
Growth and Nutrient Status of Foliage as Affected by Tree Species and Fertilization in a Fire-Disturbed Urban Forest


Abstract
:1. Introduction
2. Experimental Section
2.1. Site

2.2. Methods

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tree Species | Location | Elevation (m) | Stand Density (trees·ha−1) | DBH † (mm) | Height (m) |
|---|---|---|---|---|---|
| Liriodendron tulipifera | 35°32′34.92″ N 129°26′22.92″ E | 154 | 1666 (160) | 28.6 (2.8) | 2.87 (0.14) |
| Prunus yedoensis | 35°32′26.48″ N 129°26′16.22″ E | 121 | 1267 (160) | 49.4 (2.3) | 4.10 (0.13) |
| Quercus acutissima | 35°32′26.77″ N 125°26′15.15″ E | 118 | 1533 (66) | 26.4 (1.1) | 2.22 (0.10) |
| Pinus thunbergii | 35°32′16.36″ N 125°26′17.56″ E | 98 | 1600 (206) | 27.3 (3.7) | 2.31 (0.11) |
| Tree Species | B.D. † g·cm−3 | Sand | Silt | Clay | C | N | P mg·kg−1 | K+ | Ca2+ | Mg2+ |
|---|---|---|---|---|---|---|---|---|---|---|
| % | cmolc·kg−1 | |||||||||
| Liriodendron tulipifera | 0.90 (0.01) | 49 (4.7) | 40 (4.0) | 10 (0.7) | 3.1 (0.50) | 0.15 (0.02) | 6.0 (2.0) | 0.16 (0.02) | 1.60 (0.28) | 0.50 (0.09) |
| Prunus yedoensis | 0.89 (0.01) | 47 (1.3) | 43 (0.7) | 10 (1.2) | 3.6 (0.70) | 0.17 (0.02) | 1.2 (0.5) | 0.29 (0.11) | 1.54 (0.39) | 0.56 (0.13) |
| Quercus acutissima | 0.88 (0.02) | 43 (1.8) | 47 (2.9) | 10 (1.2) | 1.9 (0.04) | 0.08 (0.01) | 9.9 (0.7) | 0.09 (0.01) | 0.43 (0.06) | 0.19 (0.02) |
| Pinus thunbergii | 0.91 (0.01) | 63 (8.7) | 28 (8.1) | 9 (0.7) | 1.9 (0.01) | 0.07 (0.01) | 6.5 (2.6) | 0.08 (0.02) | 0.38 (0.11) | 0.16 (0.03) |

2.3. Statistical Analysis
3. Results and Discussion
3.1. Results
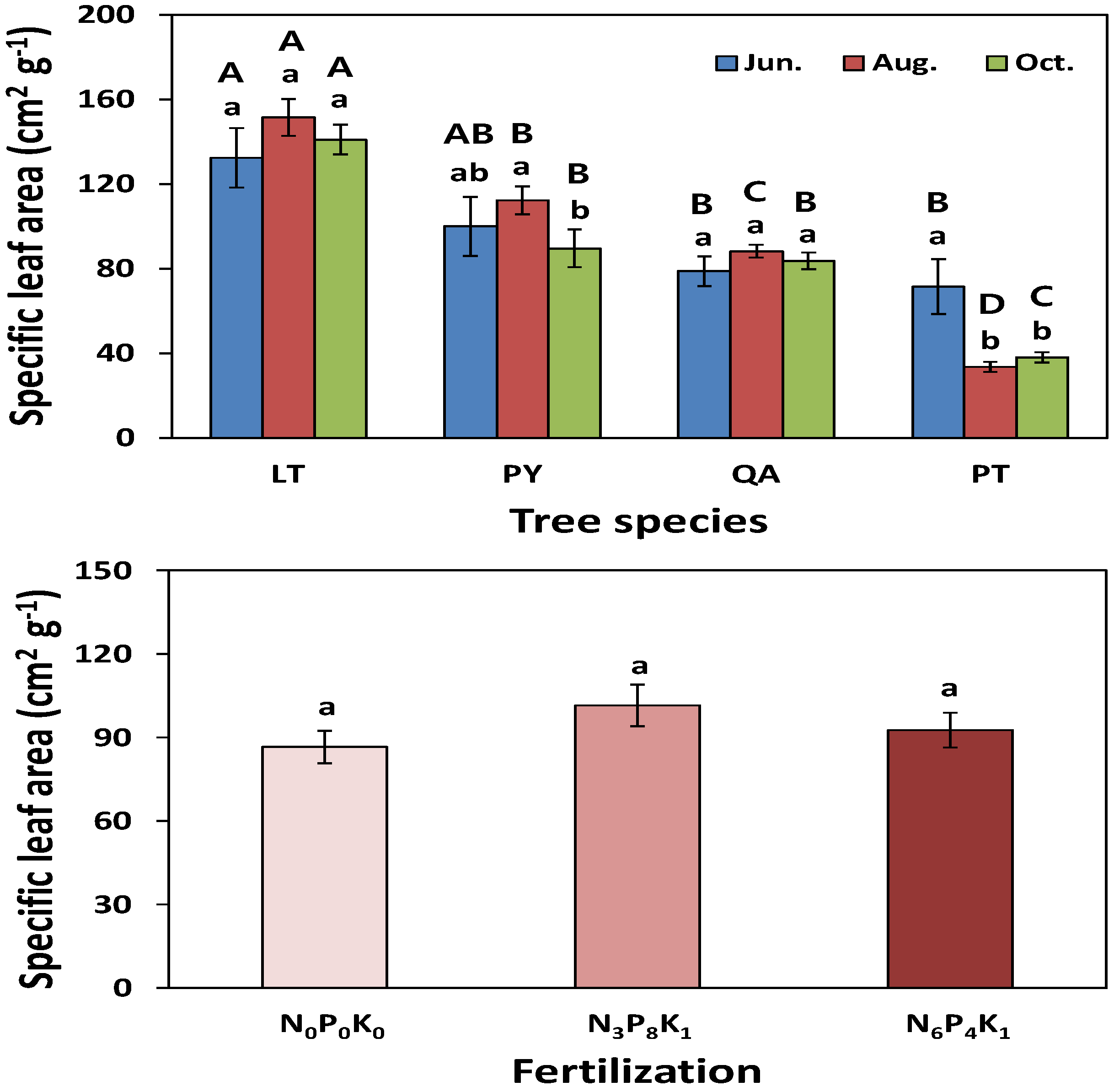
3.1.1. Growth Response
| Component | Leaf Area | Specific Leaf Area | Dry Weight | Nutrient Concentration | C/N Ratio | Nutrient Content | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C | N | P | K | C | N | P | K | |||||
| Species (S) | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Fertilizer (F) | <0.001 | 0.049 | 0.002 | 0.444 | <0.001 | <0.001 | 0.666 | <0.001 | 0.003 | <0.001 | <0.001 | 0.158 |
| Month (M) | 0.003 | 0.309 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.066 | <0.001 | <0.001 |
| S × F | 0.002 | 0.545 | 0.018 | 0.231 | 0.032 | 0.264 | 0.986 | 0.037 | 0.030 | 0.007 | 0.003 | 0.707 |
| S × M | 0.234 | 0.012 | 0.269 | <0.001 | 0.016 | 0.770 | 0.126 | 0.424 | 0.187 | 0.278 | 0.002 | <0.001 |
| F × M | 0.861 | 0.981 | 0.903 | 0.419 | 0.614 | 0.275 | 0.699 | 0.834 | 0.917 | 0.945 | 0.807 | 0.994 |
| S × F × M | 0.999 | 0.685 | 0.625 | 0.830 | 0.978 | 0.999 | 0.867 | 0.940 | 0.607 | 0.950 | 0.659 | 0.963 |


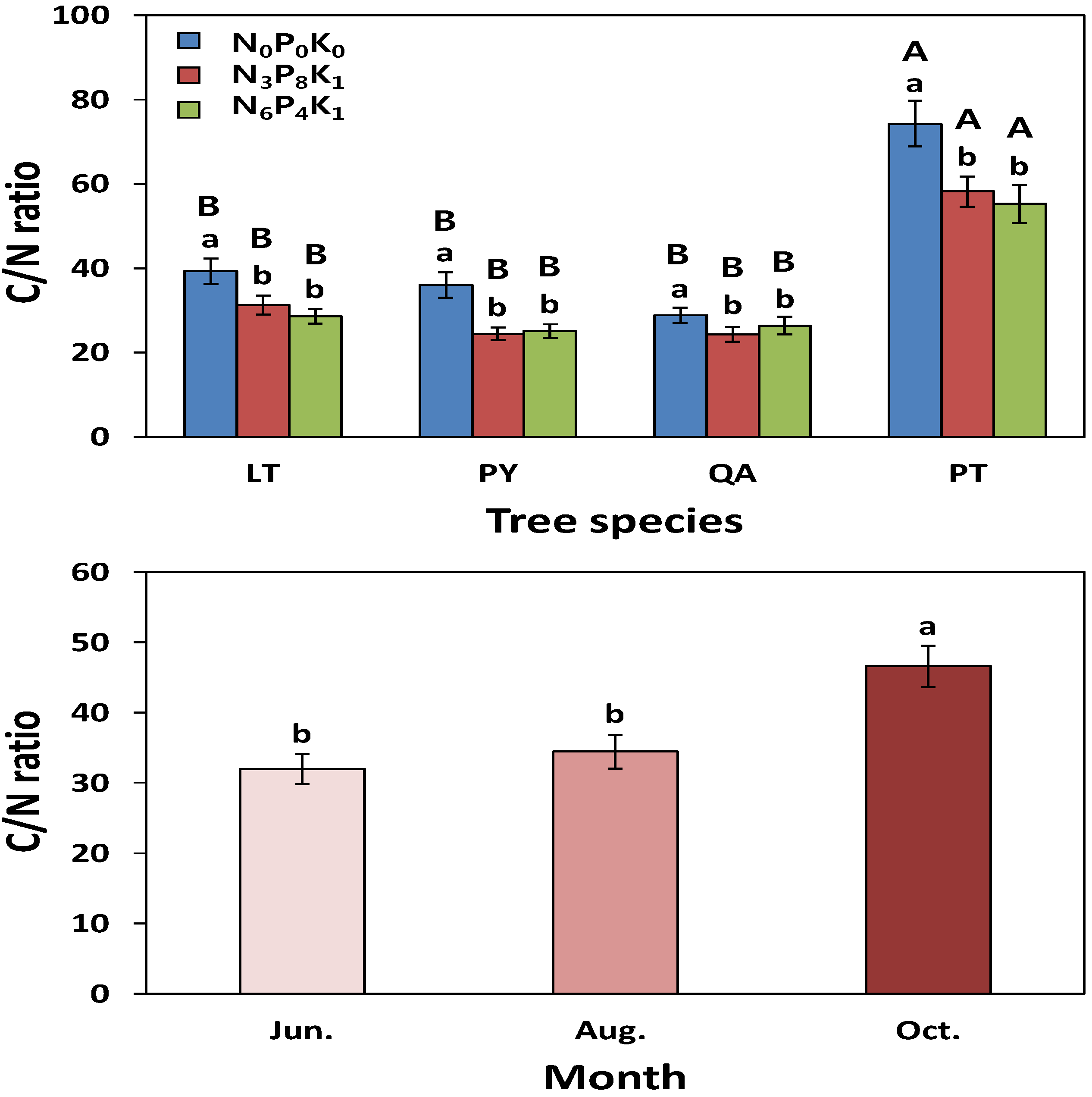
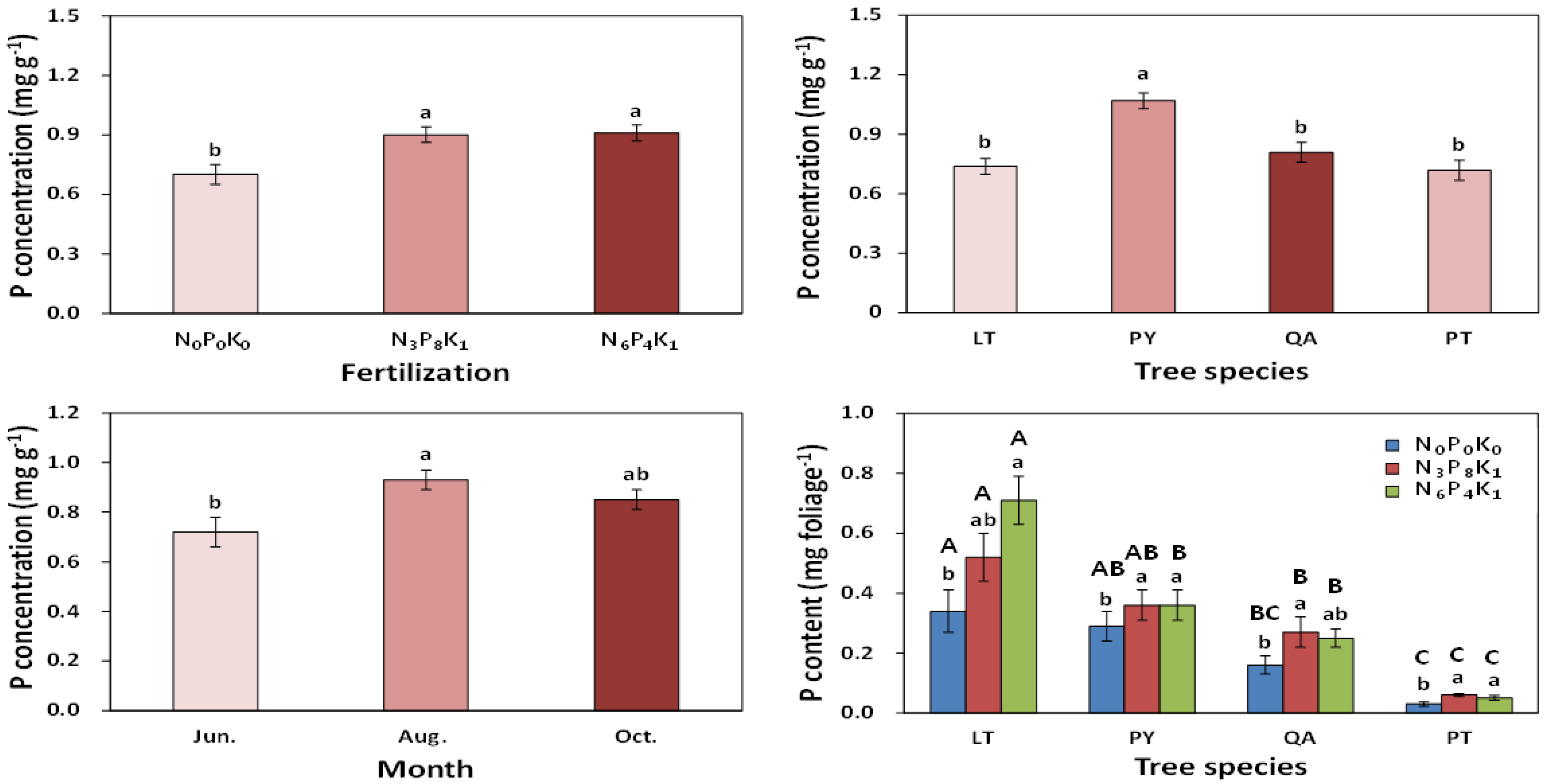
3.1.2. Nutrient Responses




3.2. Discussion
3.2.1. Growth Responses
3.2.2. Nutrient Responses
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Weetman, G.F.; Wells, C.G. Plant analyses as an aid in fertilizing forests. In Soil Testing and Plant Analysis, 3rd ed.; Westerman, R.L., Ed.; Soil Science Society of America: Madison, WI, USA, 1990; pp. 659–690. [Google Scholar]
- Barron-Gafford, G.A.; Will, R.E.; Burkes, C.; Shiver, B.; Teskey, R.O. Nutrient concentrations and content, their relation to stem growth, of intensively managed Pinus taeda and Pinus elliottii stands of different planting densities. For. Sci. 2003, 49, 291–300. [Google Scholar]
- Tausz, M.; Trummer, W.; Wonisch, A.; Goessler, W.; Grill, D.; Jiménez, M.S.; Morales, D. A survey of foliar mineral nutrient concentrations of Pinus canariensis at field plots in Tenerife. For. Ecol. Manag. 2004, 189, 49–55. [Google Scholar] [CrossRef]
- Silfverberg, K.; Moilanen, M. Long-term nutrient status of PK fertilized Scots pine stands on drained peatlands in North-Central Finland. Suo 2008, 59, 71–88. [Google Scholar]
- Pakaki, D.E.; Carreir, M.M.; Cherrier, J.; Grulke, N.E.; Jennings, V.; Princetl, S.; Pouyat, R.V.; Whitlow, T.H.; Zipperer, W.C. Coupling biogeochemical cycles in urban environments: Ecosystem services, green solutions, and misconceptions. Front. Ecol. Environ. 2011, 9, 27–36. [Google Scholar]
- Escobedo, F.J.; Nowak, D.J. Spatial heterogeneity and air pollution removal by an urban forest. Landsc. Urban Plan. 2009, 90, 102–110. [Google Scholar] [CrossRef]
- Kim, C.; Koo, K.S.; Byun, J.K.; Jeong, J.H. Post-fire effects on soil properties in red pine (Pinus densiflora) stands. Forest Sci. Tech. 2005, 1, 1–7. [Google Scholar] [CrossRef]
- Xue, L.; Li, Q.; Chen, H. Effects of a wildfire on selected physical, chemical and biochemical soil properties in a Pinus massoniana forest in south China. Forests 2014, 5, 2947–2966. [Google Scholar] [CrossRef]
- Kim, Y.S.; Byun, J.K.; Kim, C.; Park, B.B.; Kim, Y.K.; Bae, S.W. Growth response of Pinus densiflora seedlings to different fertilizer compound ratios in a recently burned area in the eastern coast of Korea. Landsc. Ecol. Eng. 2014, 10, 241–247. [Google Scholar] [CrossRef]
- Garrison, M.T.; Moore, J.A.; Shaw, T.M.; Mika, P.G. Foliar nutrient and tree growth response of mixed-conifer stands to three fertilization treatments in northeast Oregon and north central Washington. For. Ecol. Manag. 2000, 132, 183–198. [Google Scholar] [CrossRef]
- Amponsah, I.G.; Comeau, P.G.; Brockley, R.P.; Lieffers, V.J. Effects of repeated fertilization on needle longevity, foliar nutrition, effective leaf area index, and growth characteristics of lodgepole pine in interior British Columbia, Canada. Can. J. For. Res. 2005, 35, 440–451. [Google Scholar] [CrossRef]
- Miller, J.H.; Allen, H.L.; Zutter, B.R.; Zedaker, S.M.; Newbold, R.A. Soil and pine foliage nutrient responses 15 years after competing-vegetation control and their correlation with growth for 13 loblolly pine plantations in the southern United States. Can. J. For. Res. 2006, 36, 2412–2425. [Google Scholar] [CrossRef]
- Kim, C.; Ju, N.K.; Lee, H.Y.; Lee, K.S. Effects of growth, carbon and nitrogen response of foliage in a red pine stand. Korean J. Soil Sci. Fertil. 2013, 46, 9–15. [Google Scholar] [CrossRef]
- Kim, C.; Byun, J.K.; Park, J.H.; Ma, H.S. Litter fall and nutrient status of green leaves and leaf litter at various compound ratios of fertilizer in sawtooth oak stands, Korea. Ann. For. Res. 2013, 56, 339–350. [Google Scholar]
- Mugasha, A.G.; Pluth, D.J.; Macdonald, S.E. Effects of fertilization on seasonal patterns of foliar mass and nutrients of tamarack and black spruce on undrained and drained minerotrophic peatland sites. For. Ecol. Manag. 1999, 116, 13–31. [Google Scholar] [CrossRef]
- Bauer, G.; Schulze, E.-D.; Mund, M. Nutrient contents and concentrations in relation to growth of Picea abies and Fagus sylvatica along a European transect. Tree Physiol. 1997, 17, 777–786. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, L. Logic of experiments in ecology: Is pseudoreplication a pseudoissue? Oikos 2002, 94, 27–38. [Google Scholar] [CrossRef]
- SAS Institute Inc. SAS/STAT Statistical Software; Version 9.1; SAS Institute Inc.: Cary, NC, USA, 2003. [Google Scholar]
- Binkley, D.; Fisher, R. Nutrient management. In Ecology and Management of Forest Soils, 4th ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2013; pp. 254–275. [Google Scholar]
- Gough, C.M.; Seiler, J.R.; Maier, C.A. Short-term effects of fertilization on loblolly pine (Pinus taeda L.) physiology. Plant Cell Environ. 2004, 27, 876–886. [Google Scholar] [CrossRef]
- Laclau, J.P.; Almeida, J.S.R.; Gonçalves, J.L.M.; Saint-André, L.; Ventura, M.; Ranger, R.; Moreira, R.M.; Nouvellon, Y. Influence of nitrogen and potassium fertilization on leaf lifespan and allocation of above-ground growth in Eucalyptus plantations. Tree Physio. 2009, 29, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Wilson, P.J.; Thompson, K.; Hodgson, J.G. Specific leaf area and leaf dry matter content as alternative predictors of plant strategies. New Phytol. 1999, 143, 155–162. [Google Scholar] [CrossRef]
- Bert, D.; Danjon, D. Carbon concentration variations in the roots, stem and crown of mature Pinus pinaster (Ait.). For. Ecol. Manag. 2006, 222, 279–295. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, C.; Wang, X.; Quan, X. Carbon concentration variability of 10 Chinese temperate tree species. For. Ecol. Manag. 2009, 258, 722–727. [Google Scholar] [CrossRef]
- Jeong, J.; Park, J.-H.; Kim, J.-I.; Lim, J.-T.; Lee, S.-R.; Kim, C. Effects of container volumes and fertilization on red (Pinus densiflora) and black pine (Pinus thunbergii) seedlings growth. Forest Sci. Tech. 2010, 6, 80–86. [Google Scholar] [CrossRef]
- Sariyildiz, T.; Anderson, J.M. Variation in the chemical composition of green leaves and leaf litters from three deciduous tree species growing on different soil types. For. Ecol. Manag. 2005, 210, 303–319. [Google Scholar] [CrossRef]
- Hagon-Thorn, A.; Varnagiryte, I.; Nihlgard, B.; Armolaitis, K. Autumn nutrient resorption and losses in four deciduous forest tree species. For. Ecol. Manag. 2006, 228, 33–39. [Google Scholar] [CrossRef]
- O’connell, A.M.; Grove, T.S. Influence of nitrogen and phosphorus fertilizers on amount and nutrient content of litterfall in a regrowth eucalypt forest. New For. 1993, 7, 33–47. [Google Scholar] [CrossRef]
- Scott, A.; Burger, J.A.; Kaczmarek, D.J.; Kane, M.B. Growth and nutrition response of young sweetgum plantations to repeated nitrogen fertilizer on two site types. Biomass Bioenerg. 2004, 27, 313–325. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, C.; Jeong, J.; Park, J.-H.; Ma, H.-S. Growth and Nutrient Status of Foliage as Affected by Tree Species and Fertilization in a Fire-Disturbed Urban Forest. Forests 2015, 6, 2199-2213. https://doi.org/10.3390/f6062199
Kim C, Jeong J, Park J-H, Ma H-S. Growth and Nutrient Status of Foliage as Affected by Tree Species and Fertilization in a Fire-Disturbed Urban Forest. Forests. 2015; 6(6):2199-2213. https://doi.org/10.3390/f6062199
Chicago/Turabian StyleKim, Choonsig, Jaeyeob Jeong, Jae-Hyun Park, and Ho-Seop Ma. 2015. "Growth and Nutrient Status of Foliage as Affected by Tree Species and Fertilization in a Fire-Disturbed Urban Forest" Forests 6, no. 6: 2199-2213. https://doi.org/10.3390/f6062199