
Tree Species Mixtures Can Improve the Water Storage of the Litter–Soil Continuum in Subtropical Coniferous Plantations in China
Institute for Forest Resources and Environment Research Center of Guizhou Province, Plateau Mountain Forest Cultivation Key Laboratory of Guizhou Province, College of Forestry, Guizhou University, Guiyang 550025, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Forests 2023, 14(2), 431; https://doi.org/10.3390/f14020431
Submission received: 5 January 2023
/
Revised: 15 February 2023
/
Accepted: 16 February 2023
/
Published: 20 February 2023
(This article belongs to the Special Issue Biodiversity-Ecosystem Functioning Relationships in Forest Ecosystems)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Increasing evidence has shown that introducing broadleaved trees into coniferous plantations can regulate hydrologic stores and fluxes; however, the effects and regulatory mechanisms of species mixing on the water conservation capacity of the litter–soil continuum remain poorly understood, and differences among tree species may appear. Herein, we investigated and compared the water conservation capacity of the litter layer (semi-decomposed and decomposed layer) and soil layer (0–100 cm) in a monoculture plantation (Pinus massoniana) and five mixed plantations (Pinus massoniana mixed with Cercidiphyllum japonicum, Manglietia chingii, Camellia oleifera, Michelia maudiae, and Bretschneidera sinensis) and comprehensively considered their potential influencing factors. We discovered that the identity of broadleaved tree species significantly affected the water storage of litter and soil in the mixed plantations (p < 0.05). The effective water-holding capacity of the litter (13.39 t·ha−1) was low due to the coniferous litter’s simple structure and challenging breakdown, despite the fact that the litter stock of the monoculture plantation was substantially larger than that of the mixed plantation (14.72 t·ha−1). Introducing deep-rooted tree species (e.g., Bretschneidera sinensis and Camellia oleifera) into Pinus massoniana farmsteads improved the soil-pore structure and aggregate stability, thereby significantly increasing the 0–100 cm soil water storage. Furthermore, we found that litter storage, soil organic carbon, and litter thickness, as key influencing factors, have complex effects on the water storage of the litter–soil continuum. Generally, these findings demonstrated that mixed plantations can potentially improve the water conservation capacity of the litter–soil system. Nevertheless, special attention should be given to the complementarity between tree species combinations.
1. Introduction
Water resource conservation in forests can help to retain precipitation and regulate runoff [1], affecting hydrological regulation of the ecosystem and water purification [2], which is one of the vital services of terrestrial ecosystems. Climate models usually predict that there will be more drought events and changes in rainfall frequency in subtropical regions by the end of this century [3,4]; therefore, improving the forest water conservation function to cope with global climate change has a far-reaching impact. However, the effect of forest management measures on the water conservation function of subtropical plantations (e.g., through litter and soil) and its potential mechanisms are still unclear [5]. In addition, the prediction results of hydrological models in subtropical plantations are often inconsistent with experimental observations [6], which makes the impact of forest management measures on the hydrological balance (especially hydrological processes below the canopy) very uncertain.
Generally, precipitation not intercepted by the forest canopy and not evaporated back to the atmosphere in the forest moves downwards in the form of penetrating rain and trunk runoff, and finally enters the soil after redistribution by the litter layer [7]. Rainfall interception by the litter and soil layers accounts for approximately 2%–70% of the total rainfall due to the differences in tree species and rainfall conditions [8,9], and is an important source of soil water and groundwater, which indicates that the litter and soil layers have a greater interception and smaller potential evaporation of water than the canopy [10]. Previous studies have predominantly considered the water conservation factors of litter and soil separately [11,12,13,14]; however, as a soil buffer layer, the litter layer can not only lessen the erosion and compaction of soil caused by raindrop impact, thus reducing surface runoff and sedimentation, but can also help to limit soil water evaporation and enhance soil water infiltration [15,16]. In addition, litter decomposition also regulates soil ecological processes [10], including carbon and nutrient cycling, which may change soil physical properties (such as soil mechanical composition and porosity) to affect soil water retention performance [17,18]. However, the role of the litter–soil continuum in hydrological cycle processes, including infiltration, runoff, evapotranspiration, and drainage, has not been fully considered because its water storage characteristics are not clear, which tightly circumscribes our further understanding of forests’ hydrological processes.
The water conservation ability of the litter–soil continuum, as a result of long-term forest floor ecological processes, is affected by many factors, especially plants and soil [17]. Yet, there has been little documentation of the variations in the water conservation ability of the litter–soil continuum and its influencing factors under forest management practices (i.e., forming mixed plantations by replanting broadleaved trees after coniferous plantation thinning). Ecological theory shows that, compared with monoculture plantations, mixed plantations are significantly different in their methods of coping with environmental pressure [19], specifically in the differences in stand structure and the complementarity of resource acquisition and utilization (including soil nutrients and water) [20,21]. This will lead to changes in plant biomass, especially in litter and roots, and ultimately affect the water retention process, infiltration, and consumption in the litter–soil continuum [7]. In addition, interactions between litter and soil could be affected by differences in tree species. For example, tree species determine the speed of litter decomposition and the root system’s vertical distribution (shallow roots or deep roots) [22,23], thus affecting the soil characteristics related to water storage in the soil (e.g., the bulk density, porosity, and soil texture) [23]. Furthermore, tree species vary in their impact on the soil organic matter quality (e.g., the ratio of its hydrophilic and hydrophobic functional groups) and the ways in which they affect water storage in the litter–soil continuum by variations in carbon decomposition and nutrient release from litter [24]. Quantifying the interactions between litter and soil can provide essential insights to understand the changes to the forest water conservation capacity. These changes are caused by the increase in tree species diversity, which will help to formulate forest management practices according to the expected future climate change and reduce the uncertainty of the existing forest hydrological model’s predictions [25].
Pinus massoniana Lamb. (PM), as the main pioneer species for forestation in the subtropical region of China, is widely distributed due to its high economic value (e.g., timber and rosin) and ecological service function (e.g., carbon sequestration and biodiversity protection) [26]. Increasing evidence shows that with the development of single–species PM plantations, ecological problems, such as soil degradation and water and soil loss, have become prominent [27]. The establishment of mixed plantations, as a sustainable forest management measure, can provide more wood production and better water resource protection than monoculture plantations [19], and this depends on the selection of broadleaved tree species. Hence, our specific objectives were to (1) survey the difference in components of water storage in the litter–soil continuum between mixed plantations with different tree species combinations, compared with a monoculture plantation; and (2) find the major factors impacting changes in water storage in the litter–soil continuum and determine the possible interactions between them. This study may help us to further understand the water regulation services of mixed plantations and provide more effective suggestions for improving the water conservation capacity of coniferous plantations in subtropical regions.
2. Materials and Methods
2.1. Site Description
Our research was performed at the Longli National Forest Farm (LNFF) in a subtropical plantation ecosystem (N26.10°, E106.45°, 1150 m above sea level), Guizhou Province, China. Among the sites, the annual mean temperature was 15.0 °C, the mean annual accumulated precipitation was 1229.2 mm, and the mean annual relative humidity was 77%. The main soil type in the study site was Inceptisol, according to the U.S. method of soil taxonomy. This type of soil is acidic and nutrient-poor, and forms in sand shale; the average soil thickness is more than 90 m; the soil gravel content is less than 2%; soil pH levels are from 5.0–5.5; and the soil horizons were delineated as A (0–20 cm), B (20–80 cm), BC (80–100 cm), and C (>100 cm). The primary vegetation in the study site consisted of coniferous plantations and mixed conifer–broadleaf forests.
2.2. Experimental Design
The PM plantation was converted from desolate hills in 1957. In July 2004, part of the PM plantation was selectively cut at a 60% intensity and interplanted with native broadleaf trees; the experimental design was described in detail in a previous study [28]. Six forest types in similar environmental conditions (e.g., geology, soil type, and topography), such as pure PM plantation, PM and Bretschneidera sinensis (BS) mixed forest, PM and Manglietia chingii Dandy (MC) mixed forest, PM and Cercidiphyllum japonicum mixed forest, PM and Camellia oleifera Abel (CO) mixed forest, and PM and Michelia maudiae Dunn (MM) mixed forest, were utilized at the study sites. The pure stand was designated as a control compared to the other mixed stands. A more detailed description of the stand characteristics is given in Table S1.
2.3. Data Collection
For each forest type, three 20 m × 20 m plots with similar canopy structures, slopes, and light penetration were established in November 2019, and were located at least 500 m from one another to avoid spatial autocorrelation. Three duplicate sampling quadrats (0.5 m × 0.5 m) were chosen randomly along the diagonal at each plot. The samples, from the same soil and litter layers of the three duplicate sampling quadrats in each plot, were mixed to form a composite sample. The litter layers, including semi-decomposed litter (OF) and undecomposed litter (OL), were collected separately, and the litter thickness and fresh weight were recorded. The contours of the OF were broken and the color changed, but the shape and color of the OL remained unaltered. In the laboratory, the water-holding ability/capacity and dry litter mass were determined by the indoor immersion method and the drying method, respectively [7]. The wet mass of the litter reached a maximum after soaking for 24 h, and this was considered the maximum water-holding capacity of litter (MWHC) [12]; the effective water-holding capacity of litter (EWHC) was roughly 85% of MWHC in the forest base using the immersion method [29]. The MWHC and EWHC were calculated using the equations:
where W24 is the wet weight of litter dipped in a soaking bath for a day, then removed from it and weighed again after 5 min; W0 is the dry litter mass; and 0.85 is the adjustment coefficient to estimate the EWHC.
Second, undisturbed soil samples were obtained from the 0–100 cm soil layers using a 200 cm3 coring ring in 20 cm increments to determine soil water storage. The bulk density and capillary porosity of these samples were measured based on the volume–mass relationship [30] and using the cylinder soak method [31], respectively. In addition, soil-saturated water storage (SSWS; t·ha−1), capillary water storage (CWS; t·ha−1), and noncapillary water storage (NCWS; t·ha−1) were used as indicators to evaluate the soil water storage [32].
where Pt is the total porosity of the soil (%), Pa is the capillary porosity (%), Pb is the noncapillary porosity (%), and h is the thickness of the soil (m).
Finally, 1 kg of undisturbed soil was obtained by a shovel from each soil layer to determine the soil aggregates and the soil organic carbon (SOC). The potassium dichromate oxidation technique was used to calculate the SOC [33]. The dry sieving method was adopted for the separation of soil aggregates; that is, undisturbed soil was taken from each soil sample and sieved into 5 particle-size groups, including >5, 2–5, 1–2, 0.25–1, and <0.25 mm. The mean weight diameter (MWD; mm), mass fractal dimension (D), and geometric mean diameter (GMD, mm) were used to evaluate the stability of soil aggregates [26]. These parameters were calculated according to the following equations:
where Wi denotes the proportion of each aggregate class to the total (%), Xi denotes the mean diameter of the classes (mm), mi denotes the cumulative mass of the aggregates smaller than Xi, M denotes the total mass of the aggregates, and Xmax denotes the mean diameter of the greatest aggregate class.
2.4. Evaluation Method of the Water Conservation Capacity of the Litter–Soil Continuum
In this study, a coordinated comprehensive evaluation method was adopted based on multidimensional space theory to assess the water conservation capacity of various forest types [34]. Specifically, the evaluation object was regarded as a space point determined by multiple vectors, and the distance between each point was compared with the best point to eliminate the impact of dimensional differences on the evaluation results. We selected MWHC, EWHC, SSWS, CSW, and NCWS as evaluating indicators. First, dimensionless processing was performed by Formula (9) to eliminate the difference between indicators; second, Mj, the maximum value of each index, was used for comparisons to form the relative value Dij, the “matrix coordinate”; thirdly, Formula (10) was used to calculate the distance from the ith processing to the standard point; lastly, Formula (11) was used to find the sum of the distances from each evaluation indicator to the standard point. The specific calculation was performed using the following equations:
where Dij is the evaluation indicator set (i represents different forest types, and j represents different indicators), Sij is the raw dataset, Mj is the best value of the raw dataset, Pij is the distance from the ith evaluation indicator to the standard point, and M is the sum of the distances from each evaluation indicator to the standard point, representing the comprehensive evaluation score. The smaller this score is, the better.
2.5. Statistical Analysis
All statistical analyses in this study were carried out using R software version 4.1.1.
The data were firstly tested by a normal distribution test and a homogeneity of variances test. Then, the significant effects of forest type (PM, PM+BS, PM+MC, PM+CJ, PM+CO, and PM+MM) and litter layer (OL and OF) on MWHC and EWHC were determined using analysis of variance (ANOVA) followed by a least significant difference (LSD) test, as were the significant effects of forest type and soil layer (0–20, 20–40, 40–60, 60–80, and 80–100 cm) on SSWS, CWS, and NCWS. We selected MWHC, EWHC, SSWS, CSW, and NCWS as evaluating indicators in the coordinated comprehensive evaluation method. The correlations between the various environmental characteristics, including litter thickness (LT), litter stock (LS), MWD, GMD, D, and SOC, and the water conservation ability of various forest types were uncovered using redundancy analysis (RDA) and Pearson correlation analysis. The water-conserving potential of the litter–soil continuum was determined by using the random forest (RF) algorithm to identify the relative relevance of various environmental elements.
3. Results
3.1. The Water-Holding Capacity of Litter in Different Forest Types
In comparison to the OL layer, the OF layer had reduced EWHC and MWHC values (Figure 1a,b). In comparison to the other forest types, OF and OL layers’ MWHC and EWHC in PM+CO were lower. The OL layer’s EWHCs in PM, PM+BS, and PM+MM were much greater than that of PM (9.71 t·ha−1), at 14.47 and 17.05 t·ha−1, respectively.
3.2. Soil Water Storage in Different Forest Types
The SSWS and CWS in PM+CO and PM+MM at 0–20, 20–40, and 40–60 cm increased compared with those in PM (Figure 2a,b); the NCWS in PM+MM at 0–20, 20–40, and 40–60 cm was significantly higher than that in the other forest types (Figure 2c), at 148.47, 113.76, and 96.27 t·ha−1, respectively. The SSWS in PM+BS and PM+CJ at 60–80 and 80–100 cm decreased compared with that of PM; however, there was no difference in NCWS.
3.3. Comprehensive Evaluation of Water Conservation Capacity in the Litter–Soil Continuum of Different Forest Types
The contributions of litter and soil to the water conservation ability/capacity of the litter–soil continuum were significantly different among the six plantations (Figure 3). The comprehensive evaluation score of soil in PM+BS, PM+MC, and PM+CJ increased compared with that in PM, while PM+CO and PM+MM decreased. The comprehensive evaluation score of litter only increased in PM+CO compared with that in PM. Overall, the water conservation capacity of the litter–soil continuum in PM+MM, PM+BS, and PM+CJ increased compared with that of PM.
3.4. Effects of Environmental Factors on the Water Conservation Capacity of the Litter–Soil Continuum
The water conservation capacity of the litter–soil continuum between the six forest types, especially those in PM+MM and PM+CO, formed separate clusters (Figure 4a), and the investigated influencing factors explained 49.20% of the total variation. They showed that CWS, NCWS, and SSWS had positive correlations with SOC and negative correlations with D (Figure 4b); LT and LS were positively correlated with MWHC. In addition, the relative importance of each predictor was plotted (Figure 4c), revealing that LS and SOC were the two most important predictors.
4. Discussion
Forests play a vital role in regulating the water cycle of terrestrial ecosystems [35]. Generally, the relationship between the supply of and demand for forest water resources is closely related to forest changes. For example, forest disturbance will transform the forest from a water–supply area to a water-demand area and minimize the water use efficiency in downstream regions [36]. However, planned forest management measures are more conducive to improving the ability of the forest to conserve water and provide high-value hydrological ecosystem services [37]. Our results showed that mixed-species plantations with both coniferous trees and broadleaved trees were more conducive to maintaining the litter–soil continuum’s water storage compared with monoculture plantations (Figure 3), but the tree species used to cope with regional water resource pressure varied greatly, especially when the water-holding capacity of litter or soil was considered separately. In our study, the MWHC and EWHC in PM+MM were higher than those in PM (Figure 1), and PM+CO was the lowest, whether in the OL or OF layer, which may be related to the heterogeneity of litter structure in different tree species [38]. The structure included the area and density of leaves to adjust the amount and rate of water immersion to improve the MWHC of litter [12]. Although the litter stock of the OF layer in PM was the largest (Table S2), the EWHC was low because the simple structure of needles led to a lower litter thickness per unit area; therefore, its spatial hydrophobicity is not conducive to rainwater interception [16].
It is generally believed that changes in soil porosity and hydraulic conductivity caused by vegetation change affect the soil water storage [31]. The accelerated decomposition of mixed litter changes the surface soil–pore structure by increasing the SOC content in the mixed plantation to enhance soil moisture retention (Table S3) [39,40]. Our study showed that except for PM+CJ, the SSWS and CWS at 0–20 cm in other mixed plantations were significantly higher than those in PM, which showed the effect of tree species identity on surface soil water storage. In addition, the increase in soil aggregate stability caused by soil carbon accumulation reduced the impact of rainfall splash on soil [41], protected the soil-pore structure from being compacted, and increased the soil water retention capacity [11]. Interestingly, our study also found that the tree species had significant variations in the water storage of the soil profile (Figure 2). In this study, the SSWS and CWS in PM+CO and PM+MM significantly increased compared to those in PM, which may be related to the rooting depth and root depth distribution of CO and MM [42,43]. Root distribution in deep soil and the release of root exudates not only provide a rich source of organic carbon for deep soils, but also facilitate the cementation of soil particles, the formation of capillary pores, and soil moisture retention [26,44]. At the same time, better soil–pore connectivity and aggregate stability in mixed plantations may become the main channel for the movement of groundwater (Table S3), increase the recharge of groundwater, attenuate soil water evaporation, and alleviate the drying effect of plantation soil in arid and semi-arid areas [45].
Our research demonstrated that the hydrological features of the litter–soil continuum were altered by different tree species compositions (Figure 3); therefore, it is necessary to consider the selection of mixed broadleaved tree species from the perspective of coniferous plantation transformation. MM is probably suitable for a kind of broadleaved tree species mixed into PM plantations, but the mixed CO and PM plantation reduced the water-holding capacity of the litter–soil continuum, indicating synergistic or antagonistic mixed effects depending on the tree species. Globally, LS and LT have positive effects on the hydrological cycle in forests [46], which is similar to the results of this study. Our findings also suggest that LS and SOC were the major factors influencing the litter–soil continuum’s ability to store water (Figure 4), which may be due to the interplays between these two variables. On the one hand, greater litter storage can lead to more carbon input for soil [43,44]. High carbon input can improve soil characteristics by lowering soil bulk density, elevating total porosity, and enhancing field capacity (especially the stability of soil aggregates) [47]. On the other hand, higher litter stock and better soil properties can improve the rainfall interception and water-holding capacity of the litter–soil continuum [14,16], alleviate the pressure of plantations to address drought stress, and promote tree growth and biomass accumulation [48]. Furthermore, LT regulates the entry and movement of soil water by influencing the process of interception in forests, including water permeability and hydraulic conductivity. Ultimately, it affects the water conservation capacity of soil [15], which explains the phenomenon in which LT was also a key factor for the water conservation capacity of the litter–soil continuum observed in our study (Figure 4).
5. Conclusions
We concluded that in subtropical regions, compared with monoculture plantations, the mixed plantation changed the characteristics of the litter and soil properties to affect the water storage of the litter–soil continuum. Mixed planting of PM and MM promoted the decomposition of litter and the vertical distribution of the root system, thus improving the water conservation capacity of the litter–soil continuum; however, CO is not a suitable tree species for water conservation forests because of the strong hydrophobicity of its leaves. In addition, our study also found that LS, LT, and SOC were crucial to the water conservation capacity of the litter–soil continuum, which indicated that complex biogeochemical processes between the litter and soil regulate changes in forest hydrology. Overall, when determining the water conservation capacity of a mixed plantation, it is essential to take into account the comprehensive effects of tree species traits on litter and soil rather than consider them separately.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/f14020431/s1, Table S1: Stand characteristics of the experimental sites. Table S2: Effects of forest type on litter thickness and stock. Table S3: Effects of forest type on soil properties.
Author Contributions
Conceptualization, N.D. and Y.B.; methodology, N.D. and Y.B.; software, N.D. and Y.B.; validation, N.D., Y.B. and Y.Z.; formal analysis, N.D.; investigation, Y.B.; resources, Y.B.; data curation, Y.B.; writing—original draft preparation, N.D.; writing—review and editing, Y.B.; visualization, N.D. and Y.B.; supervision, Y.Z.; project administration, Y.Z.; funding acquisition, Y.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China [Grant No. 32260375], the First-Class Discipline Construction Project in Guizhou Province [GNYL (2017)007], and the 100 High-Level Innovating Project [Grant No. QKHRC-2015-4022].
Data Availability Statement
Not applicable.
Acknowledgments
We thank the Field Scientific Observation and Research Station of Longli National Forest Farm in Guizhou province.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Yu, Y.; Zhu, J.; Gao, T.; Liu, L.; Yu, F.; Zhang, J.; Wei, X. Evaluating the influential variables on rainfall interception at different rainfall amount levels in temperate forests. J. Hydrol. 2022, 615, 128572. [Google Scholar] [CrossRef]
- Young, D.; Zégre, N.; Edwards, P.; Fernandez, R. Assessing streamflow sensitivity of forested headwater catchments to disturbance and climate change in the central Appalachian Mountains region, USA. Sci. Total Environ. 2019, 694, 133382. [Google Scholar] [CrossRef] [PubMed]
- Alaniz, A.J.; Smith-Ramírez, C.; Rendón-Funes, A.; Hidalgo-Corrotea, C.; Carvajal, M.A.; Vergara, P.M.; Fuentes, N. Multiscale spatial analysis of headwater vulnerability in South-Central Chile reveals a high threat due to deforestation and climate change. Sci. Total Environ. 2022, 849, 157930. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, P.V.; Miniat, C.F.; Elliott, K.J.; Swank, W.T.; Brantley, S.T.; Laseter, S.H. Declining water yield from forested mountain watersheds in response to climate change and forest mesophication. Glob. Chang. Biol. 2016, 22, 2997–3012. [Google Scholar] [CrossRef] [PubMed]
- Del Campo, A.D.; Otsuki, K.; Serengil, Y.; Blanco, J.A.; Yousefpour, R.; Wei, X. A global synthesis on the effects of thinning on hydrological processes: Implications for forest management. For. Ecol. Manag. 2022, 519, 120324. [Google Scholar] [CrossRef]
- Haas, H.; Reaver, N.G.F.; Karki, R.; Kalin, L.; Srivastava, P.; Kaplan, D.A.; Gonzalez-Benecke, C. Improving the representation of forests in hydrological models. Sci. Total Environ. 2022, 812, 151425. [Google Scholar] [CrossRef]
- Jiang, M.; Lin, T.-C.; Shaner, P.-J.; Lyu, M.; Xu, C.; Xie, J.; Chengfang, L.; Yang, Y.-S. Understory interception contributed to the convergence of surface runoff between a Chinese fir plantation and a secondary broadleaf forest. J. Hydrol. 2019, 574, 862–871. [Google Scholar] [CrossRef]
- Helvey, J.D.; Patric, J.H. Canopy and litter interception of rainfall by hardwoods of eastern United States. Water Resour. Res. 1965, 1, 193–206. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Niu, J.; Gao, Z.; Chen, X.; Zhang, L.; Li, X.; van Doorn, N.S.; Luo, Z.; Zhu, Z. Effects of rainfall intensity and slope on interception and precipitation partitioning by forest litter layer. CATENA 2019, 172, 711–718. [Google Scholar] [CrossRef]
- Kittredge, J. The Influence of Shade and Litter of Pine on Evaporation from a Clay Loan Soil at Berkeley, California. Ecology 1954, 35, 397–405. [Google Scholar] [CrossRef]
- Chen, Y.; Wei, T.; Ren, K.; Sha, G.; Guo, X.; Fu, Y.; Yu, H. The coupling interaction of soil organic carbon stock and water storage after vegetation restoration on the Loess Plateau, China. J. Environ. Manag. 2022, 306, 114481. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Zhou, Y.; Du, J.; Zhang, X.; Di, N. Effects of a broadleaf-oriented transformation of coniferous plantations on the hydrological characteristics of litter layers in subtropical China. Glob. Ecol. Conserv. 2021, 25, e01400. [Google Scholar] [CrossRef]
- Li, X.; Niu, J.; Xie, B. Study on Hydrological Functions of Litter Layers in North China. PLoS ONE 2013, 8, e70328. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Xu, Q.; Gao, D.; Wang, T.; Sui, M.; Huang, J.; Gu, B.; Liu, F.; Jiang, J. Soil capacity of intercepting different rainfalls across subtropical plantation: Distinct effects of plant and soil properties. Sci. Total Environ. 2021, 784, 147120. [Google Scholar] [CrossRef] [PubMed]
- Marín-Castro, B.E.; Negrete-Yankelevich, S.; Geissert, D. Litter thickness, but not root biomass, explains the average and spatial structure of soil hydraulic conductivity in secondary forests and coffee agroecosystems in Veracruz, Mexico. Sci. Total Environ. 2017, 607–608, 1357–1366. [Google Scholar] [CrossRef]
- Zhao, L.; Hou, R.; Fang, Q. Differences in interception storage capacities of undecomposed broad-leaf and needle-leaf litter under simulated rainfall conditions. For. Ecol. Manag. 2019, 446, 135–142. [Google Scholar] [CrossRef]
- Biswas, A.; Si, B.C. Identifying effects of local and nonlocal factors of soil water storage using cyclical correlation analysis. Hydrol. Process. 2012, 26, 3669–3677. [Google Scholar] [CrossRef]
- Christiansen, C.T.; Haugwitz, M.S.; Priemé, A.; Nielsen, C.S.; Elberling, B.; Michelsen, A.; Grogan, P.; Blok, D. Enhanced summer warming reduces fungal decomposer diversity and litter mass loss more strongly in dry than in wet tundra. Glob. Chang. Biol. 2017, 23, 406–420. [Google Scholar] [CrossRef]
- Feng, Y.; Schmid, B.; Loreau, M.; Forrester, D.I.; Fei, S.; Zhu, J.; Tang, Z.; Zhu, J.; Hong, P.; Ji, C.; et al. Multispecies forest plantations outyield monocultures across a broad range of conditions. Science 2022, 376, 865–868. [Google Scholar] [CrossRef]
- Zhang, B.; Xu, Q.; Gao, D.; Jiang, C.; Liu, F.; Jiang, J.; Wang, T. Altered water uptake patterns of Populus deltoides in mixed riparian forest stands. Sci. Total Environ. 2020, 706, 135956. [Google Scholar] [CrossRef]
- Zhao, Y.; Liang, C.; Shao, S.; Chen, J.; Qin, H.; Xu, Q. Linkages of litter and soil C:N:P stoichiometry with soil microbial resource limitation and community structure in a subtropical broadleaf forest invaded by Moso bamboo. Plant Soil 2021, 465, 473–490. [Google Scholar] [CrossRef]
- Li, F.L.; McCormack, M.L.; Liu, X.; Hu, H.; Feng, D.F.; Bao, W.K. Vertical fine-root distributions in five subalpine forest types shifts with soil properties across environmental gradients. Plant Soil 2020, 456, 129–143. [Google Scholar] [CrossRef]
- Fernández-Alonso, M.J.; Díaz-Pinés, E.; Kitzler, B.; Rubio, A. Tree species composition shapes the assembly of microbial decomposer communities during litter decomposition. Plant Soil 2022, 480, 457–472. [Google Scholar] [CrossRef]
- Almeida, J.P.; Rosenstock, N.P.; Woche, S.K.; Guggenberger, G.; Wallander, H. Nitrophobic ectomycorrhizal fungi are associated with enhanced hydrophobicity of soil organic matter in a Norway spruce forest. Biogeosciences 2022, 19, 3713–3726. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, S.; Jones, J.; Sun, G.; Wei, X.; Ellison, D.; Archer, E.; McNulty, S.; Asbjornsen, H.; Zhang, Z.; et al. Managing the forest-water nexus for climate change adaptation. For. Ecol. Manag. 2022, 525, 120545. [Google Scholar] [CrossRef]
- Bai, Y.; Zhou, Y.; He, H. Effects of rehabilitation through afforestation on soil aggregate stability and aggregate-associated carbon after forest fires in subtropical China. Geoderma 2020, 376, 114548. [Google Scholar] [CrossRef]
- Dai, D.; Ali, A.; Huang, X.; Teng, M.; Wu, C.; Zhou, Z.; Liu, Y. Soil Available Phosphorus Loss Caused by Periodical Understory Management Reduce Understory Plant Diversity in a Northern Subtropical Pinus massoniana Plantation Chronosequence. Forests 2020, 11, 231. [Google Scholar] [CrossRef] [Green Version]
- Bai, Y.; Zhou, Y.; An, Z.; Du, J.; Zhang, X.; Chang, S.X. Tree species identity and mixing ratio affected the release of several metallic elements from mixed litter in coniferous-broadleaf plantations in subtropical China. Sci. Total Environ. 2022, 838, 156143. [Google Scholar] [CrossRef]
- Lee, R. Forest Hydrology; Columbia University Press: New York, NY, USA, 1980; ISBN 9780231882408. [Google Scholar]
- Lu, Y.; Si, B.; Li, H.; Biswas, A. Elucidating controls of the variability of deep soil bulk density. Geoderma 2019, 348, 146–157. [Google Scholar] [CrossRef]
- Mei, X.; Ma, L.; Zhu, Q.; Li, B.; Zhang, D.; Liu, H.; Zhang, Q.; Gou, Q.; Shen, M. The variability in soil water storage on the loess hillslopes in China and its estimation. CATENA 2019, 172, 807–818. [Google Scholar] [CrossRef]
- Rana, S.; Cheng, X.; Wu, Y.; Hu, C.; Jemim, R.S.; Liu, Z.; Wang, Y.; Cai, Q.; Geng, X.; Guo, X.; et al. Evaluation of soil and water conservation function in the Wugong mountain meadow based on the comprehensive index method. Heliyon 2022, 8, e11867. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.W.; Sommers, L.E. Total Carbon, Organic Carbon, and Organic Matter. In Methods of Soil Analysis: Part 3. Chemical Methods; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Eds.; Soil Science Society of America and American Society of Agronomy: Madison, WI, USA, 1996; pp. 963–1010. [Google Scholar] [CrossRef]
- Hou, G.R.; Bi, H.X.; Wei, X. Comprehensive evaluation of water conservation function of litters of robinia pseudoacacia forest lands in gully region on loess plateau. J. Soil Water Conserv. 2019, 33, 251–257. [Google Scholar]
- Jones, J.; Ellison, D.; Ferraz, S.; Lara, A.; Wei, X.; Zhang, Z. Forest restoration and hydrology. For. Ecol. Manag. 2022, 520, 120342. [Google Scholar] [CrossRef]
- Lim, C.-H.; Song, C.; Choi, Y.; Jeon, S.; Lee, W.-K. Decoupling of forest water supply and agricultural water demand attributable to deforestation in North Korea. J. Environ. Manag. 2019, 248, 109256. [Google Scholar] [CrossRef]
- Nordblom, T.L.; Hume, I.H.; Finlayson, J.D.; Pannell, D.J.; Holland, J.E.; McClintock, A.J. Distributional consequences of upstream tree plantations on downstream water users in a Public–Private Benefit Framework. Agric. Syst. 2015, 139, 271–281. [Google Scholar] [CrossRef]
- Hao, G.-Y.; Sack, L.; Wang, A.-Y.; Cao, K.-F.; Goldstein, G. Differentiation of leaf water flux and drought tolerance traits in hemiepiphytic and non-hemiepiphytic Ficus tree species. Funct. Ecol. 2010, 24, 731–740. [Google Scholar] [CrossRef]
- Gong, C.; Tan, Q.; Xu, M.; Liu, G. Mixed-species plantations can alleviate water stress on the Loess Plateau. For. Ecol. Manag. 2020, 458, 117767. [Google Scholar] [CrossRef]
- Mishra, S.; Singh, K.; Sahu, N.; Singh, S.N.; Manika, N.; Chaudhary, L.B.; Jain, M.K.; Kumar, V.; Behera, S.K. Understanding the relationship between soil properties and litter chemistry in three forest communities in tropical forest ecosystem. Environ. Monit. Assess. 2020, 191, 797. [Google Scholar] [CrossRef]
- Ma, R.; Hu, F.; Xu, C.; Liu, J.; Zhao, S. Response of soil aggregate stability and splash erosion to different breakdown mechanisms along natural vegetation restoration. CATENA 2022, 208, 105775. [Google Scholar] [CrossRef]
- Safari, M.; Ghanati, F.; Safarnejad, M.R.; Chashmi, N.A. The contribution of cell wall composition in the expansion of Camellia sinensis seedlings roots in response to aluminum. Planta 2018, 247, 381–392. [Google Scholar] [CrossRef]
- Pan, L.; Lin, W.; Yu, M.; Lie, G.; Xue, L.; Chen, H. Effects of Elevated Ozone Concentrations on Root Characteristics and Soil Properties of Elaeocarpus sylvestris and Michelia chapensis. Bull. Environ. Contam. Toxicol. 2020, 104, 682–688. [Google Scholar] [CrossRef]
- Mawodza, T.; Burca, G.; Casson, S.; Menon, M. Wheat root system architecture and soil moisture distribution in an aggregated soil using neutron computed tomography. Geoderma 2020, 359, 113988. [Google Scholar] [CrossRef]
- Wu, G.-L.; Cui, Z.; Huang, Z. Contribution of root decay process on soil infiltration capacity and soil water replenishment of planted forestland in semi-arid regions. Geoderma 2021, 404, 115289. [Google Scholar] [CrossRef]
- Liu, Y.; Shi, W.; Tao, F.; Shi, X.; Fu, B. A Global Synthesis of Multi-Factors Affecting Water Storage Capacity in Forest Canopy, Litter and Soil Layers. Geophys. Res. Lett. 2023, 50, e2022GL099888. [Google Scholar] [CrossRef]
- Liu, S.; Guo, Z.C.; Pan, Y.B.; Zhang, L.L.; Hallett, P.D.; Peng, X.H. Rare earth oxides for labelling soil aggregate turnover: Impacts of soil properties, labelling method and aggregate structure. Geoderma 2019, 351, 36–48. [Google Scholar] [CrossRef]
- Rowland, L.; da Costa, A.C.L.; Oliveira, A.A.R.; Almeida, S.S.; Ferreira, L.V.; Malhi, Y.; Metcalfe, D.B.; Mencuccini, M.; Grace, J.; Meir, P. Shock and stabilisation following long-term drought in tropical forest from 15 years of litterfall dynamics. J. Ecol. 2018, 106, 1673–1682. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Comparison of maximum water-holding capacity (a) and effective water–holding capacity (b) of litter in different forest types. Note: OF: semi-decomposed litter; OL: undecomposed litter. The lowercase letters illustrate significant differences among different forest types in the same litter layer (p < 0.05). Pinus massoniana (PM), Bretschneidera sinensis (BS), Manglietia chingii (MC), Cercidiphyllum japonicum (CJ), Camellia oleifera (CO), and Michelia maudiae (MM).
Figure 1.
Comparison of maximum water-holding capacity (a) and effective water–holding capacity (b) of litter in different forest types. Note: OF: semi-decomposed litter; OL: undecomposed litter. The lowercase letters illustrate significant differences among different forest types in the same litter layer (p < 0.05). Pinus massoniana (PM), Bretschneidera sinensis (BS), Manglietia chingii (MC), Cercidiphyllum japonicum (CJ), Camellia oleifera (CO), and Michelia maudiae (MM).
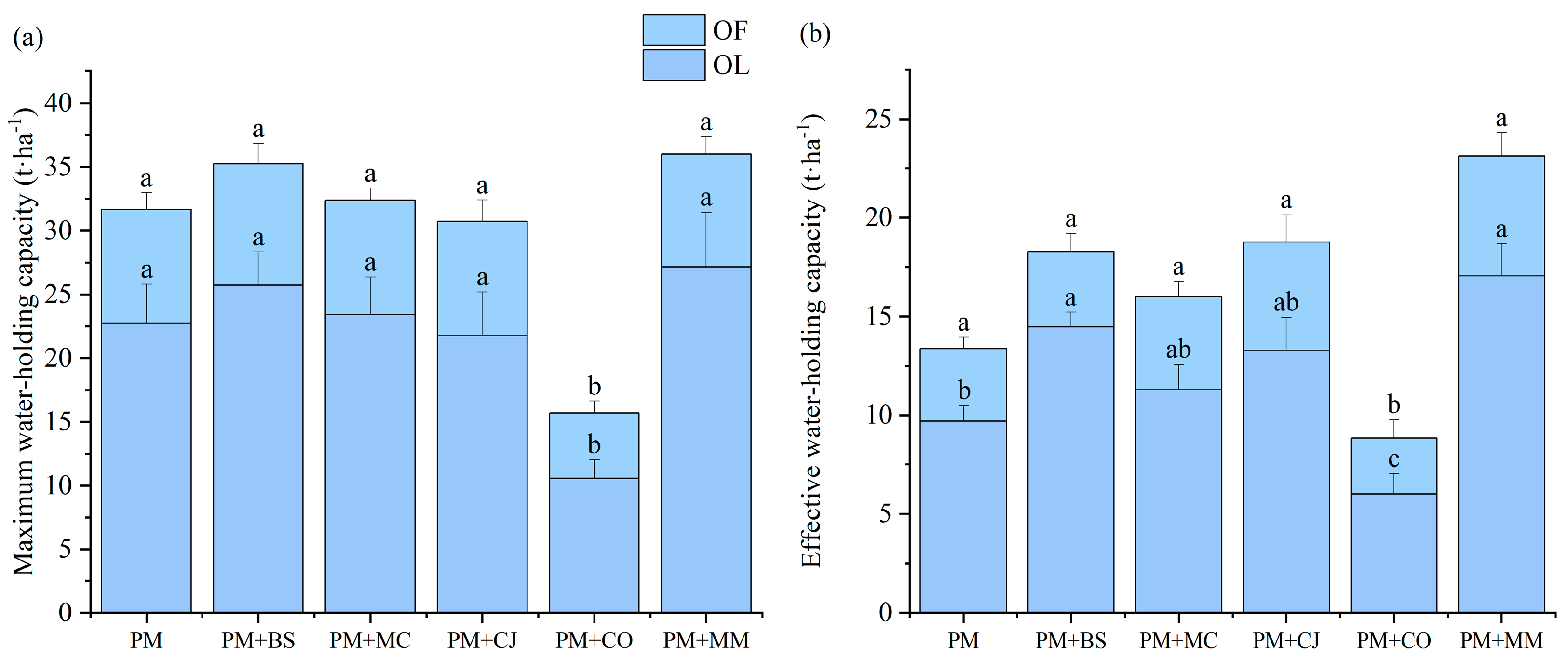
Figure 2.
(a) Soil–saturated water storage, (b) capillary water storage, and (c) noncapillary water storage in different forest types. Note: the points denote the overall mean values, and the shaded regions represent the 95% CIs. Pinus massoniana (PM), Bretschneidera sinensis (BS), Manglietia chingii (MC), Cercidiphyllum japonicum (CJ), Camellia oleifera (CO), and Michelia maudiae (MM).
Figure 2.
(a) Soil–saturated water storage, (b) capillary water storage, and (c) noncapillary water storage in different forest types. Note: the points denote the overall mean values, and the shaded regions represent the 95% CIs. Pinus massoniana (PM), Bretschneidera sinensis (BS), Manglietia chingii (MC), Cercidiphyllum japonicum (CJ), Camellia oleifera (CO), and Michelia maudiae (MM).
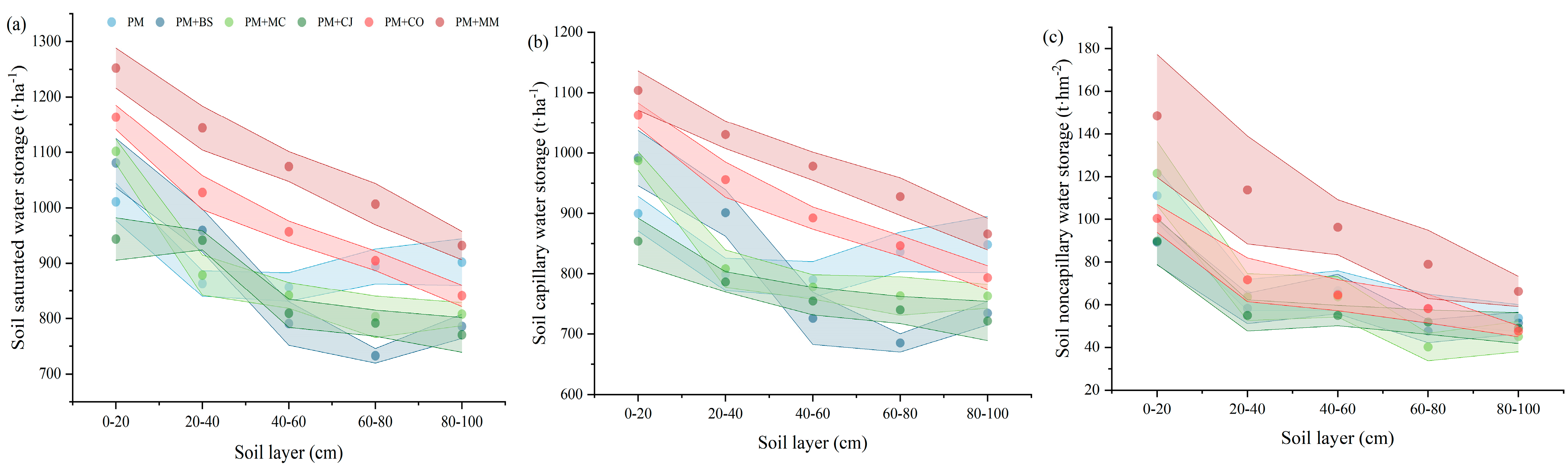
Figure 3.
Comprehensive evaluation of water capacity function in the litter–soil continuum of different forest types. Note: the lower the comprehensive evaluation score, the higher the water conservation capacity in the litter–soil continuum. Pinus massoniana (PM), Bretschneidera sinensis (BS), Manglietia chingii (MC), Cercidiphyllum japonicum (CJ), Camellia oleifera (CO), and Michelia maudiae (MM).
Figure 3.
Comprehensive evaluation of water capacity function in the litter–soil continuum of different forest types. Note: the lower the comprehensive evaluation score, the higher the water conservation capacity in the litter–soil continuum. Pinus massoniana (PM), Bretschneidera sinensis (BS), Manglietia chingii (MC), Cercidiphyllum japonicum (CJ), Camellia oleifera (CO), and Michelia maudiae (MM).
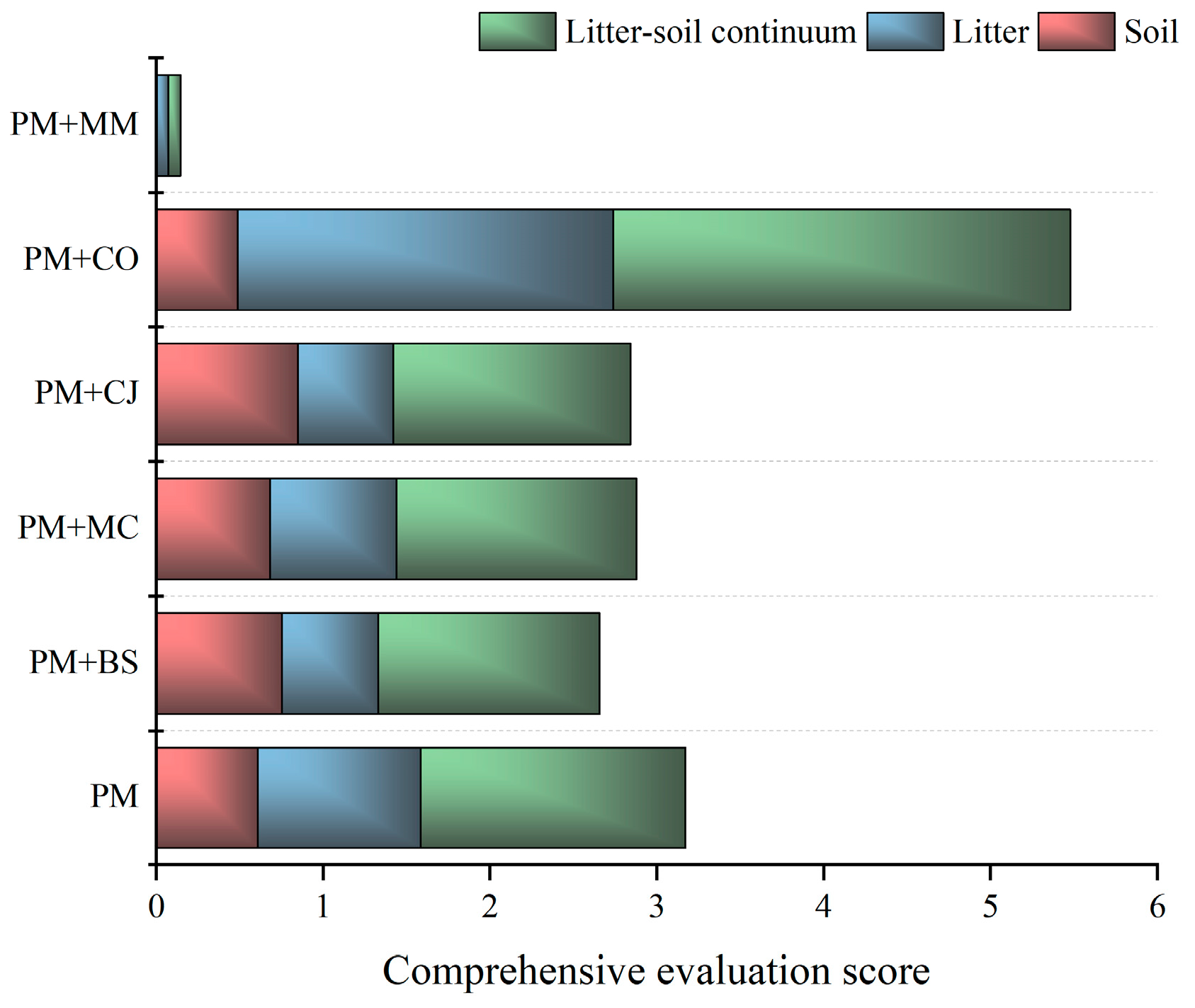
Figure 4.
Factors influencing the water conservation capacity of the litter-soil continuum. (a) Redundancy analysis (RDA) for the associations of environmental factors with water conservation capacity. (b) Correlation between environmental factors and water conservation capacity. (c) Independent effects of environmental factors on water conservation capacity. Note: SOC: soil organic carbon; LS: litter stock; LT: litter thickness; GMD: geometric mean diameter; MWD: mean weight diameter; SSWS: soil saturated water storage; D: fractal dimension; CWS: capillary water storage; MWHC: maximum water–holding capacity of litter; NCWS: noncapillary water storage; and EWHC: effective water-holding capacity of litter. Pinus massoniana (PM), Bretschneidera sinensis (BS), Manglietia chingii (MC), Cercidiphyllum japonicum (CJ), Camellia oleifera (CO), and Michelia maudiae (MM).
Figure 4.
Factors influencing the water conservation capacity of the litter-soil continuum. (a) Redundancy analysis (RDA) for the associations of environmental factors with water conservation capacity. (b) Correlation between environmental factors and water conservation capacity. (c) Independent effects of environmental factors on water conservation capacity. Note: SOC: soil organic carbon; LS: litter stock; LT: litter thickness; GMD: geometric mean diameter; MWD: mean weight diameter; SSWS: soil saturated water storage; D: fractal dimension; CWS: capillary water storage; MWHC: maximum water–holding capacity of litter; NCWS: noncapillary water storage; and EWHC: effective water-holding capacity of litter. Pinus massoniana (PM), Bretschneidera sinensis (BS), Manglietia chingii (MC), Cercidiphyllum japonicum (CJ), Camellia oleifera (CO), and Michelia maudiae (MM).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ding, N.; Bai, Y.; Zhou, Y. Tree Species Mixtures Can Improve the Water Storage of the Litter–Soil Continuum in Subtropical Coniferous Plantations in China. Forests 2023, 14, 431. https://doi.org/10.3390/f14020431
AMA Style
Ding N, Bai Y, Zhou Y. Tree Species Mixtures Can Improve the Water Storage of the Litter–Soil Continuum in Subtropical Coniferous Plantations in China. Forests. 2023; 14(2):431. https://doi.org/10.3390/f14020431
Chicago/Turabian StyleDing, Ni, Yunxing Bai, and Yunchao Zhou. 2023. "Tree Species Mixtures Can Improve the Water Storage of the Litter–Soil Continuum in Subtropical Coniferous Plantations in China" Forests 14, no. 2: 431. https://doi.org/10.3390/f14020431
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.