Effects of Cutting, Pruning, and Grafting on the Expression of Age-Related Genes in Larix kaempferi


1
State Key Laboratory of Tree Genetics and Breeding, Key Laboratory of Tree Breeding and Cultivation, National Forestry and Grassland Administration, Research Institute of Forest Ecology, Environment and Protection, Chinese Academy of Forestry, Beijing 100091, China
2
State Key Laboratory of Tree Genetics and Breeding, Key Laboratory of Tree Breeding and Cultivation, National Forestry and Grassland Administration, Research Institute of Forestry, Chinese Academy of Forestry, Beijing 100091, China
*
Authors to whom correspondence should be addressed.
Forests 2020, 11(2), 218; https://doi.org/10.3390/f11020218
Submission received: 4 December 2019
/
Revised: 1 January 2020
/
Accepted: 8 February 2020
/
Published: 14 February 2020
(This article belongs to the Section Forest Ecophysiology and Biology)
Abstract
:Grafting, cutting, and pruning are important horticultural techniques widely used in the establishment of clonal forestry. After the application of these techniques, some properties of the plants change, however, the underlying molecular mechanisms are still unclear. In our previous study, 27 age-related transcripts were found to be expressed differentially between the juvenile vegetative (1- and 2-year-old) and adult reproductive (25- and 50-year-old) phases of Larix kaempferi. Here, we re-analyzed the 27 age-related transcripts, cloned their full-length cDNA sequences, and measured their responses to grafting, cutting, and pruning. After sequence analysis and cloning, 20 transcription factors were obtained and annotated, most of which were associated with reproductive development, and six (LaAGL2-1, LaAGL2-2, LaAGL2-3, LaSOC1-1, LaAGL11, and LaAP2-2) showed regular expression patterns with L. kaempferi aging. Based on the expression patterns of these transcription factors in L. kaempferi trees subjected to grafting, cutting, and pruning, we concluded that (1) cutting and pruning rejuvenate the plants and change their expression, and the effects of cutting on gene expression are detectable within 14 years, although the cutting seedlings are still maturing during these years; (2) within three months after grafting, the rootstock is more sensitive to grafting than the scion and readily becomes mature with the effect of the scion, while the scion is not readily rejuvenated by the effect of the rootstock; and (3) LaAGL2-2 and LaAGL2-3 are more sensitive to grafting, while LaAP2-2 is impervious to it. These findings not only provide potential molecular markers to assess the state of plants but also aid in studies of the molecular mechanisms of rejuvenation.
1. Introduction
Vegetative propagation plays an important role in establishing clonal forestry, in which grafting and cutting are simple, rapid, and common methods, while pruning is a useful means of promoting the production of elite cutting wood and scions. However, with tree aging and maturing a great decline occurs in the rooting ability of cuttings [1,2]; this is the main limiting factor for the production of cutting seedlings and establishing clonal forestry. Therefore, the reversal of maturity or the maintaining of the juvenile stage are of great practical importance. In addition, grafting, cutting, and pruning are also useful techniques to rejuvenate plants [3,4]. Continuous grafting of mature scions onto juvenile rootstock can be used as a tool to rejuvenate the scions and improve their rooting ability, and this has been applied in many forest trees [4], such as Quercus acutissima [5]. After application of these horticultural techniques, plants change morphologically and physiologically, and show some juvenile characteristics, such as a stronger rooting ability [6,7]; however, the physical and molecular mechanisms underlying rejuvenation are still unclear.
Many genes have been found to function in plant aging, some of which promote maturation, such as Suppressor of Overexpression of Constans 1 (SOC1) [8], APETALA1 (AP1) [9], Squamosa-Promoter Binding Protein-Like (SPL) [10], and DAL1 (a MADS-box gene) [11], and some delay maturation or maintain the juvenile state, such as microRNA miR156 [12], miR171 [13], Tempranillo (TEM) [14], Terminal Flower 1 (TFL1) [15], MdTFL1 and MdTFL1a [16], Juvenile-to-Adult transition (JAT) [17], LaAP2L1 (a heterosis-associated AP2/EREBP transcription factor from Larix) [18], and Picea abies APETALA2-like genes [19]. In our previous study, 27 age-related transcripts were found to be expressed differentially in stems between the juvenile vegetative (1- and 2-year-old) and adult reproductive (25- and 50-year-old) phases of L. kaempferi, and some of them are homologous to the genes noted above (Figure 1), indicating that they might have similar functions in aging; meanwhile, they may be useful as molecular markers to determine the state of plants [20]. Together, these findings not only aid studies of the molecular mechanisms of aging and rejuvenation, but also provide candidate genes as molecular markers to assess the state of plants.
In fact, some have already been used as molecular markers. For example, miR156 is a potential marker of juvenility, and AP1 and SOC1 may be markers of the reproductive phase transition in avocado, mango, and macadamia plants [21]. However, the markers previously reported have generally been used to identify the state of naturally growing plants, and there has been limited use for identifying the state of plants treated with various horticultural techniques. Recently, grafting effects on gene expression at the transcriptome level have been reported in walnut [22] and tea [23] plants, providing more candidate molecular marker genes.
Here, we analyzed 27 age-related transcription factors again, cloned their full-length cDNA sequences, and measured their responses to grafting, cutting, and pruning to (1) determine if they can be used as molecular markers to determine the state of L. kaempferi trees treated with these horticultural techniques, and (2) study the molecular mechanisms of rejuvenation (Figure 1).
2. Materials and Methods
2.1. Plant Materials
The lateral branches produced in the current year from the tops of L. kaempferi trees were collected in early July of 2018 and 2019. The trees were located in Dagujia seed orchard (42°22′ N, 124°51′ E), Liaoning Province in Northeast China, and were produced from seeds, grafted seedlings, and cutting seedlings (Table 1). Rootstocks and scions from the grafted seedlings were sampled separately (Table 1). Pruning materials were sampled from four special cutting seedlings, which were pruned (hedged or sheared) and set at 1.5 m high (Table 1). After the removal of branches and needles, the stems from at least three of each kind of tree or each age category were pooled, frozen in liquid nitrogen, and stored at −80 °C until RNA extraction.
2.2. RNA Extraction and cDNA Synthesis
Total RNA was extracted with the EasyPure RNA Kit (TransGen Biotech, Beijing, China) according to the manufacturer’s protocol. A 2.5 μg aliquot of total RNA was reverse-transcribed into cDNA with the TransScript II One-step gDNA Removal and cDNA Synthesis SuperMix Kit (TransGen Biotech, Beijing, China), and subsequently diluted for gene isolation and expression analysis.
2.3. Sequence Analysis, Full-Length cDNA Cloning, and Annotation
The open reading frame (ORF) finder (https://www.ncbi.nlm.nih.gov/orffinder/) was used to identify the ORFs of the 27 transcription factors reported in our previous work [20] (Table 2). Based on the results, primers (Table S1) were designed to clone their full-length cDNA sequences (https://www.ncbi.nlm.nih.gov/tools/primer-blast/) with Platinum® Taq DNA polymerase (Invitrogen, Carlsbad, CA, USA). The PCR products were purified with a gel extraction kit (Tiangen, Beijing, China), ligated into the pEASY®-T1 simple cloning vector (TransGen Biotech, Beijing, China), and sequenced. The full-length cDNA sequences were submitted to GenBank (Table 2). Based on the results of blast analysis with Arabidopsis thaliana homologs, we assigned the Gene Ontology (GO) terms associated with the A. thaliana homologs to larch transcription factors.
2.4. Quantitative Reverse Transcription Polymerase Chain Reaction
The quantitative reverse transcription polymerase chain reaction (qRT-PCR) was used to assay the expression patterns of the identified transcription factors in different samples, with Larix kaempferi fructose-1,6-bisphosphatase 1 (LaFBP1, GenBank accession no. MN790763) (Table S2), which is expressed stably during tree aging [24], as the internal control. The qRT-PCR analysis was performed on a Bio-Rad CFX96 PCR system, using a TB Green® Premix Ex Taq™ (Tli RNase H Plus) (Takara, Shiga, Japan). Each reaction was carried out on 2 μL of diluted cDNA sample, in a total reaction system of 25 μL. The reaction procedure was set up according to the manufacturer’s protocol: 95 °C for 30 s, then 40 cycles at 95 °C for 5 s, and at 60 °C for 30 s, followed by a melting step from 65 to 95 °C. The relative expression ratio was expressed using the 2-ΔΔCt method. The expression level of each gene was standardized to the constitutive expression level of LaFBP1. The ratio between the expression levels of each age-related gene and LaFBP1 for each sample was calculated using the relative quantitative analysis method. The sample with the minimum expression level was used as a calibrator and was set to a value of 1. The qRT-PCR was performed with three technical replicates, and the data are shown as the mean ± SD.
3. Results and Discussion
3.1. Cloning and Annotation of 20 Transcription Factors
After analyzing the sequences of 27 transcription factors, we found that some had the same ORF (Table 2); in total, 21 non-redundant sequences were obtained, while 20 were cloned successfully and used for further study (Table 2). Blast analysis of the 20 cloned sequences was performed again. The results showed that the 20 transcription factors were from eight families and had 14 homologs in A. thaliana (Table 2). Among the eight families, the MADS-box family had the most members (Table 2).
Based on the names and GO annotation of their homologs in A. thaliana, they were designated and annotated (Table 2). In total, 36 GO terms in the biological process category were obtained, among which 18 were associated with processes of plant reproductive development and their hormonal and environmental control (Figure 2), such as “flower development”, “meristem maintenance”, “response to gibberellin”, and “vernalization response”, and therefore provided more information about the roles of these 20 transcription factors in the sexual reproductive development of L. kaempferi. However, further experiments were required to assess whether they showed regular expression patterns with aging and if they could be used as molecular markers to assess the state of plants.
3.2. Expression Patterns of 20 Transcription Factors during Larch Tree Aging
The qRT-PCR assays were used to reveal the expression patterns of the 20 transcription factors in 1-, 3-, 5-, 7-, 9-, 11-, and 13-year-old trees. We found that the expression level of LaAP2-2 decreased with age (Figure 3o), while that of LaAP2-1 decreased, and that of LaOZF2 increased from 1 to 5 years (Figure 3n,q) and the expression levels of LaAGL2-1, LaAGL2-2, LaAGL2-3, LaSOC1-1, LaSOC1-3, LaAGL11, and LaAGL42 increased from 1 to 7 years, while almost no changes were found in the expression levels of LaAGL2-1, LaAGL2-2, LaAGL2-3, LaSOC1-1, LaAGL11, and LaAGL42 from 7 to 13 years and a decrease occurred in the expression level of LaSOC1-3 (Figure 3).
The maturity status of a plant is dynamic and relative and can be reflected by many aspects of tree growth and development, including the changes in gene expression, morphology, and physiology. For example, in larches the capacity to produce seeds occurs at about 10-years-old, the effect of donor tree age on rooting ability is obvious, and at about 12-years-old rooting ability decreased greatly [25,26]. Based on the regular expression patterns of LaAGL2-1, LaAGL2-2, LaAGL2-3, LaSOC1-1, LaAGL11, and LaAP2-2 over the course of aging up to 13-years-old, we concluded that their expression is age-dependent. Given that the maturity status of a plant changes after the application of horticulture techniques, we speculated that the expression of these six age-related genes might reflect this change. Next, we measured the expression patterns in trees subjected to grafting, cutting, and pruning to determine the effects of these techniques on the expression of these genes.
Here, the changes in gene expression were emphasized and we tried to correlate the changes in gene expression with those in physiological status, especially flowering and rooting ability. DAL1, a MADS-box transcription factor from P. abies, is expressed increasingly from the age of 5-years-old, and its over-expression in Arabidopsis results in early flowering [11]; here the putative homolog of DAL1, LaAGL2, was identified and showed almost the same expression pattern as DAL1 (Figure 3), indicating its involvement in the transition from the juvenile vegetative to the adult reproductive phase. While the capacity to produce seeds occurs at about 10-years-old in Larix, the molecular regulatory mechanism for the vegetative to reproductive development in trees mediated by LaAGL2 is still unclear. Regarding rooting ability, it has been detected that the rooting rate of larch cutting decreased sharply at about 12-year-old [25]; further work is needed to understand how the age-related genes function in this regenerative process.
3.3. Effects of Pruning and Cutting on the Expression of Six Age-Related Genes
In 14-year-old seed seedlings, we found higher expression levels of LaAGL2-2, LaAGL2-3, LaSOC1-1, and LaAGL11 and lower levels of LaAP2-2 (Figure 4). Compared with 14-year-old seed seedlings, cutting seedlings showed a decrease in the expression levels of LaAGL2-2, LaAGL2-3, LaSOC1-1, and LaAGL11 and an increase in the level of LaAP2-2, while almost no change was found in the level of LaAGL2-1 (Figure 4). After pruning the cutting seedlings, clear changes were found in the expression levels where LaAGL2-1, LaAGL2-2, LaAGL2-3, LaSOC1-1, and LaAGL11 transcript levels decreased and LaAP2-2 transcript levels increased (Figure 4). These results suggested that the expression of these six age-related genes is affected by cutting or pruning; based on their expression patterns, we concluded that cutting and pruning rejuvenates plants, and this is indicated by the increased rooting ability of cuttings [27,28,29,30,31].
Notably, the expression levels of LaAGL2-1, LaAGL2-2, LaAGL2-3, LaSOC1-1, and LaAGL11 in the pruning materials were lower than those in cutting seedlings, and the expression levels of LaAP2-2 were higher than those in cutting seedlings (Figure 4). These results suggested that pruning further affects the expression of these six transcription factors in plants originating from cuttings. Together, we concluded that cutting rejuvenates plants, pruning maintains their juvenility [32], and the effect of cutting on gene expression can be detected 14 years later.
To further study the effect of cutting on the expression of the six age-related genes in the subsequent early years after cutting, we assessed their expression patterns in 1- and 2-year-old cutting seedlings. Compared with those in 13- and 14-year-old seed seedlings, from which the cuttings were produced in 2018 and 2017, respectively, the expression levels of LaAGL2-2, LaAGL2-3, LaSOC1-1, and LaAGL11 decreased in the 1- and 2-year-old cutting seedlings, while the expression level of LaAP2-2 increased. Almost no change in the expression level of LaAGL2-1 was found in 1-year-old cutting seedlings and 13-year-old seed seedlings, while it was lower in the 2-year-old cutting seedlings than that in 14-year-old seed seedlings (Figure 5). These results showed that cutting changes the expression of these genes immediately after the treatment and rejuvenates the plant within two years after cutting.
Moreover, we found that the expression levels of LaAGL2-1, LaAGL2-2, LaAGL2-3, LaSOC1-1, and LaAGL11 in 2-year-old cutting seedlings were lower than those in 1-year-old cutting seedlings, and the opposite pattern occurred in LaAP2-2 expression (Figure 5 and Figure S1). These results indicated that cutting affects the expression of the six age-related genes and these effects are enhanced as the cutting seedling ages.
It is known that as cutting seedlings age, the phenotypes of maturation also appear, so the question arose as to when maturation begins and when the enhancement of gene expression by the cutting effect ends. We set out to answer these questions by assessing the expression patterns of the six age-related genes in 1-, 3-, 5-, 7-, 9-, 11- and 13-year-old cutting seedlings. The lowest levels of LaAGL2-1, LaAGL2-2, and LaAGL2-3 transcripts were found in 3-year-old cutting seedlings after which the levels of these transcripts increased; the expression levels of LaAGL2-1 at 11-years-old, LaAGL2-2 and LaAGL2-3 at 9-years-old were almost the same as those at 1-year-old (Figure 6). These data indicated that, in response to cutting, the transcription of LaAGL2-1, LaAGL2-2, and LaAGL2-3 in cutting seedlings is suppressed at first, and with aging it is induced again (Figure 6). Based on the expression patterns of these transcripts, we concluded that the enhancement of gene expression by the cutting effect persists for only 2–3 years and maturation begins early. The irregular expression patterns of LaSOC1-1, LaAGL11, and LaAP2-2 during the aging of cutting seedlings cannot yet be explained (Figure 6), but these data suggested that the effects of cutting on the expression of age-related genes are complex and regulated by currently unknown mechanisms.
Based on the expression patterns of these age-related genes, we concluded that cutting rejuvenates the plant and affects gene expression; this effect is detected within 14 years, although the cutting seedlings are still maturing during these years.
3.4. Expression Patterns of the Six Age-Related Genes after Grafting
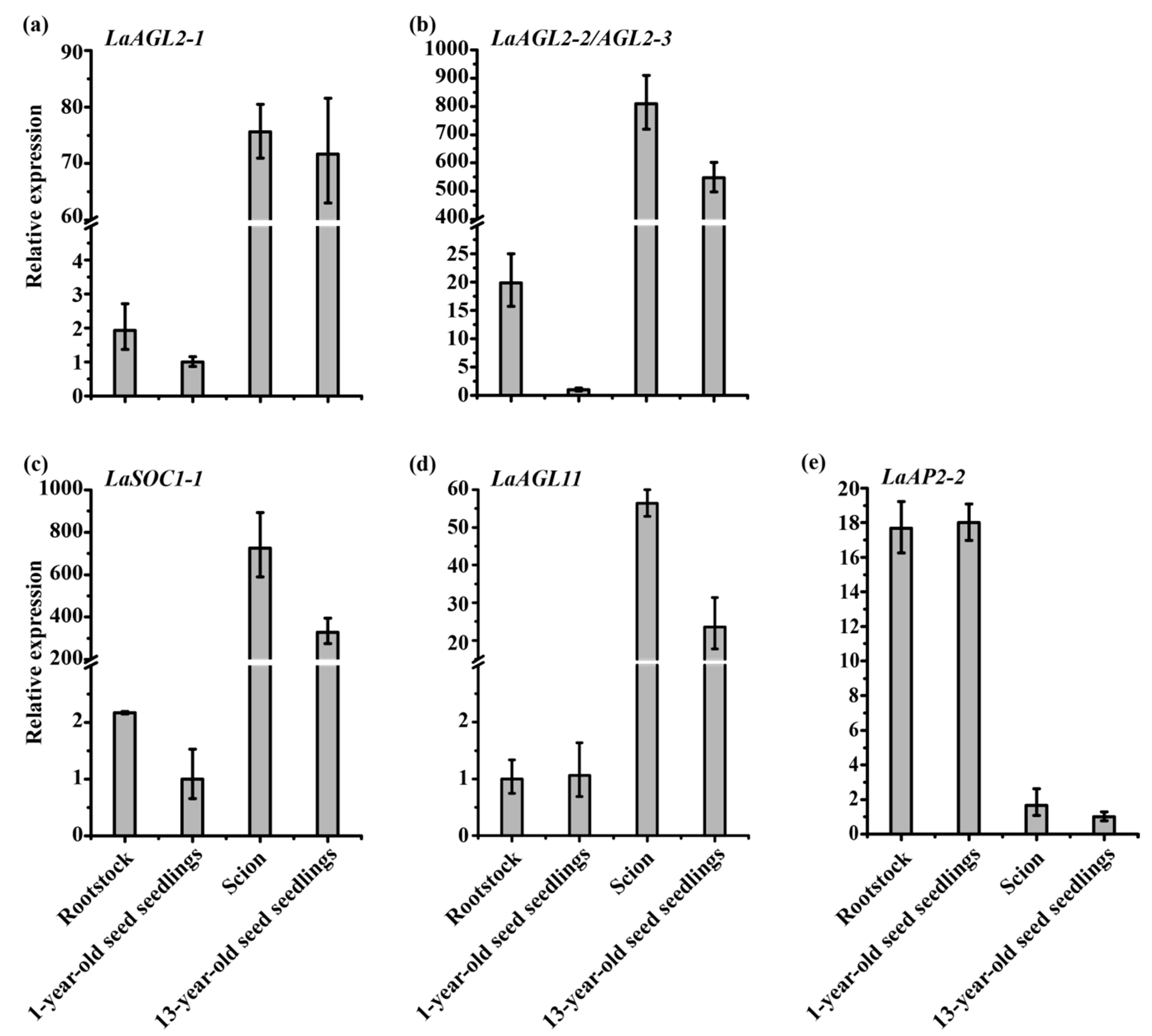
Grafting is also a useful technique that can promote the rejuvenation of many plant species, such as Sequoia sempervirens and the European larch [33,34,35]. Here, we also assessed the effect of grafting on the expression of the six age-related genes in scions and rootstocks. Compared with those in rootstocks, higher expression levels of LaAGL2-1, LaAGL2-2, LaAGL2-3, LaSOC1-1, and LaAGL11, and a lower level of LaAP2-2 were found in scions three months after grafting (Figure 7). Compared with those in 1-year-old seed seedlings, higher levels of LaAGL2-1, LaAGL2-2, LaAGL2-3, and LaSOC1-1 transcripts were found in the rootstocks, while almost no change was found in the levels of LaAGL11 and LaAP2-2 (Figure 7), showing that in rootstock, grafting increases the expression of LaAGL2-1, LaAGL2-2, LaAGL2-3, and LaSOC1-1, but has no effect on the expression of LaAGL11 and LaAP2-2. Compared with those in 13-year-old seed seedlings, higher expression levels of LaAGL2-2, LaAGL2-3, LaSOC1-1, and LaAGL11 were found in the scions, while almost no change was found in the levels of LaAGL2-1 and LaAP2-2 (Figure 7), showing that in scions, grafting increases the expression of LaAGL2-2, LaAGL2-3, LaSOC1-1, and LaAGL-11, but does not affect the expression of LaAGL2-1 and LaAP2-2. Together, these results showed that grafting affects the expression of age-related genes in the rootstock and scion and each gene has a different response to grafting.
Based on the expression patterns of these age-related genes, we concluded that (1) within three months after grafting, the rootstock is more sensitive to grafting than the scion—the rootstock readily matures with the effect of the scion, while the scion is not readily rejuvenated by the effect of the rootstock, because the expression of LaAGL2-1, LaAGL2-2, LaAGL2-3, and LaSOC1-1 increased in the rootstock and did not decrease in the scion; (2) LaAGL2-2 and LaAGL2-3 are more sensitive to grafting than the other four genes because greater changes in their expression occurred after grafting; and (3) LaAP2-2 is impervious to grafting, because almost no changes in its expression were detected in rootstock and scion.
4. Conclusions
Taken together, we conclude that grafting, cutting, and pruning change the expression of age-related genes and rejuvenate plants through currently unknown mechanisms. The identification of age-related genes helps to reveal the mechanisms of rejuvenation and the understanding of other physiological changes that occur with age, which promotes the establishment of clonal forestry.
Supplementary Materials
The following are available online at https://www.mdpi.com/1999-4907/11/2/218/s1, Figure S1: Expression patterns of LaAGL2-1 (a), LaAGL2-2 (b), LaAGL2-3 (b), LaSOC1-1 (c), LaAGL11 (d), and LaAP2-2 (e) in 1- and 2-year-old Larix kaempferi cutting seedlings (n = 3, sampled in 2018) assayed by qRT-PCR with LaFBP1 as the internal control. In 2017 and 2016 1- and 2-year-old cutting seedlings were propagated from 21- and 20-year-old seed seedlings that were sampled in 2018 when they were 22 years old, respectively. The qRT-PCR was performed with three technical replicates, and the data are shown as the mean ± SD. Table S1: Primers used for cloning 20 transcription factors, Table S2: Primers used for qRT-PCR.
Author Contributions
Y.Z. carried out the study, analyzed the data, and wrote the manuscript. Q.-L.Z. helped to analyze the data and revised the manuscript. W.-F.L. conceived and designed the study, analyzed the data and revised the manuscript. L.-W.Q. and S.-Y.H. provided suggestions on the experimental design and analyses. All authors have read and approved the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (31770714 and 31330017), the Basic Research Fund of Research Institute of Forestry (RIF2014-07), the National Transgenic Major Program (2018ZX08020-003), and the National Key R&D Program of China (2017YFD0601204-2).
Acknowledgments
The authors thank I.C. Bruce (Peking University) for critical reading of the manuscript, and the anonymous reviewers and editor for their constructive comments on an earlier version of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Díaz-Sala, C. Direct reprogramming of adult somatic cells toward adventitious root formation in forest tree species: The effect of the juvenile–adult transition. Front. Plant Sci. 2014, 5, 310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massoumi, M.; Krens, F.A.; Visser, R.G.F.; de Klerk, G.J.M. Azacytidine and miR156 promote rooting in adult but not in juvenile Arabidopsis tissues. J. Plant Physiol. 2017, 208, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Hand, P.; Besford, R.T.; Richardson, C.M.; Peppitt, S.D. Antibodies to phase related proteins in juvenile and mature Prunus avium. Plant Growth Regul. 1996, 20, 25–29. [Google Scholar] [CrossRef]
- Wendling, I.; Trueman, S.J.; Xavier, A. Maturation and related aspects in clonal forestry—part II: Reinvigoration, rejuvenation and juvenility maintenance. New For. 2014, 45, 473–486. [Google Scholar] [CrossRef]
- Moon, H.K.; Yi, J.S. Cutting propagation of Quercus acutissima clones after rejuvenation through serial grafting. Ann. Sci. For. 1993, 50, 314–318. [Google Scholar] [CrossRef]
- Wang, J.; Jiang, L.; Wu, R. Plant grafting: How genetic exchange promotes vascular reconnection. New Phytol. 2017, 214, 56–65. [Google Scholar] [CrossRef] [Green Version]
- Majada, J.; Martínez-Alonso, C.; Feito, I.; Kidelman, A.; Aranda, I.; Alía, R. Mini-cuttings: An effective technique for the propagation of Pinus pinaster Ait. New For. 2011, 41, 399–412. [Google Scholar] [CrossRef]
- Jung, J.H.; Ju, Y.; Seo, P.J.; Lee, J.H.; Park, C.M. The SOC1-SPL module integrates photoperiod and gibberellic acid signals to control flowering time in Arabidopsis. Plant J. 2012, 69, 577–588. [Google Scholar] [CrossRef]
- Murai, K.; Miyamae, M.; Kato, H.; Takumi, S.; Ogihara, Y. WAP1, a Wheat APETALA1 homolog, plays a central role in the phase transition from vegetative to reproductive growth. Plant Cell Physiol. 2003, 44, 1255–1265. [Google Scholar] [CrossRef]
- Preston, J.C.; Hileman, L.C. Functional evolution in the plant SQUAMOSA-PROMOTER BINDING PROTEIN-LIKE (SPL) gene family. Front. Plant Sci. 2013, 4, 80. [Google Scholar] [CrossRef] [Green Version]
- Carlsbecker, A.; Tandre, K.; Johanson, U.; Englund, M.; Engström, P. The MADS-box gene DAL1 is a potential mediator of the juvenile-to-adult transition in Norway spruce (Picea abies). Plant J. 2004, 40, 546–557. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.W.; Park, M.Y.; Wang, L.J.; Koo, Y.; Chen, X.Y.; Weigel, D.; Poethig, R.S. MiRNA control of vegetative phase change in trees. PLoS Genet. 2011, 7, e1002012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curaba, J.; Talbot, M.; Li, Z.; Helliwell, C. Over-expression of microRNA171 affects phase transitions and floral meristem determinancy in barley. BMC Plant Biol. 2013, 13, 6. [Google Scholar] [CrossRef] [Green Version]
- Sgamma, T.; Jackson, A.; Muleo, R.; Thomas, B.; Massiah, A. TEMPRANILLO is a regulator of juvenility in plants. Sci. Rep. 2014, 4, 3704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamed, R.; Wang, C.T.; Ma, C.; Shevchenko, O.; Dye, S.J.; Puzey, J.R.; Etherington, E.; Sheng, X.; Meilan, R.; Strauss, S.H.; et al. Populus CEN/TFL1 regulates first onset of flowering, axillary meristem identity and dormancy release in Populus. Plant J. 2010, 62, 674–688. [Google Scholar] [CrossRef]
- Mimida, N.; Kotoda, N.; Ueda, T.; Igarashi, M.; Hatsuyama, Y.; Iwanami, H.; Moriya, S.; Abe, K. Four TFL1/CEN -Like genes on distinct linkage groups show different expression patterns to regulate vegetative and reproductive development in Apple (Malus × domestica Borkh.). Plant Cell Physiol. 2009, 50, 394–412. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Ocaña, A.; García-López, M.C.; Jiménez-Ruiz, J.; Saniger, L.; Macías, D.; Navarro, F.; Oya, R.; Belaj, A.; Rosa, R.; Corpas, F.J.; et al. Identification of a gene involved in the juvenile-to-adult transition (JAT) in cultivated olive trees. Tree Genet. Genomes 2010, 6, 891–903. [Google Scholar] [CrossRef]
- Li, A.; Zhou, Y.; Jin, C.; Song, W.; Chen, C.; Wang, C. LaAP2L1, a heterosis-associated AP2/EREBP transcription factor of Larix, increases organ size and final biomass by affecting cell proliferation in Arabidopsis. Plant Cell Physiol. 2013, 54, 1822–1836. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, L.; Carlsbecker, A.; Sundås-Larsson, A.; Vahala, T. APETALA2 like genes from Picea abies show functional similarities to their Arabidopsis homologues. Planta 2007, 225, 589–602. [Google Scholar] [CrossRef]
- Xiang, W.B.; Li, W.F.; Zhang, S.G.; Qi, L.W. Transcriptome-wide analysis to dissect the transcription factors orchestrating the phase change from vegetative to reproductive development in Larix kaempferi. Tree Genet. Genomes 2019, 15, 68. [Google Scholar] [CrossRef] [Green Version]
- Ahsan, M.U.; Hayward, A.; Irihimovitch, V.; Fletcher, S.; Tanurdzic, M.; Pocock, A.; Beveridge, C.A.; Mitter, N. Juvenility and vegetative phase transition in tropical/subtropical tree crops. Front. Plant Sci. 2019, 10, 729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Q.; Bu, D.; Zhang, J.; Wu, Y.; Pei, D. The transcriptome landscape of walnut interspecies hybrid (Juglans hindsii × Juglans regia) and regulation of cambial activity in relation to grafting. Front. Genet. 2019, 10, 577. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Lu, M.; Zhu, B.; Wang, R.; Wan, X.; Deng, W.W.; Zhang, Z.Z. Integrated transcriptomic and phytochemical analyses provide insights into characteristic metabolites variation in leaves of 1-year-old grafted tea (Camellia sinensis). Tree Genet. Genomes 2019, 15, 58. [Google Scholar] [CrossRef]
- Li, W.F.; Yang, W.H.; Zhang, S.G.; Han, S.Y.; Qi, L.W. Transcriptome analysis provides insights into wood formation during larch tree aging. Tree Genet. Genomes 2017, 13, 19. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.S.; Hu, X.S.; Wang, Y.C.; Nie, J.; Liu, S.M. The effect of age and basal-cut shapes of cutting on rooting of Larix leptolepis. For. Res. 1993, 6, 627–632. [Google Scholar]
- Wang, J.H.; Sun, X.M.; Wang, X.S.; Xu, C.L.; Ding, B.; Wang, X.D. Effects of age, type of auxin and treatment concentration on rooting ability of Larix leptolepis. For. Res. 2006, 19, 102–108. [Google Scholar]
- Trueman, S.J. Clonal propagation and storage of subtropical pines in Queensland, Australia. S. Afr. For. J. 2006, 208, 49–52. [Google Scholar] [CrossRef]
- Rosier, C.L.; Frampton, J.; Goldfarb, B.; Wise, F.C.; Blazich, F.A. Stumping height, crown position, and age of parent tree influence rooting of stem cuttings of Fraser fir. HortScience 2005, 40, 771–777. [Google Scholar] [CrossRef] [Green Version]
- Aimers-Halliday, J.; Menzies, M.I.; Faulds, T.; Holden, D.G.; Low, C.B.; Dibley, M.J. Nursery systems to control maturation in Pinus radiata cuttings, comparing hedging and serial propagation. N. Z. J. For. Sci. 2003, 33, 135–155. [Google Scholar]
- Peer, K.R.; Greenwood, M.S. Maturation, topophysis and other factors in relation to rooting in Larix. Tree Physiol. 2001, 21, 267–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wendling, I.; Warburton, P.M.; Trueman, S.J. Maturation in Corymbia torelliana × C. citriodora stock plants: Effects of pruning height on shoot production, adventitious rooting capacity, stem anatomy, and auxin and abscisic acid concentrations. Forests 2015, 6, 3763–3778. [Google Scholar] [CrossRef] [Green Version]
- Cameron, R.; Harrison-Murray, R.; Fordham, M.; Judd, H.; Ford, Y.; Marks, T.; Edmondson, R. Rooting cuttings of syringa vulgaris cv. charles joly and corylus avellana cv. aurea: The influence of stock plant pruning and shoot growth. Trees 2003, 17, 451–462. [Google Scholar] [CrossRef]
- Arnaud, Y.; Franclet, A.; Tranvan, H.; Jacques, M. Micropropagation and rejuvenation of Sequoia sempervirens (Lamb) Endl: A review. Ann. Sci. For. 1993, 50, 273–295. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.C.; Chow, T.Y.; Tseng, T.C.; Kuo, C.I.; Liu, S.M.; Ngoh, M.G.; Murashige, T.; Huang, H.J. Association of mitochondrial plasmids with rejuvenation of the coastal redwood, Sequoia sempervirens (D. Don) Endl. Bot. Bull. Acad. Sin. 2003, 44, 25–30. [Google Scholar]
- Ewald, D.; Kretzschmar, U. The influence of micrografting in vitro on tissue culture behavior and vegetative propagation of old European larch trees. Plant Cell Tissue Organ Cult. 1996, 44, 249–252. [Google Scholar]
Figure 1.
Analytical flowchart. Twenty-seven transcripts differentially expressed between the juvenile vegetative (1- and 2-year-old) and adult reproductive (25- and 50-year-old) phases of Larix kaempferi, identified in a previous study [20] were analyzed again and used to identify molecular markers and assess the effects of cutting, grafting, and pruning on gene expression.
Figure 1.
Analytical flowchart. Twenty-seven transcripts differentially expressed between the juvenile vegetative (1- and 2-year-old) and adult reproductive (25- and 50-year-old) phases of Larix kaempferi, identified in a previous study [20] were analyzed again and used to identify molecular markers and assess the effects of cutting, grafting, and pruning on gene expression.

Figure 2.
Biological processes in Gene Ontology (GO) terms. Based on the results of blast analysis, the GO terms associated with the Arabidopsis thaliana homologs were assigned to the Larix kaempferi transcription factors.
Figure 2.
Biological processes in Gene Ontology (GO) terms. Based on the results of blast analysis, the GO terms associated with the Arabidopsis thaliana homologs were assigned to the Larix kaempferi transcription factors.

Figure 3.
Expression patterns of 20 transcription factors [LaCAL (a), LaAGL2-1 (b), LaAGL2-2 (c), LaAGL2-3 (c), LaSOC1-1 (d), LaSOC1-2 (e), LaSOC1-3 (f), LaSOC1-4 (g), LaAGL1 (h), LaAGL11 (i), LaAGL42 (j), LaERF017 (k), LaERF3 (l), LaSCL29 (m), LaAP2-1 (n), LaAP2-2 (o), LaHCA2 (p), LaOZF2 (q), LaTRL6 (r), LaPHL1 (s)] in 1-, 3-, 5-, 7-, 9-, 11-, and 13-year-old Larix kaempferi seed seedlings (n ≥ 6, sampled in 2019) assayed by qRT-PCR with LaFBP1 as the internal control.
Figure 3.
Expression patterns of 20 transcription factors [LaCAL (a), LaAGL2-1 (b), LaAGL2-2 (c), LaAGL2-3 (c), LaSOC1-1 (d), LaSOC1-2 (e), LaSOC1-3 (f), LaSOC1-4 (g), LaAGL1 (h), LaAGL11 (i), LaAGL42 (j), LaERF017 (k), LaERF3 (l), LaSCL29 (m), LaAP2-1 (n), LaAP2-2 (o), LaHCA2 (p), LaOZF2 (q), LaTRL6 (r), LaPHL1 (s)] in 1-, 3-, 5-, 7-, 9-, 11-, and 13-year-old Larix kaempferi seed seedlings (n ≥ 6, sampled in 2019) assayed by qRT-PCR with LaFBP1 as the internal control.

Figure 4.
Expression patterns of LaAGL2-1 (a), LaAGL2-2 (b), LaAGL2-3 (b), LaSOC1-1 (c), LaAGL11 (d), and LaAP2-2 (e) in 14-year-old Larix kaempferi pruning materials (n = 3, sampled in 2018), cutting seedlings (n = 3, sampled in 2018), and seed seedlings (n = 3, sampled in 2018) assayed by qRT-PCR with LaFBP1 as the internal control.
Figure 4.
Expression patterns of LaAGL2-1 (a), LaAGL2-2 (b), LaAGL2-3 (b), LaSOC1-1 (c), LaAGL11 (d), and LaAP2-2 (e) in 14-year-old Larix kaempferi pruning materials (n = 3, sampled in 2018), cutting seedlings (n = 3, sampled in 2018), and seed seedlings (n = 3, sampled in 2018) assayed by qRT-PCR with LaFBP1 as the internal control.

Figure 5.
Expression patterns of LaAGL2-1 (a), LaAGL2-2 (b), LaAGL2-3 (b), LaSOC1-1 (c), LaAGL11 (d), and LaAP2-2 (e) in 1- and 2-year-old Larix kaempferi cutting seedlings (n ≥ 12, sampled in 2019) and 13- and 14-year-old seed seedlings (n ≥ 6, sampled in 2019) assayed by qRT-PCR with LaFBP1 as the internal control. In 2018 and 2017 1- and 2-year-old cutting seedlings were propagated from 12-year-old seed seedlings that were sampled in 2019 when they were 13- and 14- years old, respectively.
Figure 5.
Expression patterns of LaAGL2-1 (a), LaAGL2-2 (b), LaAGL2-3 (b), LaSOC1-1 (c), LaAGL11 (d), and LaAP2-2 (e) in 1- and 2-year-old Larix kaempferi cutting seedlings (n ≥ 12, sampled in 2019) and 13- and 14-year-old seed seedlings (n ≥ 6, sampled in 2019) assayed by qRT-PCR with LaFBP1 as the internal control. In 2018 and 2017 1- and 2-year-old cutting seedlings were propagated from 12-year-old seed seedlings that were sampled in 2019 when they were 13- and 14- years old, respectively.

Figure 6.
Expression patterns of LaAGL2-1 (a), LaAGL2-2 (b), LaAGL2-3 (b), LaSOC1-1 (c), LaAGL11 (d), and LaAP2-2 (e) in 1-, 3-, 5-, 7-, 9-, 11-, and 13-year-old Larix kaempferi cutting seedlings (n ≥ 5, sampled in 2019) assayed by qRT-PCR with LaFBP1 as the internal control.
Figure 6.
Expression patterns of LaAGL2-1 (a), LaAGL2-2 (b), LaAGL2-3 (b), LaSOC1-1 (c), LaAGL11 (d), and LaAP2-2 (e) in 1-, 3-, 5-, 7-, 9-, 11-, and 13-year-old Larix kaempferi cutting seedlings (n ≥ 5, sampled in 2019) assayed by qRT-PCR with LaFBP1 as the internal control.

Figure 7.
Expression patterns of LaAGL2-1 (a), LaAGL2-2 (b), LaAGL2-3 (b), LaSOC1-1 (c), LaAGL11 (d), and LaAP2-2 (e) in 3-month-old grafted seedlings (n = 6, sampled in 2019) and 1- and 13-year-old seed seedlings (n = 11, sampled in 2019) assayed by qRT-PCR with LaFBP1 as the internal control.
Figure 7.
Expression patterns of LaAGL2-1 (a), LaAGL2-2 (b), LaAGL2-3 (b), LaSOC1-1 (c), LaAGL11 (d), and LaAP2-2 (e) in 3-month-old grafted seedlings (n = 6, sampled in 2019) and 1- and 13-year-old seed seedlings (n = 11, sampled in 2019) assayed by qRT-PCR with LaFBP1 as the internal control.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Information on samples.
| Samples | Age | Other Information |
|---|---|---|
| Seed Seedlings | 1 year | These seedlings were grown from seeds |
| 3 years | ||
| 5 years | ||
| 7 years | ||
| 9 years | ||
| 11 years | ||
| 13 years | ||
| 14 years | ||
| Grafted Seedlings | 3 months | Rootstocks were from 1-year-old and scions from 12-year-old seed seedlings |
| Cutting Seedlings | 1 year | Cuttings were sampled from 12-year-old seed seedlings |
| 2 years | ||
| 3 years | - | |
| 5 years | - | |
| 7 years | - | |
| 9 years | - | |
| 11 years | - | |
| 13 years | - | |
| 14 years | Cuttings were sampled from 8-year-old seed seedlings | |
| Pruning Materials | 14 years | Cutting seedlings were pruned; they were propagated from cuttings from 8-year-old seed seedlings |
- indicates no age information.
Table 2.
Analysis of 27 transcripts expressed differentially between juvenile vegetative (1- and 2-year-old) and adult reproductive (25- and 50-year-old) phases of Larix kaempferi [20].
Table 2.
Analysis of 27 transcripts expressed differentially between juvenile vegetative (1- and 2-year-old) and adult reproductive (25- and 50-year-old) phases of Larix kaempferi [20].
| Family | Name | Transcript ID | Accession Number |
|---|---|---|---|
| MADS-box | LaCAL | comp81209_c0_seq1 | MN790743 |
| LaAGL2-1 | comp125095_c0_seq5 | MN790744 | |
| comp126977_c0_seq13 | |||
| LaAGL2-2 | comp125095_c0_seq9 | MN790745 | |
| comp126977_c0_seq2 | |||
| comp126977_c0_seq5 | |||
| LaAGL2-3 | comp126977_c0_seq16 | MN790746 | |
| LaSOC1-1 | comp128412_c0_seq11 | MN790747 | |
| LaSOC1-2 | comp128412_c0_seq23 | MN790748 | |
| LaSOC1-3 | comp128412_c0_seq8 | MN790749 | |
| comp128471_c0_seq5 | |||
| comp128471_c0_seq24 | |||
| comp128471_c0_seq9 | |||
| LaSOC1-4 | comp129709_c0_seq16 | MN790750 | |
| NC | comp129709_c0_seq2 | ||
| LaAGL1 | comp129017_c0_seq15 | MN790751 | |
| LaAGL11 | comp128471_c0_seq14 | MN790752 | |
| LaAGL42 | comp128471_c0_seq18 | MN790753 | |
| ERF | LaERF017 | comp124322_c0_seq3 | MN790754 |
| LaERF3 | comp129386_c0_seq9 | MN790755 | |
| GRAS | LaSCL29 | comp114072_c0_seq3 | MN790756 |
| AP2 | LaAP2-1 | comp128327_c0_seq16 | MN790757 |
| LaAP2-2 | comp122930_c0_seq2 | MN790758 | |
| DOF | LaHCA2 | comp120092_c0_seq3 | MN790759 |
| C3H | LaOZF2 | comp111742_c0_seq2 | MN790760 |
| MYB_Related | LaTRFL6 | comp128112_c0_seq2 | MN790761 |
| G2-Like | LaPHL1 | comp130729_c0_seq1 | MN790762 |
NC indicates that the full-length cDNA for this transcript was not cloned successfully. LaCAL, Larix kaempferi CAULIFLOWER; LaAGL2-1, L. kaempferi AGAMOUS-Like 2-1; LaAGL2-2, L. kaempferi AGAMOUS-Like 2-2; LaAGL2-3, L. kaempferi AGAMOUS-Like 2-3; LaSOC1-1, L. kaempferi SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1-1; LaSOC1-2, L. kaempferi SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1-2; LaSOC1-3, L. kaempferi SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1-3; LaSOC1-4, L. kaempferi SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1-4; LaAGL1, L. kaempferi AGAMOUS-Like 1; LaAGL11, L. kaempferi AGAMOUS-Like 11; LaAGL42, L. kaempferi AGAMOUS-Like 42; LaERF017, L. kaempferi Ethylene-Responsive Transcription Factor ERF017; LaERF3, L. kaempferi Ethylene-Responsive Transcription Factor 3; LaSCL29, L. kaempferi Scarecrow-Like 29; LaAP2-1, L. kaempferi APETALA 2-1; LaAP2-2, L. kaempferi APETALA 2-2; LaHCA2, L. kaempferi High Cambial Activity 2; LaOZF2, L. kaempferi Oxidation-Related Zinc Finger 2; LaTRFL6, L. kaempferi TRF-Like 6; LaPHL1, L. kaempferi PHR1-Like 1.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhang, Y.; Zang, Q.-L.; Qi, L.-W.; Han, S.-Y.; Li, W.-F. Effects of Cutting, Pruning, and Grafting on the Expression of Age-Related Genes in Larix kaempferi. Forests 2020, 11, 218. https://doi.org/10.3390/f11020218
AMA Style
Zhang Y, Zang Q-L, Qi L-W, Han S-Y, Li W-F. Effects of Cutting, Pruning, and Grafting on the Expression of Age-Related Genes in Larix kaempferi. Forests. 2020; 11(2):218. https://doi.org/10.3390/f11020218
Chicago/Turabian StyleZhang, Yao, Qiao-Lu Zang, Li-Wang Qi, Su-Ying Han, and Wan-Feng Li. 2020. "Effects of Cutting, Pruning, and Grafting on the Expression of Age-Related Genes in Larix kaempferi" Forests 11, no. 2: 218. https://doi.org/10.3390/f11020218
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.