
Roe Deer, Lithuania’s Smallest and Most Abundant Cervid


Nature Research Centre, Akademijos 2, 08412 Vilnius, Lithuania
Forests 2024, 15(5), 767; https://doi.org/10.3390/f15050767
Submission received: 29 March 2024
/
Revised: 23 April 2024
/
Accepted: 25 April 2024
/
Published: 27 April 2024
(This article belongs to the Special Issue Wildlife Ecology and Conservation in Forest Habitats)
Abstract
:A review of 113 scientific and other publications on the smallest and most abundant deer in Lithuania, the roe deer (Capreolus capreolus), is presented, along with an analysis based on compound annual population growth rates of population numbers, hunting bags, and roadkill. This review covers the species’ history in the country from the last glaciation onwards, the changes in numbers from 1934 to 2023, hunting bag changes from 1965 to 2022, roadkill numbers from 2002 to 2022, data on roe deer reproduction, habitat selection, genetic diversity, pathogens, and damage to forest stands. It also provides an overview of species management and selection for trophies. Despite the exponential increase in roe deer numbers since 1990 and the dominance of this species in roadkill, even on urban roads, the number and density of animals and the damage they cause to forest stands are relatively low compared to other European countries. Within the observed period, drops in numbers were related to harsh winters in 1969/1970 and 1995/1996. Poaching, especially coinciding with weakened enforcement during the period of political–economic transition in the early 1990s, has also had a negative impact on population numbers, as have recent increases in the number of large predators. Population growth over the recent period does not correlate with hunting mortality, which has remained stable at an average of 16.9% since 2002.
Keywords:
Capreolus capreolus; abundance; hunting; roadkill; population management; genetics; pathogens; Lithuania1. Introduction
With a vast range in the Palearctic region, the roe deer (Capreolus capreolus) inhabits most of Europe, excluding Ireland, southern islands such as Sicily, Corsica, Sardinia, Cyprus, and the far northern latitudes [1,2,3]. The species has been present in Europe for over half a million years, enduring through glacial and interglacial periods, and it has been one of the most abundant mammals in the Late Quaternary [4].
Randi et al. highlighted that genetically, European roe deer can be categorized into three haplogroups: one in the west (mainly Iberia), another in the east (Balkans and Greece), and a third in the central part of the continent (spanning across Europe, including the eastern region); however, this study did not include samples from Lithuania [5]. Subsequently, Lorenzini and Lovari [6] classified Lithuanian roe deer into a central–northern group, alongside animals from Romania, Turkey, Greece, and Poland. Roe deer populations in eastern and central parts of Europe exhibit the highest genetic diversity, likely attributable to introgression from the Siberian roe deer (Capreolus pygargus) [7].
The roe deer is a generalist browser, but they also eat grasses and sedges. Species occupies “deciduous wooded areas with glades and clearings, as well as in fields interspersed with wood patches, mostly in temperate climates” [1]. However, in areas with small forest fragments and intensive agriculture, the so-called “field roe deer” ecotype evolved [8]. Its biology differs from that of woodland roe deer in several respects [9]. Field roe deer foraging in agricultural areas outside forests can have substantial impacts on crops [10]. The field roe deer ecotype is also widespread in Lithuania [11,12,13].
Roe deer are the most abundant deer in the mid-latitudes [1,14], and management of this species is therefore linked to ecological, economic, and other aspects [15]. Recently, the management of the species has faced a number of challenges [16,17] due to the overabundance of ungulates in Europe [18], including human safety issues due to the increasing number of wildlife and vehicle accidents, even in urban areas [19].
Roe deer are important as browsers, playing a significant role in forest regeneration [20]. Furthermore, they can serve as a model species for studying the impacts of climate change [21] and landscape change [22]. In heterogeneous European landscapes, roe deer could be utilized as an indicator species [23]. Despite any disagreement with the authors’ position, roe deer have been recognized as ecosystem engineers [24].
Roe deer have been a good indicator of political and economic change in the post-1990 post-socialist countries of Eastern and Central Europe [25,26]. Currently, the management of roe deer populations in European National Parks has shown the need for an integrated management approach [27].
However, many aspects of roe deer ecology and management in Lithuania are either unexplored or inaccessible to non-native speakers. The aim of this review is therefore to provide a comprehensive overview of roe deer in Lithuania, including the history of the species, its current status and abundance, ecology, and other aspects of the species’ management, based on published sources. Changes in the management of the species in relation to legislation are assessed. In the absence of publications on population dynamics, we have supplemented the review with trend analysis. Management options have been analyzed in the light of the political and governance changes that have occurred in the country since the 1960s.
2. Materials and Methods
2.1. Scope of Literature Sources and Filtering of References
Given that many of the references on roe deer are in the national language and those previously published are in Russian, we carried out several literature searches. The two main terms to be included in the search were “roe deer” as a species and “Lithuania” as a geographical reference. In some references, Lithuania was not included in the search terms because all Baltic countries were included.
Further on, we used search terms including biology, ecology, abundance, diet, reproduction, hunting, management, and roadkill of roe deer (Table 1).
Three search engines were used to search for published information sources: the Web of Knowledge, Google Scholar, and Google. The latter was needed for books published in Lithuanian. Our search period covered the period from 1900 to 2024. Boolean combinations of terms were used, such as
also using the same terms in Lithuanian and Russian languages.
(roe deer OR Capreolus capreolus) AND (history OR survey OR hunting OR roadkill OR monitoring OR management) AND Lithuania,
Two problems with the key term “roe deer” in Lithuanian were the fact that it has the same species name in Latvian and that it is popular as a surname or company name. For this reason, the resulting reference lists were manually sorted to exclude sources not related to the animal.
Student works, such as Master’s and Bachelor’s theses, were critically analyzed and singled out because they did not contain original information on the species. The literature sources that did not provide original information and relied on other sources were also excluded. Thus, the list of sources to be reviewed included 114 sources. The remainder of the cited sources are related to other countries.
2.2. Study Site and Sources of Empirical Data
The study area with its forest cover, main cities, and the main roads is shown in Figure 1. Data on the recent forest cover and forest cover dynamics are taken from the Official Statistics Portal [28,29]. Data on regulations concerning roe deer management were collected from official sources [30,31,32,33,34]. The hunting periods for roe deer are taken from the hunting calendar [35].
The size of the national roe deer herd (number of animals surveyed) and the number of hunting bags (number of animals hunted per season) were used to describe population dynamics and trends. We used official data on the number of surveyed animals [36] and data on the hunting bag [37], previously available in the archives of the Ministry of the Environment of the Republic of Lithuania, with data for the period 2015–2023 only being available on the internet.
The data on roadkill of roe deer were provided by the Lithuanian Police Traffic Supervision Service, comprising the sum of reported roadkill documented in officers’ reports. In the past decade, the verification of roadkill has been conducted by the nearest hunting clubs. Additionally, unregistered roadkill numbers of roe deer were compiled by professional biologists from the Nature Research Centre. These figures represent the sum of animals found roadkilled by scientists but not documented in the reports of police officers.
Until 1990, the main survey method was snow-tracking, adjusted according to the number of animals recorded visually. Game counts in the snow by foresters and hunters were carried out in a coordinated manner on the same day throughout the country, and the results obtained were aggregated at several levels. Subsequently, after the 1990s, the population size was taken as the sum of the animals present in all hunting areas in the country, as reported by hunting clubs. In the last decade, wildlife cameras were used by nearly all clubs, however, there are no publications on the roe deer numbers. After 2018, the snow-track index [38] has again been used to analyze population trends, but not the number.
We understand that official figures may be biased, but this is the only source of game numbers. These data have been used in publications covering Europe and the Baltic States [14,16,26,27,39], Lithuania [11], and at the local level [40].
Data on roe deer damage were taken from the annual reports [41].
2.3. Data Analyses
We analyzed trends in population numbers, hunting bag numbers, and roadkill numbers using the compound annual growth rate (CAGR) and regression analysis. Various time periods were selected to represent different population phases.
CAGR was expressed as the number required for the population or hunting bag to change from the beginning value to an end value. We used the following expression:
where IV is the initial value, EV is the end or final value, and n is the number of years [42]. Larger CAGR values indicate faster growth, while negative CAGR values indicate a reduction in numbers over the specified time period.
We employed basic statistics, including Pearson correlation and the standard error of the mean, for our analysis. Regression analysis was conducted to determine the best fit to the raw data based on the least squares criteria, utilizing either linear, power, or exponential regression models. The best regression model was selected using the coefficient of determination, R2.
All calculations were done in MS Excel, ver. 16.0.5387.1000 and Statistica for Windows, ver. 6.0 [43].
3. Results
3.1. Historic Data on Presence of Roe Deer in Lithuania
As pointed out by Sommer et al. [44], “the European roe deer is a typical faunal element of the Holocene”, and it was present in Europe for over 600,000 years, surviving several glaciations in refugia as were other ungulate species [45]. During the last glaciation period, the distribution range of roe deer was at its minimum [44]. Central and northern Europe were subsequently recolonized after the glaciation, as fossils of the species older than 11,000–12,000 years are not present [6]. The current distribution of the species is limited by permafrost, leading to the absence of roe deer beyond the Arctic Circle in Scandinavia [5]. Furthermore, forest re-establishment after deglaciation was followed by the expansion of the species’ range.
While the teeth of ungulates were commonly found in Mesolithic burials across Europe [46], those of roe deer were not discovered in Lithuania [47]. From the Early Neolithic to the Early Bronze Age, the number of findings of roe deer bones and teeth was considerably lower compared to those of moose (Alces alces) and red deer (Cervus elaphus) [48]. According to these authors, roe deer bones accounted for only 1.54% of the animal remains in the Early Neolithic, 2.99% in the Middle Neolithic, 2.32–6.66% in the Late Neolithic periods, and 3.93% in the Early Bronze Age. Furthermore, based on stable isotopes, there is no evidence of roe deer presence in the diet of humans during that period [49].
From the Late Paleolithic to the Middle Bronze Age, hunting was the main economic activity in Lithuania, but roe deer were not often hunted—hunting was focused on the largest ungulates such as reindeer (Rangifer tarandus), moose, and red deer [50]. As the landscape changed and forests expanded from the river valleys into the rest of the country, the composition of game also changed, but roe deer were again in the minority.
As the Iron Age progressed, the importance of hunting gradually diminished with the appearance of other sources of meat, such as domestic animals [50]. M. Vitkūnas [51] demonstrated that the proportion of game animal bones in archaeological findings decreased over time: it was up to 35.4% in the last millennium BC, then decreased to 27.5% in the first 400 years AD, further dropping to up to 7% before the end of the first millennium AD, and finally to 3.1% until 1400 AD. The proportion of roe deer bones in the latter case ranged from 3% to 3.7% of all bones at several sites. This suggests that either roe deer were not numerous or larger game were the main target of hunting [52]. Additionally, roe deer were not numerous in later periods, as evidenced by materials from the 14th to 17th centuries excavated in the castle of Vilnius, the capital city, where roe deer bones were absent [53].
In the 15th and 16th centuries, roe deer were a common and widespread animal in Lithuania [54,55,56]. In the 16th and 19th centuries, their numbers declined due to the colder climate [57].
Towards the 20th century, roe deer distribution declined and became fragmented due to uncontrolled hunting [11] and the influence of predators [58]. In Lithuania, a decline in roe deer numbers due to both of these causes was reported at the end of the 18th century [59]. In the second half of the 19th century, roe deer in Lithuania became rare or endangered [11,60].
3.2. Numbers of Roe Deer in Lithuania from the 20th Century to the 1990s
At the European level, the roe deer population started to increase again in the 20th century [1]. However, in Lithuania, the trends were different. Poaching and wolf population growth led to a critical decline in roe deer numbers during the First World War [11]. It was not until 1928 that it was concluded that roe deer were no longer threatened with extinction due to their rapid reproduction and ability to adapt to their environment [61].
In the 1930s, the number of roe deer increased (Figure 2), from 13,930 in 1934 to 35,030 in 1949 [11]. The compound growth rate of the roe deer population during this period was CAGR = 0.202. However, during the Second World War and the post-war years, the number of roe deer decreased drastically due to poaching and preying by wolves. In 1948, only 8500 roe deer remained, CAGR = −0.146. Wolf numbers at the end of the 1940s were at about 1700 [59].
The next decade, 1948–1958, was characterized by a slow increase in roe deer numbers (CAGR = 0.084); despite the extermination, numbers of wolves remained high, and poaching still had a substantial influence. Subsequently, numbers increased more rapidly (CAGR = 0.138 during 1959–1968), reaching 70,800 individuals in 1968. The growth of the population in some regions halted even without roe deer being hunted [62], followed by a reduction in breeding intensity [63].
The harsh and deep snow winter of 1969/1970 resulted in a significant decrease (CAGR = −0.20) in roe deer numbers, declining from 68,500 in 1969 to 44,000 in 1971 [11]. Subsequently, roe deer numbers remained at the same level (CAGR = −0.002) until 1991, when the population was 44,400 roe deer.
3.3. Numbers of Roe Deer in Lithuania during 1990–2023
In the last decade of the 20th century, roe deer numbers remained nearly stable (CAGR = 0.003), at just over 40,000 individuals [11]. A reduction in roe deer numbers in 1996–1997 was attributed to intensive hunting (see below), an increase in wolf numbers, and increased poaching associated with weakened controls during the political–economic transition [26,64,65]. Additionally, the harsh and deep snow winter of 1995/1996 resulted in a decrease in roe deer numbers from 41,000 to 36,500 in the country. However, locally, the loss ranged from 20% to 50%, and in some forests, it amounted to up to 80% of the roe deer population [66].
In general, after 1998, the increase in the roe deer herd in Lithuania lasted until 2020 (see Figure 2). The growth rate was not very steep (CAGR = 0.059), with the fastest growth occurring during the years 2014–2020 (CAGR = 0.084). The roe deer population growth after 1990 is best described as exponential (R2 = 0.94), as indicated by the equation
where x is the year.
3.4. Roe Deer Hunting
The first period of legal hunting of roe deer in the 20th century was 1936–1939, during which time the number of individuals hunted increased from 486 to 868 and their share in the population was 1.96%–2.48%. Following the subsequent decline in the population, roe deer hunting only resumed in 1965 (Figure 3).
From 1965 to 1996, the numbers of roe deer bags remained stable (CAGR = 0.010), averaging about 6500 ± 500 annually, with a minimum of 910 in 1970 and a maximum of 10,844 in 1976, followed by a second drop in 1979. Meanwhile, the proportion of the population that hunted fluctuated widely (see Figure 4), averaging 14.5% ± 1.3%. However, it reached a minimum of 1.5% in 1970, dropped to 5.1% in 1979, and peaked at 28.3% in 1993. From 1993 to 1997, the proportion of hunted roe deer sharply decreased (CAGR = −0.882). In 1997, under a hunting ban for roe deer, only two individuals were reported to have been hunted.
From 1999 to 2019, the increase in the roe deer hunting bag was steady (CAGR = 0.115), with only a small drop in 2011 (see Figure 3). During this period, the hunting bag size increased from 3558 to 31,564 individuals, with the latter number representing the absolute maximum. The increase in the hunting bag (CAGR = 0.053) from 1999 to 2019 was consistent with the population increase. Therefore, the proportion of the population that hunted remained stable from 2002 onwards, averaging 16.9%. There was a drop in roe deer hunting bag size and proportion of the population hunted from 2020 to 2022 (see Figure 3 and Figure 4); however, there are no data on changes in hunting effort.
3.5. Roe Deer Roadkill in 2002–2022
Roe deer are the most abundant species among ungulate roadkill incidents [67]. Out of 50,679 animals killed on the roads of Lithuania during 2002–2022, 32,944 were ungulates, including 32,707 cervids. Among these, 29,199 collisions were registered as cases of roe deer roadkill, while 57 incidents went unreported to the Lithuanian Police Traffic Supervision Service. The correlation between roadkill incidents and population numbers is very high (r = 0.94, p < 0.001). Extrapolating from unregistered roe deer roadkill numbers based on the sampling effort, it is estimated that approximately 9200 individuals were not recorded by the Service during 2002–2022 [67].
The changes in roadkilled roe deer numbers on main, national, regional, and urban roads, based on [19], are presented in Table 2. An exponential increase in registered roe deer roadkill numbers, from 150 in 2002 to over 3500 individuals in 2019, 2021, and 2022, was significant (R2 = 0.97) and best expressed on urban roads.
The decrease in roadkill incidents in 2020, observed on main, national, and regional roads, can be attributed to COVID-19-related human and vehicular mobility restrictions [68]. However, on urban roads, the number of roe deer roadkill incidents increased during the COVID-19 restrictions [19]. This increase was one of the most notable consequences of the so-called “anthropause” [69].
Two patterns of roe deer roadkill were observed: an increase in the proportion of roe deer in the total roadkill (Table 2) and the habitat distribution of roadkill, which was not related to forest habitats (Figure 5). The proportion of roe deer in the total roadkill exceeded 50% after 2014, rising to nearly 75% in 2022. Decreases in this proportion in 2008, 2009, and 2013 were related to intensive roadkill studies conducted at the Nature Research Centre, which focused on registering the number of smaller mammals [70].
Among other cervids, roe deer roadkill incidents were the least related to forest habitats [67]. With the increase in roe deer numbers roadkilled in urban areas [19], the proportion of roadkill in forested areas decreased, particularly in recent years. From 2018 to 2022, the proportion of roe deer roadkilled in forest habitats was consistently less than 20% (see Table 2).
This pattern is most evident in the southwestern, western, and central–northern parts of the country, where forest cover is below average [67], creating conditions conducive to the existence of the field roe deer ecotype [11,71]. These animals exhibit morphometric differences [12] and genetic distinctions [72] from forest-dwelling roe deer. Field roe deer are also common in northeast Poland [8,9]. The formation of this ecotype depended on scarce forest cover and the abundance of forage in agricultural areas [10]. Landscape changes in post-Soviet countries [16,25,26] have favored the formation or support of the field ecotype of roe deer.
The influence of cervid roadkill on population management in Lithuania has been studied for nearly two decades [73]. While roadkill incidents were very important for moose during years of low population density and limited hunting [74], this was not the case for red deer, as roadkill numbers are very limited [75]. Roadkill incidents involving roe deer are the most notable in proportion to the hunting bag of all cervids, averaging 7.7%. However, this proportion increased to 16.5%–16.6% in 2020–2021 [67].
There is no single solution to the problem of roadkill; therefore, in Lithuania, several approaches were investigated, including the following:
- The use of chemical repellent to deter ungulates from entering main roads through gaps in wildlife fencing was investigated as a quick solution before the fence was repaired [76]. It was demonstrated that the reduction in ungulate entry, particularly in roe deer, exceeded 40% when using Wam Porocol®, Witasek PflanzenSchutz GmbH, Feldkirchen, Austria.
- Understanding the temporal patterns of roadkill incidents [77,78,79] reveals that the majority of casualties occur during dusk in May and September, primarily between 8:00 p.m. and 10:00 p.m., and on weekends (Friday to Sunday) [80]. However, roe deer are not the main contributors to human casualties.
- Testing the effectiveness of wildlife fencing as a mitigation tool to prevent ungulates from entering roads has been conducted [84].
- Identifying intersections between urban corridors and wildlife serves as a framework for planning wildlife-vehicle collision mitigation measures [85].
- Given limitations in the use of wildlife fences, there is a need to improve driver awareness regarding hotspot locations and the most dangerous times for collisions [84]. Fencing on main roads can lead to the redistribution of ungulate roadkill toward roads characterized by lower speed and traffic intensity.
3.6. Environmental Parameters and Roe Deer Ecology
The increase in roe deer numbers and harvest has been observed across much of Europe, particularly in the western part [14]. The synchrony of harvested numbers across the continent suggests common factors affecting abundance. Hagen et al. [86] found “a significant positive correlation between the North Atlantic Oscillation winter index (December–March) and annual changes in roe deer bag records for Czech Republic, Germany, Latvia, Lithuania, Norway, and Sweden”. They concluded that periodic fluctuations in population size and harvest are induced by the interplay of environmental factors and human-induced changes.
In Lithuania, roe deer ecology is inadequately understood, with much of the available data being over three decades old [11]. Previous studies concluded that roe deer densities are directly proportional to land fertility and the proportion of deciduous trees in forest stands, while inversely proportional to woodland cover. Land fertility and afforestation are indirect factors influencing the protective qualities and fodder basis of roe deer habitats [87,88]. This suggests that the increased anthropogenic nature of the landscape contributes to higher ungulate densities in Lithuania [89], a phenomenon reflected in their territorial movements [90].
Predation, particularly by wolves, and mortality during severe winters have the most substantial negative impact on roe deer abundance [64,65,66,91,92] The winters of 1969/1970 and 1995/1996 serve as notable examples of such negative consequences for roe deer [11]. Snow cover exceeding 40 cm was considered detrimental to the species survival [93].
The influence of environmental parameters, particularly the composition of forest stands, on roe deer abundance has been studied since the late 1960s [94], with a focus on the food resources available to roe deer and their impact on forest stands. Subsequent analyses of the roe deer diet in relation to forest cover [95,96] not only determined the species composition of plants eaten (including 20 species of trees and shrubs and 95 herbaceous plant species), but also assessed the importance of agricultural habitats [97,98]. Supplementary food sources from agricultural fields were acknowledged as the main factor contributing to increased cervid abundance in the country after the 1980s [89,90]. Another important source of food for roe deer was the remnants of forest cutting [99], which were also processed into special pellets [100].
In the 1990s, the distribution of roe deer across various landscape types and forest stands was assessed based on winter pellet counts [101], revealing the highest importance of forest stands over 20 years old, as well as young pine stands on coastal and sandy plains [102]. Additionally, the spatial distribution of roe deer was found to be dependent on the presence of undergrowth and dwarf shrubs [103].
Roe deer have been observed as city-dwelling animals [104]. Subsequently, the various effects of urbanization on cervids, including roe deer, were analyzed by Brazaitis et al. [105]. It is reasonable to assume that roe deer numbers in urban areas have been increasing over the last decade, with a significant turning point being the limitations on human mobility during the COVID-19 pandemic [19,68].
Data on the reproduction of roe deer in Lithuania are outdated, primarily referencing the 1970s [11]. Calves are typically born in May–June [59]. The average number of embryos per pregnant doe was recorded at 1.9, with 96.6% of adult females breeding [63,106]. Regarding the presence of yellow bodies in roe deer ovaries, 12.6% of females had one, 61.8% had two, 22.3% had three, and 3.3% had four. Older roe deer (6+ years old) tended to have more yellow bodies, averaging 2.5, compared to middle-aged (3–5 years old) and young (1–2 years old) individuals, with 2.2 and 1.9 yellow bodies, respectively [11]. In other countries, large litters (>3 offspring) in roe deer are exceptions, but they might have the potential to further increase populations if monitored more frequently [107].
There are few publications on the field roe deer ecotype in Lithuania, although its formation in Southwest Lithuania began in 1965–1967 [12]. This region is characterized by very rich soil and minor forest cover, comprising only 9% of the area. Field roe deer are distinguished by their larger body mass, ranging from 20.1 to 20.4 kg in juvenile females and males, to 27.7–31.0 kg in adult animals, respectively. In contrast, adult forest roe deer do have a body mass of 26.1 kg, and bucks 29.1 kg [88], resulting in a difference of 1.6–1.8 kg. In Poland, field roe deer were reported to be even heavier than forest ones, by 2.2 kg for males and 3.2 kg for females [108]. Additionally, the cranium and trophy value of field roe deer were reported to be larger [109].
3.7. Genetic Diversity of Lithuanian Roe Deer
Genetic analysis conducted at various spatial scales across Central and West Europe revealed the presence of mtDNA from the Siberian roe deer. In the Pannonian region of Hungary, this species was represented in nearly 3% of samples [110]. Conversely, in northeastern Poland, mtDNA from the Siberian roe deer was found in half of the investigated roe deer [111]. These findings suggest either natural hybridization in the past or the translocation of Siberian roe deer by humans. Interestingly, no genetic differences were observed between field and forest roe deer by these authors, indicating that the species’ ability to thrive in such diverse environments relies on phenotypic plasticity.
Three studies conducted on a continental scale have assessed genetic diversity patterns across the range of roe deer, incorporating samples from Lithuania. In the Eastern European deme, which spans Belarus, Lithuania, Latvia, and the European part of Russia, the highest number of haplotypes of the Siberian roe deer was identified [7]. It was observed that the introgression of the Siberian lineage decreased from eastern to central Europe [112]. These patterns likely depend on the survival of roe deer during the last glacial maximum across a wide range, including two northern refugia. During recolonization, the Baltic Sea has been regarded as a barrier to gene flow [113]. Consequently, the genetic makeup of the species in Lithuania holds interest. However, research on this subject has been notably limited thus far.
Using RAPD (random amplified polymorphic DNA) and enzyme studies, researchers identified nine polymorphic loci and confirmed differences in genetic variability among local roe deer populations [114]. Subsequently, analysis of data from the 457 bp mtDNA control region sequences revealed widespread introgression of Siberian roe deer, with three out of six identified haplotypes being specific to roe deer from Lithuania [115]. Further analysis of microsatellite and D-loop sequences demonstrated a high level of molecular genetic variation in roe deer populations in Lithuania [116].
Genetic differentiation between the field and forest ecotypes, as assessed using mtDNA and AMOVA, was found to be not significant. This includes differences in genetic diversity parameters such as heterozygosity and allelic richness [71]. The authors concluded that both field and forest ecotypes of roe deer in Lithuania are interbreeding and share a recent common gene pool.
3.8. Pathogens of Lithuanian Roe Deer
Roe deer can suffer from very dangerous diseases such as foot-and-mouth disease, brucellosis, tuberculosis, and rabies [58], and they have a variety of endoparasites [117]. Infectious diseases in roe deer have not been studied [11].
Pathogens of roe deer were analyzed during three periods. In the first period, the focus was on coccidian and helminth pathogens. Seven coccidian species were identified, including Eimeria lituanica n.sp., discovered in roe deer [118]. The most frequently encountered coccidians in roe deer were Eimeria capreoli and E. ponderosa [119]. In the internal organs of roe deer, 27 species of helminths were identified, including 3 flukes, 3 tapeworms, and 22 roundworms [120,121,122,123]. However, this represented only a fraction of the helminth fauna known to infect roe deer at that time, with 68 species identified in the Soviet Union [58]. The most frequently found roundworms were Chabertia ovina (prevalence—80% of investigated roe deer), Bunostomum trigonocephalum (25%), and Trichocephalum capreoli (14%). Ch. ovina and Varestrongylus capreoli were the most pathogenic, particularly in cases of intense infection [123]. The helminth fauna of roe deer is influenced by various factors, including the age of the animals, the availability of food, the prevalence of helminths in domestic livestock, and the density of the roe deer herd.
In the second period, beginning in the late 1990s, investigations into the muscle parasites of roe deer were primarily conducted. Four species of Sarcocystis were identified from the cysts, including S. capreolicanis, S. cf. hofmanni, and S. gracilis [124]. The prevalence of Sarcocystis infection among roe deer was 87.5% in central, northern, and eastern Lithuania. The number of cases with intense infection in roe deer was higher than in other ungulates, with a median intensity of 26.8% [125]. The main limitation of this period was the conventional method of light microscopy, which was not supported by molecular methods.
The prevalence of Sarcocystis infection, ranging from 94.4% to 100.0% in various muscle groups of roe deer, was confirmed in the third period of investigations [126]. This study marked the last investigation solely based on light microscopy. Molecular methods confirmed the presence of six Sarcocystis species in roe deer from Lithuania: S. capreolicanis, S. gracilis, S. linearis, S. oviformis, S. silva, and S. entzerothi [127], with the latter described as a new species [128].
In the last decade, molecular methods have been instrumental in identifying pathogens associated with cervids. Two Babesia species, B. microti and B. venatorum, were identified in ticks, Ixodes ricinus and Dermacentor reticulatus, collected from cervids [129]. Other pathogens identified in roe deer include Anaplasma phagocytophilum, with a prevalence of 47%, and Bartonella spp., with a prevalence of 15.7% [130]. Using nested PCR targeting different parts of the 18S rRNA gene of Babesia spp., these pathogens were detected in roe deer with a prevalence of 70.8%, identifying Babesia capreoli, B. odocoilei, B. venatorum, and B. divergens [131]. According to the 16S–23S rRNA ITS region, samples of Babesia from wild animals, including roe deer, were similar to B. shoenbuchensis, B. chomelii, and B. birtlesi [132].
Roe deer have been shown to be susceptible to hepatitis E virus (HEV), with an overall prevalence of IgG antibodies being 1.20%. Such a low prevalence suggests that roe deer are accidental hosts [133]. However, a later finding revealed that 22.58% of the roe deer samples tested positive for HEV RNA using primers targeting the ORF1 fragment. This underscores the need for further studies to understand HEV transmission between animals and humans in the Baltic countries [134].
3.9. Roe Deer Damage to Forest Stands
The most substantial damage caused by roe deer to forest saplings and undergrowth has been reported in deciduous forests, particularly affecting oak, ash, maple, and, to some extent, spruce. Studies from the 1960s and 1970s indicated that intact oak trees accounted for only 0.5%–2.0% of the total [135]. In addition to the damaged area (Figure 1), it was reported that 36%–39% of trees were damaged in 1969, 32% in 1975, 30% in 1980, and 39% in 1981. Later, the intensity of damages was not presented in the annual reports [41].
In mixed forests, roe deer damage is relatively low, while pure pine forests are generally unaffected. Damage to agriculture is considered insignificant, even in cases where the animals graze in fields throughout the year [11].
In general, roe deer damage is significantly lower compared to that caused by moose or red deer. To mitigate damage to forests, various protective measures have been developed and tested in the country [136]: different methods for maintaining the most valuable tree species, fencing of critical areas, implementation of chemical and biotechnical measures, and control of deer populations.
Despite increasing roe deer numbers, their damage decreased significantly (Figure 6), from 12,114 ha in 1967 to 489 ha in 1997 (CAGR = −0.101). After 1998, deer species are no longer analyzed separately (State Forest Service 2024). It should be noted that, during the same period, damage caused by red deer and moose increased [74,75].
Damage caused by all cervid species reached its maximum in 1990–1992, covering 23–24,000 ha of forest, later gradually decreasing to 2–3000 ha after 2014 (Figure 6). Scientific investigations of roe deer damage in the last 40 years are absent. Sabalinkiene et al. found the negative influence of ungulates on natural forest regeneration and afforestation even under cervid densities not exceeding the highest optimal numbers [137].
3.10. Roe Deer Population Management
Roe deer management in Lithuania is based on regulating their numbers through legal and administrative measures, which underwent several changes during the analyzed period [11,74,75]. The main management method is hunting [16]. However, roe deer are not the main commercial target on hunting grounds [40,138].
During the first two periods (Table 3), roe deer hunting was either forbidden or hunters were not actually interested in increasing the hunting bag. Later, after introducing the so-called “norms” or allowable maximum density of cervids (Table 4), overhunting of roe deer as well as of moose and red deer [74,75] occurred due to concerns about the perceived threat of cervids to forest regeneration and quality.
Institutional and legal changes also influenced roe deer population management. In 1989, wildlife management was transferred to the Ministry of Forestry, leading to a period where the highest proportion of the roe deer population was hunted, particularly during the early years of independence. Norms of allowable density of ungulates, incorporated into legal acts [30,145], mandated hunters to regulate cervid populations. According to these norms, the density of one species of cervids could be increased if the numbers of other species were reduced. For instance, one moose was considered equivalent to three red deer, and one red deer was equivalent to four roe deer. However, the allowable density of roe deer (Table 4) was multiplied by two in forests smaller than 300 ha.
The last three periods were pivotal for the dynamics of roe deer numbers. The implementation of the Law on Hunting in 2002, which allowed the renting of hunting lands for extended periods, enabled hunter organizations to pursue long-term wildlife management strategies [144]. Consequently, the numbers of other deer species besides roe deer also increased [74,75]. The lifting of limits on roe deer hunting and the removal of binding restrictions on selective roe deer hunting, with the only remaining constraint being the length of the hunting season, stabilized roe deer numbers in recent years at approximately 170 thousand.
4. Discussion
We do not dispute the official roe deer numbers, as in many countries, official statistics and bag records are the primary sources of data [16,17,26]. According to Burbaitė and Csányi, the actual roe deer population size in Europe may be around 1.5 times larger than the official data suggest [14]. In the 1980s and 1990s, the real number of roe deer in Lithuania could have been 1.3–1.6 times higher than officially reported, depending on hunters’ reluctance to hand over hunted animals to the state [11]. While roe deer hunting was restricted, from the 1990s onwards, the reported number of roe deer may have been increased to achieve a higher limit. However, these expert opinions have not been thoroughly verified at the country level.
The variation in roe deer population density across Europe is considerable, ranging from 11 to 5380 individuals per 100 km2 [146]. In Lithuania, the density falls towards the lower end of this spectrum, averaging around 347 individuals per 100 km2, with observed limits between 100 and 886 individuals per 100 km2 of the forest area [11]. Specifically, in the continental pine forests of Lithuania, roe deer densities are comparatively low, ranging from 30 to 100 individuals per 100 km2 [93], while in the mixed forests of southern Lithuania, densities are higher, averaging about 150–160 individuals per 100 km2 [40].
There are no publications on the density of the population in the country, taking into account the changes in the area of the country between 1923 and 1945. In 1937, the total forest area in Lithuania was 9064.82 km2, while the country area in 1939 was 55,265.82 km2 (https://osp.stat.gov.lt/statistiniu-rodikliu-analize#/, accessed on 12 January 2024). After 1945, the area of Lithuania remained stable at 65,300 km2. Thus, in 1937–1939, the density of roe deer was 48–63 ind. per 100 km2 of the country, or 295–386 ind. per 100 km2 of the forest area. After the minimum densities found in 1948, 13 ind. per 100 km2 of the country, or 66 ind. per 100 km2 of the forest area, later roe deer densities increased. In 1961, these indices were 46 and 195; in 1983, 60 and 214; in 2002, 106 and 345; in 2013, 181 and 555, respectively. Maximum densities reached 276 ind. per 100 km2 of the country, or 837 ind. per 100 km2 of the forest area in 2020. Therefore, roe deer has consistently been the most abundant cervid in the country [74,75], outnumbering moose 8–12 times and red deer 2–4 times in 2012–2022. In 2017, the size of the roe deer bag in Lithuania was second to Poland, with 26,592 individuals harvested, surpassing Latvia and Estonia, which reported 22,135 and 6264 individuals, respectively [147]. Therefore, we can conclude that the roe deer population in Lithuania is robust.
Studies conducted in eastern Poland, where field roe deer populations are abundant, much like in the southwestern part of Lithuania, have revealed that these populations are particularly vulnerable to poaching, which is identified as the primary source of mortality [148]. Similarly, in the intensively managed cultural landscape of the Czech Republic, predation by red foxes (Vulpes vulpes) and the use of harvesting machines have been identified as important factors contributing to fawn mortality [149]. Although poaching specifically targeting field roe deer was not mentioned as a specific threat [98], guidance notes on the protection of game fauna during agricultural operations were issued in Lithuania [150].
One of the distinctive features of ungulate population management in the country is the selective hunting of ungulate males, a practice that has been in place for over 40 years [151,152,153]. Selective hunting and the criteria for selecting deer were established based on the age variability of cervid antlers [154], and were later formalized into legal requirements [145], including the mandate to pass an exam for permission to hunt adult animals [155]. The requirement for selective hunting was only lifted in 2021 (see Table 3).
In the case of roe deer, a similar strategy of enhancing the value of trophy morphometric parameters was not employed in Latvia or Estonia, the other Baltic countries with similar game fauna and environments [156]. As a result, the largest roe deer trophies are found in Lithuania, and the trophy values of younger roe deer bucks in Lithuania are comparable to the best mature individuals in Estonia and Latvia, despite the lower availability of good quality food.
One potential direction for further research on roe deer ecology and population management could be to assess the relationship between the abundance of several deer species, which has only been considered in terms of permissible densities in Lithuania thus far. A study conducted in northern Poland suggests that red deer may outcompete roe deer, and high densities of red deer could have a negative impact on roe deer numbers and spatial distribution [157]. While red deer numbers in Lithuania are increasing [75], this relationship has not yet been analyzed at various spatial scales.
5. Conclusions
The review concludes that the methodologies used in the studies analyzed had a number of shortcomings, such as:
- The survey of roe deer numbers may not provide an accurate picture, even with broadly coordinated snow or visual surveys, due to potential overlap in roe deer herds between surveyed areas.
- Determining habitat preferences from pellet counts may be biased by differences in pellet decomposition rates and detectability in open and forest habitats.
- Attribution of deer damage to specific deer species lacks sufficient documentation in the cited sources. Methods such as animal tracks and feces in damaged stands, as well as measuring the thickness of browsed shoots, still leave space for bias due to the presence and numbers of different deer species in the area.
- The review highlights the scarcity of research on roe deer biology and ecology in recent decades, except in areas related to road ecology and pathogens.
Funding
This research was conducted under long-term research program of Nature Research Centre with no external funding.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
We thank Jolanta Urbelionytė, from the Ministry of the Environment of the Republic of Lithuania, and Virgilijus Vasiliauskas, from the State Forest Service, for their help with roe deer survey and damage data. We acknowledge the help of Deivis Kulvietis and Artūras Kajėnas from the Lithuanian Police Traffic Supervision Service for their help with deer roadkill. Andrius Kučas provided maps of roe deer roadkill.
Conflicts of Interest
The author declares no conflicts of interest.
References
- Andersen, R.; Duncan, P.; Linnell, J.D. (Eds.) The European Roe Deer: The Biology of Success; Scandinavian University Press: Oslo, Norway, 1998; pp. 1–376. [Google Scholar]
- Wilson, D.E.; Reeder, D.M. (Eds.) Mammal Species of the World: A Taxonomic and Geographic Reference, 3rd ed.; JHU Press: Baltimore, MD, USA, 2005; pp. 1–2147. [Google Scholar]
- Lorenzini, R.; Hewison, M.; Gaillard, J.M.; Garofalo, L.; Rossi, L.; Morellet, N.; Verheyden, H.; Lovari, S.; Lister, A.M.; Mattioli, S. European roe deer Capreolus capreolus (Linnaeus, 1758). In Handbook of the Mammals of Europe; Springer International Publishing: Cham, Switzerland, 2022; pp. 165–195. [Google Scholar]
- Sommer, R.S.; Fahlke, J.M.; Schmölcke, U.; Benecke, N.; Zachos, F.E. Quaternary history of the European roe deer Capreolus capreolus. Mamm. Rev. 2009, 39, 1–16. [Google Scholar] [CrossRef]
- Randi, E.; Alves, P.C.; Carranza, J.; Milošević-Zlatanović, S.; Sfougaris, A.; Mucci, N. Phylogeography of roe deer (Capreolus capreolus) populations: The effects of historical genetic subdivisions and recent nonequilibrium dynamics. Mol. Ecol. 2004, 13, 3071–3083. [Google Scholar] [CrossRef] [PubMed]
- Lorenzini, R.; Lovari, S. Genetic diversity and phylogeography of the European roe deer: The refuge area theory revisited. Biol. J. Linn. Soc. 2006, 88, 85–100. [Google Scholar] [CrossRef]
- Plis, K.; Niedziałkowska, M.; Borowik, T.; Lang, J.; Heddergott, M.; Tiainen, J.; Bunevich, A.; Šprem, N.; Paule, L.; Danilkin, A.; et al. Mitochondrial DNA diversity and the population genetic structure of contemporary roe deer (Capreolus capreolus) in Europe. Mamm. Biol. 2022, 102, 1743–1754. [Google Scholar] [CrossRef]
- Kałuziński, J. Dynamics and structure of a field roe deer population. Acta theriol. 1982, 27, 385–408. [Google Scholar] [CrossRef]
- Pielowski, Z. Some aspects of population structure and longevity of field roe deer. Acta Theriol. 1984, 29, 17–33. [Google Scholar] [CrossRef]
- Putman, R.J. Foraging by roe deer in agricultural areas and impact on arable crops. J. Appl. Ecol. 1986, 23, 91–99. [Google Scholar] [CrossRef]
- Baleišis, R.; Bluzma, P.; Balčiauskas, L. Lietuvos kanopiniai žvėrys [Ungulates of Lithuania], 3rd ed.; Akstis: Vilnius, Lithuania, 2003; pp. 1–217. (In Lithuanian) [Google Scholar]
- Pėtelis, K.; Brazaitis, G. Morphometric data on the field ecotype roe deer in Southwest Lithuania. Acta Zool. Litu. 2003, 13, 61–64. [Google Scholar] [CrossRef]
- Sabalinkiene, G.; Danusevicius, D.; Manton, M.; Brazaitis, G.; Simkevicius, K. Differentiation of European roe deer populations and ecotypes in Lithuania based on DNA markers, cranium and antler morphometry. Silva Fenn. 2017, 51, 1743. [Google Scholar] [CrossRef]
- Burbaitė, L.; Csányi, S. Roe deer population and harvest changes in Europe. Est. J. Ecol. 2009, 58, 169–180. [Google Scholar] [CrossRef]
- Cederlund, G.; Bergqvist, J.; Kjellander, P.; Gill, R.; Gaillard, J.M.; Boisaubert, B.; Ballon, P.; Duncan, P. Managing roe deer and their impact on the environment: Maximising the net benefits to society. In The European Roe Deer: The Biology of Success; Scandinavian University Press: Oslo, Norway, 1998; pp. 337–372. [Google Scholar]
- Apollonio, M.; Andersen, R.; Putman, R. (Eds.) European Ungulates and Their Management in the 21st Century; Cambridge University Press: New York, NY, USA, 2010; pp. 578–604. [Google Scholar]
- Carpio, A.J.; Apollonio, M.; Acevedo, P. Wild ungulate overabundance in Europe: Contexts, causes, monitoring and management recommendations. Mammal Rev. 2021, 51, 95–108. [Google Scholar] [CrossRef]
- Valente, A.M.; Acevedo, P.; Figueiredo, A.M.; Fonseca, C.; Torres, R.T. Overabundant wild ungulate populations in Europe: Management with consideration of socio-ecological consequences. Mammal Rev. 2020, 50, 353–366. [Google Scholar] [CrossRef]
- Balčiauskas, L.; Kučas, A.; Balčiauskienė, L. Mammal Roadkills in Lithuanian Urban Areas: A 15-Year Study. Animals 2023, 13, 3272. [Google Scholar] [CrossRef] [PubMed]
- Partl, E.; Szinovatz, V.; Reimoser, F.; Schweiger-Adler, J. Forest restoration and browsing impact by roe deer. Forest Ecol. Manag. 2002, 159, 87–100. [Google Scholar] [CrossRef]
- Davis, M.L.; Stephens, P.A.; Kjellander, P. Beyond climate envelope projections: Roe deer survival and environmental change. J. Wildl. Manag. 2016, 80, 452–464. [Google Scholar] [CrossRef]
- Maublanc, M.L.; Bideau, E.; Launay, C.; Monthuir, B.; Gerard, J.F. Indicators of ecological change (IEC) as efficient tools for managing roe deer populations: A case study. Eur. J. Wildl. Res. 2016, 62, 189–197. [Google Scholar] [CrossRef]
- Breyne, P.; Mergeay, J.; Casaer, J. Roe deer population structure in a highly fragmented landscape. Eur. J. Wildl. Res. 2014, 60, 909–917. [Google Scholar] [CrossRef]
- Martin, J.; Vourc’h, G.; Bonnot, N.; Cargnelutti, B.; Chaval, Y.; Lourtet, B.; Goulard, M.; Hoch, T.; Plantard, O.; Hewison, A.M.; et al. Temporal shifts in landscape connectivity for an ecosystem engineer, the roe deer, across a multiple-use landscape. Landscape Ecol. 2018, 33, 937–954. [Google Scholar] [CrossRef]
- Bragina, E.V.; Ives, A.R.; Pidgeon, A.M.; Kuemmerle, T.; Baskin, L.M.; Gubar, Y.P.; Piquer-Rodríguez, M.; Keuler, N.S.; Petrosyan, V.G.; Radeloff, V.C. Rapid declines of large mammal populations after the collapse of the Soviet Union. Conserv. Biol. 2015, 29, 844–853. [Google Scholar] [CrossRef] [PubMed]
- Bragina, E.V.; Ives, A.R.; Pidgeon, A.M.; Balčiauskas, L.; Csányi, S.; Khoyetskyy, P.; Kysucká, K.; Lieskovsky, J.; Ozolins, J.; Randveer, T.; et al. Wildlife population changes across Eastern Europe after the collapse of socialism. Front. Ecol. Environ. 2018, 16, 77–81. [Google Scholar] [CrossRef]
- Calkoen, S.T.S.V.; Muhlbauer, L.; Andren, H.; Apollonio, M.; Balčiauskas, L.; Belotti, E.; Carranza, J.; Cottam, J.; Filli, F.; Gatiso, T.T.; et al. Ungulate management in European national parks: Why a more integrated European policy is needed. J. Environ. Manag. 2020, 260, 110068. [Google Scholar] [CrossRef]
- Official Statistics Portal. Forests. Available online: https://osp.stat.gov.lt/lietuvos-aplinka-zemes-ukis-ir-energetika-2021/aplinka/miskai (accessed on 10 January 2024).
- Official Statistics Portal. Lietuvos Miškingumas Pagal Apskritis [Lithuania’s Forest Cover by County]. Available online: https://osp.stat.gov.lt/infografikas-lietuvos-miskingumas-pagal-apskritis (accessed on 10 January 2024).
- LR Aplinkos Ministro 2005 m. Kovo 21 d. Įsakymas nr. D1-162. Dėl Medžioklėtvarkos Projektų Rengimo, Derinimo, Teikimo Tvirtinti ir Tvirtinimo Tvarkos Aprašo Patvirtinimo. [Regarding the Preparation, Coordination, Submission for Approval of Hunting Regulation Projects and Approval of the Description of the Approval Procedure]. Available online: https://e-seimas.lrs.lt/portal/legalAct/lt/TAD/TAIS.252979?jfwid=9iufxps7h (accessed on 10 January 2024).
- Lietuvos Respublikos Aplinkos Ministras Įsakymas dėl Lietuvos Respublikos Aplinkos Ministro 2000 m Birželio 27, D. Įsakymo nr. 258 “Dėl Medžioklės Lietuvos Respublikos Teritorijoje Taisyklių Patvirtinimo” Pakeitimo 2018 m. Spalio 18 d. Nr. D1-892 [Regarding Approval of Rules for Hunting in the Territory of the Republic of Lithuania]. Available online: https://e-seimas.lrs.lt/portal/legalAct/lt/TAD/38fc75e0d2d011e8a82fc67610e51066 (accessed on 10 January 2024).
- Lietuvos Respublikos Aplinkos Ministras. 2018 m. Gegužės 7 d. Įsakymas Nr. D1-365 “Dėl Lietuvos Respublikos Aplinkos Ministro 2000 m. Birželio 27 d. Įsakymo Nr. 258 “Dėl Medžioklės Lietuvos Respublikos Teritorijoje Taisyklių Patvirtinimo” Pakeitimo”. [Change Regarding Approval of Rules for Hunting in the Territory of the Republic of Lithuania]. Available online: https://www.e-tar.lt/portal/legalAct.html?documentId=9379426051dd11e884cbc4327e55f3ca (accessed on 10 January 2024).
- Lietuvos Respublikos Aplinkos Ministras. 2020 m. Gegužės 8 d. Įsakymas Nr. D1-268 “Dėl Lietuvos Respublikos Aplinkos Ministro 2000 m. Birželio 27 d. Įsakymo Nr. 258 “Dėl Medžioklės Lietuvos Respublikos Teritorijoje Taisyklių Patvirtinimo” pakeitimo”. [Change Regarding Approval of Rules for Hunting in the Territory of the Republic of Lithuania]. Available online: https://www.e-tar.lt/portal/legalAct.html?documentId=c4cf1e70910a11ea9515f752ff221ec9 (accessed on 10 January 2024).
- Lietuvos Respublikos Aplinkos Ministras. 2020 m. Spalio 14 d. Įsakymas Nr. D1-631 “Dėl Medžioklės Lietuvos Respublikos Teritorijoje Taisyklių Patvirtinimo” [Regarding the Approval of Hunting Rules in the Territory of the Republic of Lithuania]. Available online: https://www.e-tar.lt/portal/legalAct.html?documentId=348629a00e1211ebb74de75171d26d52 (accessed on 11 February 2024).
- Medžioklės Kalendorius 2024 m. Available online: https://medziokleskalendorius.lt/ (accessed on 10 January 2024).
- Medžiojamųjų Žvėrių Apskaita [Game Survey]. Available online: https://am.lrv.lt/lt/veiklos-sritys-1/gamtos-apsauga/medziokle/medziojamuju-zveriu-apskaita (accessed on 1 February 2024).
- Sumedžioti Žvėrys ir Paukščiai [Hunted Animals and Birds]. Available online: https://am.lrv.lt/lt/veiklos-sritys-1/gamtos-apsauga/medziokle/sumedzioti-zverys-ir-pauksciai/ (accessed on 1 February 2024).
- Medžiojamųjų Žvėrių Apskaita Pagal Pėdsakus Sniege [Game Snow-Track Counts]. Available online: https://vstt.lrv.lt/lt/veiklos-sritys/biologines-ivairoves-apsauga/zveriu-apskaita/medziojamuju-zveriu-apskaita-pagal-pedsakus-sniege/ (accessed on 1 February 2024).
- Kawata, Y.; Ozolins, J.; Andersone-Lilley, Z. An analysis of the game animal population data from Latvia. Baltic For. 2008, 14, 75–86. [Google Scholar]
- Barkauskas, T.; Belova, O.; Griciuvienė, L. Differences in ungulate population use in different hunting ground units in Lithuania. Baltic For. 2020, 26, 514. [Google Scholar] [CrossRef]
- Valstybinė Miškų Tarnyba. Miškų Sanitarinė Būklė [Sanitary Condition of Forests]. Available online: https://amvmt.lrv.lt/lt/veiklos-sritys/misko-sanitarine-apsauga/misku-sanitarine-bukle (accessed on 5 January 2024).
- Compound Annual Growth Rate—CAGR. Available online: https://www.investopedia.com/terms/c/cagr.asp#what-is-compound-annual-growth-rate--cagr (accessed on 20 December 2023).
- TIBCO Software Inc. Data Science Textbook. Available online: https://docs.tibco.com/data-science/textbook (accessed on 15 January 2024).
- Sommer, R.S.; Zachos, F.E. Fossil evidence and phylogeography of temperate species: ‘glacial refugia’ and post-glacial recolonization. J. Biogeogr. 2009, 36, 2013–2020. [Google Scholar] [CrossRef]
- Niedziałkowska, M.; Jędrzejewska, B.; Honnen, A.C.; Otto, T.; Sidorovich, V.E.; Perzanowski, K.; Skog, A.; Hartl, G.B.; Borowik, T.; Bunevich, A.N.; et al. Molecular biogeography of red deer Cervus elaphus from eastern Europe: Insights from mitochondrial DNA sequences. Acta Theriol. 2011, 56, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Grünberg, J.M. Animals in Mesolithic burials in Europe. Anthropozoologica 2013, 48, 231–253. [Google Scholar] [CrossRef]
- Rimkus, T. In search of Lithuania coastal Mesolithic: Review of current data and the aims of an ongoing research project. Fornvännen 2019, 114, 1–11. [Google Scholar]
- Daugnora, L.; Girininkas, A. Osteoarcheologija Lietuvoje; Savastis: Vilnius, Lithuania, 1996; pp. 1–198. [Google Scholar]
- Simčenka, E.; Kozakaitė, J.; Piličiauskienė, G.; Gaižauskas, L.; Piličiauskas, G. Human Diet During the Stone Age and Early Metal Period (7000–1 cal BC) in Lithuania: An update. Radiocarbon 2022, 64, 1171–1189. [Google Scholar] [CrossRef]
- Girininkas, A.; Daugnora, L. Hunting in the territory of Lithuania from the Late Palaeolithic to the Middle Ages. In Hunting in Northern Europe until 1500 AD: Old Traditions and Regional Developments, Continental Sources and Continental Influences; Grimm, O., Schmölcke, U., Eds.; Neumünster: Wachholtz, Germany, 2013; pp. 567–595. [Google Scholar]
- Vitkūnas, M. Medžioklė XIII–XIV a. (tyrinėjimų Pietryčių Lietuvoje duomenimis) [Hunting in the 13th–14th Centuries (on the research data in Southeastern Lithuania)]. Istorija. Liet. Aukštųjų Mokykl. Moksl. Darb. 2009, 76, 3–17. (In Lithuanian) [Google Scholar]
- Volkaitė-Kulikauskienė, R. Lietuviai IX–XII Amžiais; Mintis: Vilnius, Lietuva, 1970; pp. 59–60. [Google Scholar]
- Daugnora, L.; Piličiauskienė, G. XIV–XVII a. osteologinės medžiagos, 1988–1990 m. iškastos Vilniaus žemutinės pilies teritorijoje, analizė [The analysis of the osteological material of the 14th–17th c. excavated in 1988–1990 in the territory of the Vilnius Lower Castle]. Liet. Archeol. 2005, 28, 207–216. (In Lithuanian) [Google Scholar]
- Rzączyñski, G. Historia Naturalis Curiosa Regni Poloniae, Magni Ducatus Lithvaniae; Typis Collegii Soc. Jesu: Sandomiria, Poland, 1721; pp. 1–489. [Google Scholar]
- Ładowski, S. Hystoria Naturalna Królestwa Polskiego; W Drukarni Uprzywilejowaney Ignacego Grebla: Kraków, Poland, 1783; pp. 1–206. [Google Scholar]
- Eichwald, E. Naturhistorische Skizze von Lithauen, Volhynien und Podolien; Zawadzki: Wilna, Lithuania, 1830; pp. 1–256. [Google Scholar]
- Paaver, K.L. Formirovanie Teriofauny i Izmenchivost’mlekopitayushchikh Pribaltiki v Golotsene (Formation of the Theriofauna and the Variability of Baltic Mammals in the Holocene); Akad. Nauk ESSR: Tartu, Estonia, 1965; pp. 1–494. [Google Scholar]
- Heptner, V.G.; Nasimovič, A.A.; Bannikov, A.G.E. Mammals of the Soviet Union: Volume 1, Ungulates; Model Press: New Delhi, India, 1989; pp. 243–301. [Google Scholar]
- Prūsaitė, J. (Ed.) . Lietuvos Fauna. Žinduoliai [Fauna of Lithuania. Mammals]; Mokslas: Vilnius, Lithuania, 1988; pp. 214–218. (In Lithuanian) [Google Scholar]
- Plater, A. Spis Zwierząt Ssących, Ptaków i Ryb Krajowych; Zawadzki: Wilno, Lithuania, 1852; pp. 1–160. [Google Scholar]
- Ivanauskas, T. Mūsų medžiojamieji gyvuliai. Stirna. Mūsų rytojus 1928, 6. Available online: https://dokumen.pub/tadas-ivanauskas-gyvenimas-ir-veikla.html (accessed on 15 January 2024).
- Prusaite, J.; Blazys, A.; Mickus, A.; Bluzma, P. Number dynamics and structure of the unhunted roe deer population. Lith. Acad. Sci. Ser. C 1973, 62, 115–125. [Google Scholar]
- Prusaite, J.A.; Blazys, A.S.; Baleisis, R.M. Intensity of breeding and fecundity of the European roe deer in North Lithuania. Liet. TSR Moksl. Akad. Darb. Ser. C 1977, 3, 105–110. [Google Scholar]
- Bluzma, P. Estimation of the state of lynx and wolf populations in Lithuania. Acta Zool. Litu. 1999, 9, 35–41. [Google Scholar] [CrossRef]
- Bluzma, P. Lietuvos medžiojamieji žinduoliai: Populiacijų dinamika ir dabartinė būklė. Theriol. Litu. 2001, 1, 12–21. [Google Scholar]
- Bluzma, P.; Baleišis, R. The cervid (Cervidae) monitoring in Lithuania (1993–1998): Abundance, dynamics and distribution. Acta Zool. Litu. 1999, 9, 61–70. [Google Scholar] [CrossRef]
- Balčiauskas, L.; Kučas, A.; Balčiauskienė, L. The Impact of Roadkill on Cervid Populations in Lithuania. Forests 2023, 14, 1224. [Google Scholar] [CrossRef]
- Balčiauskas, L.; Stratford, J.; Kučas, A.; Balčiauskienė, L. Lockdown’s Silver Lining? Different Levels of Roadkill during the COVID-19 Times in Lithuania. Animals 2023, 13, 2918. [Google Scholar] [CrossRef]
- Perkins, S.E.; Shilling, F.; Collinson, W. Anthropause Opportunities: Experimental Perturbation of Road Traffic and the Potential Effects on Wildlife. Front. Ecol. Evol. 2022, 10, 833129. [Google Scholar] [CrossRef]
- Balčiauskas, L.; Stratford, J.; Balčiauskienė, L.; Kučas, A. Importance of professional roadkill data in assessing diversity of mammal roadkills. Transp. Res. D Transp. Environ. 2020, 87, 102493. [Google Scholar] [CrossRef]
- Narauskaitė, G.; Pėtelis, K.; Maksvytis, M. Šilutė region seacoast roe deer Capreolus capreolus L. population quality. Acta Biol. Univ. Daugavp. 2011, 11, 29–34. [Google Scholar]
- Narauskaitė, G.; Danusevičius, D.; Østbye, K.; Pėtelis, K. Genetic Differentiation of Field and Forest Ecotypes of Roe Deer (Capreolus capreolus L.) in Lithuania Based on DNA Markers. Rural. Dev. 2013, 6, 376–381. [Google Scholar]
- Balčiauskas, L. The influence of roadkill on protected species and other wildlife in Lithuania. In Proceedings of the 2011 International Conference on Ecology and Transportation (ICOET 2011), Seattle, WA, USA, 21–25 August 2011; pp. 647–655. Available online: https://trid.trb.org/view/1347982 (accessed on 20 December 2023).
- Balčiauskas, L.; Kawata, Y.; Balčiauskienė, L. Moose Management Strategies under Changing Legal and Institutional Frameworks. Sustainability 2020, 12, 8482. [Google Scholar] [CrossRef]
- Balčiauskas, L.; Kawata, Y. Red Deer in Lithuania: History, Status and Management. Sustainability 2022, 14, 14091. [Google Scholar] [CrossRef]
- Balčiauskas, L.; Jasiulionis, M. Reducing the incidence of mammals on public highways using chemical repellent. Balt. J. Road Bridge E 2012, 7, 92–97. [Google Scholar] [CrossRef]
- Ignatavicius, G.; Valskys, V. The influence of time factors on the dynamics of roe deer collisions with vehicles. Landsc. Ecol. Eng. 2018, 14, 221–229. [Google Scholar] [CrossRef]
- Kučas, A.; Balčiauskas, L. Temporal patterns of ungulate-vehicle collisions in Lithuania. J. Environ. Manag. 2020, 273, 111172. [Google Scholar] [CrossRef] [PubMed]
- Galinskaitė, L.; Ignatavičius, G. Dynamics of Roe Deer (Capreolus capreolus) Vehicle Collisions in Lithuania: Influence of the Time Factors. Int. J. Transp. Veh. Eng. 2022, 16, 108–112. [Google Scholar]
- Balčiauskas, L.; Kučas, A.; Balčiauskienė, L. Roadkill Patterns on Workdays, Weekends and Long Weekends: Anticipating the Implications of a Four-Day Work Week. Diversity 2024, 16, 84. [Google Scholar] [CrossRef]
- Balčiauskas, L.; Wierzchowski, J.; Kučas, A.; Balčiauskienė, L. Habitat Suitability Based Models for Ungulate Roadkill Prognosis. Animals 2020, 10, 1345. [Google Scholar] [CrossRef] [PubMed]
- Galinskaitė, L.; Ulevičius, A.; Valskys, V.; Samas, A.; Busher, P.E.; Ignatavičius, G. The Influence of Landscape Structure on Wildlife–Vehicle Collisions: Geostatistical Analysis on Hot Spot and Habitat Proximity Relations. ISPRS Int. J. Geo-Inf. 2022, 11, 63. [Google Scholar] [CrossRef]
- Kučas, A.; Balčiauskas, L. Roadkill-Data-Based Identification and Ranking of Mammal Habitats. Land 2021, 10, 477. [Google Scholar] [CrossRef]
- Kučas, A.; Balčiauskas, L. Impact of Road Fencing on Ungulate-Vehicle Collisions and Hotspot Patterns. Land 2021, 10, 338. [Google Scholar] [CrossRef]
- Kučas, A.; Balčiauskas, L.; Lavalle, C. Identification of Urban and Wildlife Terrestrial Corridor Intersections for Planning of Wildlife-Vehicle Collision Mitigation Measures. Land 2023, 12, 758. [Google Scholar] [CrossRef]
- Hagen, R.; Heurich, M.; Storch, I.; Hanewinkel, M.; Kramer-Schadt, S. Linking annual variations of roe deer bag records to large-scale winter conditions: Spatio-temporal development in Europe between 1961 and 2013. Eur. J. Wildl. Res. 2017, 63, 97. [Google Scholar] [CrossRef]
- Marma, B.; Padaiga, V. Stirnų mitybos fiziologijos klausimu. Girios 1975, 5, 24–25. [Google Scholar]
- Bluzma, P. Lietuvos Stirnos. Mūsų Gamt. 1976, 7–9, 12. [Google Scholar]
- Balčiauskas, L. Антрoпoгеннoсть ландшафта Литвы как фактoр высoкoй численнoсти кoпытных [Anthropogenicity of the landscape of Lithuania as a factor of high abundance of ungulates]. Весци Ан Бсср Сер. Биoл. 1988, 2, 105–107. (In Russian) [Google Scholar]
- Balčiauskas, L. Сеть трoп кoпытных как oтражение перемещений живoтных и испoльзoвания ими oбжитoй территoрии [Network of the paths of ungulates as a reflection of the animal movements and use of the inhabited territory]. In Млекoпитающие в Культурнoм Ландшафте Литвы; Ekologijos Institutas: Vilnius, Lithuania, 1990; pp. 131–137. (In Russian) [Google Scholar]
- Marma, B. Stirnų kritimų priežastys ir pasekmės. Girios 1970, 9, 17–19. [Google Scholar]
- Падаига, В. Фактoры, oпределяющие зимние падежы кoсуль. Зooлoгичеслий Журнал 1971, 10, 1546–1552. [Google Scholar]
- Belova, O. The impact of herbivorous mammals on woody vegetation in the different stages of forest succession. Baltic For. 2012, 18, 100–110. [Google Scholar]
- Padaiga, V. Stirnos ir kietieji lapuočiai. Girios 1967, 1, 10–11. [Google Scholar]
- Балейшис, Р.М.; Прусайте, Я.А. Питание еврoпейскoй кoсули в небoльшoм лиственнoм лесу Севернoй Литвы в 1975–1976 г.г. Труды Академии Наук Литoвскoй Сср Серия В 1980, 1, 85–91. [Google Scholar]
- Прусаите, Я.; Балеишис, Р.; Блузма, П. Сoстав кoрмв еврoпейскoй кoсули в зависимoсти oт лесистoсти oбитаемoй теритoрии. Труды Академии Наук Литoвскoй Сср. Серия В 1983, 4, 84–98. [Google Scholar]
- Блузма, П.П.; Балейшис, Р.M. Значение агрoценoзoв для кoсули в Литве. In Охoтoустрoйствo в Специализирoваннoм Леснoм Хoзяйстве; LMŪMTI: Kaunas-Girionys, Lithuania, 1983; pp. 64–66. [Google Scholar]
- Блузма, П.; Балейшис, Р. Еврoпейская кoсуля в агрoценoзах Литвы. In Млекoпитающие в культурнoм ландшафте Литвы; Mokslas: Vilnius, Lithuania, 1990; pp. 138–145. [Google Scholar]
- Tauginas, J. Žmogaus ūkinės veiklos miške įtaka elninių žvėrių gausumui [Impact of human economic activities in the forest on the abundance of cervids. Girios 1977, 4, 15–16. (In Lithuanian) [Google Scholar]
- Priedytis, A.; Tauginas, J. Feeding effectiveness of granules, made from summer cutting waste of aspen, in the winter ration of roe deer. Liet. TSR Moksl. Akad. Darbai. Ser. C 1981, 3, 149–158. [Google Scholar]
- Bluzma, P.; Baleišis, R.; Balčiauskas, L. Численнoсть и рaспределение oленьих (Cervidae) нa зимних пaстбищaх рaзличных лaндшaфтoв Литвы [Abundance and distribution of cervids in winter stands of various landscapes of Lithuania]. In Млекoпитaющие B Культурнoм Лaндшaфте Литвы; Ekologijos Institutas: Vilnius, Lithuania, 1990; pp. 122–130. (In Russian) [Google Scholar]
- Bluzma, P.; Baleisis, R.; Balciauskas, L. Significance of the stand composition to cervid (Cervidae) winter distribution in the forests of various Lithuanian landscapes. Ekologija 1996, 3, 61–66. [Google Scholar]
- Baleisis, R.; Bluzma, P.; Balciauskas, L. Deers (Cervidae) distribution in winter depending on the amount of dwarf shrubs in the forest areas of different landscapes in Lithuania. Ekologija 1995, 2, 45–48. [Google Scholar]
- Baranauskas, K.; Balčiauskas, L.; Mažeikytė, R. Vilnius city theriofauna. Acta Zool. Litu. 2005, 15, 228–238. [Google Scholar] [CrossRef]
- Brazaitis, G.; Pėtelis, K.; Žalkauskas, R.; Belova, O.; Danusevičius, D.; Marozas, V.; Narauskaitė, G. Landscape effect for the Cervidaes Cervidae in human-dominated fragmented forests. Eur. J. Forest Res. 2014, 133, 857–869. [Google Scholar] [CrossRef]
- Прусайте, Я.А.; Блажис, А.С.; Балейшис, Р.М. Влияние эксплуатции различнoй интенсивнoсти на численнoсть и прирoст пoпуляции еврoпейскoй кoсули. In II съезд Всесoюзнoгo Териoлoгическoгo Общества; USSR: Nauka, Moskva, 1978; pp. 251–252. [Google Scholar]
- Flajšman, K.; Pokorny, B.; Chirichella, R.; Bottero, E.; Mattioli, L.; Apollonio, M. I can produce more offspring as you can imagine: First records on exceptionally large litters in roe deer in central/southern Europe. Eur. J. Wildl. Res. 2017, 63, 42. [Google Scholar] [CrossRef]
- Fruzinski, B.; Kaluzinski, J.; Baksalary, J. Weight and body measurements of forest and field roe deer. Acta Theriol. 1982, 27, 479–488. [Google Scholar] [CrossRef]
- Pėtelis, K. The estimation of trophy maturity on roe deer field ecotype. Miškininkystė 1997, 1, 123–127. [Google Scholar]
- Markov, G.; Zvychaynaya, E.; Danilkin, A.; Kholodova, M.; Sugar, L. Genetic diversity and phylogeography of roe deer (Capreolus capreolus L.) in different biogeographical regions in Europe. CR Acad. Bulg. Sci. 2016, 69, 579–584. [Google Scholar]
- Olano-Marin, J.; Plis, K.; Sönnichsen, L.; Borowik, T.; Niedziałkowska, M.; Jędrzejewska, B. Weak population structure in European roe deer (Capreolus capreolus) and evidence of introgressive hybridization with Siberian roe deer (C. pygargus) in northeastern Poland. PLoS ONE 2014, 9, e109147. [Google Scholar] [CrossRef] [PubMed]
- Plis, K.; Niedziałkowska, M.; Borowik, T.; Lang, J.; Heddergott, M.; Tiainen, J.; Bunevich, A.; Šprem, N.; Paule, L.; Danilkin, A.; et al. Pan-European phylogeography of the European roe deer (Capreolus capreolus). Ecol. Evol. 2022, 12, e8931. [Google Scholar] [CrossRef] [PubMed]
- Niedziałkowska, M.; Plis, K.; Marczuk, B.; Lang, J.; Heddergott, M.; Tiainen, J.; Danilkin, A.; Kholodova, M.; Zvychaynaya, E.; Kashinina, N.; et al. Genetic diversity and complex structure of the European Roe Deer population at a continental scale. J. Mamm. 2024, 105, 73–84. [Google Scholar] [CrossRef]
- Pūraitė, I.; Paulauskas, A.; Sruoga, A. Analysis of genetic diversity of roe deer (Capreolus capreolus L.) in Lithuania using RAPD and allozyme systems. Biologija 2013, 59, 29–38. [Google Scholar] [CrossRef]
- Pūraitė, I.; Paulauskas, A.; Sruoga, A.; Prakas, P. Mitochondrial DNA variation in roe deer population from Lithuania. Balk. J. Wildl. Res. 2014, 1, 13–19. [Google Scholar] [CrossRef]
- Pūraitė, I.; Paulauskas, A.; Sruoga, A. Genetic analysis of roe deer (Capreolus capreolus L.) using DNA markers. Вестник Охoтoведения 2014, 11, 226–230. [Google Scholar]
- González, S.; del Rio, M.L.; Díez-Baños, N.; Martínez, A.; Hidalgo, M.d.R. Contribution to the Knowledge of Gastrointestinal Nematodes in Roe Deer (Capreolus capreolus) from the Province of León, Spain: An Epidemiological and Molecular Study. Animals 2023, 13, 3117. [Google Scholar] [CrossRef] [PubMed]
- Arnastauskienė, T. Coccidia and Coccidioses of Domestic and Wild Animals in Lithuania; Mokslas: Vilnius, Lithuania, 1985; pp. 1–176. [Google Scholar]
- Arnastauskienė, T. Coccidial fauna of roe deer (Capreolus capreolus) in Lithuania. Acta Parasitol. Litu. 1974, 12, 27–37. [Google Scholar]
- Kazlauskas, Y.; Arnastauskene, T. Helminths of wild artiodactyls and hares common to man and domestic animals in Lithuania. Acta Parasitol. Lith. 1969, 9, 61–67. [Google Scholar]
- Падаига, В.; Марма, Б. Зараженoсть кoсуль паразитами в звисимoсти oт плoтнoсти пoпуляции oбитания. Зooлoгичеслий журнал 1970, 9, 283–287. [Google Scholar]
- Shlikas, A.; Kazlauskas, Y. Survival of Trichuris eggs from roe-deer and hares in Lithuania. Acta Parasitol. Litu. 1970, 10, 75–80. [Google Scholar]
- Kazlauskas, J.; Pužauskas, R. On the factors affecting the distribution of roe-deer helminths in Lithuania. Acta Parasitol. Litu. 1974, 12, 87–91. [Google Scholar]
- Kutkienė, L. The species composition of European roe deer (Capreolus capreolus) Sarcocystis in Lithuania. Acta Zool. Litu. 2001, 11, 97–101. [Google Scholar] [CrossRef]
- Malakauskas, M.; Grikienienė, J. Sarcocystis infection in wild ungulates in Lithuania. Acta Zool. Litu. 2002, 12, 372–380. [Google Scholar] [CrossRef]
- Januškevičius, V.; Januškevičienė, G.; Paulauskas, V.; Petkevičius, S. Prevalence and intensity of Sarcosporidia infection in wild cloven-hoofed hunted for food in Lithuania. Vet. Zootec. 2016, 73, 27–31. [Google Scholar]
- Rudaitytė-Lukošienė, E.; Delgado de Las Cuevas, G.E.; Prakas, P.; Calero-Bernal, R.; Martínez-González, M.; Strazdaitė-Žielienė, Ž; Servienė, E.; Habela, M.A.; Butkauskas, D. Sarcocystis spp. diversity in the roe deer (Capreolus capreolus) from Lithuania and Spain. Parasitol. Res. 2020, 119, 1363–1370. [Google Scholar] [CrossRef] [PubMed]
- Prakas, P.; Rudaitytė, E.; Butkauskas, D.; Kutkienė, L. Sarcocystis entzerothi n. sp. from the European roe deer (Capreolus capreolus). Parasitol. Res. 2017, 116, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Ražanskė, I.; Radzijevskaja, J.; Paulauskas, A.; Sruoga, A. Babesia spp. in questing ticks and ticks parasitizing cervids. In The Vital Nature Sign; Vytautas Magnus University: Kaunas, Lithuania, 2013; p. 31. [Google Scholar]
- Paulauskas, A.; Morkutė, K.; Ražanskė, I.; Radzijevskaja, J. Infection of tick-borne pathogens of games in Lithuania. In 5th International Hunting and Game Management Symposium; Kusza, S., Jâvor, A., Eds.; Licium-Art Kft.: Debrecen, Hungary, 2016; pp. 10–12. [Google Scholar]
- Paulauskas, A.; Ražanskė, I.; Radzijevskaja, J.; Olav, R.; Morkutė, K. Prevalence and diversity of Babesia spp. In wild cervids. In German Society for Parasitology: Program and Abstracts; University of Göttingen: Göttingen, Germany, 2016; p. 115. [Google Scholar]
- Vasaitytė, A.; Klepeckienė, K.; Ražanskė, I.; Radzijevskaja, J.; Paulauskas, A. Investigation of Bartonella spp. In domestic and wild animals from Lithuania. In Smart Bio: ICSB 2nd International Conference, Abstracts Book; Vytautas Magnus University: Kaunas, Lithuania, 2018; p. 195. [Google Scholar]
- Spancerniene, U.; Buitkuviene, J.; Grigas, J.; Pampariene, I.; Salomskas, A.; Cepuliene, R.; Žymantienė, J.; Stankevicius, A. Seroprevalence of hepatitis E virus in Lithuanian domestic pigs and wildlife. Acta Vet. Brno 2017, 85, 319–327. [Google Scholar] [CrossRef]
- Spancerniene, U.; Grigas, J.; Buitkuviene, J.; Zymantiene, J.; Juozaitiene, V.; Stankeviciute, M.; Razukevicius, D.; Zienius, D.; Stankevicius, A. Prevalence and phylogenetic analysis of hepatitis E virus in pigs, wild boars, roe deer, red deer and moose in Lithuania. Acta Vet. Scand. 2018, 60, 13. [Google Scholar] [CrossRef] [PubMed]
- Padaiga, V. Miškas ir Elniniai [Forest and Cervids]; (In Lithuanian). Mintis: Vilnius, Lithuania, 1969; pp. 1–122. [Google Scholar]
- Падайга, В.И. Зимние прoстранственнoе распределение oленей в лесных oхoтничьих угoдиях. Тр. Литoвскoгo НИИЛХ 1973, 14, 337–351. [Google Scholar]
- Sabalinkiene, G.; Simkevicius, K.; Petelis, K.; Stankeviciute, J.; Talijunas, D. High densities of Cervidae effect to forest regeneration in mixed broadleaf forest in south part of Lithuania. Agrofor 2016, 1, 95–102. [Google Scholar] [CrossRef]
- Saveljev, A.P.; Lissovsky, A.A.; Kozlov, Y.A. Comparative analysis of the lists of hunting mammals of the countries of the Baltic region and Belarus. Russian J. Theriol. 2020, 19, 65–70. [Google Scholar] [CrossRef]
- Žemulis, F. Medžioklė Tarybiniais Laikais ir Dabar [Hunting in Soviet Times and Now]. 2012. Available online: https://www.delfi.lt/grynas/aplinka/medziokle-tarybiniais-laikais-ir-dabar.d?id=57145619 (accessed on 17 February 2024).
- Padajga, V.I. Methods of Regulating Deer Populations in Intensive Forestry; LitNIILH: Kaunas, Lithuania, 1970; pp. 1–32. [Google Scholar]
- Padaiga, V. Экoлoгические oснoвы управления численнoстью oленьих в Литoвскoй ССР [Ecological Fundamentals for the Management of Cervid Number in the Lithuanian SSR.] Habil. Ph.D. Thesis, University of Tartu, Tartu, Estonia, 1984. [Google Scholar]
- Ribikauskas, V. Medžioklė yra Medžioklė. Available online: https://www.manoukis.lt/mano-ukis-zurnalas/2014/02/medziokle-yra-medziokle/ (accessed on 17 February 2024).
- Balciauskas, L. Game survey as a mean of sustainable population management. In Proceedings of the International Symposium “Rational Management of Cervids in Forest Habitats”, Šiauliai, Lithuania, 28–30 January 2004. [Google Scholar]
- Lietuvos Respublikos Medžioklės Įstatymas [Hunting Law of the Republic of Lithuania]. Available online: https://e-seimas.lrs.lt/portal/legalAct/lt/TAD/TAIS.169653/asr (accessed on 15 February 2024).
- Padaiga, V. Medžioklės Ūkio Biologiniai Pagrindai; Žiburys: Vilnius, Lithuania, 1996; pp. 1–210. [Google Scholar]
- Melis, C.; Jędrzejewska, B.; Apollonio, M.; Bartoń, K.A.; Jędrzejewski, W.; Linnell, J.D.; Kojola, I.; Kusak, J.; Adamic, M.; Ciuti, S.; et al. Predation has a greater impact in less productive environments: Variation in roe deer, Capreolus capreolus, population density across Europe. Global Ecol. Biogeogr. 2009, 18, 724–734. [Google Scholar] [CrossRef]
- Linnell, J.D.; Cretois, B.; Nilsen, E.B.; Rolandsen, C.M.; Solberg, E.J.; Veiberg, V.; Kaczensky, P.; Van Moorter, B.; Panzacchi, M.; Rauset, G.R.; et al. The challenges and opportunities of coexisting with wild ungulates in the human-dominated landscapes of Europe’s Anthropocene. Biol. Conserv. 2020, 244, 108500. [Google Scholar] [CrossRef]
- Sönnichsen, L.; Borowik, T.; Podgórski, T.; Plis, K.; Berger, A.; Jędrzejewska, B. Survival rates and causes of mortality of roe deer Capreolus capreolus in a rural landscape, eastern Poland. Mamm. Res. 2017, 62, 141–147. [Google Scholar] [CrossRef]
- Cukor, J.; Havránek, F.; Vacek, Z.; Bukovjan, K.; Podrázský, V.; Sharma, R.P. Roe deer (Capreolus capreolus) mortality in relation to fodder harvest in agricultural landscape. Mammalia 2019, 83, 461–469. [Google Scholar] [CrossRef]
- Baleišis, R.; Bluzma, P.; Prūsaitė, J.; Vėžys, P. Medžiojamosios Faunos Apsauga Žemės Ūkio Darbų Metu (Metodiniai Nurodymai); Lietuvos TSR Medžiotojų ir Žvejų Draugija: Vilnius, Lithuania, 1980; pp. 1–16. [Google Scholar]
- Zažeckis, V.; Baleišis, R. Kanopinių Žvėrių Atrankinė Medžioklė (Metodiniai Nurodymai); Gamtos Apsaugos Komitetas Prie Lietuvos TSR Ministrų Tarybos: Vilnius, Lithuania, 1975; pp. 1–14. [Google Scholar]
- Baleišis, R.; Bluzma, P.; Matuzevičius, A.; Zažeckis, V. Elninių Žvėrių Selekcinė Medžioklė (Metodiniai Nurodymai); Lietuvos TSR Medžiotojų ir Žvejų Draugija: Vilnius, Lithuania, 1979; pp. 1–36. [Google Scholar]
- Балейшис, Р.М.; Блузма, П.П. Результаты селекциoннoгo oтстрела oленъих (Cervidae) в Литoвскoй ССР. In Териoлoгические Исследoвания в Литве; Mokslas: Vilnius, Lietuva, 1988; pp. 84–89. [Google Scholar]
- Балейшис, Р.М.; Блузма, П.П. Вoзрастная изменчивoсть рoгoв кoсули в Литве. Бюл. Мoск. О-Ва Испытателей Прирoды. Отд. Биoл. 1981, 86, 3–10. [Google Scholar]
- Medžiotojų Selekcininkų Kvalifikacijos Suteikimo Tvarka. 2008. Available online: https://www.e-tar.lt/portal/lt/legalAct/TAR.A053608065EC (accessed on 1 February 2016).
- Balčiauskas, L.; Varanauskas, R.; Bukelskis, E. Impact of selective hunting on the trop hy size of roe deer: Baltic example. North-West. J. Zool. 2017, 13, 118–127. [Google Scholar]
- Borkowski, J.; Banul, R.; Jurkiewicz-Azab, J.; Hołdyński, C.; Święczkowska, J.; Nasiadko, M.; Załuski, D. There is only one winner: The negative impact of red deer density on roe deer numbers and distribution in the Słowiński National Park and its vicinity. Ecol. Evol. 2021, 11, 6889–6899. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Map of Lithuania showing forests, cities, and main roads in 2021. The inset indicates the position of the country in Europe. Note: Between 1934 and 1940, the territorial boundaries of the country were altered in the western and southeastern regions due to political factors.
Figure 1.
Map of Lithuania showing forests, cities, and main roads in 2021. The inset indicates the position of the country in Europe. Note: Between 1934 and 1940, the territorial boundaries of the country were altered in the western and southeastern regions due to political factors.

Figure 2.
Changes of the roe deer numbers in Lithuania, 1934–2023, based on [11,36,59]. Data for 1940–1942 and 1944–1946 are not available, while data for 1943 are not reliable.


Figure 4.
Proportion of the roe deer hunting bag from the population numbers in Lithuania, 1965–2022. Based on [11,36,37,59].

Figure 5.
Roe deer roadkill in respect to forest habitat, 2002–2022 [67]. One dot represents one roadkill.
Figure 5.
Roe deer roadkill in respect to forest habitat, 2002–2022 [67]. One dot represents one roadkill.

Figure 6.
Dynamics of roe deer-induced forest damage in Lithuania from 1967 to 1997, measured by the extent of damaged areas (ha) [41].
Figure 6.
Dynamics of roe deer-induced forest damage in Lithuania from 1967 to 1997, measured by the extent of damaged areas (ha) [41].


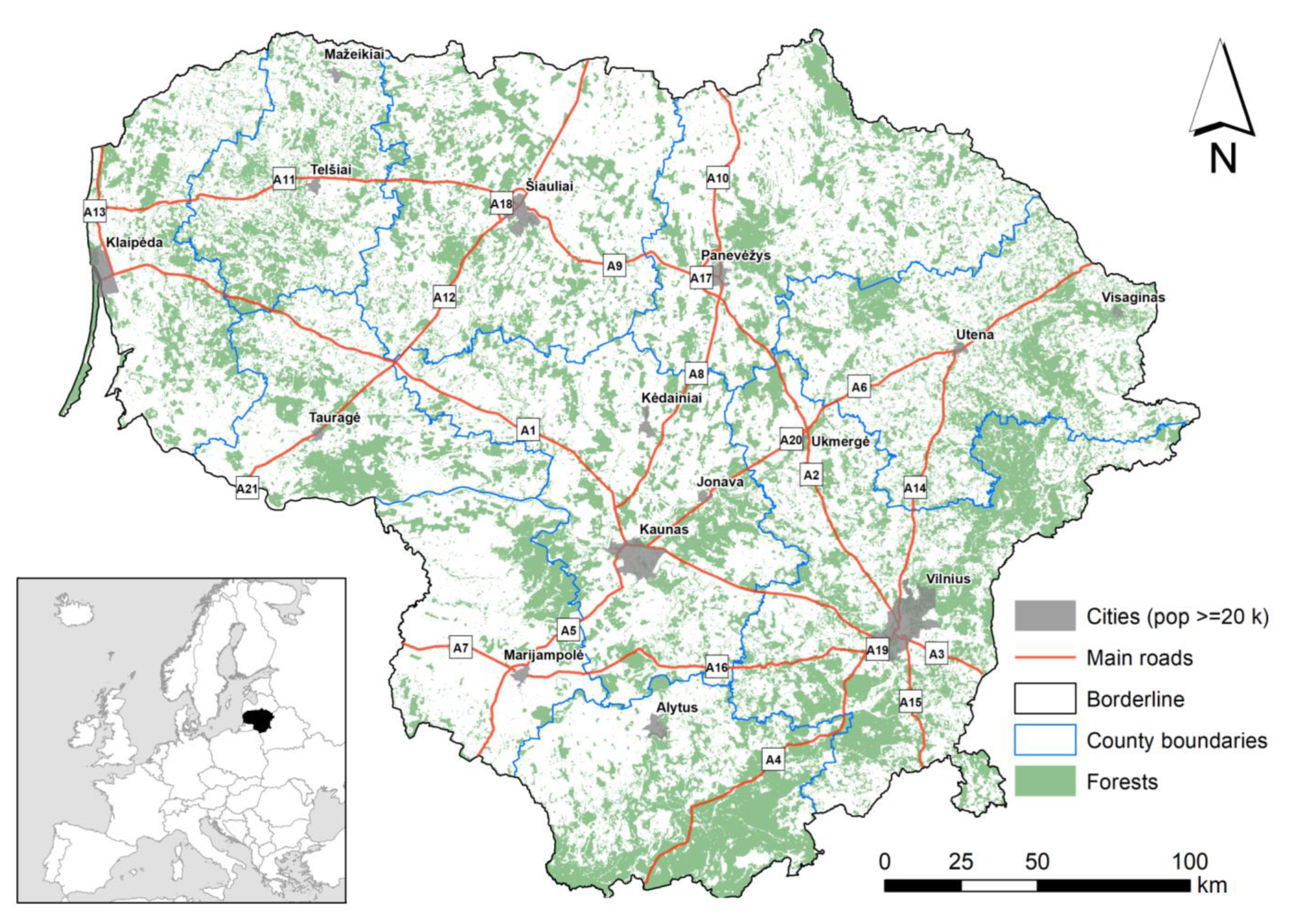{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Key terms used in the literature search.
| Key Term | Definition | Notes on Usage |
|---|---|---|
| Roe deer | Capreolus capreolus, member of the Cervidae family | We also used “stirna” (Lithuanian) and “Evropejskaja kosulia” (Russian) |
| Lithuania | Territorial scope of the search | We also used “Lietuva”, “Baltic countries” and “Baltic states”, as well as “Baltijos šalys” |
| History | Period between last glaciation and world War II | Includes archaeological findings and archives |
| Management | Includes hunting, survey, and monitoring | The hunting yield and the abundance of species were associated with alterations in the legal framework |
Table 2.
Roe deer roadkill on the main, national, regional, and urban roads in 2002–2022, and roe deer proportion of the roadkilled mammals: TM, N—the total number of roadkilled mammals; RD main—number of roadkilled roe deer on the main, RD nat—national, RD reg—regional, RD urb—urban roads; RD, n—total of roadkilled roe deer; RD, %—percent of roadkilled roe deer from all roadkilled mammals, RDF, %—percent of roe deer roadkilled in the forest habitat.
Table 2.
Roe deer roadkill on the main, national, regional, and urban roads in 2002–2022, and roe deer proportion of the roadkilled mammals: TM, N—the total number of roadkilled mammals; RD main—number of roadkilled roe deer on the main, RD nat—national, RD reg—regional, RD urb—urban roads; RD, n—total of roadkilled roe deer; RD, %—percent of roadkilled roe deer from all roadkilled mammals, RDF, %—percent of roe deer roadkilled in the forest habitat.
| Year | TM, N | RD Main | RD Nat | RD Reg | RD Urb | RD, n | RD, % | RDF, % |
|---|---|---|---|---|---|---|---|---|
| 2002 | 373 | 42 | 93 | 15 | 150 | 40.2 | 29.3 | |
| 2003 | 447 | 52 | 92 | 23 | 167 | 37.4 | 22.9 | |
| 2004 | 571 | 104 | 128 | 33 | 265 | 46.4 | 26.2 | |
| 2005 | 641 | 102 | 152 | 41 | 295 | 46.0 | 29.6 | |
| 2006 | 781 | 105 | 201 | 49 | 355 | 45.5 | 28.8 | |
| 2007 | 1625 | 158 | 257 | 79 | 41 | 535 | 32.9 | 30.8 |
| 2008 | 1534 | 145 | 258 | 84 | 36 | 523 | 34.1 | 23.2 |
| 2009 | 2774 | 159 | 255 | 73 | 44 | 531 | 19.1 | 22.8 |
| 2010 | 1350 | 135 | 308 | 95 | 69 | 607 | 45.0 | 19.5 |
| 2011 | 1265 | 129 | 244 | 94 | 78 | 545 | 43.1 | 23.6 |
| 2012 | 1379 | 175 | 326 | 87 | 83 | 671 | 48.7 | 25.0 |
| 2013 | 2120 | 180 | 408 | 126 | 111 | 825 | 38.9 | 22.3 |
| 2014 | 2810 | 283 | 525 | 229 | 161 | 1198 | 42.6 | 23.4 |
| 2015 | 2824 | 336 | 679 | 295 | 176 | 1486 | 52.6 | 21.7 |
| 2016 | 3327 | 403 | 988 | 402 | 282 | 2075 | 62.4 | 22.1 |
| 2017 | 2554 | 341 | 735 | 295 | 223 | 1593 | 62.4 | 21.9 |
| 2018 | 4065 | 548 | 1283 | 576 | 463 | 2870 | 70.6 | 19.6 |
| 2019 | 5027 | 733 | 1495 | 724 | 700 | 3652 | 72.6 | 18.2 |
| 2020 | 4717 | 505 | 1060 | 503 | 1325 | 3393 | 71.9 | 18.5 |
| 2021 | 5281 | 513 | 1357 | 696 | 1281 | 3847 | 72.8 | 17.8 |
| 2022 | 4902 | 573 | 1255 | 615 | 1230 | 3673 | 74.9 | 16.8 |
Table 3.
Roe deer population management in Lithuania by time periods.
| Years | Period | Short Description | References |
|---|---|---|---|
| 1948–1964 | Hunting ban | During the period of the post-war hunting ban, the roe deer population increased 6.7 times (see Figure 2). | [11,36,59] |
| 1965–1988 | Central regulation | The hunted roe deer were delivered to state-owned meat factories; therefore, hunter interest was kept to minimum. | [139] |
| 1986–1994 | Overhunting | Based on the opinion that deer overabundance is threatening forests, the hunting bag was set at over 20% of the winter population (Figure 4). This period ended with a decrease in the population and a temporary ban on roe deer hunting in 1997. | [140,141,142] |
| 1990–1994 | Hunting reorganization | In the reorganization period following Lithuania’s independence, administrative and legal changes negatively affected roe deer numbers. Overhunting was driven by norms indicating maximum density limits for deer in forests. | [30,143] |
| 1995 | Hunt regulations | The hunting of roe deer in the Republic of Lithuania is regulated by the Adopted Regulation for Hunting and the Rules of Hunting. Roe deer hunting quotas are set annually. | [34] |
| 2002 | Law on Hunting | Concepts of hunting, hunting plot units, and selectional hunting established. Hunting grounds rent terms set for no shorter than 10 years. | [144] |
| 2020 | Limits lifted | From 2020 onwards, roe deer can be hunted without limits. | [34] |
| 2021 | Selective deer hunting lifted | There are no mandatory regulations specifically addressing selective deer hunting; the only restriction pertains to the length of the hunting season. Currently, roe deer bucks can be hunted from 1 May to 31 October, while does and calves can be hunted from 1 October to 31 January. | [34] |
Table 4.
Norms of allowable maximum density of roe deer, red deer, and moose in the forests of Lithuania (1995), which become Norms of recommended maximum density in 2005 [30].
Table 4.
Norms of allowable maximum density of roe deer, red deer, and moose in the forests of Lithuania (1995), which become Norms of recommended maximum density in 2005 [30].
| Forest Type | Number of Animals (Per 1000 ha of Forest) | ||
|---|---|---|---|
| Moose | Red Deer | Roe Deer | |
| Deciduous forests with admixture of coniferous trees | 4 | 15 | 55 |
| Mixed coniferous forests with admixture of deciduous stands | 3 | 12 | 45 |
| Mixed coniferous forests with scarce deciduous admixture | 2 | 12 | 34 |
| Pure pine forests | 1 | 6 | 14 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Balčiauskas, L. Roe Deer, Lithuania’s Smallest and Most Abundant Cervid. Forests 2024, 15, 767. https://doi.org/10.3390/f15050767
AMA Style
Balčiauskas L. Roe Deer, Lithuania’s Smallest and Most Abundant Cervid. Forests. 2024; 15(5):767. https://doi.org/10.3390/f15050767
Chicago/Turabian StyleBalčiauskas, Linas. 2024. "Roe Deer, Lithuania’s Smallest and Most Abundant Cervid" Forests 15, no. 5: 767. https://doi.org/10.3390/f15050767
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.