Cellulolytic Bacteria Associated with the Gut of Dendroctonus armandi Larvae (Coleoptera: Curculionidae: Scolytinae)


Abstract
:1. Introduction
2. Experimental Section
2.1. Insect Collection and Dissection
2.2. Isolation and Counting of Cellulolytic Microorganisms
2.3. CMC Assay
2.4. ARDRA
2.5. Molecular and Phylogenetic Analysis
2.6. Operational Taxonomic Units (OTU)
2.7. Morphological and Physiological Characteristic Tests
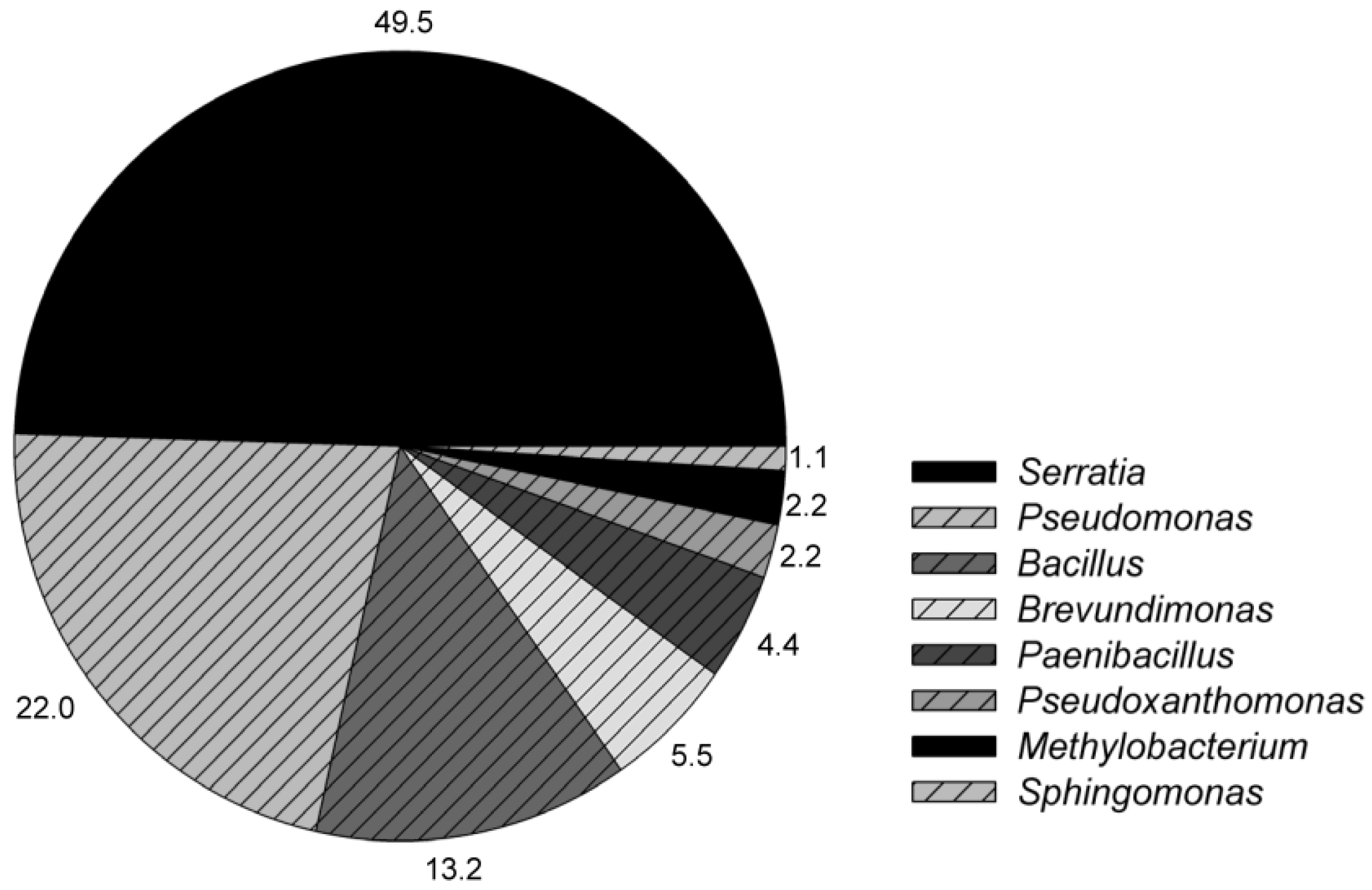
3. Results and Discussion



{kind=link}
{kind=link}
{kind=link}
| Representative strains | Closest known taxa | Colony | Cell morphology | Pellicle | Flagella | Spore | G * | Numnero of isolates | |
|---|---|---|---|---|---|---|---|---|---|
| Medium 2a | Medium 2b | ||||||||
| A | Serratia | Pink | Rods, 0.39–0.72 × 0.92–6.43 μm | + | Peritrichous | − | − | 2 | 5 |
| B | Serratia | Pink | Rods, 0.31–0.66 × 0.96–5.54 μm | + | Peritrichous | − | − | 11 | 20 |
| C | Pseudomonas | Ivory | Rods, 0.35–0.66 × 1.08–2.75 μm | − | ND | − | − | 10 | 10 |
| D | Bacillus | Ivory | Rods, 0.38–0.67 × 0.84–3.24 μm | + | Peritrichous | + | + | 2 | 10 |
| F | Paenibacillus | Pink | Rods, 0.64–1.32 × 2.12–6.10 μm | − | Peritrichous | + | + | 0 | 4 |
| H | Sphingomonas | Yellow | Rods, 0.32–0.59 × 0.98–10.62 μm | − | Peritrichous | + | − | 0 | 1 |
| I | Serratia | Pink | Rods, 0.36–0.70× 0.91–4.98 μm | + | Peritrichous | − | − | 2 | 5 |
| J | Brevundimonas | Pale yellow | Rods, 0.61–1.13× 1.64–3.50 μm | − | Peritrichous | + | − | 0 | 2 |
| M | Brevundimonas | Ivory | Rods, 0.28–0.50× 1.16–3.40 μm | − | Peritrichous | + | − | 2 | 1 |
| N | Pseudoxanthomonas | Yellow | Rods, 0.21–0.38× 1.05–2.50 μm | + | Polar | − | − | 1 | 1 |
| P | Methylobacterium | Pale yellow | Rods, 0.32–0.50× 0.89–2.40 μm | − | Polar or subpolar | − | − | 2 | 0 |

4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Purves, C.B. Chemical Nature of Cellulose and its Derivatives. In Cellulose and Cellulose Derivatives, Part 1; Spurlin, H.M., Grafflin, M.W., Eds.; Interscience: New York, NY, USA, 1954; pp. 29–98. [Google Scholar]
- Ghio, S.; di Lorenzo, G.S.; Lia, V.; Talia, P.; Cataldi, A.; Grasso, D.; Campos, E. Isolation of Paenibacillus sp. and Variovorax sp. Strains from decaying woods and characterization of their potential for cellulose deconstruction. Int. J. Biochem. Mol. Biol. 2012, 3, 352–364. [Google Scholar]
- Sun, J.Z.; Scharf, M. Exploring and integrating cellulolytic systems of insects to advance biofuel technology. Insect Sci. 2010, 17, 163–165. [Google Scholar] [CrossRef]
- Huang, S.; Sheng, P.; Zhang, H. Isolation and identification of cellulolytic bacteria from the gut of Holotrichia parallela larvae (coleoptera: Scarabaeidae). Int. J. Mol. Sci. 2012, 13, 2563–2577. [Google Scholar] [CrossRef]
- Ni, J.; Tokuda, G. Lignocellulose-degrading enzymes from termites and their symbiotic microbiota. Biotechnol. Adv. 2013, 31, 838–850. [Google Scholar] [CrossRef]
- Haack, R.A.; Slansky, F., Jr. Nutritional Ecology of Wood-Feeding Coleoptera, Lepidoptera, and Hymenoptera. In Nutritional Ecology of Insects, Mites, Spiders, and Related Invertebrates; Slansky, F., Jr., Rodriguez, J., Eds.; Wiley: New York, NY, USA, 1987; pp. 449–486. [Google Scholar]
- Coulson, R.; Stark, R. Integrated management of bark beetles. In Bark Beetles in North American Conifers: A System for the Study of Evolutionary Biology; Mitton, J.B., Sturgeon, K.B., Eds.; University of Texas Press: Austin, TX, USA, 1982; pp. 315–349. [Google Scholar]
- Chen, H.; Tang, M.; Liu, L.; Wang, H.; Li, Z. Cytochemical localization of acid phosphatase activity in tissues of Pinus armandi infected by Leptographium qinlingensis. Symbiosis (Rehovot) 2007, 43, 65–70. [Google Scholar]
- Delalibera, I.; Handelsman, J., Jr.; Raffa, K.F. Contrasts in cellulolytic activities of gut microorganisms between the wood borer, Saperda vestita (coleoptera: Cerambycidae), and the bark beetles, Ips pini and Dendroctonus frontalis (coleoptera: Curculionidae). Environ. Entomol. 2005, 34, 541–547. [Google Scholar] [CrossRef]
- Morales-Jiménez, J.; Zúñiga, G.; Ramírez-Saad, H.C.; Hernández-Rodríguez, C. Gut-associated bacteria throughout the life cycle of the bark beetle Dendroctonus rhizophagus Thomas and Bright (Curculionidae: Scolytinae) and their cellulolytic activities. Microb. Ecol. 2012, 64, 268–278. [Google Scholar]
- Hu, X.; Wang, C.; Chen, H.; Ma, J. Differences in the structure of the gut bacteria communities in development stages of the Chinese white pine beetle (Dendroctonus armandi). Int. J. Mol. Sci. 2013, 14, 21006–21020. [Google Scholar] [CrossRef]
- Teather, R.M.; Wood, P.J. Use of congo red-polysaccharide interactions in enumeration and characterization of cellulolytic bacteria from the bovine rumen. Appl. Environ. Microbiol. 1982, 43, 777–780. [Google Scholar]
- Weisurg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar]
- Martínez-Murcia, A.; Acinas, S.; Rodriguez-Valera, F. Evaluation of prokaryotic diversity by restrictase digestion of 16S rDNA directly amplified from hypersaline environments. FEMS Microbiol. Ecol. 1995, 17, 247–255. [Google Scholar]
- Cole, J.R.; Wang, Q.; Cardenas, E.; Fish, J.; Chai, B.; Farris, R.J.; Kulam-Syed-Mohideen, A.S.; McGarrell, D.M.; Marsh, T.; Garrity, G.M.; et al. The ribosomal database project: Improved alignments and new tools for rRNA analysis. Nucleic Acids Res. 2009, 37, D141–D145. [Google Scholar] [CrossRef]
- Wheeler, D.L.; Barrett, T.; Benson, D.A.; Bryant, S.H.; Canese, K.; Chetvernin, V.; Church, D.M.; DiCuccio, M.; Edgar, R.; Federhen, S.; et al. Database resources of the national center for biotechnology information. Nucleic Acids Res. 2007, 35, D5–D12. [Google Scholar] [CrossRef]
- Edgar, R.C. Muscle: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. Mega5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Schloss, P.D.; Handelsman, J. Introducing DOTUR, a computer program for defining operational taxonomic units and estimating species richness. Appl. Environ. Microbiol. 2005, 71, 1501–1506. [Google Scholar] [CrossRef]
- Gregersen, T. Rapid method for distinction of gram-negative from gram-positive bacteria. Eur. J. Appl. Microbiol. Biotechnol. 1978, 5, 123–127. [Google Scholar] [CrossRef]
- Smibert, R. Phenotypic Characterization. In Methods for General and Molecular Bacteriology; Murray, R.G.E., Wood, W.A., Eds.; American Society for Microbiology: Washington, DC, USA, 1994. [Google Scholar]
- Cazemier, A.E.; Verdoes, J.C.; Reubsaet, F.A.; Hackstein, J.H.; van der Drift, C.; den Camp, H.J.O. Promicromonospora pachnodae sp. nov., a member of the (hemi) cellulolytic hindgut flora of larvae of the scarab beetle Pachnoda marginata. Anton. Leeuw. Int. J. G. 2003, 83, 135–148. [Google Scholar] [CrossRef]
- Petersen, L.M.; Tisa, L.S.S. Friend or foe? A review of the mechanisms that drive Serratia towards diverse lifestyles. Can. J. Microbiol. 2013, 59, 627–640. [Google Scholar] [CrossRef]
- Yilmaz, H.; Sezen, K.; Kati, H.; Demirbað, Z. The first study on the bacterial flora of the European spruce bark beetle, Dendroctonus micans (Coleoptera: Scolytidae). Biologia 2006, 61, 679–686. [Google Scholar] [CrossRef]
- Vasanthakumar, A.; Delalibera, I.; Handelsman, J.; Klepzig, K.D.; Schloss, P.D.; Raffa, K.F. Characterization of gut-associated bacteria in larvae and adults of the southern pine beetle, Dendroctonus frontalis Zimmermann. Environ. Entomol. 2006, 35, 1710–1717. [Google Scholar] [CrossRef]
- Anand, A.A.P.; Vennison, S.J.; Sankar, S.G.; Prabhu, D.I.G.; Vasan, P.T.; Raghuraman, T.; Geoffrey, C.G.; Vendan, S.E. Isolation and characterization of bacteria from the gut of Bombyx mori that degrade cellulose, xylan, pectin and starch and their impact on digestion. J. Insect Sci. 2010, 10. [Google Scholar] [CrossRef]
- Palleroni, N.J. The Pseudomonas story. Environ. Microbiol. 2010, 12, 1377–1383. [Google Scholar] [CrossRef]
- Brodey, C.L.; Rainey, P.B.; Tester, M.; Johnstone, K. Bacterial blotch disease of the cultivated mushroom is caused by an ion channel forming lipodepsipeptide toxin. Mol. Plant-Microbe Interact. 1991, 4, 407–411. [Google Scholar] [CrossRef]
- Young, J. Drippy gill: A bacterial disease of cultivated mushrooms caused by Pseudomonas agaricin sp. N. Z. J. Agric. Res. 1970, 13, 977–990. [Google Scholar] [CrossRef]
- Kodama, K.; Kimura, N.; Komagata, K. Two new species of pseudomonas: P. Oryzihabitans isolated from rice paddy and clinical specimens and P. Luteola isolated from clinical specimens. Int. J. Syst. Evol. Microbiol. 1985, 35, 467–474. [Google Scholar]
- Meyers, M.; Poffe, R.; Verachtert, H. Properties of a cellulolytic pseudomonas. Antonie van Leeuwenhoek 1984, 50, 301–301. [Google Scholar] [CrossRef]
- Sindhu, S.; Dadarwal, K. Chitinolytic and cellulolytic Pseudomonas sp. Antagonistic to fungal pathogens enhances nodulation by Mesorhizobium sp. Cicer in chickpea. Microbiol. Res. 2001, 156, 353–358. [Google Scholar] [CrossRef]
- Trivedi, N.; Gupta, V.; Kumar, M.; Kumari, P.; Reddy, C.; Jha, B. An alkali-halotolerant cellulase from Bacillus flexus isolated from green seaweed Ulva lactuca. Carbohydr. Polym. 2011, 83, 891–897. [Google Scholar] [CrossRef]
- Yang, J.K.; Zhang, J.J.; Yu, H.Y.; Cheng, J.W.; Miao, L.H. Community composition and cellulase activity of cellulolytic bacteria from forest soils planted with broad-leaved deciduous and evergreen trees. Appl. Microbiol. Biotechnol. 2013. [Google Scholar] [CrossRef]
- Okeke, B.C.; Lu, J. Characterization of a defined cellulolytic and xylanolytic bacterial consortium for bioprocessing of cellulose and hemicelluloses. Appl. Biochem. Biotechnol. 2011, 163, 869–881. [Google Scholar] [CrossRef]
- Ferreira Filho, A.S.; Quecine, M.C.; Bogas, A.C.; de Barros Rossetto, P.; de Souza Lima, A.O.; Lacava, P.T.; Azevedo, J.L.; Araújo, W.L. Endophytic methylobacterium extorquens expresses a heterologous â-1,4-endoglucanase A (EglA) in Catharanthus roseus seedlings, a model host plant for xylella fastidiosa. World J. Microbiol. Biotechnol. 2012, 28, 1475–1481. [Google Scholar] [CrossRef]
- Wenzel, M.; Schönig, I.; Berchtold, M.; Kämpfer, P.; König, H. Aerobic and facultatively anaerobic cellulolytic bacteria from the gut of the termite Zootermopsis angusticollis. J. Appl. Microbiol. 2002, 92, 32–40. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hu, X.; Yu, J.; Wang, C.; Chen, H. Cellulolytic Bacteria Associated with the Gut of Dendroctonus armandi Larvae (Coleoptera: Curculionidae: Scolytinae). Forests 2014, 5, 455-465. https://doi.org/10.3390/f5030455
Hu X, Yu J, Wang C, Chen H. Cellulolytic Bacteria Associated with the Gut of Dendroctonus armandi Larvae (Coleoptera: Curculionidae: Scolytinae). Forests. 2014; 5(3):455-465. https://doi.org/10.3390/f5030455
Chicago/Turabian StyleHu, Xia, Jiamin Yu, Chunyan Wang, and Hui Chen. 2014. "Cellulolytic Bacteria Associated with the Gut of Dendroctonus armandi Larvae (Coleoptera: Curculionidae: Scolytinae)" Forests 5, no. 3: 455-465. https://doi.org/10.3390/f5030455