walton, M.; Samonte-Tan, G.; Primavera, J.; Edwards-Jones, G.; Le Vay, L. Are mangroves worth replanting? The direct economic benefits of a community-based reforestation project. Environ. Conserv. 2006, 33, 335–343. [Google Scholar] [CrossRef]
- Primavera, J.H.; Rollon, R.N.; Samson, M.S. The pressing challenges of mangrove rehabilitation: Pond reversion and coastal protection. In Ecohydrology and Restoration; Chicharo, L., Zalewski, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2011; pp. 217–244. [Google Scholar]
- Andradi-Brown, D.A.; Howe, C.; Mace, G.M.; Knight, A.T. Do mangrove forest restoration or rehabilitation activities return biodiversity to pre-impact levels? Environ. Evid. 2013, 2, 20. [Google Scholar] [CrossRef] [Green Version]
- López-Portillo, J.; Lewis III, R.R.; Saenger, P.; Rovai, A.; Koedam, N.; Dahdouh-Guebas, F.; Agraz-Hernández, C.; Rivera-Monroy, V.H. Mangrove Forest Restoration and Rehabilitation. In Mangrove Ecosystems: A Global Biogeographic Perspective—Structure, Function, and Services; Rivera-Monroy, V.H., Lee, S.Y., Kristensen, E., Twilley, R.R., Eds.; Springer: Cham, Switzerland, 2017; pp. 301–345. [Google Scholar]
- Carugati, L.; Gatto, B.; Rastelli, E.; Lo Martire, M.; Coral, C.; Greco, S.; Danovaro, R. Impact of mangrove forests degradation on biodiversity and ecosystem functioning. Sci. Rep. 2018, 8, 13298. [Google Scholar] [CrossRef] [Green Version]
- Ellison, A.M.; Felson, A.J.; Friess, D.A. Mangrove Rehabilitation and Restoration as Experimental Adaptive Management. Front. Mar. Sci. 2020, 7, 327. [Google Scholar] [CrossRef]
- Ellison, A.M. Mangrove Restoration: Do we know enough? Restor. Ecol. 2000, 8, 219–229. [Google Scholar] [CrossRef]
- Iftekhar, M.S. Functions and development of reforested mangrove areas: A review. Int. J. Biodivers. Sci. Manag. 2008, 4, 1–14. [Google Scholar] [CrossRef]
- IUCN Mass Mangrove Restoration: Driven by Good Intentions But Offering Limited Results. 2017. Available online: https://www.iucn.org/news/forests/201702/mass-mangrove-restoration-driven-good-intentions-offering-limited-results (accessed on 10 May 2020).
- Kodikara, K.A.S.; Mukherjee, N.; Jayatissa, L.P.; Dahdouh-Guebas, F.; Koedam, N. Have mangrove restoration projects worked? An in-depth study in Sri Lanka. Restor. Ecol. 2017, 25, 705–716. [Google Scholar] [CrossRef]
- Lee, S.Y.; Hamilton, S.; Barbier, E.B.; Primavera, J.; Lewis, R.R. Better restoration policies are needed to conserve mangrove ecosystems. Nat. Ecol. Evol. 2019, 3, 870–872. [Google Scholar] [CrossRef] [PubMed]
- Ross, M.S.; Ruiz, P.L.; Telesnicki, G.J.; Meeder, J.F. Estimating above-ground biomass and production in mangrove communities of Biscayne national Park, Florida (USA). Wetl. Ecol. Manag. 2001, 9, 27–37. [Google Scholar] [CrossRef]
- Alongi, D.M. Mangrove forests: Resilience, protection from tsunamis, and responses to global climate change. Estuar. Coast. Shelf Sci. 2008, 76, 1–13. [Google Scholar] [CrossRef]
- Onrizal, O.; Ahmad, A.G.; Mansour, M. Assessment of Natural Regeneration of Mangrove Species at Tsunami Affected Areas in Indonesia and Malaysia. IOP Conf. Ser. Mater. Sci. Eng. 2017, 180, 012045. [Google Scholar] [CrossRef]
- Bayraktarov, E.; Saunders, M.I.; Abdullah, S.; Mills, M.; Beher, J.; Possingham, H.P.; Mumby, P.J.; Lovelock, C.E. The cost and feasibility of marine coastal restoration. Ecol. Appl. 2016, 26, 1055–1074. [Google Scholar] [CrossRef]
- Fickert, T. Better resilient than resistant—Regeneration Dynamics of storm-disturbed mangrove forests on the Bay Island of Guanaja (Honduras) during the first two decades after Hurricane Mitch (October 1998). Diversity 2018, 10, 8. [Google Scholar] [CrossRef] [Green Version]
- Pasch, R.J.; Avila, L.A.; Guiney, J.L. Atlantic Hurricane Season of 1998. Mon. Weather Rev. 2001, 129, 3085–3123. [Google Scholar] [CrossRef] [Green Version]
- Cahoon, D.R.; Hensel, P.; Rybczyk, J.; McKee, K.; Proffitt, C.E.; Perez, B. Mass tree mortality leads to mangrove peat collapse at Bay Islands, Honduras after Hurricane Mitch. J. Ecol. 2003, 91, 1093–1105. [Google Scholar] [CrossRef]
- Guiney, J.L.; Lawrence, M.B. Preliminary Report: Hurricane Mitch 22 October–5 November 1998; National Hurricane Center: Miami, FL, USA, 1999. [Google Scholar]
- Vanselow, K.A.; Kolb, M.; Fickert, T. Destruction and Regeneration of Terrestrial, Littoral and Marine Ecosystems on the Island of Guanaja/Honduras seven years after Hurricane Mitch. Erdkunde 2007, 61, 358–371. [Google Scholar] [CrossRef] [Green Version]
- Fickert, T.; Grüninger, F. Floristic zonation, vegetation structure, and plant diversity patterns within a Caribbean mangrove and swamp forest on the Bay Island of Utila (Honduras). Ecotropica 2010, 16, 73–92. [Google Scholar]
- Valade, I.; Grelot, J.; Laurent, J.M. Esquema Director de Manejo Ambietal de las Islas de la Bahía, Honduras; Repiublica de Honduras: Islas de La Bahia, Honduras, 2002. [Google Scholar]
- Doyle, T.W.; Michot, T.C.; Roetker, F.; Sullivan, J.; Melder, M.; Handley, B.; Balmat, J. Hurricane Mitch: Landscape Analysis of Damaged Forest Resources of the Bay Islands and Caribbean Coast of Honduras, USGS Open File Report 03-175. 2002.
- Mueller-Dombois, D.; Ellenberg, H. Aims and Methods of Vegetation Ecology; John Wiley & Sons: New York, NY, USA, 1974. [Google Scholar]
- Cintrón, G.; Schaeffer-Novelli, Y. Methods for studying mangrove structure. In The Mangrove Ecosystem: Research Methods; Snedaker, S.C., Snedaker, J.G., Eds.; UNESCO: Paris, France, 1984; pp. 91–113. [Google Scholar]
- Balick, M.J.; Nee, M.H.; Daniel, E.A. Checklist of the Vascular Plants of Belize; Memoirs of the New York Botanical Garden 85: New York, NY, USA, 2000. [Google Scholar]
- Ter Braak, C.J.F.; Šmilaur, P. CANOCO for Windows Version 4.5; Biometris, Plant Research International: Wageningen, The Netherlands, 2002. [Google Scholar]
- Lepš, J.; Šmilauer, P. Multivariate Analysis of Ecological Data Using CANOCO; Cambridge University Press: New York, NY, USA, 2003. [Google Scholar]
- Saenger, P. Mangrove Ecology, Silviculture and Conservation; Kluwer: Dordrecht, The Netherlands; Boston, MA, USA; London, UK, 2002. [Google Scholar]
- Vanselow, K.A. Die Ruhe nach dem Sturm—Eine Zeitreihenanalyse zur Entwicklung der Vegetationsbedeckung auf der Karibikinsel Guanaja nach Hurrikan Mitch. Mitt. Fränkischen Geogr. Ges. 2018, 63, 111–121. [Google Scholar]
- Winograd, M. Sustainability and vulnerability indicators for decision making: Lessons learned from Honduras. Int. J. Sustain. Dev. 2007, 10, 93–105. [Google Scholar] [CrossRef]
- Wanless, H.R.; Parkinson, R.W.; Tedesco, L.P. Sea level control on stability of Everglades wetlands. In Everglades: The Ecosystem and Its Restoration; Davis, S.M., Ogden, J.C., Eds.; St. Lucie Press: Boca Raton, FL, USA, 1994; pp. 199–223. [Google Scholar]
- Bandeira, S.; Balidy, H. Limpopo Estuary Mangrove Transformation, Rehabilitation and Management. In Estuaries: A Lifeline of Ecosystem Services in the Western Indian Ocean; Diop, S., Scheren, P., Machiwa, J., Eds.; Springer: Heidelberg/Berlin, Germany, 2016; pp. 227–237. [Google Scholar]
- Mbense, S.; Rajkaran, A.; Bolosha, U.; Adams, J. Rapid colonization of degraded mangrove habitat by succulent salt marsh. S. Afr. J. Bot. 2016, 107, 129–136. [Google Scholar] [CrossRef]
- Asbridge, E.; Lucas, R.; Rogers, K.; Accad, A. The extent of mangrove change and potential for recovery following severe Tropical Cyclone Yasi, Hinchinbrook Island, Queensland, Australia. Ecol. Evol. 2018, 8, 10416–10434. [Google Scholar] [CrossRef] [Green Version]
- Schaeffer-Novelli, Y.; Soriano-Sierra, E.; Vale, C.; Bernini, E.; Rovai, A.; Pinheiro, M.; Schmidt, A.; de Almeida, R.; Júnior, C.; Menghini, R.; et al. Climate changes in mangrove forests and salt marshes. Braz. J. Oceanogr. 2016, 64, 37–52. [Google Scholar] [CrossRef] [Green Version]
- McKee, K.L.; McGinnis II, T.E. Hurricane Mitch: Effects on Mangrove Soil Characteristics and Root Contributions to Soil Stabilization; USGS Open File Report 03-178: Lafayette, IN, USA, 2002. [Google Scholar]
- Lacerda, L.D. Mangrove Ecosystems: Function and Management; Springer: Berlin, Germany, 2002. [Google Scholar]
- Hossain, M.D.; Nuruddin, A.A. Soil and mangrove: A Review. J. Environ. Sci. Technol. 2016, 9, 198–207. [Google Scholar] [CrossRef] [Green Version]
- McDonald, K.O.; Webber, D.F.; Webber, M.K. Mangrove forest structure under varying environmental conditions. Bull. Mar. Sci. 2003, 73, 491–505. [Google Scholar]
- Hseu, Z.Y.; Chen, Z.S. Monitoring the changes of redox potential, pH and electric conductivity of the mangrove soils in Northern Taiwan. Proc. Natl. Sci. Counc. Repub. China Part B 2000, 24, 143–150. [Google Scholar]
- Imbert, D.; Rousteau, A.; Scherrer, P. Ecology of Mangrove Growth and Recovery in the Lesser Antilles: State of Knowledge and Basis for Restoration Projects. Restor. Ecol. 2000, 8, 230–236. [Google Scholar] [CrossRef]
- Milbrandt, E.C.; Tinsley, M.N. The role of saltwort (Batis maritima L.) in regeneration of degraded mangroive forests. Hydrobiologia 2006, 568, 369–377. [Google Scholar] [CrossRef]
- McKee, K.L.; Rooth, J.E.; Feller, I.C. Mangrove recruitment after forest disturbance is facilitated by herbaceous species in the Caribbean. Ecol. Appl. 2007, 17, 1678–1693. [Google Scholar] [CrossRef] [PubMed]
- González, C.; Urrego, L.E.; Martínez, J.I.; Polanía, J.; Yokoyama, Y. Mangrove dynamics in the southwestern Caribbean since the ‘Little Ice Age’: A history of human and natural disturbances. Holocene 2010, 20, 849–861. [Google Scholar] [CrossRef]
- Biswas, S.R.; Khan, M.S.I.; Mallik, A.U. Invaders’ control on post-disturbance succession in coastal mangroves. J. Plant Ecol. 2012, 5, 157–166. [Google Scholar] [CrossRef]
- Polidoro, B.A.; Carpenter, K.E.; Collins, L.; Duke, N.C.; Ellison, A.M.; Ellison, J.C.; Farnsworth, E.J.; Fernando, E.S.; Kathiresan, K.; Koedam, N.E.; et al. The Loss of Species: Mangrove extinction Risk and Geographic Areas of global concern. PLoS ONE 2010, 5, e10095. [Google Scholar] [CrossRef]
- Chowdhury, R.R.; Uchida, E.; Chen, L.; Osorio, V.; Yoder, L. Anthropogenic Drivers of Mangrove Loss. Patterns and Implications for Livelihoods. In Mangrove Ecosystems: A Global Biogeographic Perspective—Structure, Function, and Services; Rivera-Monroy, V.H., Lee, S.Y., Kristensen, E., Twilley, R.R., Eds.; Springer: Cham, Switzerland, 2017; pp. 275–300. [Google Scholar]
- Feller, I.C.; Friess, D.A.; Krauss, K.W.; Lewis III, R.R. The state of the world’s mangroves in the 21st century under climate change. Hydrobiologia 2017, 803, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Thomas, N.; Lucas, R.; Bunting, P.; Hardy, A.; Rosenqvist, A.; Simard, M. Distribution and drivers of global mangrove forest change, 1996–2010. PLoS ONE 2017, 12, e0179302. [Google Scholar] [CrossRef] [Green Version]
- Lewis, R.R. Ecological engineering for successful management and restoration of mangrove forests. Ecol. Eng. 2005, 24, 403–418. [Google Scholar] [CrossRef]
- Dale, P.E.R.; Knight, J.M.; Dwyer, P.G. Mangrove rehabilitation: A review focusing on ecological and institutional issues. Wetl. Ecol. Manag. 2014, 22, 587–604. [Google Scholar] [CrossRef] [Green Version]
- Rey, J.R.; Carlson, D.B.; Brockmeyer Jr., R. E. Coastal wetland management in Florida: Environmental concerns and human health. Wetl. Ecol. Manag. 2012, 20, 197–211. [Google Scholar] [CrossRef]
- Brown, B.; Fadilla, R.; Nurdin, Y.; Soulsby, I.; Ahmad, R. Community based ecological mangrove rehabilitation (CBEMR) in Indonesia. Sapiens 2014, 7, 53–64. [Google Scholar]
- Ferreira, A.C.; Ganade, G.; de Attayde, J.L. Restoration versus natural regeneration in a neotropical mangrove: Effects on plant biomass and crab communities. Ocean Coast. Manag. 2015, 110, 38–45. [Google Scholar] [CrossRef]
- Bosire, J.O.; Dahdouh-Guebas, F.; Walton, M.; Crona, B.I.; Lewis, R.R.; Field, C.; Kairo, J.G.; Koedam, N. Functionality of restored mangroves: A review. Aquat. Bot. 2008, 89, 251–259. [Google Scholar] [CrossRef] [Green Version]
- Primavera, J.; Esteban, J. A review of mangrove rehabilitation in the Philippines: Successes, failures and future prospects. Wetl. Ecol. Manag. 2008, 16, 345–358. [Google Scholar] [CrossRef]
- Barnuevo, A.; Asaeda, T.; Sanjaya, K.; Kanesaka, Y.; Fortes, M. Drawbacks of mangrove rehabilitation schemes: Lessons learned from the large-scale mangrove plantations. Estuar. Coast. Shelf Sci. 2017, 198, 432–437. [Google Scholar] [CrossRef]
- Rovai, A.; Soriano-Sierra, E.; Pagliosa, P.; Cintrón, G.; Schaeffer-Novelli, Y.; Menghini, R.; Coelho-Jr, C.; Horta, P.; Lewis, R.; Simonassi, J.; et al. Secondary succession impairment in restored mangroves. Wetl. Ecol. Manag. 2012, 20, 447–459. [Google Scholar] [CrossRef]
- Granado, R.; Pinto Neta, L.C.; Nunes-Freitas, A.F.; Voloch, C.M.; Lira, C. Assessing Genetic Diversity after Mangrove Restoration in Brazil: Why Is It So Important? Diversity 2018, 10, 27. [Google Scholar] [CrossRef] [Green Version]
- Valenzuela, R.B.; Yeo-Chang, Y.; Park, M.S.; Chun, J.N. Local People’s Participation in Mangrove Restoration Projects and Impacts on Social Capital and Livelihood: A Case Study in the Philippines. Forests 2020, 11, 580. [Google Scholar] [CrossRef]
- Babak, K.; Hashim, R. Mangrove restoration without planting. Ecol. Eng. 2011, 37, 387–391. [Google Scholar]
- Crouzeilles, R.; Ferreira, M.S.; Chazdon, R.L.; Lindenmayer, D.B.; Sansevero, J.B.B.; Monteiro, L.; Iribarrem, A.; Latawiec, A.E.; Strassburg, B.B.N. Ecological restoration success is higher for natural regeneration than for active restoration in tropical forests. Sci. Adv. 2017, 3, e1701345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schönig, D. Resilience-Based Restoration of Mangrove Social-Ecological Systems—A Case Study from the Gulf of Guayaquil, Ecuador. Master’s Thesis, Master thesis submitted in partial fulfillment of the requirements for the degree of Master of Science in Forest Ecology and Management, University Freiburg, Freiburg, Germany, 2014. [Google Scholar]
- Romañach, S.; Deangelis, D.; Koh, H.; Li, Y.; Teh, S.; Sulaiman, R.; Zhai, L. Conservation and restoration of mangroves: Global status, perspectives, and prognosis. Ocean Coast. Manag. 2018, 154, 72–82. [Google Scholar] [CrossRef]
Figure 1.
Shaded relief of Guanaja (SRTM data); highlighted in green are mangrove areas; those observed in the long-term monitoring project [37] are labeled in bold letters; the three most severely disturbed ones considered in this paper are labeled in red (Mangrove Bight (MB), Savannah Bight (SB), West End South (WES)); the climate chart of the neighboring island of Roatan displays the overall climatic conditions of the Bay Islands.
Figure 1.
Shaded relief of Guanaja (SRTM data); highlighted in green are mangrove areas; those observed in the long-term monitoring project [37] are labeled in bold letters; the three most severely disturbed ones considered in this paper are labeled in red (Mangrove Bight (MB), Savannah Bight (SB), West End South (WES)); the climate chart of the neighboring island of Roatan displays the overall climatic conditions of the Bay Islands.

Figure 2.
Shaded relief of Guanaja (SRTM data) with the categorization of mangrove regeneration in 2005; bar plots visualize the development of mean ground cover of certain functional groups at MB, SB, and WES during the study period; photographs illustrate the degree of disturbance seven years after Hurricane Mitch (adapted and modified from [37]).
Figure 2.
Shaded relief of Guanaja (SRTM data) with the categorization of mangrove regeneration in 2005; bar plots visualize the development of mean ground cover of certain functional groups at MB, SB, and WES during the study period; photographs illustrate the degree of disturbance seven years after Hurricane Mitch (adapted and modified from [37]).
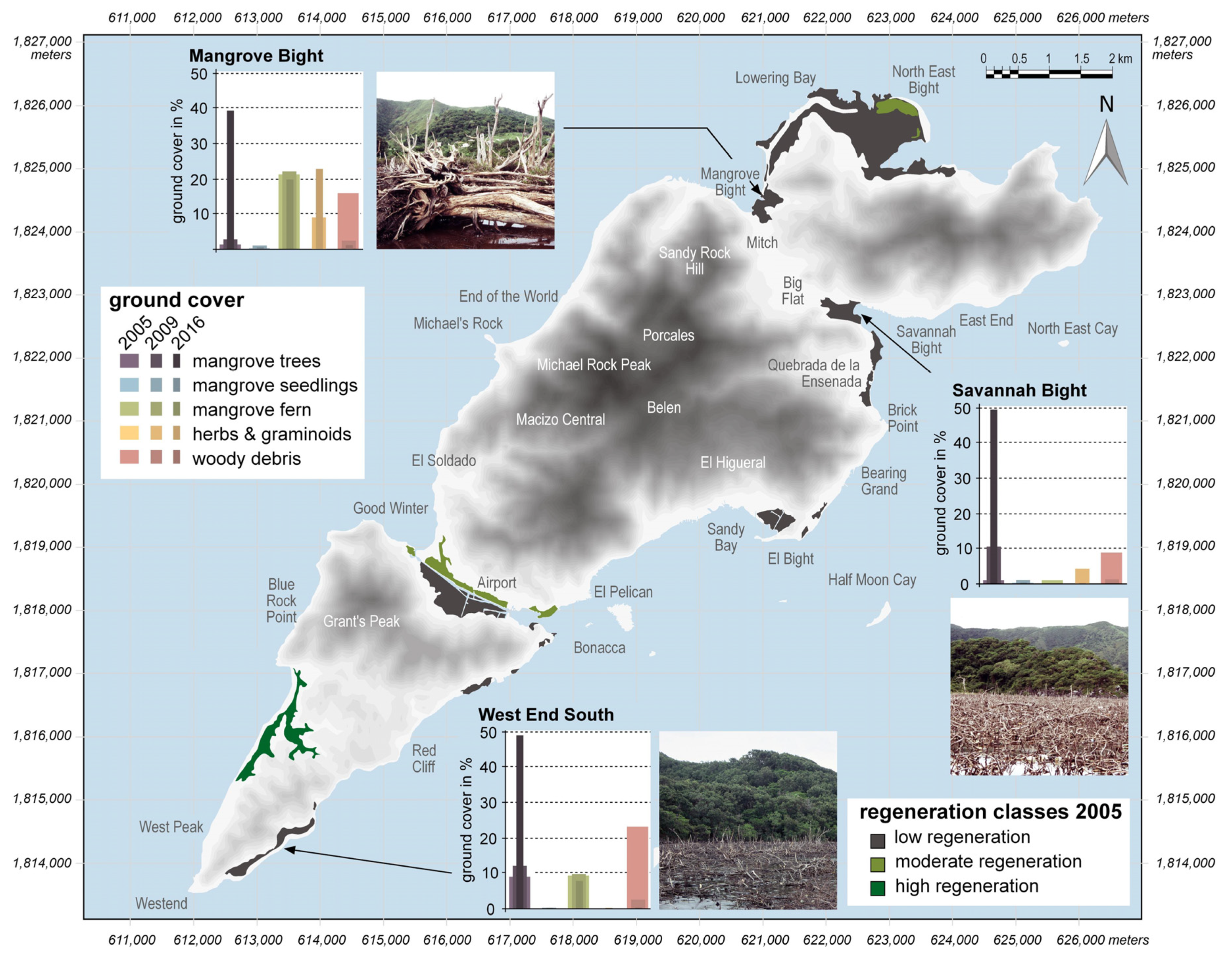
Figure 3.
Boxplots of the soil parameters of temperature, pH value, salinity, and redox potential at MB, SB, and WES.
Figure 3.
Boxplots of the soil parameters of temperature, pH value, salinity, and redox potential at MB, SB, and WES.
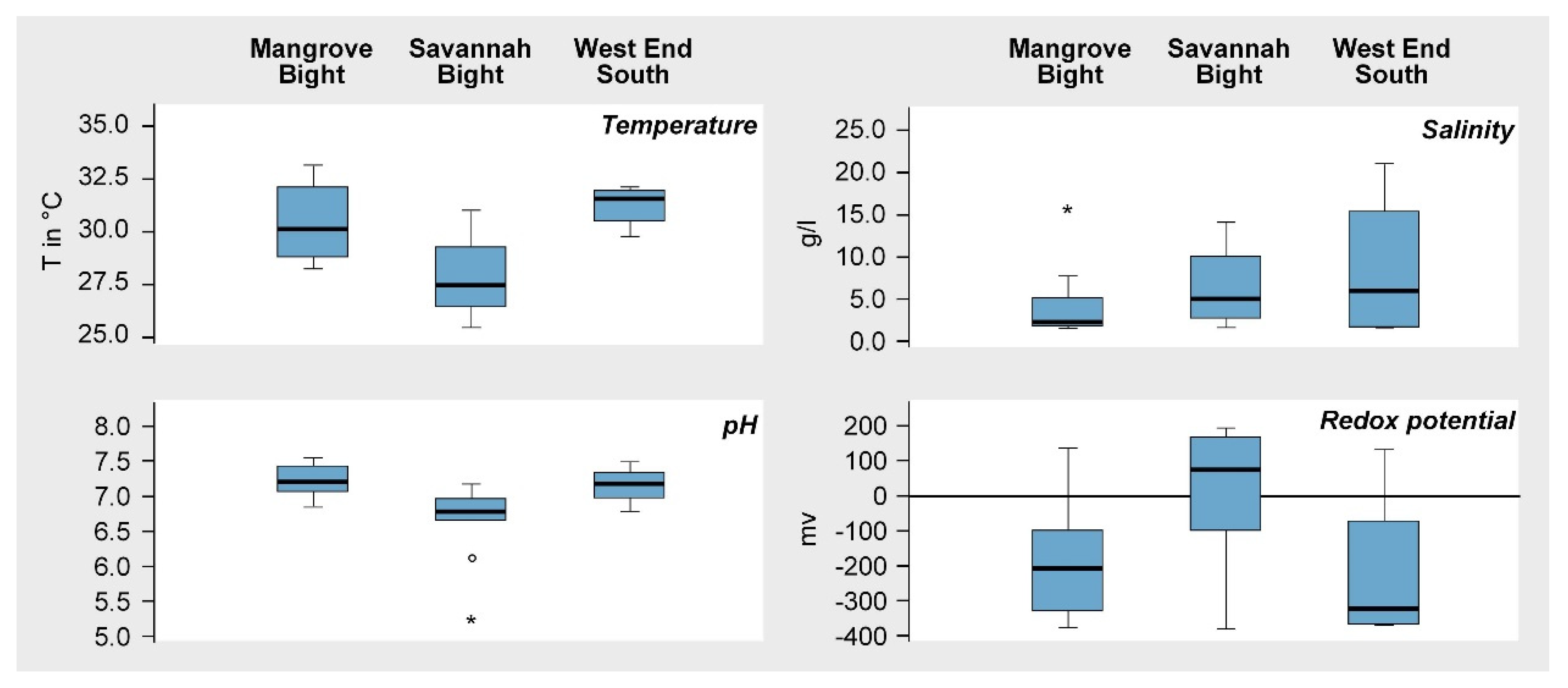
Figure 4.
Vegetation development documented along the line transects at MB, SB, and WES for the three different sampling dates in 2005, 2009, and 2016. Due to space limitations, only one transect (of the two) is displayed for MB and WES.
Figure 4.
Vegetation development documented along the line transects at MB, SB, and WES for the three different sampling dates in 2005, 2009, and 2016. Due to space limitations, only one transect (of the two) is displayed for MB and WES.
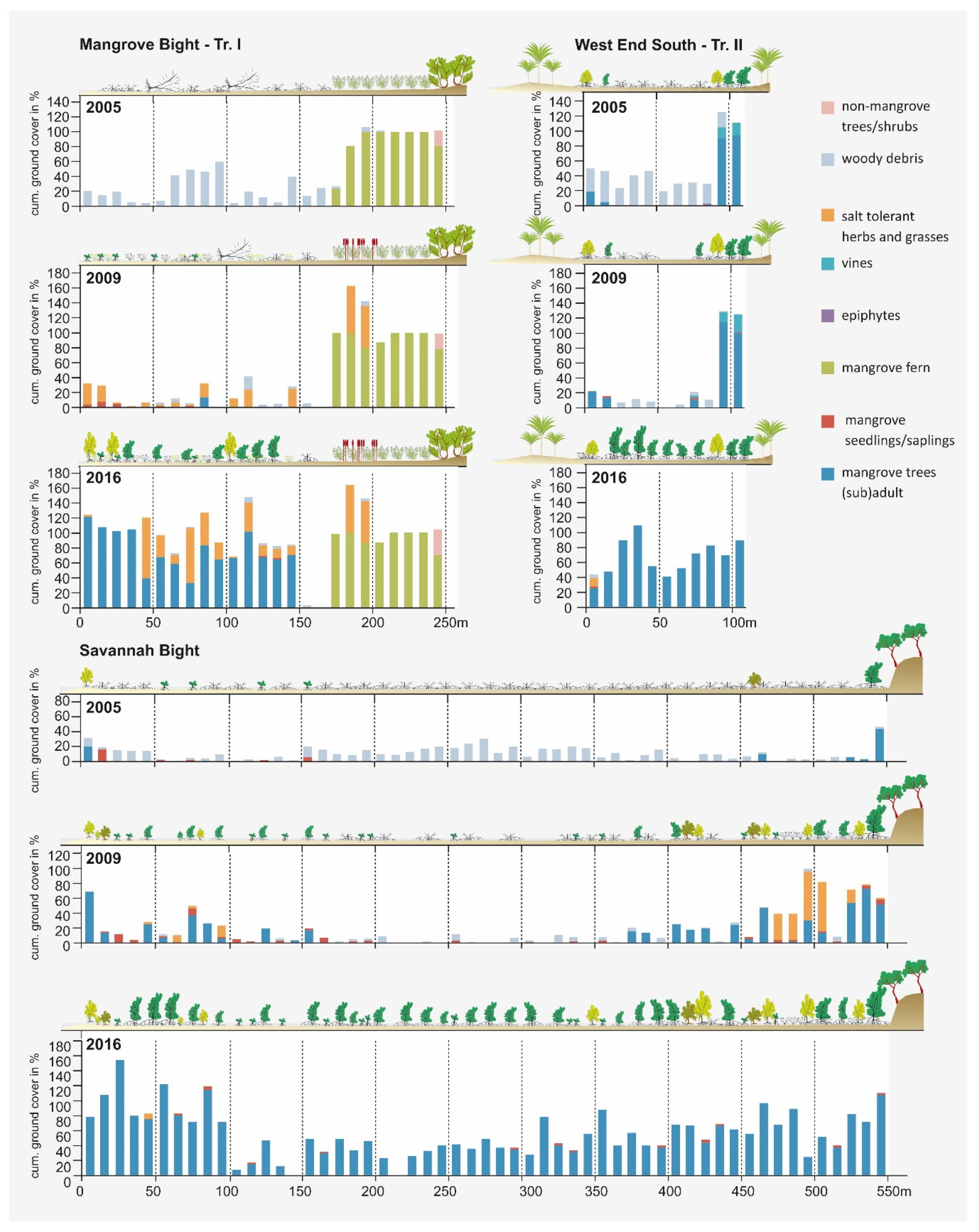
Figure 5.
Mean seedling numbers per 100 m of the three mangrove tree species present on Guanaja.

Figure 6.
PCA scatterplot encompassing all 10m-transect segments of all sampling dates (2005, 2009, 2016) for MB, SB, and WES; for sake of clarity, data are presented separately for the different sampling dates (adapted and modified from [37]).
Figure 6.
PCA scatterplot encompassing all 10m-transect segments of all sampling dates (2005, 2009, 2016) for MB, SB, and WES; for sake of clarity, data are presented separately for the different sampling dates (adapted and modified from [37]).

© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Fickert, T. To Plant or Not to Plant, That Is the Question: Reforestation vs. Natural Regeneration of Hurricane-Disturbed Mangrove Forests in Guanaja (Honduras). Forests 2020, 11, 1068. https://doi.org/10.3390/f11101068
AMA Style
Fickert T. To Plant or Not to Plant, That Is the Question: Reforestation vs. Natural Regeneration of Hurricane-Disturbed Mangrove Forests in Guanaja (Honduras). Forests. 2020; 11(10):1068. https://doi.org/10.3390/f11101068
Chicago/Turabian StyleFickert, Thomas. 2020. "To Plant or Not to Plant, That Is the Question: Reforestation vs. Natural Regeneration of Hurricane-Disturbed Mangrove Forests in Guanaja (Honduras)" Forests 11, no. 10: 1068. https://doi.org/10.3390/f11101068
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.