
Floristic Legacies of Historical Land Use in Swedish Boreo-Nemoral Forests: A Review of Evidence and a Case Study on Chimaphila umbellata and Moneses uniflora
Department of Ecology, Environment and Plant Sciences, Stockholm University, 10691 Stockholm, Sweden
Forests 2022, 13(10), 1715; https://doi.org/10.3390/f13101715
Submission received: 5 September 2022
/
Revised: 3 October 2022
/
Accepted: 14 October 2022
/
Published: 18 October 2022
(This article belongs to the Section Forest Biodiversity)
Abstract
:Many forests throughout the world contain legacies of former human impacts and management. This study reviews evidence of floristic legacies in the understory of Swedish boreo-nemoral forests, and presents a case study on two currently declining forest plants, suggested to have been favored by historical use of forests. The review provides evidence of forest remnant populations of 34 grassland species. Thus, many floristic legacies have their main occurrence in semi-natural grasslands, but maintain remnant populations in forests, in some cases more than 100 years after grazing and mowing management have ceased. Despite less information on true forest understory plants appearing as legacies of historical human use of boreo-nemoral forests, a putative guild of such species is suggested. The case study on two species, Chimaphila umbellata and Moneses uniflora (Pyroleae, Ericaceae) suggests that both species are currently declining, mainly due to modern forestry and ceased livestock grazing in forests. Chimaphila maintains remnant populations during decades, due to its extensive clonal capacity and its long-lived ramets. Moneses is more sensitive, due to a lower stature, weaker clonal capacity and short-lived ramets, flowering only once during their lifetime. Thus, Moneses have more transient occurrences, and will decline rapidly under deteriorating conditions.
1. Introduction
Historical legacies of former human activities are common in forests throughout the world, e.g., [1,2,3,4]. In addition to material remains of these activities, associated with for example settlements, e.g., [5,6,7,8], these legacies may be manifested biologically. Many species, especially plants, remain as a historical signal reflecting past human activities and management regimes, e.g., [9,10,11,12]. This holds true also for boreal forests in Scandinavia, particularly for the southern transition zone, usually termed the boreo-nemoral zone [13], which harbored the largest human population historically. Historical legacies in present-day boreal and boreo-nemoral tree communities are well-known and have been recognized in several studies, e.g., [14,15,16]. In contrast, legacies in the boreal and boreo-nemoral forest understory have been focus of less research [17], for example in comparison with still remaining open habitats such as pastures and meadows, e.g., [18]. Generally, ecological perspectives on historical use of forests are often overlooked, not only in Scandinavia but all over Europe [19].
Boreo-nemoral forests in Scandinavia have been much influenced by humans ever since the arrival of agriculture around 4000 BC [20]. The impact has since that time more or less continually increased [21], although interrupted by periods of population decline, for example due to wars, famine and disease [22]. Forests were cleared and burned to make room for agriculture, i.e., crop fields and hay-meadows for production of winter fodder, and forests were used for livestock grazing. Slash-and-burn cultivation was practiced regionally [23,24]. Controlled burning was also used to improve grazing conditions in forests [25]. Production of winter fodder was conducted not only in open meadows but also by harvesting twigs and leaves, both in remaining forested areas [26] and in wooded meadows [27,28]. Wetlands, i.e., mires, bogs and shores along streams and lakes were particularly important for production of winter fodder [29,30]. Livestock grazed in forests outside the infields of the farms [31,32,33]. The impact of livestock was massive [34]. The forests in southern Sweden during the 19th century have been described as “a large cow pasture” [35] (p. 395). In addition, forests were used for production of charcoal, tar and potash, e.g., [2,36]. During the 17th to 19th centuries, in areas close to iron production sites, most wood biomass was used for production of charcoal [37,38]. Moreover, one should not forget the people themselves. In large parts of southern and central Sweden, forests which are today used for production of timber and pulp and almost devoid of permanent human inhabitants, were just over a century ago populated by lots of people [39,40,41], smallholder farmers and crofters, maintaining their livelihood by growing some crops, keeping some livestock, managing small gardens, in addition working at nearby located manors, mills, mines, or ironworks.
This plethora of historical impacts largely ceased during a period from the late 19th to mid-20th century, primarily due to a modernization of agriculture and the introduction of modern production forestry, but also promoted by industrialization and urbanization [21]. This led to a landscape transformation which can be illustrated by the changing area of semi-natural grasslands. These grasslands were managed by grazing or mowing, they were not much influenced by fertilizers, sowing or plowing, and they usually have a long history of management, for centuries or in some cases even millennia [18,42]. Semi-natural grasslands typically harbor a very high local-scale plant species richness, e.g., [43]. In one study area in southern Sweden, it was estimated that over 96% of semi-natural grasslands in 1900 was lost in 2013, most of them transformed to forest [44]. The area used for forest grazing has decreased with over 98% since 1927 [45].
The forest cover in southern Sweden has increased during the last 100 years [46], and the same holds for the standing biomass of wood in these expanding forests [47,48]. The forests we have today are generally much denser, darker and less disturbed than they have been for a very long time, perhaps ever. Increased shading from a dense tree canopy has been identified as one major driver behind recent changes in the boreal forest understory, together with increased nitrogen deposition and climate warming [49]. There is an ongoing decline of several plant species in the boreo-nemoral forest zone. The species in the forest understory that have decreased most during the 20th century are those with a small stature, i.e., low-growing species that are particularly sensitive to increased shade, either from the canopy or from taller understory species [50].
As a consequence of these changes in the boreo-nemoral forest environment, we may expect that there are floristic legacies of the historical, no longer occurring, human use of the forests (Figure 1). The main objective of this paper is to review and synthesize studies on floristic legacies in Swedish boreo-nemoral forest understory.
The rationale for studying floristic legacies is that they contribute to the biological diversity in forest ecosystems, and that they can be regarded as biological cultural heritage, e.g., [17,34,51]. In addition to being interesting in their own right, such legacies are also informative for developing appropriate conservation plans, e.g., [52], and they may be useful for predicting the future of forest ecosystems, e.g., [53,54]. The paper ends with a brief discussion on the question if, and if so, how, these legacies should be preserved.
This paper also has a second objective. The species comprising the floristic historical legacies in boreo-nemoral forest understory can be seen as belonging to three different categories. Firstly, there are species which have their main occurrence in open grassland systems, and which maintain populations in the current forest environment. Secondly, there are species which are not found in open grassland systems—they are true forest species—but they were nevertheless favored by the historical impacts creating more open, lighter, and disturbed forests. As the first group of species so far has received more attention than the second, this paper also contains a case study comparing two species belonging to this latter category.
A third category of floristic legacies is cultivated plants at forest settlements such as crofts and small farms, abandoned during the period of transformation of forest land use between the late 19th and early 20th century. Many of these cultivated plants are still found at the same sites, sometimes as the only remaining trace of the former settlement. Such cultivated plants, the most common being Ribes uva-crispa L. (gooseberry), Syringa vulgaris L. (lilac), and Malus domestica Borkh. (apple), are often encountered in boreo-nemoral forests, e.g., [41,55]. However, cultivated garden plants are beyond the scope of this paper, and are not treated further.
Before reviewing evidence regarding floristic legacies, and presenting the case study, below follows a brief overview of ecological mechanisms behind floristic legacies, and some remarks on methodological issues.
2. Ecological Mechanisms behind Floristic Legacies
In Longman Dictionary of Contemporary English, legacy is defined as “a situation that exists as a result of things that happened at an earlier time”. Applied in the context here, this mean that some previous (henceforth “historical”) ecological condition—such as any of the plethora of effects mediated by human actions listed above in the Introduction—have promoted establishment or an increasing population of a certain plant species. Furthermore, in order to be meaningful, identifying a species as a legacy presumes that the species would not exhibit its current occurrence, unless the historical ecological condition had existed.
Given this broad definition, there are two main mechanisms behind floristic legacies. Firstly, the species still persists even though the particular ecological conditions necessary for its establishment and population growth have ceased to exist. For example, we may envisage a species that established at a site as a direct result of effects of livestock grazing in the forest, the species would otherwise not have been there, but the species still persists at the site even though livestock grazing has for long ceased.
The second mechanism is when the ecological conditions altered by historical land use still persists, affecting the current flora, even though the agents causing the altered ecological conditions have ceased to exist. For example, we may envisage a historical management regime altering soil conditions and that these soil conditions persist and affect the current plant species composition. Several studies conducted in forest of continental Europe have documented this phenomenon [5,56,57,58]. Although one could perhaps say the legacy in this case is the ecological condition rather than the species associated it, this semantic issue is here disregarded, and records due to this mechanism are recognized as floristic legacies.
The first mechanism—a species is present despite the ecological conditions favoring it has ceased to exist—is often referred to as “remnant populations” [59,60]. By definition, a positive population growth rate (λ > 1) implies an increasing population, while λ < 1 implies that the population is decreasing, and that it will ultimately go extinct. Incorporating a spatial component, maintenance of a local population can be achieved even if λ < 1, namely if there is a flow of individuals into the population from elsewhere; the local population being a sink population [61]. Many species have a patchy distribution composed of several more or less interconnected local populations, a metapopulation [62]. Some of these local populations may be sink populations, even though the metapopulation as a whole is persistent. However, as legacies refer to a time sequence of events (past and present), one needs to incorporate a temporal component. Eriksson [59] defined the concept remnant population as a local population which is maintained despite λ < 1, without being a sink population. In order to make sense, this concept necessitates defining a timescale which is considered relevant. Many plants have life stages that can be very long-lived, viewed in a timescale of decades and even centuries. For example, this holds for many clonal plants [63] and for plants with seed banks [64]. Given that this corresponds to a timescale often used when investigating vegetation changes, recognizing remnant populations does make sense.
Remnant populations can be seen as “sink populations in time”. While an ordinary sink population is maintained by individuals moving in space from a local population with λ > 1 to the sink population, remnant populations are maintained by individuals “moving in time” from a temporal stage with λ > 1 to a later temporal stage with λ < 1 (Figure 2). Potentially, this may enable the population to again achieve λ > 1 if the conditions improve, a phenomenon referred to as remnant population dynamics [59]. However, we here focus on the remnant populations as such, having λ < 1. Species with remnant populations contribute to an extinction debt [65], and the expected time to extinction for a remnant population corresponds to the time it takes for the historical signal to vanish, i.e., the historical time depth for the floristic legacy.
An additional way of looking at mechanisms behind floristic legacies is to use the niche concept. There are several different definitions of ecological niche (see [66] for an overview). The most common conception is that the ecological niche encapsulates the ecological parameter space within which the species can exist, i.e., have a population growth rate λ ≥ 1 [67]. This is based on Hutchinson [68], who also introduced the useful distinction between fundamental and realized niches. The fundamental niche includes all aspects of the ecological parameter space in the absence of other species, whereas the realized niche is a part of the fundamental niche restricted by interactions with other species [66]. Even though Hutchinson [68] stressed competition as the major interspecific interaction, the factors restricting the fundamental niche may be other interactions, such herbivory, or even abiotic conditions. In addition, a species’ fundamental niche may differ among life cycle stages. This has been termed “ontogentic niche shifts” [69]. One example of such ontogenetic niche shifts in boreal forest is understory Vaccinium shrubs, where the conditions required for seedling recruitment are partly or fully outside the niche for adults [70]. Indeed, Grubb [71] stressed that understanding the dynamics of plant communities may require that a regeneration niche is recognized, specifying the totality of conditions needed for recruiting new individuals to a population. These conditions may be completely different from the conditions required for an established adult plant to thrive.
By definition, floristic legacies occur because historical land use once made it possible for species to express their fundamental niche at sites where they otherwise would not have been able to establish and survive. The fact that the species still remain there reflects that the current ecological parameter space is within the fundamental niche for at least a part of the plant’s life cycle. Understanding the mechanism behind a legacy therefore includes identifying which part of the life cycle that is outside the ecological parameter space, and why this is so.
3. Some Remarks on Methodological Issues
Considering that we generally lack time series data on population occurrences covering both the historical conditions and the present ones, there are methodological problems to detect floristic legacies empirically. A record of a plant species in the field (for example as a putative remnant population) may reflect that the species is present because of historical conditions, and has remained at the site long after these conditions have changed, or that the species has dispersed there later, and may be a sink population established from a source population elsewhere.
Distinguishing between these alternatives is difficult. One method is to couple present-day floristic composition with the site’s history as revealed by historical maps. This method is commonly used in research conducted in semi-natural grasslands where the impact of historical management on present-day diversity and species composition has been shown in many studies [18, and references therein]. Thus, historical maps, or other sources of historical information, can be used to identify historical ecological conditions in present-day forests, and these can be compared with data on species’ current occurrence and performance.
Another approach is to conduct demographic studies combined with modelling, in order to estimate how population growth rate varies with environmental conditions reflecting different land use trajectories, e.g., [72,73,74]. If no demographic data is available, there are other ways to detect a likely remnant population, for example based on the existence of life-cycle stages enabling population persistence even in the absence of seed production and seedling recruitment, such as subterranean propagating structures and rhizomes, re-sprouting ability, and seed banks [75].
For floristic legacies resulting from persistent ecological conditions (e.g., soil) caused by historical management, the problem is not to associate the ecological conditions with the presence of certain species (this can be studied by experiments). The problem is rather to show that it was the historical management per se that caused the ecological conditions, and that these conditions do not occur irrespective of the historical management.
In light of these methodological difficulties, one needs to accept an element of subjectivity in assessing floristic legacies, based on what is the most reasonable and likely interpretation. This is something the ecologist has in common with historians trying to assess causation in historical processes, e.g., [76]; the conditions or events considered are no longer possible to observe, and experiments can rarely be employed. In the review below, it is thus unavoidable that there is an element of interpretation when assessing whether records of species in forests are legacies of historical land use.
4. Grassland Species as Floristic Legacies of Historical Land Use in Forests
Table 1 contains a list of grassland species referred to in the studies reviewed below in this section. In order to avoid over-representing grassland species recorded in forests, a conservative criterion was used. This criterion was derived from a commonly used overview of semi-natural grassland plants in Sweden [77], which lists species according to their dependence of grassland management, grazing or mowing, for maintaining populations, i.e., exhibiting λ ≥ 1. Four categories of grassland species were recognized, defined based on their sensitivity to abandonment of management. The categories A, B and C include species which decline after abandonment, with A being those responding most rapidly, B is intermediate, and C being the least sensitive, responding last. Table 1 only lists grassland plants assigned to the categories A and B. Thus, “grassland plants” henceforth mean species having their core occurrence in semi-natural grasslands.
The strongest evidence for floristic legacies comes from studies of present-day forest sites with a known history as pasture and meadow, and where the studies include control sites with similar forest lacking this history. One study [78] investigated species composition at forest clear-cut sites with a historical background as hay-meadow, based on maps from the late 19th century, and compared them with adjacent sites which were not used as meadow historically. It was found that clear-cuts with a history as meadow had 36% higher species richness. Overall, 48 species were significantly or close to significantly associated with clear-cuts with a history as meadow. Seven of these were species (using the conservative definition) categorized as grassland plants (Table 1). In a similar study [60], species composition was compared between forest sites with a history as meadow (also based on late 19th century maps) and sites which have had a continuous coniferous forest cover. The species composition at the forest sites was also compared with present-day species-rich semi-natural grasslands, managed by livestock grazing. The species-richness was 30% higher at forest sites with a history as meadow. Overall, 77 species were significantly or close to significantly associated with forests with a history as meadow, and nine of these were species categorized as grassland plants (Table 1). The forest sites with a history as meadow were also more similar to current grasslands than forests with continuous coniferous cover. The authors of both these studies interpreted the results as evidence of remnant populations.
Even if this interpretation seems reasonable, the result that the overall species richness was higher at the clear-cut/forest sites with a history as meadow (i.e., not only for grassland plants) illustrates one of the methodological problems mentioned in the previous section. The sites used as meadow historically may have been chosen by the farmers because they were initially identified as more productive than adjacent land. Alternatively, the meadows were improved as a result of the management, and this also affected species richness. Historical meadows were exceptionally rich in plant species, e.g., [27,79] and management of wooded meadows improved soil conditions because deep tree roots increased the available nutrient pool, nutrients were recycled to the top soil after leaf shedding, and leaf litter created favorable habitat for earthworms [36] (p. 204). Although there is no additional information enabling us to distinguish between these alternative explanations, the association between historical management and current species richness found [60,78] holds as a legacy of historical management.
Combining historical maps with current records of species occurrences can provide a basis for modelling how spatial dynamics of species develop over time. This approach was used to examine the influence of land-use history on the distribution of Succisa pratensis, a perennial often found abundantly in semi-natural grasslands with long history of management [72]. The main conclusion was that the current distribution, which included present-day forest where historical management ceased 100–150 years ago, reflected the landscape in the 19th century. Incorporating assumptions of dispersal rates, and assuming “weak” dispersal, led to the conclusion that the current occurrence pattern of S. pratensis is strongly affected by the historical (19th century) landscape. Assuming a higher dispersal rate, the historical effect became weaker, but was still present. Overall, the expected time it would take for the regional population of S. pratensis to reach something close to an equilibrium with the landscape configuration of habitats was found to be in the magnitude of several centuries. Considering the rate of historical and ongoing landscape changes, S. pratensis is thus likely to never reach such an equilibrium.
These results are in line with other studies that have examined to what extent present-day landscape configuration of grasslands is related to site-specific species richness. The prediction from theory is that both site area and connectivity of grasslands have a positive effect of their species richness. Two studies [80,81] conducted in Swedish and Estonian semi-natural grasslands, respectively, found that neither present-day area nor connectivity were related to site-specific species richness. In contrast, when using maps depicting grassland configuration 50–100 years ago, connectivity (both studies) and area [81] turned out to be strongly positively related to species richness. This suggests that there are time-lags in the correspondence between landscape habitat configuration and species occurrences, and that these time-lags are in magnitude of 50–100 years. It was estimated that there was an extinction debt comprising about 40% of the grassland species [81]. Thus, extinction (albeit slow) is currently a dominating process. This was also the conclusion for Succisa pratensis [72]. The current dynamics of this species was dominated by population extinction processes.
Demographic analyses of the grassland perennial Primula veris was conducted to estimate population growth rate in populations at sites where grassland management had ceased, with varying time since abandonment [74]. A general conclusion was that the time to extinction ranged from decades to centuries. Field studies were employed to validate the results from the demographic analyses, confirming that P. veris occurred abundantly even at forest sites where grassland management had ceased in the early 20th century. Thus, as in Succisa pratensis, a large fraction of occurrences of P. veris in the modern forested landscape most likely consists of remnant populations.
There are also studies with the limitation that they focused on surveys of sites with a known history of human management, but lacking controls and demographic modelling. One study [82] surveyed forest sites with a history as semi-natural grassland, i.e., pasture or meadow, managed until 60–100 ago. The surveys were compared with investigated records from still managed semi-natural grasslands in the same landscape. The objective was to assess which grassland species were still present in the forests, and whether they had declined. Of 67 species for which data allowed analysis, 44 species had declined but were still present at the former grassland sites. Of these, 27 species are categorized as grassland plants (Table 1).
A similar study [32] focused on five selected species (Table 1). For each of these species, the number of patches (a localized occurrence of the species) was counted within a study area of c. 2 km2 comprising both still managed grassland and forest which in 1854 was documented as pasture. For all five species, the majority (between 62% and 91%) of the patches were located in forest. As the study area was quite small, sink populations cannot be ruled out. However, considering that the abundance of patches was much higher in the forest environment, and referring to the time-lags mentioned above [80,81], it is likely that the forest patches were mostly remnant populations.
Another study [83] was conducted on the annual Rhinanthus minor, and the two perennials Campanula rotundifolia and Primula veris using two field study areas, both comprising still managed semi-natural grasslands and present-day forest on land that previously (in the 1940s) were managed as grassland. The species’ occurrences were investigated along a hypothetical successional gradient from open localities to localities with full forest cover. The annual R. minor rapidly disappeared after abandonment of grassland management, but the two perennials C. rotundifolia and P. veris maintained populations along the successional gradient, for C. rotundifolia even in the present-day closed forest. For both these species the local population size and flowering frequency declined along the successional gradient. This study also included seed addition experiments (estimating germination, but not seedling survival), and the results suggested that both species were capable of germinating also in the closed forest environment. Thus, as with the example above [32], sink populations cannot be ruled out, but the interpretation is that the distribution pattern largely reflects occurrence of remnant populations.
Still another study [84] surveyed land that formerly was used as meadow or field, and after abandonment in the early 20th century now has encroached to forest. This study did not present a complete species list, but listed ten grassland species recorded at the former meadows (Table 1). A notable observation from this study was that two of the grassland species (Polygala vulgaris and Ajuga pyramidalis) were significantly more abundant at former crop fields than on former meadows. As these crop fields are not likely to have harbored these species at the time when the fields were managed for growing crops, this provides circumstantial evidence that the species have later colonized suitable locations nearby their presumed original occurrence in the meadows. This suggests that some species are able to maintain populations at refuges where they were not present originally, but which are located in the near surroundings of the original sites. This implies spatial dynamics among small suitable sites in the forest environment, promoting maintenance of grassland species, if not exactly at the same site as the historical management, so in the vicinity.
A similar interpretation was made in a study of historical management other than livestock grazing and mowing [85], investigating species richness and species occurrences at charcoal production sites, remaining as so-called charcoal kiln platforms. Charcoal production was massive historically (until early 20th century) in many areas of present-day forest, and charcoal kiln platforms are usually recognizable as circular areas (about 10 m in diameter) with an aberrant flora. It was found that charcoal kiln platforms harbored a higher species richness than adjacent control sites, and several species occurred there significantly more frequent. One group of species which was more frequent at charcoal kiln platforms was low-abundant (occurring at ≤ 9 of the 50 sites investigated) species categorized as grassland plants; 11 species according to the conservative definition (Table 1). The interpretation was that charcoal kiln platforms, which have different soil conditions than adjacent controls (higher pH and concentrations of P, K, Mg and Ca), and a lower abundance of the otherwise dominating ericaceous shrubs (Vaccinium spp., Calluna vulgaris (L.) Hull, provide small-scale refuges for grassland species which historically occurred in the semi-open, disturbed, and grazed forests used for charcoal production.
In a companion study [41], some records on remaining grassland plants were made in a re-survey otherwise focusing on cultivated plants at abandoned crofts, housing people formerly working in the region as miners, or as farmers part-time working with for example charcoal production. These crofts were abandoned between the 19th century and early 20th century, and were surveyed for the first time in 1967. The whole survey was repeated in 2019. Four grassland species were recorded both 1967 and 2019 (Table 1), tentatively interpreted as remnant populations.
Thus, in all, Table 1 lists 34 grassland species, selected based on a conservative criterion for having their core abundance in managed semi-natural grasslands, for which the reviewed studies provide evidence that they occur as floristic legacies from historical land use in present-day forests. However, note that for two of the 34 species, the categorization as grassland plants has been questioned (see Table 1). The strength of evidence is variable, both because of the different design of the studies reviewed, and because of the varying number of studies with evidence for each individual species.
As the categorization used for grassland plants [77] was based on the species’ presumed more or less rapid disappearance after abandonment, it may seem contradictory that these species maintain occurrences even long after abandonment of grassland management at sites which now has turned into production forests. However, even if the predicted rate of decline of these species after abandonment of grassland management may have been exaggerated, the categorization of these species as having their core occurrence in semi-natural grasslands is valid.
Relaxing the criterion, for example including category C [77] would have increased the number of species listed as putative floristic legacies. For example, using this relaxed criterion on the results from Ref. [78] would have increased the number of species from seven (Table 1) to 15. For Ref. [82], the number of species would increase from 27 to 41. In a survey of 50 sites in a boreo-nemoral forest area, there were in total 127 understory species recorded [85]. If figures in this magnitude are used as a baseline, and acknowledging that presenting exact figures is difficult as the surveys were conducted in different study areas, this review suggests that a considerable fraction (in the magnitude of at least a third) of the understory plant species richness in boreo-nemoral forests may consist of floristic legacies of historical forest land use. In addition, ecological conditions remaining from historical land use (meadows, charcoal production) are associated with a higher local species richness, also including other species than grassland plants.
Even with the caveats associated with presenting exact figures in mind, the general conclusion is that boreo-nemoral forests contain numerous floristic legacies of grassland species, reflecting these forests’ historical land use, for hay-production, livestock grazing, and for production of charcoal, and that this legacy makes up a large fraction of the present-day plant species diversity in these forests.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Grassland plants for which there is evidence that they occur as legacies of historical land use in boreo-nemoral forests (Marked with X). The category “grassland plants” was based on [77]. For species marked with * the categorization has been questioned [82,84,85]. “Reference” is according to the number in the reference list.
Table 1.
Grassland plants for which there is evidence that they occur as legacies of historical land use in boreo-nemoral forests (Marked with X). The category “grassland plants” was based on [77]. For species marked with * the categorization has been questioned [82,84,85]. “Reference” is according to the number in the reference list.
| Species | References | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| [78] | [60] | [72] | [74] | [82] | [32] | [83] | [84] | [85] | [41] | |
| Ajuga pyramidalis L. | X | X | X | X | ||||||
| Alchemilla glaucescens Wallr. | X | |||||||||
| Antennaria dioica (L.) Gaertn. | X | |||||||||
| Anthoxanthum odoratum L. | X | X | X | X | ||||||
| Campanula rotundifolia L. | X | X | X | X | X | |||||
| Carex leporina L. | X | X | ||||||||
| Carex pallescens L. | X | X | ||||||||
| Carex pilulifera L. | X | |||||||||
| Carex spicata Huds. | X | |||||||||
| Cerastium fontanum Baumg. | X | |||||||||
| Danthonia decumbens (L.) DC. | X | X | ||||||||
| Euphrasia stricta J.P. Wolff ex J.F. Lehm. | X | |||||||||
| Festuca ovina L. | X | |||||||||
| Festuca rubra L. | X | |||||||||
| Juncus filiformis L. | X | |||||||||
| Lathyrus linifolius * (Reichard) Bässler | X | X | ||||||||
| Leucanthemum vulgare Lam. | X | X | X | X | ||||||
| Lotus corniculatus L. | X | X | X | X | ||||||
| Luzula campestris (L.) DC. | X | X | ||||||||
| Pilosella officinarum Vaill. | X | X | ||||||||
| Pimpinella saxifraga L. | X | X | ||||||||
| Plantago lanceolata L. | X | X | X | |||||||
| Polygala vulgaris L. | X | X | X | X | ||||||
| Primula veris L. | X | X | X | X | X | |||||
| Ranunculus acris L. | X | X | X | X | ||||||
| Ranunculus auricomus L. | X | |||||||||
| Rhinanthus minor L. | X | |||||||||
| Succisa pratensis Moench | X | X | ||||||||
| Trifolium pratense L. | X | X | X | |||||||
| Trifolium repens L. | X | X | ||||||||
| Veronica chamaedrys L. | X | X | X | X | ||||||
| Veronica officinalis * L. | X | X | ||||||||
| Veronica serpyllifolia L. | X | |||||||||
| Viola canina L. | X | |||||||||
5. Forest Species as Floristic Legacies of Historical Land Use in Forests
As mentioned in the Introduction, it is well-known that tree composition, structure and diversity of present-day boreo-nemoral forests maintain features reflecting their history, e.g., [15,34,51,86], particularly referring to deciduous tree species [35,87,88,89]. For subordinate trees and shrubs, historical land use affects the present-day composition of a guild of woody fleshy-fruited plants [90]. Species richness of this guild was higher at sites with a history as open grassland, and five species were positively associated with this land-use history (Lonicera xylosteum L., Prunus padus L., Ribes alpinum L., Rubus idaeus L., and Viburnum opulus L.). Additionally, for other taxa than vascular plants, current occurrences have been suggested to reflect forest history, for example for fungi and bryophytes [91], insects [92] and birds [93,94]. Time-lags similar to those described above for grassland species have been documented for various epiphytes on oaks, where their occurrence reflects the historical rather than the current composition and distribution of the oak stands [95,96].
Although not much studied, scattered in the literature there are also putative cases of historical cultural impacts on distribution and abundance of forest understory plants. Actaea spicata L., Lathyrus vernus (L.) Bernh., Galium odoratum (L.) Scop., and Festuca altissima All. are examples of plants whose occurrences may reflect forest history [92]. The three first (A. spicata, L. vernus, G. odoratum) are possible legacies from historical wooded meadows [27,79,97]. A study of L. vernus along its northern distribution range suggested that current occurrences partly reflect forest history [98]. However, for F. altissima it may rather be the lack of strong historical management impact of grazing or mowing that promotes its current occurrence [99]. Other tentative examples are plants suggested to have been favored by livestock grazing, e.g., Lycopodium spp., Pyrola spp., Goodyera repens (L.) R.Br. and Linnaea borealis L. [100], Carex digitata L., Luzula pilosa (L.) Willd., Maianthemum bifolium (L.) F.W. Schmidt and Melampyrum spp. [34], Cephalanthera rubra (L.) Rich. [101], and Pulsatilla vernalis (L.) Mill. [102,103]. Diphasiastrum [Lycopodium] complanatum (L.) Holub may have been additionally favored by intentional burning [34,104]. Both the latter species have been recorded as declining in southeastern Sweden, with c. 52% (Diphasiastrum) and 46% (Pulsatilla) between 1929 and re-surveys conducted from the 1990s onward [105]. Goodyera repens and Pulsatilla vernalis are red-listed in Sweden as Vulnerable (VU) and Endangered (EN), respectively [106].
Generally, however, studies of understory forest plants as historical land-use legacies are not as common as for the grassland species reviewed in the previous section. To balance this relative paucity of research, below follows a synthesis of published and new information on two forest understory species, whose present-day occurrences in boreo-nemoral forests are likely floristic legacies of historical land use [17,101]: Chimaphila umbellata (L.) Barton (Figure 3) and Moneses uniflora (L.) A. Gray (Figure 4). Comparing these two species will also illustrate how differences in the plant life cycle influence species response to a changing forest environment, i.e., responses to the abandonment of historical land use and its replacement with modern production forestry.
6. The Case Study
6.1. Description of the Species
Chimaphila umbellata and Moneses uniflora (henceforth referred to by their genus name only) belong to the tribe Pyroleae of the family Ericaceae. Within this tribe, Chimaphila and Moneses are closely related sister taxa [107]. Species of Pyroleae form intricate associations with mycorrhizal fungi with varying degrees of mycoheterotrophy [108], i.e., they derive carbon from a fungal source. As far as known, all species of Pyroleae possess partial mycoheterotrophy (or mixotrophy) [109], except Pyrola aphylla Sm. which is fully mycoheterotrophic [110]. This means that they use fungal carbon only during parts of their life, or, if throughout their life, only partially. All Pyroleae have so-called dust seeds [111], i.e., minute seeds produced in vast numbers with a very small and undifferentiated embryo, often lacking any nutrient reserves. Pyroleae have subterranean juveniles which during development, taking place below ground during several years, act as parasites on mycorrhizal fungi [109,112]. Thus, germination and early development from dust seeds of the Pyroleae are completely dependent on fungal associates supplying carbon and other nutrients.
Chimaphila (Figure 3) is a perennial evergreen dwarf shrub with leathery leaves, and with an extensive capacity of clonal growth with long creeping subterranean rhizomes [113,114]. It may have over 50 m of interconnected rhizomes belonging to the same clone [115] (clones are henceforth referred to as “genets”). The shoots (henceforth “ramets”) are typically 10–20 cm high. Flowering ramets usually have 2–4, sometimes up to 6 flowers. The ramets are iteroparous, i.e., they may produce flowers during several years. It has been suggested that the ramets have a life span of up to six years [113], but considering that they have a woody base and can flower repeatedly, it seems likely that they may reach a higher age. The flowers produce nectar, and they are pollinated by bumblebees [116]. A capsule contains c. 7900 seeds, and a ramet may thus produce over 25,000 seeds [117]. Chimaphila utilizes a wide range of fungi during germination, but shows a tendency of becoming more specialized during juvenile ontogeny [118]. Examples of common fungi in developing juveniles were the basidomycetes Cortinarius and Piloderma [118]. Chimaphila is mycoheterotrophic only as juvenile, adults are autotrophic [107,109]. Chimaphila has a circumboreal distribution [119]. In Sweden it occurs mainly in the boreo-nemoral zone, most typically in pine forest heaths on sandy soils, but also in mixed pine-spruce forests [114]. Chimaphila is red-listed as Endangered (EN) in Sweden [106].
Moneses (Figure 4) is a low growing perennial herb, with overwintering green leaves, almost in a rosette, close to the ground. Although the extent has not been studied in detail, Moneses is clonal. It has only short rhizomes [120], but spreads clonally by producing ramets from horizontally growing roots [121]. After 2–4 years, probably after reaching over a certain size threshold, a ramet produces one single flower bud, which lives over the winter to produce a flower the following year [122]. After completion of seed production, the ramet dies, without producing any lateral shoots [122]. Moneses is buzz-pollinated by bumblebees [116]. A capsule (and thus a ramet) produces c. 7300 seeds [117]. Moneses utilizes a range of fungi during germination, and as Chimaphila, it shows a tendency of becoming more specialized during ontogeny [118]. Examples of common fungi in developing juveniles were the ascomycete Meliniomyces and the basidiomycetes Russula and Tylospora [118]. Adults have been found autotrophic [109], but other studies suggest that also adults are partially mycoheterotrophic, ultimately becoming specialized on one genus of fungi, Tylospora [123]. Moneses has a wide distribution in the northern hemisphere, where it occurs in slightly moist coniferous forests with moss-dominated ground cover and in forests with sandy soils, but it is also occasionally found in ditches along road verges and at former fields now covered with Norway spruce (Picea abies (L.) H. Karst.) but with only a sparse cover of other field layer plants [105,124]. Thus, it seems able to exploit sites which have been relatively recently disturbed. Moneses occurs over almost the whole of Sweden except at high altitudes and in the southernmost region [125].
6.2. Population Decline
From the province of Uppland, southeastern Sweden, the occurrences of Chimaphila have declined by 81% between 1929 and the 1990s [105]. As being red-listed as Endangered, Chimaphila has received some special attention in floristic surveys. A summary was made of the state-of-the-art for Sweden based on surveys conducted from the 1990s onwards [101]. Re-surveys (on average with a 10-year interval) suggested that the majority of the populations were either extinct or had decreased (Figure 5), and more than half of the population extinctions were associated with modern forest management (clear-cuts, plantations) or by increasing dominance of Vaccinium and graminoids.
A study was conducted on a selection of Chimaphila populations in two provinces of Sweden, Uppland and Södermanland (southeastern Sweden), based on field studies in 2013 [114]. In 2021, a re-survey was made of these populations (Appendix A, Table A1). The distribution of extinct, decreasing and increasing populations was similar to results for Sweden as a whole (Figure 5), wherefore it is reasonable that the results [114] on the relationships between populations and environmental factors are generally representative for Chimaphila. The average number of ramets in 2013 of populations that were extinct 2021 was significantly smaller than those populations still persisting (75 vs. 313; p = 0.0466; Appendix A Table A1). Both population size (number of ramets) and fruit-set in Chimaphila were negatively affected by competition from dominating Vaccinium myrtillus L. and graminoids, and flowering was negatively affected by shading from the canopy [114]. Increasing soil nitrogen was associated with decreasing seed production [114]. Figure 6 shows the forest structure where Chimaphila thrives, an open-canopy pine forest with low cover of those otherwise dominating graminoid and Vaccinium competitors. This site harbored the largest Chimaphila population (>3500 ramets) in the re-survey.
The data presented above suggests that Chimaphila is currently declining generally, but occasionally have large and even increasing populations. The majority of the occurrences of Chimaphila exhibit features suggesting that they are remnant populations, slowly declining from a time where conditions were more favorable.
For Moneses, there is less available data. To provide an overview of the development of Moneses abundance and distribution, one has to rely on several sources, including a field survey conducted in 2022 in the same two Swedish provinces, Uppland and Södermanland, as was used for Chimaphila. There are published floras for both these provinces, useful as a starting point. In Uppland, where the surveys were conducted mainly during 1990s, Moneses was recorded as still relatively common [105], although the authors of the flora suggested that Moneses is probably declining rapidly. In the flora of Södermanland [124], based on surveys conducted mainly during the 1980s, Moneses was recorded as relatively rare. It was remarked that most populations were small, and that Moneses has declined sharply ever since the 19th century when it was regarded as common, a decline the authors suggest is due to the abandonment of livestock grazing in forests.
Sweden has a citizen science based information site of species occurrences, “Artportalen” [126], where records of species are reported. The Swedish Association for Botany (Svenska Botaniska Föreningen) in 2015 selected Moneses as the “species of the year”, encouraging botanists to report finds of the species. Moneses may be easy to overlook when it is not flowering and is thus potentially under-reported, but this call is likely to have increased the intensity and accuracy of the reporting.
Finds reported in Artportalen from 2015 onwards, from Uppland and Södermanland, was downloaded. A selection of sites of Moneses reported in Artportalen were also re-surveyed in 2022.
There were a total of 133 records of Moneses in Artportalen for these two provinces (a record meaning a site-specific finding of Moneses at one year during 2015–2021), distributed among 79 different sites. The first author of the flora of Södermanland, Hans Rydberg, one of the most experienced field botanists in this province, on request gave the following information: “I almost never encounter Moneses anymore, and when it occasionally happens there are only a few tiny plants” [127]. Furthermore, of all records of Moneses downloaded from Artportalen 2015–2021, 75% was either just a notification that this species was present, or a record of a small patch with ≤ 20 flowering ramets.
The results of the re-survey conducted 2022 are summarized Figure 7 and in Appendix A, Table A2. Of the 25 re-surveyed sites, Moneses was found at four sites, two interpreted as increasing and two as decreasing. These four were all at sites where Moneses was previously (2015–2016) recorded as having a relatively large population (at least 50 flowering ramets, or a total of 450 ramets). All smaller populations were gone in 2022. Among the 21 sites where Moneses were extinct, four had been subjected to clear-cutting. Thus, Moneses disappears even if the forest still stands. During the surveys, a search was conducted in the surroundings of each of the investigated sites, to look for previously unreported populations. Not one single occurrence was recorded.
The largest Moneses population (in terms of flowering ramets) was found at the same location as for Chimaphila (Figure 6), a semi-open forests with a limited cover of Vaccinium and graminoids. It seems reasonable that large populations of Moneses persist because the ecological conditions resemble those which formerly were common due to livestock grazing in forestsas remarked in [124].
This data, despite its limitations, suggests that Moneses is continually declining. In contrast to Chimaphila, where a majority of sites persist several decades even though declining in size, most Moneses populations are gone. This difference is accentuated by the fact that the time interval between the surveys for Moneses was shorter than for Chimaphila. Thus, the capacity of Moneses to maintain local populations is seemingly more limited than for Chimaphila, and therefore regional persistence of Moneses is more dependent on continuous establishment new genets.
In order to understand this difference between the two species, we need to look into their life histories.
6.3. Comparison of Life Histories
In order to maintain an established genet, the production of new ramets must balance the loss of ramets dying. A difference between the species is that the ramets in Chimaphila live longer. Although no demographic study has been made, the suggestion of a ramet life span up to six years for Chimaphila [113] rather seems as an underestimate, as remarked above. In Moneses, reproductive ramets die after completion of fruit production. A crude estimate is that this occurs in the ramet’s third year of life, on average (B1 in Appendix B), in line with the suggestion [122] that their life span is 2–4 years. In addition, the capacity of clonal propagation in Chimaphila, with its extensive rhizome system [115], exceeds the corresponding capacity of Moneses, which propagate by horizontal roots. These roots lie near the ground surface [121], which may be the reason why Moneses thrives at sites where mosses dominate the ground floor [105,124], i.e., at sites with limited root competition for space. Competing with roots of graminoids, especially, may be difficult for Moneses. The ramets of Chimaphila also have their leaves placed higher than the rosette of leaves in Moneses. Altogether, this implies that genets are larger and considerably more long-lived in Chimaphila than in Moneses, and that Moneses is even more sensitive to competition by potential dominants (Vaccinium spp. and graminoids) in the forest understory.
At the genet level, a balanced population requires that each reproductive genet on average produces one new genet during it life. Both Chimaphila and Moneses produce a vast number of tiny dust seeds, but Chimaphila has the highest seed production. The yearly seed production per seed producing ramet is about 3.4 times higher than in Moneses [117], and to this one must add that Chimaphila ramets can reproduce during several years.
It might be expected that these tiny dust seeds have long dispersal ranges, but in fact over 95% of dust seeds are deposited within 5 m from the seeds source [117]. Thus, the likelihood of a seed finding a suitable site for recruiting a new genet at another location is very small, despite this vast seed production. For both species, suitable sites for germination and juvenile growth depend on presence of fungi, from which the growing juvenile is totally dependent on for extracting resources. Using field experiments at what was judged as potentially suitable sites, seed germination and growth of subterranean juveniles was investigated over three years [118]. Moneses had a higher germination rate than Chimaphila. Of germinated Chimaphila seeds, only 4.2% reached any of the larger size classes of subterranean juveniles. In contrast, 51% of the germinated Moneses seeds reached the larger size classes.
One may speculate that the higher growth capacity of the subterranean juveniles in Moneses is due to this species ultimately becomes more specialized in its use of fungi: Tylospora spp. [123]. Despite its disadvantage in reducing the range of potential hosts, specialization usually implies increasing efficiency of the parasitism [128,129]. For example, the full mycoheterotroph Monotropa hypopitys (Ericaceae), specialized on fungi of the genus Tricholoma [130], also has a conspicuously high growth capacity of its subterranean juveniles [118].
Both Chimaphila and Moneses thrive in open canopy forest where the potential understory dominants, Vaccinium spp. and graminoids, are not present, or have a limited occurrence (e.g., Figure 6). Historically managed forests created these conditions; wood was harvested without large clear-cuts and livestock grazing counteracted the expansion of the potential understory dominants [34]. Both species suffer in modern production forests, which are darker and denser. The suggested interpretation is that the present occurrences of both Chimaphila and Moneses are legacies of historical use of forests, the most important being grazing by livestock.
However, there is a difference in how these legacies are manifested. In comparison with Chimaphila, the life span of both ramets and genets in Moneses is shorter; its growth form with rosettes close to the ground and clonal spread by horizontal roots make it more sensitive to expansion of competitive dominants in the understory. This makes adult Moneses populations less capable of persisting under deteriorating conditions. On the other hand, if seeds are deposited at a suitable site, Moneses may be more capable at recruiting new genets than Chimaphila, due to the higher germination rate and growth capacity of its subterranean juveniles.
The current occurrence of Chimaphila shows all signs of being composed mainly of remnant populations (Figure 2 and Figure 5). While still being present at many locations, most of these populations are in state of slow decline. When one encounters Chimaphila in the field, it is likely that the ecological conditions necessary for persistence of this species are no longer existing at that particular site. In contrast, Moneses is more sensitive to deteriorating local conditions, and show signs of responding more rapidly. Moneses appears more like a “fugitive” species with transient occurrences, exploiting sites (at least in any significant numbers) which still maintain ecological conditions similar to those which historically prevailed at the time when livestock grazed the forests. However, if the niche requirements for adults are expressed at only few sites in modern production forests, the likelihood that Moneses manage to disperse to and recruit at these sites is extremely small. As also several of the large populations from the earlier records (2015 onwards) were gone in 2022, as indicated by the re-survey data (Appendix A, Table A2), Moneses runs the risk of complete disappearance, and at a faster rate than Chimaphila.
7. Discussion
The main conclusions from this review are the following: (i) There are numerous floristic legacies of historical land use in the understory of boreo-nemoral forests, and these legacies make up a considerable fraction of the current understory plant species richness. (ii) The species considered are both those which have their core occurrence in open grassland systems and species which are regarded as true forest species, but which were favored by historical land use. (iii) Many of the species that occur as legacies have remnant populations sensu [59], but there are also species whose occurrence reflects the existence of ecological conditions resembling the historical forest structure, although the agents causing these ecological conditions are no longer acting. (iv) There are cases where local-scale species richness, taken as a whole, is a legacy of historical activities in forests. (v) Two of the forest species appearing as floristic legacies are Chimaphila umbellata and Moneses uniflora. Both species are declining, but due to differences in their life histories, it is expected that Chimaphila will maintain its populations longer despite deteriorating conditions, while Moneses is more dependent on local conditions resembling what was prevalent historically.
In light of the history of Swedish boreo-nemoral forests, briefly summarized in the Introduction, these legacies reflect that forests for not so long time ago (less than a century) were used very differently from today. Smallholder farmers were still relatively common in forest landscapes until the post-World War II rationalization of Swedish agriculture [131]. Charcoal production occurred to some extent until the 1950s [37]. Livestock grazing in forests were still occurring regionally at least until the 1970s [34]. Although forestry with plantations and clear-cutting practices started to become used by large forest owners (state and private) in northern Sweden during the first part of the 19th century [132], selective cutting was not fully replaced with clear-cutting before the mid-1900s [133]. In southern Sweden, where more forest was privately owned by farmers, this shift may not have been fully implemented until the 1970s [134]. Thus, the conditions characteristic of boreo-nemoral forests historically were quite common until no more than 50–60 years ago. Especially on less productive land, these features take time to vanish, even if the previous management practices have ceased to exist. This is the general explanation for the floristic legacies presented in this review.
For the grassland plants appearing in forests as legacies, the most reasonable explanation is that a part of the former species-pool of the managed grasslands remain at the same location, or in its vicinity, as remnant populations. As forest grazing by livestock covered the largest area of historical forest management regimes [34], this stands out as a dominating factor. The effect of forest grazing was that open-habitat species occurring in the surrounding landscape expanded into forested areas, where they to some extent remain until today [34], i.e., the group of species listed in Table 1. Additionally, for the true forest species, conditions caused by livestock grazing stands out as the dominating explanation behind these species’ presence as legacies. Forest grazing has been suggested to increase the local plant species richness in forests [34,135,136] and one contemporary study from Finland found such a relationship [137]. However, as mentioned previously, the effects on the flora of livestock grazing in boreal and boreo-nemoral forests have not been much studied. Considering the plethora of studies of grazing effects in various grasslands [18], this lack of research may seem puzzling.
The reason why many forest species were favored by forest grazing by livestock is that such forests were lighter (semi-open canopy), with small-scale disturbances, and less dominance of Vaccinium shrubs and graminoids. Similar effects may derive from intentional burning to improve grazing conditions [25,100]. These conditions are expected to allow small-statured, less competitive species to thrive. One may tentatively suggest a guild of forest understory species whose current occurrences at least regionally may be legacies of forest grazing, for example including Lycopodium spp., Diphasiastrum complanatum, Pyrola spp., Linnaea borealis, Maianthemum bifolium, Pulsatilla vernalis, the orchids Goodyera repens and Cephalanthera rubra, and the two species in the case study, Chimaphila umbellata and Moneses uniflora. Further studies are however needed to examine the extent to which these species (and others) should be regarded as legacies of historical livestock grazing in boreo-nemoral forests.
The case study focusing on the two latter species was motivated by the lack of such studies. At least in the provinces examined, Södermanland and Uppland, the results suggest that both species appear as legacies. Both species are in a process of ongoing decline, and for Chimaphila the majority of sites investigated harbor remnant populations. For Moneses, the interpretation was that declining populations (i.e., where the local ecological parameter space is partly outside this species’ fundamental niche) disappear quite rapidly. The more limited clonal ability of Moneses and the short life span of ramets do not promote survival of declining adult genets for long under deteriorating conditions. It may seem unlikely that dispersed seeds of Moneses are able to find suitable sites for establishing new populations to compensate for genet mortality. Instead, maintenance of local populations of Moneses depends on ecological conditions resembling those which were common in historical forests, semi-openness and a sparse cover of Vaccinium spp. and graminoids, and a ground dominated by mosses.
It may also be that the mycorrhizal fungi associated with Chimaphila and Moneses have declined. The diversity of ectomycorrhizal fungi has been found to decrease with increasing nitrogen load [138,139], and several of the mycorrhizal fungi most important to Chimaphila and Moneses depend on decaying logs, for example Tylospora spp. (especially important for Moneses), Piloderma spp., and Russula spp. [140,141]. As the amount of dead wood is reduced in production forests [142], this also indirectly affects Chimaphila and Moneses negatively. One can speculate that similar mechanisms operate in spore forming plants with non-photosynthetic subterranean and mycorrhizal gametophytes, e.g., Lycopodium clavatum [143], which in addition to ectomycorrizal fungi associate with Glomerian AM fungi [144]. AM fungi are typically considered not to be common in boreal forests, e.g., [145,146], but a considerable diversity of AM fungi was discovered in boreo-nemoral forests with a species-rich understory of herbs [147], probably associated with these forest’s former use as wooded meadows. Thus, the historical use may have opened niche space for AM fungi in boreal and boreo-nemoral forests, thus favoring plants such as L. clavatum dependent on these fungi.
If the main conclusions of this review are correct (there are lots of floristic legacies in understory of present-day boreo-nemoral forest), what are their implications? Is it important, and if so, for what?
These legacies will ultimately disappear under current forest management regimes, even though it may take a long time. It is easy to argue that floristic legacies of historical land use are interesting in their own right, as biological cultural heritage [17,51], and also just because one of the basic aims of ecology is to understand the mechanisms behind the occurrence and distribution of species. For forest ecosystems, historical aspects have often been neglected, at least in the context of conservation biology [17,19]. If nothing else, this paper’s main conclusion implies that the historical use of boreo-nemoral forest is a necessary component of any study aiming to understand and explain the current biodiversity in these forests. A focus only on contemporary ecological conditions will not be sufficient. Much of the focus in forest conservation revolves around preserving untouched forests in nature reserves and National Parks. However, if the history of human use of forests is neglected, this may lead to unexpected decline of many forest species even if the forest are protected from clear-cutting and logging [34].
Many of the legacies are species that have their core occurrence in ecological systems which are not forest, for example open grassland systems. They can be considered as being left behind when management changed during the last 50–100 years. Vellend [148] pointed out that biodiversity research often has an unfortunate value-driven bias, which for forest conservation biology may imply that species that are not seen as belonging to the natural forest ecosystems are overlooked. Obviously, such a value-driven bias would lead to very misleading conclusions on current patterns of forest biodiversity.
For true forest species, such as the ones in the case study, Chimaphila umbellata and Moneses uniflora, insights on their dependence on historical use of forests comes out as more essential, i.e., if we strive at preserving these species for the future. In contrast to species which have the core abundance elsewhere, these species will not survive unless the ecological conditions of the forests permit them to do so. This is a key question for conservation biology in forest ecosystems. One currently much discussed method is retention forestry, where parts of forest stands are left unlogged [149]. Overall, the focus of retention forestry has not been on forest understory plants whose occurrence reflect historical land use in forest, e.g., [150]. Some studies suggest that the positive effects of retention forestry on maintaining understory richness is quite limited, e.g., [151], whereas other studies suggest that this method is generally positive for maintaining species diversity, also of understory plants, including open-habitat species [152]. For example, stands of the large shrub Corylus avellana L. are commonly encountered in forests which were previously used a wooded meadow, and these stands have been found to associate with a high understory species richness [153]. Retaining such stands would thus have positive effects also for open-habitat understory plants.
Other actions would be to identify and preserve forest stands which still maintain features of former forests, i.e., a relatively open canopy of trees with variable ages, and introducing livestock grazing [34]. Unfortunately, this is probably not easily compatible with commercial forestry, at least as long as production forestry focuses on wood biomass rather than wood quality.
A final remark is that if conditions improve (from the perspective of the species), the existence of floristic legacies may function as foci for expansion and colonization of new sites by the species, e.g., [102]. Thus, knowledge of the historical background behind present-day biodiversity patterns may be important for future biodiversity, for example in the context of climate warming, e.g., [53]. Research on floristic legacies of historical land use in boreo-nemoral forests are thus not only providing examples of biological cultural heritage and, for some species, knowledge useful for conservation, it may also turn out as relevant for understanding the future of biodiversity in these forests.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data for the review are from the original publications which can be found in the reference list, or presented in Appendix A and Appendix B.
Acknowledgments
I am grateful to L. Glav Lundin for field assistance and to A. Lundell for access to survey data.
Conflicts of Interest
The author declares no conflict of interest.
Appendix A
Table A1.
Results from a re-survey conducted in 2021 of Chimaphila umbellata populations recorded in 2013 [114]. In 2013, 38 sites were investigated, among which Chimaphila was found at 34 sites. These 34 sites were re-surveyed in July 2021. The exact location and identity of four of these sites were however uncertain and they were therefore excluded. Of the 30 remaining sites, Chimaphila was not found at eight sites, and the populations were interpreted as extinct. The survey-data is listed below, divided into those sites where the population had increased (Site 1–6), decreased (Site 7–22), and was extinct (Site 23–30). The average number of ramets in 2013 in still persistent populations (Site 1–22) was 313 (range 7–1776), and in extinct populations (Site 23–30) it was 75 (range 1–314) (Z = 1.99, p = 0.0466; Mann–Whitney U-test). ‘Change’ is the ratio #ramets 2021/#ramets 2013.
Table A1.
Results from a re-survey conducted in 2021 of Chimaphila umbellata populations recorded in 2013 [114]. In 2013, 38 sites were investigated, among which Chimaphila was found at 34 sites. These 34 sites were re-surveyed in July 2021. The exact location and identity of four of these sites were however uncertain and they were therefore excluded. Of the 30 remaining sites, Chimaphila was not found at eight sites, and the populations were interpreted as extinct. The survey-data is listed below, divided into those sites where the population had increased (Site 1–6), decreased (Site 7–22), and was extinct (Site 23–30). The average number of ramets in 2013 in still persistent populations (Site 1–22) was 313 (range 7–1776), and in extinct populations (Site 23–30) it was 75 (range 1–314) (Z = 1.99, p = 0.0466; Mann–Whitney U-test). ‘Change’ is the ratio #ramets 2021/#ramets 2013.
| Populations Persisting 2021 | ||||||
| Site | Name | Coordinates WGS84 | #Ramets 2013 | #Ramets 2021 | Change | |
| Lat | Long | |||||
| Increasing populations | ||||||
| 1 | Bollmora | 59.239331 | 18.235026 | 7 | 10 | 1.43 |
| 2 | Munsö Ekerö | 59.414600 | 17.584391 | 339 | 561 | 1.65 |
| 3 | Snöbergen | 59.544097 | 17.968348 | 13 | 15 | 1.15 |
| 4 | Lapp | 60.115844 | 18.659704 | 51 | 133 | 2.61 |
| 5 | Billudden | 60.637188 | 17.461401 | 1208 | 3578 | 2.96 |
| 6 | Notören | 60.567220 | 17.441189 | 437 | 788 | 1.80 |
| Average change | 1.93 | |||||
| Decreasing populations | ||||||
| 7 | Tullgarn | 58.978209 | 17.601575 | 1776 | 592 | 0.33 |
| 8 | Svarvartorp Ekerö | 59.303610 | 17.701306 | 96 | 11 | 0.11 |
| 9 | Södergården | 59.184542 | 18.366396 | 98 | 75 | 0.76 |
| 10 | Lanan | 59.178118 | 18.323817 | 147 | 64 | 0.44 |
| 11 | Lilla Gräskärret 1 | 59.676369 | 17.558602 | 900 | 647 | 0.72 |
| 12 | Lilla Gräskärret 2 | 59.677099 | 17.557752 | 218 | 48 | 0.22 |
| 13 | Bålstaåsen | 59.575964 | 17.521484 | 140 | 133 | 0.95 |
| 14 | Rosersberg | 59.580045 | 17.892885 | 45 | 4 | 0.09 |
| 15 | Tilskogen | 59.609193 | 17.751514 | 80 | 48 | 0.60 |
| 16 | Österändan | 59.834283 | 18.817657 | 18 | 10 | 0.55 |
| 17 | Långängen | 60.121074 | 18.628193 | 133 | 74 | 0.56 |
| 18 | Hummelfjärden | 60.333915 | 18.422524 | 60 | 21 | 0.35 |
| 19 | Vendelheden | 60.158122 | 17.510899 | 23 | 15 | 0.65 |
| 20 | Billudden/Udden | 60.657148 | 17.508934 | 515 | 286 | 0.56 |
| 21 | Kapplasse | 60.577296 | 17.825108 | 557 | 341 | 0.61 |
| 22 | Slada 1 | 60.532624 | 18.017672 | 20 | 6 | 0.30 |
| Average change | 0.48 | |||||
| Populations extinct 2021 | ||||||
| Site | Name | Coordinates WGS84 | #Ramets 2013 | |||
| Lat | Long | |||||
| 24 | Åva träsk | 59.162262 | 18.342956 | 5 | ||
| 25 | Ekillabadet | 59.606646 | 17.508787 | 4 | ||
| 26 | Hebbo | 59.515183 | 17.533661 | 314 | ||
| 27 | Jomale telemast | 60.497423 | 18.379444 | 19 | ||
| 28 | Vendelheden 2 | 60.157306 | 17.508011 | 25 | ||
| 29 | Slada 2 | 60.534323 | 18.013214 | 125 | ||
| 30 | Skånsta | 59.491324 | 18.324522 | 106 | ||
Table A2.
Results from a re-survey conducted in 2022 of Moneses uniflora populations.
| Site | Name | Year | Coordinates WGS84 | Earlier Records | Re-Survey 2022 | Change | ||
|---|---|---|---|---|---|---|---|---|
| Lat | Long | #P | #FR/R | #FR/VR | ||||
| 1 | Idbäcken | 2015 | 60.595086 | 17.591258 | 2 | 700 R | 301 FR + 996 VR | Increasing |
| 2 | Billudden | 2017 | 60.640076 | 17.465936 | 1 | 50 FR | 355 FR + 750 VR | Increasing |
| 3 | Stigenberg | 2015 | 60.590743 | 17.477950 | 3 | 1340 R | 50 FR + 164 VR | Decreasing |
| 4 | Pärlmossen | 2015 | 60.500548 | 17.473303 | 1 | 450 R | 15 FR + 37 VR | Decreasing |
| 5 | Skirkällan | 2018 | 59.311823 | 16.671995 | 5 | 477 R | 0 | Extinct |
| 6 | Saldalen | 2015 | 59.155962 | 16.363755 | 8 | 59 FR | 0 | Extinct * |
| 7 | Strandstuviken 1 | 2016 | 58.719304 | 17.081295 | 1 | >10 FR | 0 | Extinct |
| 8 | Strandstuviken 2 | 2019 | 58.719269 | 17.081241 | 2 | 12 FR | 0 | Extinct |
| 9 | Örstigsnäs | 2016 | 58.727422 | 17.091121 | 1 | 5 R | 0 | Extinct |
| 10 | Dyvikskärret | 2016 | 58.935340 | 17.585522 | 2 | 19 FR | 0 | Extinct |
| 11 | Åtorpsmossen | 2016 | 58.978506 | 17.589190 | 1 | 150 R | 0 | Extinct * |
| 12 | Tullgarnskogen | 2016 | 58.976070 | 17.568387 | 1 | 2 R | 0 | Extinct |
| 13 | Svartbäcken | 2017 | 59.164998 | 18.204130 | 1 | 1 FR | 0 | Extinct |
| 14 | Tistelkullen | 2015 | 59.745384 | 18.335471 | 1 | 2 FR | 0 | Extinct |
| 15 | Kvarntorpet | 2015 | 59.744616 | 18.335733 | 1 | 5 FR | 0 | Extinct |
| 16 | Ladängssjön | 2015 | 59.860466 | 18.528562 | 1 | 20 FR | 0 | Extinct |
| 17 | Stornotsand | 2018 | 60.190373 | 18.778001 | 1 | 15 FR | 0 | Extinct |
| 18 | Finkarbo | 2015 | 60.523351 | 17.544233 | 2 | 50 FR | 0 | Extinct * |
| 19 | Postmästarhage | 2015 | 60.543124 | 17.474053 | 3 | 200 FR | 0 | Extinct * |
| 20 | Mullbro mossar | 2016 | 60.543450 | 17.476038 | 1 | 200 FR | 0 | Extinct |
| 21 | Mararna | 2015 | 60.618052 | 17.598409 | 8 | 908 R | 0 | Extinct |
| 22 | Stadsskogen | 2015 | 59.838504 | 17.622772 | 2 | 2 FR | 0 | Extinct |
| 23 | Byholma | 2014 | 60.115404 | 18.809443 | 1 | 11 FR | 0 | Extinct |
| 24 | Backby | 2019 | 60.178772 | 18.770642 | 1 | 2 FR | 0 | Extinct |
| 25 | Boda | 2017 | 60.194603 | 18.735252 | 1 | 5 FR | 0 | Extinct |
The records extracted from ‘Artportalen’ (www.artportalen.se, accessed on 5 March 2022) were from 2015 onwards, from the provinces of Södermanland and Uppland. There were many different rapporteurs. The data reported included inconsistencies, since the figure reported for ‘size of the population’ (number of ramets) may refer either to the number of flowering ramets (which is what most easily can be recorded in the field), or total number of ramets, including also vegetative ramets.
In the table below, sites with several patches have been clustered because it was difficult to repeat the initial survey’s delimitation of patches (#P denotes the number of patches in the initial survey). Coordinates refer to a mid-point among patches. #ramets denote either the number of flowering ramets (FR), vegetative ramets (VR), or ramets irrespective of whether vegetative and flowering ramets were distinguished (R). The re-survey recorded the total number of flowering ramets (FR) and the total number of vegetative ramets (VR).
From the province of Södermanland, there were 41 records (in 10 of these, Moneses was just ‘noted’, i.e., no population counts). The records were distributed among 18 sites (some included several patches, and some were visited several years). The number of records for each year was 16 (2015), 9 (2016), 5 (2017), 5 (2018), 3 (2019) and 3 (2020). Nine sites were re-surveyed in June 2022 (Site 5–13).
From the province of Uppland, there were 92 records (in 26 of these, Moneses was just ‘noted’, i.e., no population counts). The records were distributed among 61 sites (some included several patches, and some were visited several years). The number of records for each year was 58 (2015), 9 (2016), 13 (2017), 5 (2018), 4 (2019), 2 (2020), and 1 (2021). A total of 13 sites were re-surveyed in June 2022 (Site 1–4 and 14–22). In addition, three sites known to the author but not recorded in Artportalen were re-surveyed in June 2022 (Site 23–25).
‘Change’ is the interpretation of the fate of the population. An asterisk indicates that the site was a clear-cut in 2022.
Appendix B
- B1.
- The age of flowering in ramets of Moneses uniflora.
Data from Moneses uniflora populations recorded in 2022 was used to make a crude estimate of the age of flowering for Moneses ramets. Note that “age 1” refers to the first year of life, “age 2” to second year of life, etc. The number of flowering and vegetative ramets was counted at sites where Moneses occurred in 2022. The fraction of flowering ramets was 0.270 (based on 2668 ramets).
Under the simplified assumption that genets are in a steady state condition, i.e., new ramets exactly replace the ones that flower and then die, and there is no ramet mortality before flowering (after which the ramet dies), the age of flowering becomes 1/0.270 = 3.7 years (i.e., they flower during their third-fourth year).
This assumption is however not likely to be valid. Genets may be in an expanding phase (relatively more vegetative ramets) or a retarding phase (relatively fewer ramets). Furthermore, it is most likely that some ramets die before reaching the flowering stage. Since no demographic study has been conducted, we can only assume a mortality rate. For example, if 20% of all ramets die before reaching the stage of flowering, the age of flowering becomes 0.8/0.270 = 3.0 years. If 30% of ramets die before reaching the stage of flowering, the age of flowering becomes 0.7/0.270 = 2.6 years.
Acknowledging that the estimate is uncertain, these rough calculations suggest that Moneses ramets typically flower during their third year (±one year), i.e., in line the suggestion by Warming [122].
References
- Josefsson, T.; Gunnarson, B.; Liedgren, L.; Bergman, I.; Östlund, L. Historical human influence on forest composition and structure in boreal Fennoscandia. Can. J. For. Res. 2010, 40, 872–884. [Google Scholar] [CrossRef]
- Iles, L. The role of metallurgy in transforming global forests. J. Archaeol. Method Th. 2016, 23, 1219–1241. [Google Scholar] [CrossRef]
- Jamrichová, E.; Hédl, R.; Kolář, J.; Tóth, P.; Bobek, P.; Hajnalová, M.; Procházka, J.; Kadlec, J.; Szabó, P. Human impact on open temperate woodlands during the middle Holocene in Central Europe. Rev. Palaeobot. Palynol. 2017, 245, 55–68. [Google Scholar] [CrossRef]
- Roberts, P.; Hunt, C.; Arroyo-Kalin, M.; Evans, D.; Boivin, N. The deep human prehistory of global tropical forests and its relevance for modern conservation. Nat. Plants 2017, 3, 17093. [Google Scholar] [CrossRef] [PubMed]
- Dupouey, J.L.; Dambrine, E.; Laffite, D.; Moares, C. Irreversible impact of past land use on forest soils and biodiversity. Ecology 2002, 83, 2978–2984. [Google Scholar] [CrossRef]
- Erickson, C.L. The domesticated landscape of the Bolivian Amazon. In Time and Complexity in Historical Ecology; Balée, W., Erickson, C.L., Eds.; Columbia University Press: New York, NY, USA, 2006; pp. 235–278. [Google Scholar]
- Svensson, E.; Bodin, S.; Hulling, H.; Pettersson, S. The crofter and the iron works: The material culture of structural crisis, identity and making of a living on the edge. Int. J. Hist. Archaeol. 2009, 13, 183–205. [Google Scholar] [CrossRef]
- Carso, J.F.; Watling, J.; Mayle, F.E.; Whitney, B.S.; Iriarte, J.; Prümers, H.; Soto, J.D. Pre-Columbian land use in the ring-ditch region of the Bolivian Amazon. Holocene 2015, 25, 1285–1300. [Google Scholar] [CrossRef] [Green Version]
- Bellemare, J.; Motzkin, G.; Foster, D.R. Legacies of the agricultural past in the forested present: An assessment of historical land-use effect on rich mesic forests. J. Biogeogr. 2002, 29, 1401–1420. [Google Scholar] [CrossRef] [Green Version]
- Hermy, M.; Verheyen, K. Legacies of the past in the present-day forest biodiversity: A review of past land-use effects on forest plant species composition and diversity. Ecol. Res. 2007, 22, 361–371. [Google Scholar] [CrossRef]
- Plue, J.; Hermy, M.; Verheyen, K.; Thuillier, P.; Saguez, R.; Decocq, G. Persistent changes in forest vegetation and seed bank 1600 years after human occupation. Landsc. Ecol. 2008, 23, 673–688. [Google Scholar] [CrossRef]
- Bodin, S.C.; Molino, J.-F.; Odonne, G.; Bremond, L. Unraveling pre-Columbian occupation patterns in the tropical forests of French Guiana using an anthracological approach. Veg. Hist. Archaeobot. 2020, 29, 567–580. [Google Scholar] [CrossRef]
- Sjörs, H. The background: Geology, climate and zonation. In Swedish Plant Geography; Rydin, H., Snoeijs, P., Diekmann, M., Eds.; Acta Phytogeographica Suecica: Uppsala, Sweden, 1999; pp. 5–14. [Google Scholar]
- Östlund, L.; Zackrisson, O.; Axelsson, A.-L. The history and transformation of a Scandinavian boreal forest landscape since the 19th century. Can. J. For. Res. 1997, 27, 1198–1206. [Google Scholar] [CrossRef]
- Lindbladh, M.; Bradshaw, R. The origin of present forest composition and pattern in southern Sweden. J. Biogeogr. 1998, 25, 463–477. [Google Scholar] [CrossRef]
- Ericsson, S.; Östlund, L.; Axelsson, A.-L. A forest of grazing and logging: Deforestation and reforestation history of a boreal landscape in central Sweden. New For. 2000, 19, 227–240. [Google Scholar] [CrossRef]
- Eriksson, O. What is biological cultural heritage and why should we care about it? An example from Swedish rural landscapes and forests. Nat. Conserv. 2018, 28, 1–32. [Google Scholar] [CrossRef]
- Eriksson, O.; Cousins, S.A.O. Historical landscape perspectives on grasslands in Sweden and the Baltic region. Land 2014, 3, 300–321. [Google Scholar] [CrossRef]
- Agnoletti, M.; Santoro, A. Cultural values and sustainable forest management: The case of Europe. J. For. Res. 2015, 20, 438–444. [Google Scholar] [CrossRef]
- Berglund, B.E. The Cultural Landscape during 6000 years in Southern Sweden. Ecol. Bull. 1991, 41, 1–495. [Google Scholar]
- Myrdal, J.; Morell, M. (Eds.) The Agrarian History of Sweden. From 4000 BC to AD 2000; Nordic Academic Press: Lund, Sweden, 2011. [Google Scholar]
- Lagerås, P. The Ecology of Expansion and Abandonment: Medieval and Post-Medieval Land-Use and Settlement Dynamics in a Landscape Perspective; Swedish National Heritage Board: Stockholm, Sweden, 2007. [Google Scholar]
- Emanuelsson, M.; Segerström, U. Medieval slash-and-burn cultivation: Strategic or adapted land use in the Swedish mining district? Environ. Hist. 2002, 8, 173–196. [Google Scholar] [CrossRef]
- Vestbö-Franzén, Å. Farming by fire in north-eastern Småland. Bebygg. Tidskr. 2019, 77, 8–21. [Google Scholar]
- Granström, A.; Niklasson, M. Potentials and limitations for human control over historic fire regimes in boreal forest. Philos. Trans. R. Soc. B 2008, 383, 2353–2358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slotte, H. Harvesting of leaf-hay shaped the Swedish landscape. Landsc. Ecol. 2001, 16, 691–702. [Google Scholar] [CrossRef]
- Hæggström, C.-A. Vegetation and soil of the wooded meadows in Nåtö, Åland. Acta Bot. Fenn. 1983, 120, 1–66. [Google Scholar]
- Eriksson, O. Origin and development of managed meadows in Sweden. A review. Rural Landsc. Soc. Environ. Hist. 2020, 7, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Segerström, U.; Emanuelsson, M. Extensive forest grazing and hay-making on mires: Vegetation changes in south-central Sweden due to land use since the Medieval times. Veg. Hist. Archaeobot. 2002, 11, 181–190. [Google Scholar] [CrossRef]
- Elveland, J. Wet hay-meadows in northern Sweden—Their history and present status. Svensk Bot. Tidskr. 2015, 109, 292–336. [Google Scholar]
- Dahlström, A. Pastures, Livestock Number and Grazing Pressure 1620–1850. Ecological Aspects of Grazing History in South-Central Sweden. Ph.D. Thesis, Swedish University of Agricultural Sciences, Uppsala, Sweden, 2006. (In Swedish with English Summary). [Google Scholar]
- Dahlström, A.; Cousins, S.A.O.; Eriksson, O. The history (1620–2003) of land use, people and livestock, and the relationship to present plant species diversity in a rural landscape in Sweden. Environ. Hist. 2006, 12, 191–212. [Google Scholar] [CrossRef]
- Kardell, Ö. Swedish forestry, forest pasture grazing by livestock and game browsing pressure since 1900. Environ. Hist. 2016, 22, 561–587. [Google Scholar] [CrossRef]
- Westin, A.; Lennartsson, T.; Ljung, T. Skogsbeten Och Bondeskogar: Historia, Ekologi, Natur—Och Kulturmiljövård; Centrum för biologisk mångfald: Uppsala, Sweden, 2022. (In Swedish) [Google Scholar]
- Nilsson, S.G. The changing structure and tree composition in the traditionally grazed forests in the parish Stensbrohult. Svensk Bot. Tidskr. 2006, 100, 393–412. [Google Scholar]
- Emanuelsson, U. The Rural Landscape of Europe: How Man Has Shaped European Nature; The Swedish Research Council Formas: Stockholm, Sweden, 2009. [Google Scholar]
- Arpi, G. Den Svenska Järnhanteringens Träkolsförsörjning 1830–1950; Jernkontorets Bergshistoriska Skriftserie N:r 14: Stockholm, Sweden, 1951. (In Swedish) [Google Scholar]
- National Atlas of Sweden. Swedish Mining and Metalworking; Norstedts Förlagsgrupp: Stockholm, Sweden, 2011. [Google Scholar]
- Gadd, C.-J. The agricultural revolution in Sweden, 1700–1870. In The Agrarian History of Sweden. From 4000 BC to AD 2000; Myrdal, J., Morell, M., Eds.; Nordic Academic Press: Lund, Sweden, 2011; pp. 118–164. [Google Scholar]
- Svensson, E.; Amundsen, H.R.; Holm, I.; Hulling, H.; Johansson, A.; Löfgren, J.; Nilsson, P.; Nilsson, S.; Pettersson, S.; Stensby, V. Empowering marginal lifescapes: The heritage of crofters in between the past and the present. Int. J. Herit. Stud. 2018, 24, 17–34. [Google Scholar] [CrossRef] [Green Version]
- Eriksson, O.; Glav Lundin, L. ‘Gooseberry is the only thing left’—A study of declining biological cultural heritage at abandoned crofts in the province of Södermanland, Sweden. Int. J. Herit. Stud. 2020, 26, 1061–1076. [Google Scholar] [CrossRef] [Green Version]
- Eriksson, O.; Arnell, M.; Lindholm, K.-J. Historical ecology of Scandinavian infield systems. Sustainability 2021, 13, 817. [Google Scholar] [CrossRef]
- Wilson, J.B.; Peet, R.K.; Dengler, J.; Pärtel, M. Plant species richness: The world records. J. Veg. Sci. 2012, 23, 796–802. [Google Scholar] [CrossRef]
- Cousins, S.A.O.; Auffret, A.G.; Lindgren, J.; Tränk, L. Regional-scale land-cover change during the 20th century and its consequences for biodiversity. Ambio 2015, 44 (Suppl. S1), 17–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swedish Board of Agriculture. Plan för Odlingslandskapets Biologiska Mångfald 2019. Available online: https://www2.jordbruksverket.se/download/18.36d57baa168c704154d46f04/1549611543321/ra19_1.pdf (accessed on 7 October 2021).
- O’Dwyer, R.; Marquer, L.; Trondman, A.-K.; Jönsson, A.M. Spatially continuous land-cover reconstructions through the Holocene in southern Sweden. Ecosystems 2021, 24, 1450–1467. [Google Scholar] [CrossRef]
- Hedwall, P.-O.; Brunet, J.; Nordin, A.; Berg, J. Changes in the abundance of keystone forest floor species in response to changes of forest structure. J. Veg. Sci. 2013, 24, 296–306. [Google Scholar] [CrossRef]
- Lindbladh, M.; Axelsson, A.-L.; Hultberg, T.; Brunet, J.; Felton, A. From broadleaves to spruce: The borealization of southern Sweden. Scand. J. For. Res. 2014, 29, 686–696. [Google Scholar] [CrossRef]
- Hedwall, P.-O.; Uria-Diez, J.; Brunet, J.; Gustafsson, L.; Axelsson, A.-L.; Strengbom, J. Interactions between local and global drivers determine long-term trends in boreal forest understorey vegetation. Global Ecol. Biogeogr. 2021, 30, 1765–1780. [Google Scholar] [CrossRef]
- Sundberg, S. Boreal plant decline in southern Sweden during the twentieth century. New J. Bot. 2014, 4, 76–84. [Google Scholar] [CrossRef]
- Ljung, T.; Lennartsson, T.; Westin, A. Inventering Av Biologiskt Kulturarv; Riksantikvarienämbetet: Stockholm, Sweden, 2015. [Google Scholar]
- Szabó, P.; Hedl, R. Advancing the integration of history and ecology for conservation. Conserv. Biol. 2011, 25, 680–687. [Google Scholar] [CrossRef]
- Vellend, M.; Brown, C.D.; Kharouba, H.M.; McCune, J.L.; Myers-Smith, I.H. Historical ecology: Using unconventional data sources to test for effects of global environmental change. Am. J. Bot. 2013, 100, 1294–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perring, M.P.; de Frenne, P.; Baeten, L.; Maes, S.L.; Depauw, L.; Blondeel, H.; Carón, M.M.; Verheyen, K. Global environmental change effects on ecosystems: The importance of land-use legacies. Global Change Biol. 2016, 22, 1361–1371. [Google Scholar] [CrossRef] [PubMed]
- Svedjemyr, O. The flora at abandoned crofts in the parish of Malexander, Östergötland, Sweden. Svensk Bot. Tidskr. 1986, 80, 9–15. [Google Scholar]
- Koerner, W.; Dupouey, J.L.; Dambrine, E.; Benoît, M. Influence of past land use on the vegetation and soils of present day forest in the Vosges mountains, France. J. Ecol. 1997, 85, 351–358. [Google Scholar] [CrossRef]
- Dambrine, E.; Dupouey, J.-L.; Laüt, L.; Humbert, L.; Thinon, M.; Beaufils, T.; Richard, H. Present forest biodiversity patterns in France related to former Roman agriculture. Ecology 2007, 88, 1430–1439. [Google Scholar] [CrossRef]
- Criscuoli, I.; Alberti, G.; Baronti, S.; Favilli, F.; Martinez, C.; Calzolari, C.; Pusceddu, E.; Rumpel, C.; Viola, R.; Miglietta, F. Carbon sequestration and fertility after centennial time scale incorporation of charcoal into soil. PLoS ONE 2014, 3, e91114. [Google Scholar] [CrossRef]
- Eriksson, O. Regional dynamics in plants: A review of evidence for remnant, source-sink and metapopulations. Oikos 1996, 77, 248–258. [Google Scholar] [CrossRef]
- Milberg, P.; Bergman, K.-O.; Jonason, D.; Karlsson, J.; Westerberg, L. Land-use history influence the vegetation in coniferous production forests in southern Sweden. For. Ecol. Manag. 2019, 440, 23–30. [Google Scholar] [CrossRef]
- Pulliam, H.R. Sources, sinks, and population regulation. Am. Nat. 1988, 132, 652–661. [Google Scholar] [CrossRef]
- Hanski, I.; Gilpin, M. Metapopulation dynamics: Brief history and conceptual domain. Biol. J. Linn. Soc. 1991, 42, 3–16. [Google Scholar] [CrossRef]
- De Witte, L.C.; Stöcklin, J. Longevity of clonal plants: Why it matters and how to measure it. Ann. Bot. 2010, 106, 859–870. [Google Scholar] [CrossRef] [PubMed]
- Fenner, M.; Thompson, K. The Ecology of Seeds; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Hylander, K.; Ehrlén, J. The mechanisms causing extinction debts. Trends Ecol. Evol. 2013, 28, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Chase, J.M.; Leibold, M.A. Ecological Niches: Linking Classical and Contemporary Approaches; University of Chicago Press: Chicago, IL, USA, 2003. [Google Scholar]
- Soberón, J.; Nakamura, M. Niches and distributional areas: Concepts, methods, and assumptions. Proc. Natl. Acad. Sci. USA 2009, 106, 19644–19650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutchinson, G.E. Concluding remarks. Cold Spring Harb. Sym. 1957, 22, 415–427. [Google Scholar] [CrossRef]
- Sillett, K.B.; Foster, S.A. Ontogenetic niche shifts in two populations of juvenile threespine stickleback, Gasterosteus aculeatus, that differ in pelvic spine morphology. Oikos 2000, 91, 468–476. [Google Scholar] [CrossRef]
- Eriksson, O. Ontogenetic niche shifts and their implications for recruitment in three clonal Vaccinium shrubs: Vaccinium myrtillus, Vaccinium vitis-idaea, and Vaccinium oxycoccos. Can. J. Bot. 2002, 80, 635–641. [Google Scholar] [CrossRef]
- Grubb, P.J. The maintenance of species-richness in plant communities: The importance of the regeneration niche. Biol. Rev. 1977, 52, 107–145. [Google Scholar] [CrossRef]
- Herben, T.; Münzbergová, Z.; Mildén, M.; Ehrlén, J.; Cousins, S.A.O.; Eriksson, O. Long-term spatial dynamics of Succisa pratensis in a changing rural landscape: Linking dynamical modelling with historical maps. J. Ecol. 2006, 94, 131–143. [Google Scholar] [CrossRef]
- Dahlgren, J.; Ehrlén, J. Incorporating environmental change over succession in an integral projection model of population dynamics of a forest herb. Oikos 2011, 120, 1183–1190. [Google Scholar] [CrossRef]
- Lehtilä, K.; Dahlgren, J.; García, M.B.; Leimu, R.; Syrjänen, K.; Ehrlén, J. Forest succession and population viability of grassland plants: Long repayment of extinction debt in Primula veris. Oecologia 2016, 181, 125–135. [Google Scholar] [CrossRef]
- Eriksson, O. Functional roles of remnant populations in communities and ecosystems. Global Ecol. Biogeogr. 2000, 9, 443–449. [Google Scholar] [CrossRef]
- Leuridan, B.; Froeyman, A. On lawfulness in history and historiography. Hist. Theory 2012, 51, 172–192. [Google Scholar] [CrossRef]
- Ekstam, U.; Forshed, N. Om Hävden Upphör: Kärlväxter Som Indikatorarter i Ängs—Och Hagmarker; Naturvårdsverket: Stockholm, Sweden, 1992. (In Swedish) [Google Scholar]
- Jonason, D.; Ibbe, M.; Milberg, P.; Tunér, A.; Westerberg, L.; Bergman, K.-O. Vegetation in clear-cuts depends on previous land use: A century-old grassland legacy. Ecol. Evol. 2014, 4, 4287–4295. [Google Scholar] [CrossRef] [PubMed]
- Palmgren, A. Studier öfver löfängsområdena på Åland. III Statistisk undersökning av floran. Acta Soc. Pro. Fauna Flora Fenn. 1916, 42, 477–633. (In Swedish) [Google Scholar]
- Lindborg, R.; Eriksson, O. Historical landscape connectivity affects present plant species richness. Ecology 2004, 85, 1840–1845. [Google Scholar] [CrossRef]
- Helm, A.; Hanski, I.; Pärtel, M. Slow response of plant species richness to habitat loss and fragmentation. Ecol. Lett. 2006, 9, 72–77. [Google Scholar] [CrossRef]
- Johansson, V.A.; Cousins, S.A.O.; Eriksson, O. Remnant populations and plant functional traits in abandoned semi-natural grasslands. Folia Geobot. 2011, 46, 165–179. [Google Scholar] [CrossRef]
- Lindborg, R.; Cousins, S.A.O.; Eriksson, O. Plant species response to land use change—Campanula rotundifolia, Primula veris and Rhinanthus minor. Ecography 2005, 28, 29–36. [Google Scholar] [CrossRef]
- Dahlström, A.; Borgegård, S.-O.; Rydin, H. The vascular flora after 50 and 90 years of succession in abandoned infields in Kilsbergen, south-central Sweden. Svensk Bot. Tidskr. 1998, 91, 211–226. [Google Scholar]
- Eriksson, O.; Glav Lundin, L. Legacies of historic charcoal production affect the forest flora in a Swedish mining district. Nord. J. Bot. 2021, 39, e003312. [Google Scholar] [CrossRef]
- Angelstam, P.; Andersson, K.; Isacson, M.; Gavrilov, D.V.; Axelsson, R.; Bäckström, M.; Degerman, E.; Elbakidze, M.; Kazakova-Apkarimova, E.Y.; Satrz, L.; et al. Learning about the history of landscape use for the future: Consequences for ecological and social systems in Swedish Bergslagen. Ambio 2013, 42, 146–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindbladh, M. The influence if former land-use on vegetation and biodiversity in the boreo-nemoral zone of Sweden. Ecography 1999, 22, 485–498. [Google Scholar] [CrossRef]
- Eliasson, P.; Nilsson, S.G. You should hate young oaks and young noblemen—The environmental history of oaks in eighteenth- and nineteenth-century Sweden. Environ. Hist. 2002, 7, 659–677. [Google Scholar] [CrossRef]
- Eriksson, S.; Skånes, H.; Hammer, M.; Lönn, M. Current distribution of older and deciduous forests as legacies from historical use patterns in a Swedish boreal landscape (1725–2007). For. Ecol. Manag. 2010, 260, 1095–1103. [Google Scholar] [CrossRef]
- Arnell, M.; Cousins, S.A.O.; Eriksson, O. Does historical land use affect the regional distribution of fleshy-fruited wood plants? PLoS ONE 2019, 14, e0225791. [Google Scholar] [CrossRef] [Green Version]
- Berg, Å.; Gärdenfors, U.; Hallingbäck, T.; Norén, M. Habitat preferences of red-listed fungi and bryophytes in woodland key habitats in southern Sweden—Analyses of data from a national survey. Biodivers. Conserv. 2002, 11, 1479–1503. [Google Scholar] [CrossRef]
- Nilsson, S.G.; Hedin, J.; Niklasson, M. Biodiversity and its assessment in boreal and nemoral forests. Scand. J. For. Res. 2001, 3, 10–26. [Google Scholar] [CrossRef] [Green Version]
- Jansson, G.; Angelstam, P. Threshold levels of habitat composition for the presence of the long-tailed tit (Aegithalos caudatus) in a boreal landscape. Landsc. Ecol. 1999, 14, 283–290. [Google Scholar] [CrossRef]
- Wiktander, U.; Olsson, O.; Nilsson, S.G. Seasonal variation in home-range size, and habitat area requirement of the lesser spotted woodpecker (Dendrocopos minor) in southern Sweden. Biol. Conserv. 2001, 100, 387–395. [Google Scholar] [CrossRef]
- Ranius, T.; Eliasson, P.; Johansson, P. Large-scale occurrence patterns of red-listed lichens and fungi on old oaks are influenced both by current and historical habitat density. Biodivers. Conserv. 2008, 17, 2371–2381. [Google Scholar] [CrossRef] [Green Version]
- Johansson, V.; Snäll, T.; Ranius, T. Estimates of connectivity reveal non-equilibrium epiphytes occurrence patterns almost 180 years after habitat decline. Oecologia 2013, 172, 607–615. [Google Scholar] [CrossRef] [PubMed]
- Sjöbeck, M. Vång och utmark i Skånes skogsbygd. In Sveriges Natur; Svenska Naturskyddsföreningens årsbok; Svenska Naturskyddsföreningen: Stockholm, Sweden, 1966; pp. 149–182. (In Swedish) [Google Scholar]
- Greiser, C.; Hylander, K.; Meineri, E.; Luoto, M.; Ehrlén, J. Climate limitation at the cold edge: Contrasting perspectives from species distribution modelling and a transplant experiment. Ecography 2020, 43, 637–647. [Google Scholar] [CrossRef]
- Brunet, J. Environmental and historical factors limiting the distribution of rare forest grasses in south Sweden. For. Ecol. Manag. 1993, 61, 263–275. [Google Scholar] [CrossRef]
- Ljung, T. Fäbodskogen Som Biologiskt Kulturarv; Centrum för biologisk mångfald: Uppsala, Sweden, 2011. [Google Scholar]
- Sundberg, S. Present status for Cephalanthera rubra and Chimaphila umbellata in Sweden. Svensk Bot. Tidskr. 2017, 111, 90–104. [Google Scholar]
- Kiedrzyński, M.; Zielińska, K.M.; Kiedrzyńska, E.; Rewicz, A. Refugial debate: On small sites according to their function and capacity. Evol. Ecol. 2017, 31, 815–827. [Google Scholar] [CrossRef] [Green Version]
- Sandström, A.; Svensson, B.; Milberg, P. An example of how to build conservation evidence from case studies: Fire and raking to enhance Pulsatilla vernalis populations. J. Nat. Conserv. 2017, 36, 58–64. [Google Scholar] [CrossRef] [Green Version]
- Oinonen, E. Sporal regeneration of ground pine (Lycopodium complanatum L.) in southern Finland in the light of the dimensions and the age of its clones. Acta For. Fenn. 1967, 83, 1–85. [Google Scholar] [CrossRef] [Green Version]
- Jonsell, L. (Ed.) Upplands Flora; SBF-förlaget: Uppsala, Sweden, 2010. (In Swedish) [Google Scholar]
- SLU Artdatabanken. Rödlistade Arter i Sverige 2020; The Swedish Red List, in Swedish with English Summary; SLU: Uppsala, Sweden, 2020. [Google Scholar]
- Lallemand, F.; Puttsepp, Ü.; Lang, M.; Luud, A.; Courty, P.-E.; Palancade, C.; Selosse, M.-A. Mixotrophy in Pyroleae (Ericaceae) from Estonian boreal forest does not vary with light or tissue age. Ann. Bot. 2017, 120, 361–371. [Google Scholar] [CrossRef]
- Merckx, V.S.F.T. (Ed.) Mycoheterotrophy: The Biology of Plants Living on Fungi; Springer: New York, NY, USA, 2013. [Google Scholar]
- Johansson, V.A.; Mikusinska, A.; Ekblad, A.; Eriksson, O. Partial mycoheterotrophy in Pyroleae: Nitrogen and carbon stable isotope signatures during development from seedling to adult. Oecologia 2015, 177, 203–211. [Google Scholar] [CrossRef]
- Hynson, N.A.; Preiss, K.; Gebauer, G.; Bruns, T.D. Isotopic evidence of full and partial myco-heterotrophy in the plant tribe Pyroleae (Ericaceae). New Phytol. 2009, 182, 719–726. [Google Scholar] [CrossRef]
- Eriksson, O.; Kainulainen, K. The evolutionary ecology of dust seeds. Perspect. Plant Ecol. Evol. Syst. 2011, 13, 73–87. [Google Scholar] [CrossRef]
- Johansson, V.A.; Eriksson, O. Recruitment limitation, germination of dust seeds, and early development of underground seedlings in six Pyroleae species. Botany 2013, 91, 17–24. [Google Scholar] [CrossRef]
- Warming, E. Om Jordudløbere; In Danish with English Summary; Det Kgl. Danske Videnskabernes Selskabs Skrifter, Naturvidenskabelig og Mathematisk Afdelning, 8, Række II. 6.: Copenhagen, Denmark, 1918. [Google Scholar]
- Lundell, A.; Cousins, S.A.O.; Eriksson, O. Population size and reproduction in the declining endangered forest plant Chimaphila umbellata in Sweden. Folia Geobot. 2015, 50, 13–23. [Google Scholar] [CrossRef]
- Antos, J.A.; Zobel, D.B.; Fischer, D.G. Belowground morphology and population dynamics of two forest understory herbs of contrasting growth forms. Botany 2021, 99, 569–580. [Google Scholar] [CrossRef]
- Knudsen, J.T.; Olesen, J.M. Buzz-pollination and patterns in sexual traits in north European Pyrolaceae. Am. J. Bot. 1993, 80, 900–913. [Google Scholar] [CrossRef]
- Johansson, V.A.; Müller, G.; Eriksson, O. Dust seed production and dispersal in Swedish Pyroleae species. Nord. J. Bot. 2014, 32, 209–214. [Google Scholar] [CrossRef] [Green Version]
- Johansson, V.A.; Bahram, M.; Tedersoo, L.; Kõljalg, U.; Eriksson, O. Specificity of fungal associations of Pyroleae and Monotropa hypopitys during germination and seedling development. Mol. Ecol. 2017, 26, 2591–2604. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.-W.; Zhou, J.; Peng, H.; Freudenstein, J.V.; Milne, R.I. Relationships between Tertiary relict and circumboreal woodland floras: A case study in Chimaphila (Ericaeae). Ann. Bot. 2019, 123, 1089–1098. [Google Scholar] [CrossRef]
- Massicote, H.B.; Melwille, L.H.; Tackaberry, L.E.; Peterson, R.L. A comparative study of mycorrhizas in several genera of Pyroleae (Ericaceae) from western Canada. Botany 2008, 86, 610–622. [Google Scholar] [CrossRef]
- Warming, E. Om Skudbygning, Overvintring Og Foryngelse; Aftryck af Naturhistorisk Forenings Festskrift. Kgl. Hof-Bogtrykkeri: Kjøbenhavn, Denmark, 1884. (In Danish) [Google Scholar]
- Warming, E. The structure and biology of arctic flowering plants: Ericineae (Ericaceae, Pirolaceae) 1. Morphology and Biology. Arb. Fra Den Bot. Have I København 1908, 43, 1–71. [Google Scholar]
- Hynson, N.A.; Bidartondo, M.I.; Read, D.J. Are there geographic mosaics of mycorrhizal specificity and partial mycoheterotrophy? A case study in Moneses uniflora (Ericaceae). New Phytol. 2015, 208, 1003–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rydberg, H.; Wanntorp, H.-E. Sörmlands Flora; Botaniska Sällskapet: Stockholm, Sweden, 2001. (In Swedish) [Google Scholar]
- Johansson, V.A.; Lundell, A.; Eriksson, O. Ecology of ericaceous mycoheterotrophs in Sweden. Svensk Bot. Tidskr. 2017, 111, 240–263. [Google Scholar]
- Artportalen. Available online: www.artportalen.se (accessed on 15 March 2022).
- Rydberg, H. (Linnaeus University). Personal communication, 2021.
- Thompson, J.N. The Coevolutionary Process; University of Chicago Press: Chicago, IL, USA, 1994. [Google Scholar]
- Leake, J.R.; Cameron, D.D. Physiological ecology of mycoheterotrophy. New Phytol. 2010, 185, 601–605. [Google Scholar] [CrossRef] [PubMed]
- Bidartondo, M.; Bruns, T.D. Extreme specificity in epiparasitic Monotropoideae (Ericaceae): Widespread phylogenetic and geographical structure. Mol. Ecol. 2001, 10, 2285–2295. [Google Scholar] [CrossRef] [Green Version]
- Flygare, I.A.; Isacson, M. The tension between modernity and reality. In The Agrarian History of Sweden. From 4000 BC to AD 2000; Myrdal, J., Morell, M., Eds.; Nordic Academic Press: Lund, Sweden, 2011; pp. 214–256. [Google Scholar]
- Lundmark, H.; Östlund, L.; Josefsson, T. Continuity forest or second-generation forest? Historical aerial photos provide evidence of early clear-cutting in northern Sweden. Silva Fenn. 2021, 51, 10460. [Google Scholar] [CrossRef]
- Lundmark, H.; Josefsson, T.; Östlund, L. The history of clear-cutting in northern Sweden: Driving forces and myths in boreal silviculture. For. Ecol. Manag. 2013, 307, 112–122. [Google Scholar] [CrossRef]
- Östlund, L. (Swedish University of Agricultural Sciences). Personal communication, 2022.
- Steen, E. Effects of Grazing on Swedish Vegetation; In Swedish with English summary; Statens Jordbruksförsök Meddelande nr 89: Uppsala, Sweden, 1958. [Google Scholar]
- Plieninger, T.; Hartel, T.; Martín-López, B.; Beaufoy, G.; Bergmeier, E.; Kirby, K.; Montero, M.J.; Moreno, G.; Oteros-Rozas, E.; van Uytvanck, J. Wood-pastures of Europe: Geographic coverage, social-ecologcial values, conservation management, and policy implications. Biol. Conserv. 2015, 190, 70–79. [Google Scholar] [CrossRef]
- Oldén, A.; Raatikainen, K.J.; Tervonen, K.; Halme, P. Grazing and soil pH are biodiversity drivers of vascular plants and bryophytes in boreal wood-pastures. Agri. Ecosyst. Environ. 2016, 222, 171–184. [Google Scholar] [CrossRef]
- Lilleskov, E.A.; Fahey, T.J.; Horton, T.R.; Lovett, G.M. Belowground ectomycorrhizal fungal community change over a nitrogen deposition gradient in Alaska. Ecology 2002, 83, 104–115. [Google Scholar] [CrossRef]
- Davey, M.L.; Skogen, M.J.; Heegaard, E.; Halvorsen, R.; Kauserud, H.; Ohlson, M. Host and tissue variations overshadow the response of boreal moss-associated fungal communities to increased nitrogen load. Mol. Ecol. 2017, 26, 571–588. [Google Scholar] [CrossRef]
- Rajala, T.; Tuomivirta, T.; Pennanen, T.; Mäkipää, R. Habitat models of wood-inhabiting fungi along a decay gradient of Norway spruce logs. Fungal Ecol. 2015, 18, 48–55. [Google Scholar] [CrossRef]
- Mäkipää, R.; Rajala, T.; Schigel, D.; Rinne, K.T.; Pennanen, T.; Abrego, N.; Ovaskainen, O. Interactions between soil- and dead wood-inhabiting fungal communities during the decay of Norway spruce logs. ISME J. 2017, 11, 1964–1974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jonsson, B.G.; Kruys, N.; Ranius, T. Ecology of species living on dead wood—Lessons from dead wood management. Silva Fenn. 2005, 39, 289–309. [Google Scholar] [CrossRef] [Green Version]
- Whittier, D.P. Red light inhibition of spore germination in Lycopodium clavatum. Am. Fern J. 2008, 98, 194–198. [Google Scholar] [CrossRef]
- Winther, J.L.; Friedman, W.E. Arbuscular mycorrhizal association in Lycopodiaceae. New Phytol. 2008, 177, 790–801. [Google Scholar] [CrossRef]
- Johnson, D.; Ijdo, M.; Genney, D.R.; Anderson, I.C.; Alexander, I.J. How do plants regulate the function, community structure, and diversity of mycorrhizal fungi? J. Exp. Bot. 2005, 56, 1751–1760. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: Oxford, UK, 2008. [Google Scholar]
- Öpik, M.; Moora, M.; Zobel, M.; Saks, Ü.; Wheatley, R.; Wright, F.; Daniell, T. High diversity of arbuscular mycorrhizal fungi in a boreal herb-rich coniferous forest. New Phytol. 2008, 179, 867–876. [Google Scholar] [CrossRef]
- Vellend, M. The behavioral economics of biodiversity conservation scientists. Philos. Top. 2019, 47, 219–237. [Google Scholar] [CrossRef]
- Gustafsson, L.; Baker, S.C.; Bauhus, J.; Beese, W.J.; Brodie, A.; Kouki, J.; Lindenmayer, D.B.; Lõhmus, A.; Martínez Pastur, G.; Messier, C.; et al. Retention forestry to maintain multifunctional forests: A world perspective. BioScience 2012, 62, 633–645. [Google Scholar] [CrossRef] [Green Version]
- Koivula, M.; Vanha-Majamaa, I. Experimental evidence on biodiversity impacts of variable retention forestry, prescribed burning, and deadwood manipulation in Fennoscandia. Ecol. Process. 2020, 9, 11. [Google Scholar] [CrossRef] [Green Version]
- Johnson, S.; Strengbom, J.; Kouki, J. Low levels of tree retention do not mitigate the effects of clearcutting on ground vegetation dynamics. For. Ecol. Manag. 2014, 330, 67–74. [Google Scholar] [CrossRef]
- Fedrowitz, K.; Koricheva, J.; Baker, S.C.; Lindenmayer, D.B.; Palik, B.; Rosenvald, R.; Beese, W.; Franklin, J.F.; Kouki, J.; Macdonald, E.; et al. Can retention forestry help conserve biodiversity? A meta-analysis. J. Appl. Ecol. 2014, 51, 1669–1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koorem, K.; Moora, M. Positive association between understory species richness and a dominant shrub species (Corylus avellana) in a boreonemoral spruce forest. For. Ecol. Manag. 2010, 260, 1407–1413. [Google Scholar] [CrossRef]
Figure 1.
The background of floristic legacies in boreo-nemoral forests. (A) A gate in a stone wall enclosing a former hay-meadow on Singö, province of Uppland, Sweden. Forest grazing was conducted in the foreground of the gate and the stone wall. The site is slowly reverting back to forest. (B) An old storage building for hay in the meadow enclosed by the stone-wall in (A). The grass sward still maintains a rich grassland flora. (Photos: the author, July 2022).
Figure 1.
The background of floristic legacies in boreo-nemoral forests. (A) A gate in a stone wall enclosing a former hay-meadow on Singö, province of Uppland, Sweden. Forest grazing was conducted in the foreground of the gate and the stone wall. The site is slowly reverting back to forest. (B) An old storage building for hay in the meadow enclosed by the stone-wall in (A). The grass sward still maintains a rich grassland flora. (Photos: the author, July 2022).

Figure 2.
A graphic illustration of the difference between source-sink and remnant populations (“source-sink populations in time”). Adapted from Ref. [59].
Figure 2.
A graphic illustration of the difference between source-sink and remnant populations (“source-sink populations in time”). Adapted from Ref. [59].
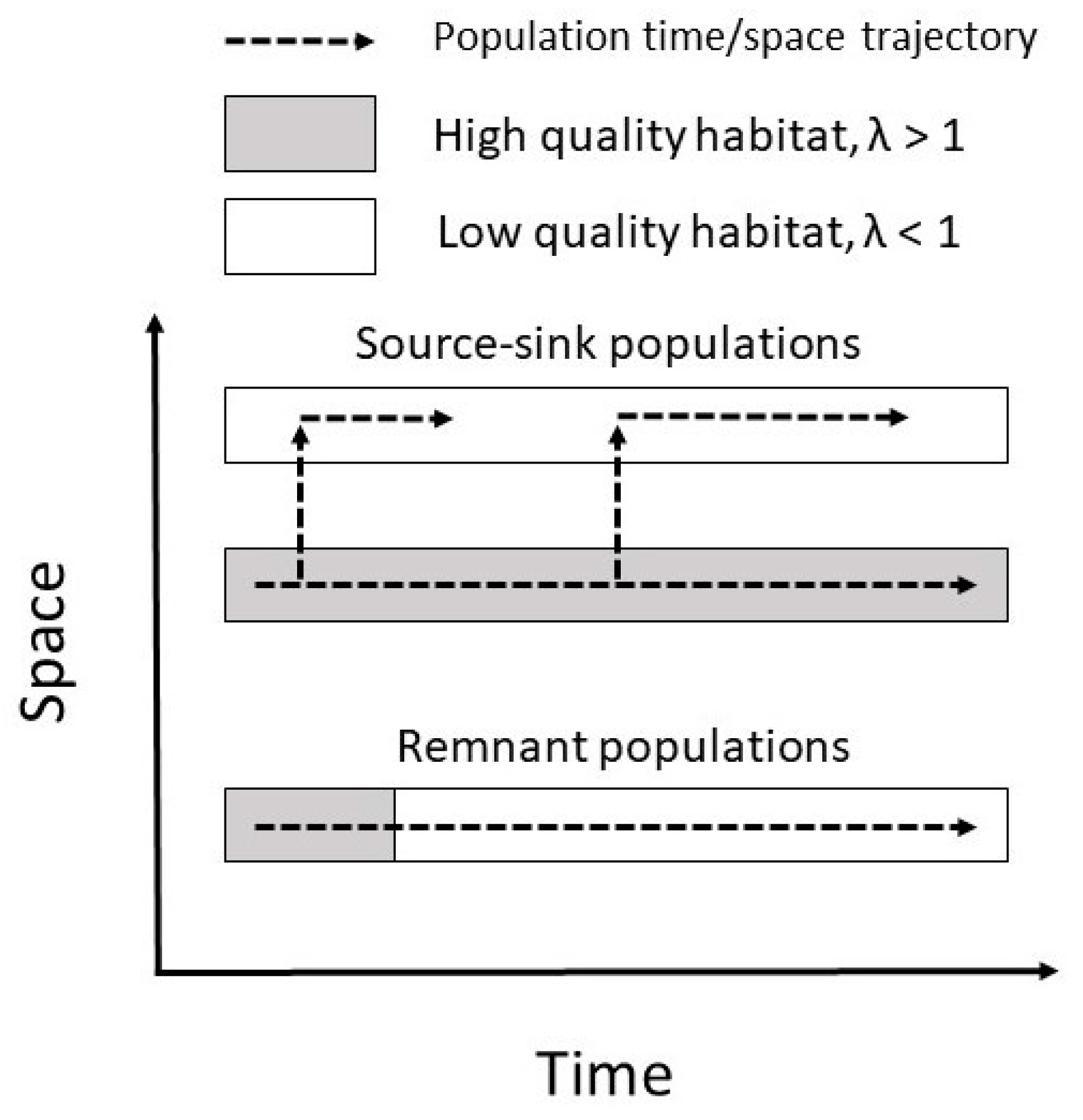
Figure 3.
Chimaphila umbellata, a species favored by historical human use of boreo-nemoral forests, mainly livestock grazing. (Photo: Anna Lundell, July 2013).
Figure 3.
Chimaphila umbellata, a species favored by historical human use of boreo-nemoral forests, mainly livestock grazing. (Photo: Anna Lundell, July 2013).

Figure 4.
Moneses uniflora, a species favored by historical human use of boreo-nemoral forests, mainly livestock grazing. (Photo: the author, June 2022).
Figure 4.
Moneses uniflora, a species favored by historical human use of boreo-nemoral forests, mainly livestock grazing. (Photo: the author, June 2022).

Figure 5.
The fate of Chimaphila umbellata populations based on two sources. Black bars: based on surveys of more than 600 populations from the 1990s onwards and re-surveys on average 10 years later [101]. Grey bars: this study, based on a re-survey in 2021 of 30 populations recorded in 2013 [114].
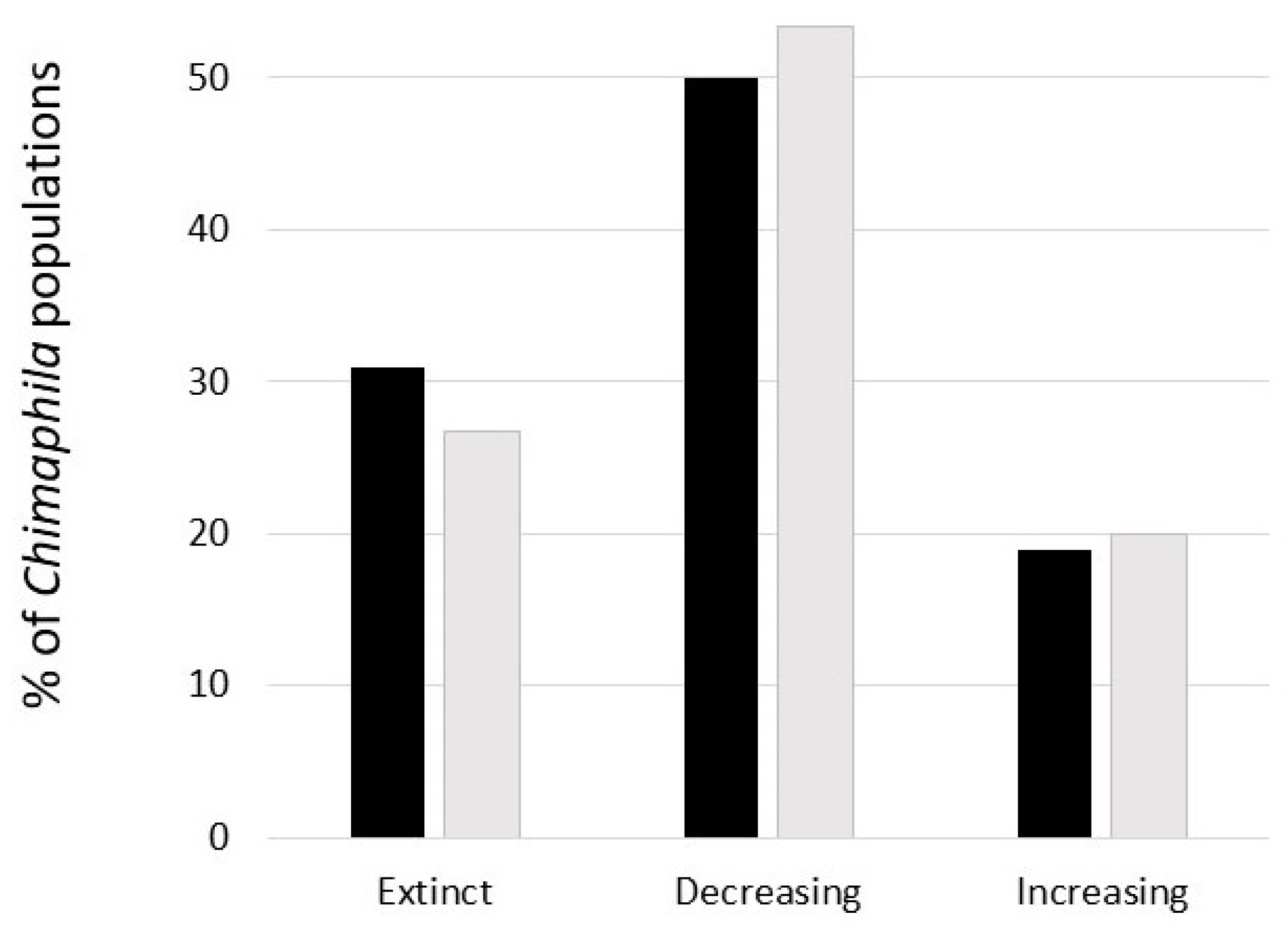
Figure 6.
A forest where Chimaphila umbellata and Moneses uniflora thrive. The site at Billudden, province of Uppland, harboring the largest populations of these species recorded in this study. (Photo: the author, June 2021).
Figure 6.
A forest where Chimaphila umbellata and Moneses uniflora thrive. The site at Billudden, province of Uppland, harboring the largest populations of these species recorded in this study. (Photo: the author, June 2021).

Figure 7.
The fate of Moneses uniflora populations in 2022 in comparison with surveys conducted between 2014 and 2019 (n = 25).
Figure 7.
The fate of Moneses uniflora populations in 2022 in comparison with surveys conducted between 2014 and 2019 (n = 25).
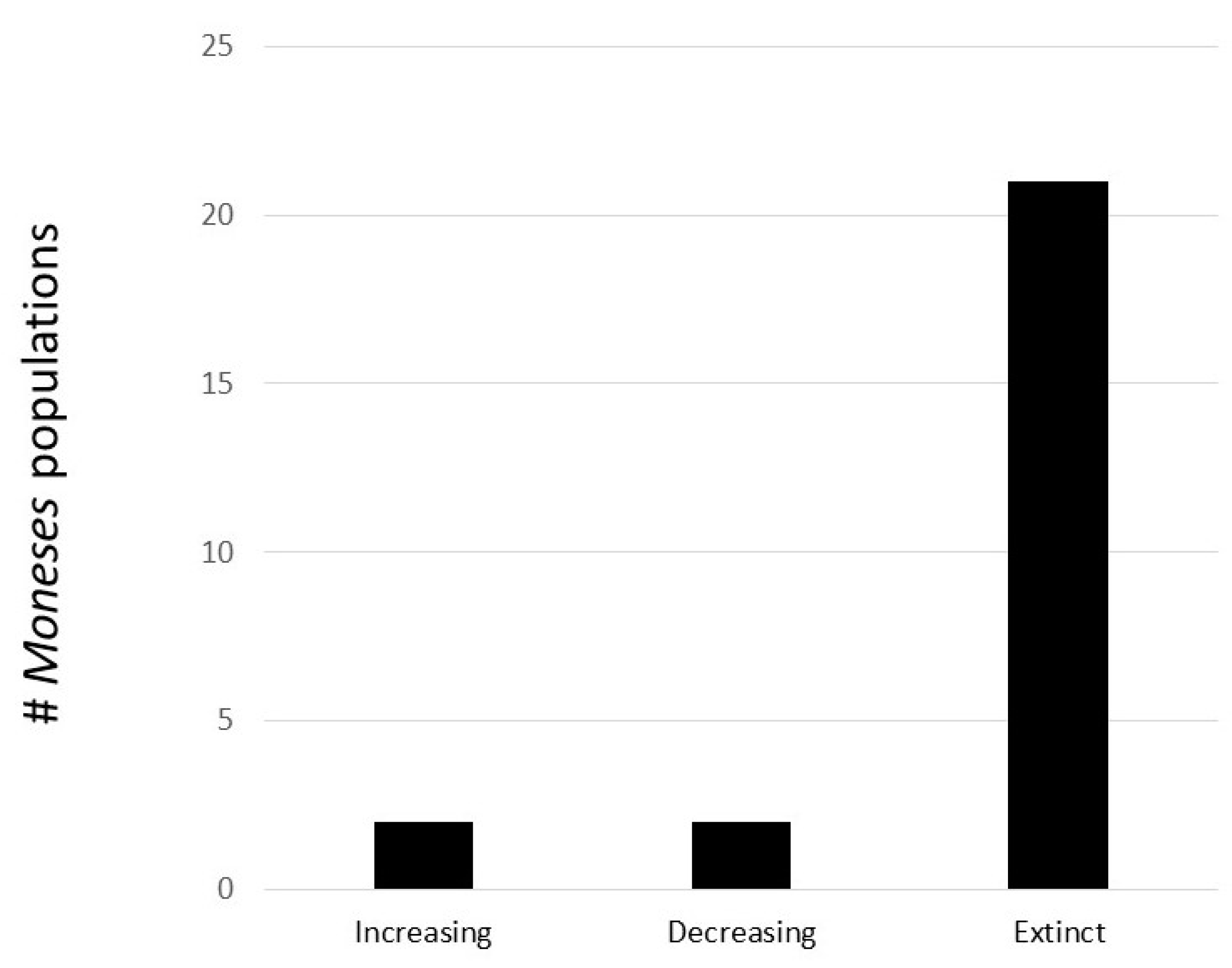
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Eriksson, O. Floristic Legacies of Historical Land Use in Swedish Boreo-Nemoral Forests: A Review of Evidence and a Case Study on Chimaphila umbellata and Moneses uniflora. Forests 2022, 13, 1715. https://doi.org/10.3390/f13101715
AMA Style
Eriksson O. Floristic Legacies of Historical Land Use in Swedish Boreo-Nemoral Forests: A Review of Evidence and a Case Study on Chimaphila umbellata and Moneses uniflora. Forests. 2022; 13(10):1715. https://doi.org/10.3390/f13101715
Chicago/Turabian StyleEriksson, Ove. 2022. "Floristic Legacies of Historical Land Use in Swedish Boreo-Nemoral Forests: A Review of Evidence and a Case Study on Chimaphila umbellata and Moneses uniflora" Forests 13, no. 10: 1715. https://doi.org/10.3390/f13101715
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.