
Sub-Soiling and Genotype Selection Improves Populus Productivity Grown on a North Carolina Sandy Soil

Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description, Design, and Establishment
2.2. Data Collection for Tree Growth and Melampsora spp. Rust Resistance Index
2.3. LAI, Chlorophyll Content, and Foliar N
2.4. Statistical Analyses
3. Results
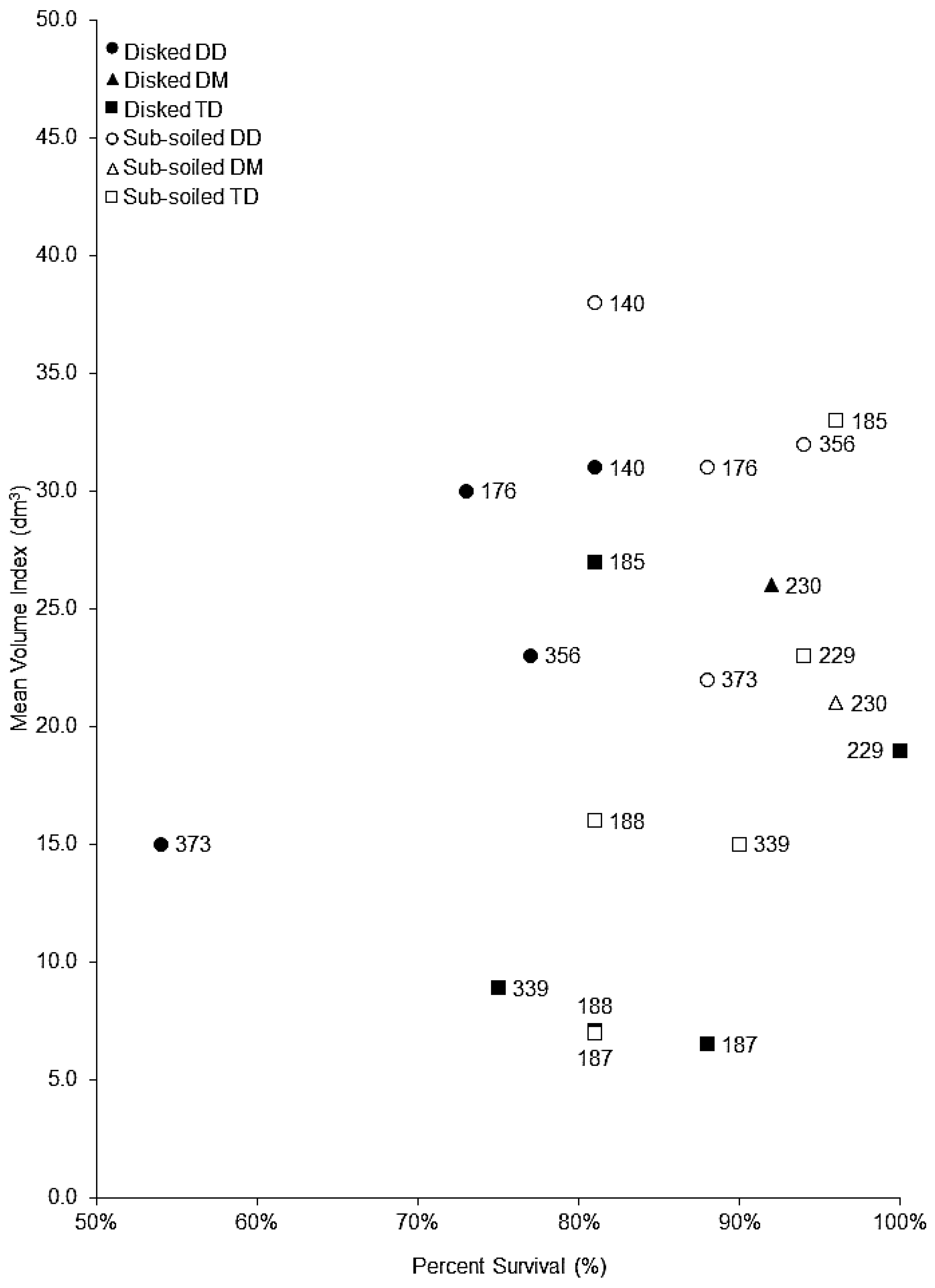
3.1. Tillage and Genotype Selection
3.2. Chlorophyll Content, Foliar N, and Volume Index
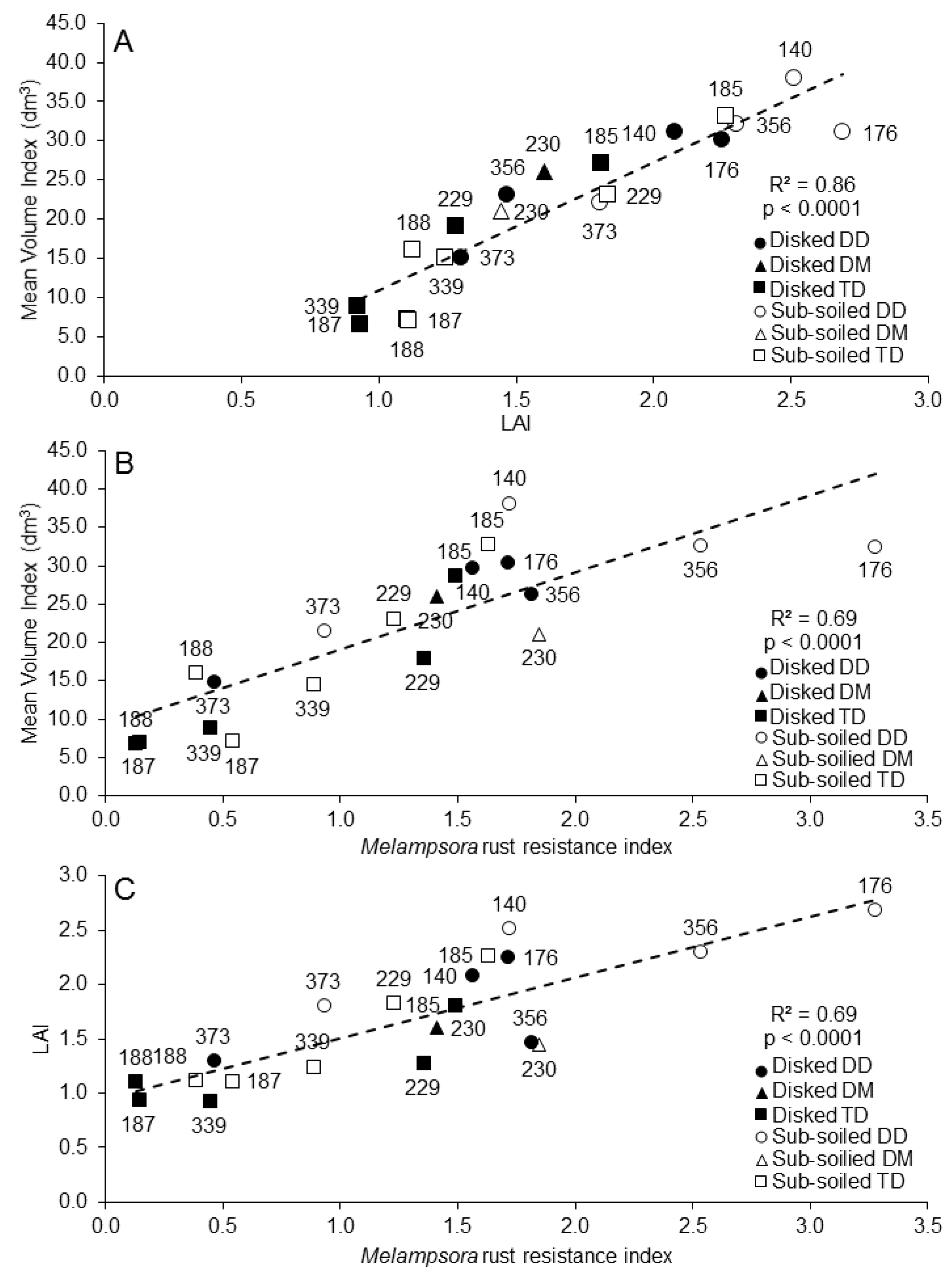
3.3. Melampsora Rust Resistance, LAI, and Volume Index
4. Discussion
4.1. Tillage and Genotype Selection
4.2. Chlorophyll Content, Foliar N, and Volume Index
4.3. Melampsora Rust Resistance, LAI, and Volume Index
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- European Commission. Directive 2009/28/EC of the European Parliament and of the Council—on the Promotion of the Use of Energy from Renewable Sources and Amending and Subsequently Repealing Directives 2001/77/EC and 2003/30/EC; European Commission: Brussels, Belgium, 2009; p. 62. [Google Scholar]
- Sikkema, R.; Steiner, M.; Junginger, M.; Hiegl, W.; Hansen, M.T.; Faaij, A. The European wood pellet markets: Current status and prospects for 2020. Biofuel Bioprod. Biorefin. 2011, 5, 250–278. [Google Scholar] [CrossRef]
- Federal Register. Regulation of Fuels and Fuel Additives: Changes to Renewable Fuel Standard; Federal Register: Washington, DC, USA, 26 March 2010. [Google Scholar]
- Lewis, S.M.; Kelly, M. Mapping the potential for biofuel production on marginal lands: Differences in definitions, data and models across scales. ISPRS Int. J. Geo-Inf. 2014, 3, 430–459. [Google Scholar] [CrossRef]
- Scarlat, N.; Banja, M. Possible impact of 2020 bioenergy targets on European Union land use. A scenario-based assessment from national renewable energy action plans proposals. Renew. Sustain. Energy Rev. 2013, 18, 595–606. [Google Scholar]
- Abt, R.C.; Abt, K.L. Potential impact of bioenergy demand on the sustainability of the southern forest resource. J. Sustain. For. 2013, 32, 175–194. [Google Scholar] [CrossRef]
- U.S. Department of Energy. U.S. Billion-Ton Update: Biomass Supply for a Bioenergy and Bioproducts Industry. Available online: http://www1.eere.energy.gov/bioenergy/pdfs/billion_ton_update.pdf (accessed on 17 March 2016).
- Campbell, J.E.; Lobell, D.B.; Genova, R.C.; Field, C.B. The global potential of bioenergy on abandoned agriculture lands. Environ. Sci. Technol. 2008, 42, 5791–5794. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, H.K.; Salmon, J.M. Mapping the world’s degraded lands. Appl. Geogr. 2015, 57, 12–21. [Google Scholar] [CrossRef]
- Eisenbies, M.H.; Vance, E.D.; Aust, W.M.; Seiler, J.R. Intensive utilization of harvest residues in southern pine plantations: Quantities available and implications for nutrient budgets and sustainable site productivity. BioEnerg. Res. 2009, 2, 90–98. [Google Scholar] [CrossRef]
- Hinchee, M.; Rottmann, W.; Mullinax, L.; Zhang, C.; Chang, S.; Cunningham, M.; Pearson, L.; Nehra, N. Short-rotation woody crops for bioenergy and biofuels applications. In Vitro Cell Dev. Biol. Plant 2009, 45, 619–629. [Google Scholar] [CrossRef] [PubMed]
- Zalesny, R.S., Jr.; Cunningham, M.W.; Hall, R.B.; Mirck, J.; Rockwood, D.L.; Stanturf, J.A.; Volk, T.A. Woody Biomass from Short Rotation Energy Crops. In Sustainable Production of Fuels, Chemicals, and fibers from Forest Biomass; Zhu, J., Zhang, X., Pan, X., Eds.; ACS Symposium Series: Washington, DC, USA, 2011; pp. 27–63. [Google Scholar]
- Dickmann, D.I. Silviculture and biology of short-rotation woody crops in temperate regions: Then and now. Biomass Bioenergy 2006, 30, 696–705. [Google Scholar] [CrossRef]
- Heilman, P.E.; Stettler, R.F. Genetic variation and productivity of Populus trichocarpa and its hybrids. II. Biomass production in a 4-year plantation. Can. J. For. Res. 1985, 15, 384–388. [Google Scholar]
- Ceulemans, R.; Scarascia-Mugnozza, G.; Wiard, B.M.; Braatne, J.H.; Hinckley, T.M.; Stettler, R.F.; Heilman, P.E. Production physiology and morphology of Populus species and their hybrids grown under short rotation. I. Clonal comparisons of 4-year growth and phenology. Can. J. For. Res. 1992, 22, 1937–1948. [Google Scholar]
- Heilman, P.E.; Xie, F. Influence of nitrogen on growth and productivity of short-rotation Populus trichocarpa × Populus deltoides hybrids. Can. J. For. Res. 1993, 23, 1863–1869. [Google Scholar] [CrossRef]
- Heilman, P.E.; Ekuan, G.; Fogle, D. Above- and below-ground biomass and fine roots of 4-year-old hybrids of Populus trichocarpa × Populus deltoides and parental species in short-rotation culture. Can. J. For. Res. 1994, 24, 1186–1192. [Google Scholar] [CrossRef]
- Hansen, E.A.; McLaughlin, R.A.; Pope, P.E. Biomass and nitrogen dynamics of hybrid poplar on two different soils: Implications for fertilization strategy. Can. J. For. Res. 1988, 18, 223–230. [Google Scholar] [CrossRef]
- Netzer, D.A.; Tolsted, D.N.; Ostry, M.E.; Isebrands, J.G.; Riemenschneider, D.E.; Ward, K.T. Growth, Yield, and Disease Resistance of 7- to 12-Year-Old Poplar Clones in the North Central United States; General Technical Report GTR-NC-229; Department of Agriculture, Forest Service: St. Paul, MN, USA, 2002. [Google Scholar]
- Zalesny, R.S., Jr.; Hall, R.B.; Zalesny, J.A.; McMahon, B.G.; Berguson, W.E.; Stanosz, G.R. Biomass and genotype × environment interactions of Populus energy crops in the Midwestern United States. Bioenergy Res. 2009, 2, 106–122. [Google Scholar] [CrossRef]
- Czapowskyj, M.M.; Safford, L.O. Site preparation, fertilization, and 10-year yields of hybrid poplar on a clearcut forest site in eastern Maine, USA. New For. 1993, 7, 331–344. [Google Scholar]
- Blow, F.E. Hybrid poplar performance in tests in the Tennessee Valley. J. For. 1948, 46, 493–499. [Google Scholar]
- Blackmon, B.G. Effects of fertilizer nitrogen on tree growth, foliar nitrogen, and herbage in eastern cottonwood plantations. Soil Sci. Soc. Am. J. 1977, 41, 992–995. [Google Scholar] [CrossRef]
- Foster, G.S.; Rousseau, R.J.; Nance, W.L. Eastern cottonwood clonal mixing study: Intergenotypic competition effects. For. Ecol. Manag. 1998, 112, 9–22. [Google Scholar] [CrossRef]
- Cao, Q.V.; Durand, K.M. A growth and yield model for improved eastern cottonwood plantations in the lower Mississippi Delta. South J. Appl. For. 1991, 15, 213–216. [Google Scholar]
- Moreau, B.; Gardiner, E.S.; Stanturf, J.A.; Fisher, R.K. Estimating Leaf Nitrogen of Eastern Cottonwood Trees with a Chlorophyll Meter; General Technical Report SRS-92; Southern research station, Department of Agriculture, Forest Service: Asheville, NC, USA, 2004; pp. 487–491. [Google Scholar]
- Coleman, M.D.; Coyle, D.R.; Blake, J.; Britton, K.; Buford, M.; Campbell, R.G.; Wullschleger, S. Production of Short-Rotation Woody Crops Grown with a Range of Nutrient and Water Availability: Establishment Report and First-Year Responses; General Technical Report SRS-72; Savannah River, Deptartment of Agriculture, Forest Service: New Ellenton, SC, USA, 2004. [Google Scholar]
- Coyle, D.R.; Coleman, M.D.; Durant, J.A.; Newman, L.A. Multiple factors affect pest and pathogen damage on 31 Populus clones in South Carolina. Biomass Bioenergy 2006, 30, 759–768. [Google Scholar] [CrossRef]
- Coyle, D.R.; Coleman, M.D.; Durant, J.A.; Newman, L.A. Survival and growth of 31 Populus clones in South Carolina. Biomass Bioenergy 2006, 30, 750–758. [Google Scholar] [CrossRef]
- Coyle, D.R.; Coleman, M.D. Forest production responses to irrigation and fertilization are not explained by shifts in allocation. For. Ecol. Managy 2005, 208, 137–152. [Google Scholar] [CrossRef]
- Coyle, D.R.; Aubrey, D.P.; Siry, J.P.; Volfovicz-Leon, R.R.; Coleman, M.D. Optimal nitrogen application rates for three intensively-managed hardwood tree species in the southeastern USA. For. Ecol. Manag. 2013, 303, 131–142. [Google Scholar] [CrossRef]
- Kaczmarek, D.J.; Coyle, D.R.; Coleman, M.D. Survival and growth of a range of Populus clones in central South Carolina U.S.A. through age ten: Do early assessments reflect longer-term survival and growth trends? Biomass Bioenergy 2013, 49, 260–272. [Google Scholar]
- Rousseau, R.J.; Adams, J.P.; Wilkerson, D.W. Nine-Year Performance of a Variety of Populus Taxa on an Upland Site in Western Kentucky; General Technical Report SRS-GTR-175; Southern research station, Department of Agriculture, Forest Service: Asheville, NC, USA, 2013; pp. 299–305. [Google Scholar]
- Jeffreys, J.P.; Land, S.B., Jr.; Schultz, E.B.; Londo, A.J. Clonal Tests of New Cottonwood Selections from the Southeast; General Technical Report SRS-92; Southern Research Station, Department of Agriculture, Forest Service: Asheville, NC, USA, 2006; pp. 229–233. [Google Scholar]
- Shifflett, S.D.; Hazel, D.W.; Frederick, D.J.; Nichols, E.G. Species Trials of Short Rotation Woody Crops on Two Wastewater Application Sites in North Carolina, U.S.A. BioEnergy Res. 2014, 7, 157–173. [Google Scholar] [CrossRef]
- Ghezehei, S.B.; Nichols, E.G.; Hazel, D.W. Early Clonal Survival and Growth of Poplars Grown on North Carolina Piedmont and Mountain Marginal Lands. BioEnergy Res. 2015, 1–11. [Google Scholar] [CrossRef]
- Hansen, E.A.; Ostry, M.E.; Johnson, W.D.; Tolsted, D.N.; Netzer, D.A.; Berguson, W.E.; Hall, R.B. Field Performance of Populus in Short-Rotation Intensive Culture Plantations in the North-Central U.S.; Research Paper NC-320; Department of Agriculture, Forest Service: St. Paul, MN, USA, 1994. [Google Scholar]
- Hansen, E.A.; Netzer, D.A.; Woods, R.F. Tillage superior to no-till for establishing hybrid poplar plantations. Tree Plant. Notes 1986, 37, 6–10. [Google Scholar]
- Morris, L.A.; Lowery, R.F. Influence of site preparation on soil conditions affecting stand establishment and tree growth. South J. Appl. For. 1988, 12, 170–178. [Google Scholar]
- Fox, T.R.; Burger, J.A.; Kreh, R.E. Effects of site preparation on nitrogen dynamics in the southern Piedmont. For. Ecol. Manag. 1986, 15, 241–256. [Google Scholar] [CrossRef]
- Kasmioui, O.E.; Ceulemans, R. Financial analysis of the cultivation of short rotation woody crops for bioenergy in Belgium: Barriers and opportunities. BioEnergy Res. 2013, 6, 336–350. [Google Scholar] [CrossRef]
- Van den Berg, A.K.; Perkins, T.D. Evaluation of a portable chlorophyll meter to estimate chlorophyll and nitrogen contents in sugar maple (Acer saccharum Marsh.). leaves. For. Ecol. Manag. 2004, 200, 113–117. [Google Scholar] [CrossRef]
- Chang, S.X.; Robison, D.J. Nondestructive and rapid estimation of hardwood foliar nitrogen status using the SPAD-502 chlorophyll meter. For. Ecol. Manag. 2003, 181, 331–338. [Google Scholar] [CrossRef]
- Pinkard, E.A.; Patel, V.; Mohammed, C. Chlorophyll and nitrogen determination for plantation-grown Eucalyptus nitens and E. globulus using a non-destructive meter. For. Ecol. Manag. 2006, 223, 211–217. [Google Scholar]
- DeBell, D.S.; Clendenen, G.W.; Harrington, C.A.; Zasada, J.C. Tree growth and stand development in short-rotation Populus plantings: 7-year results for two clones at three spacings. Biomass Bioenergy 1996, 11, 253–269. [Google Scholar] [CrossRef]
- Stettler, R.F.; Fenn, R.C.; Heilman, P.E.; Stanton, B.J. Populus trichocarpa × Populus deltoides hybrids for short rotation culture: Variation patterns and 4-year field performance. Can. J. For. Res. 1988, 18, 745–753. [Google Scholar] [CrossRef]
- Zalesny, J.A.; Zalesny, R.S.; Coyle, D.R.; Hall, R.B. Growth and biomass of Populus irrigated with landfill leachate. For. Ecol. Manag. 2007, 248, 143–152. [Google Scholar] [CrossRef]
- Abrahamson, L.P.; White, E.H.; Nowak, C.A.; Briggs, R.D.; Robison, D.J. Evaluating hybrid poplar clonal growth potential in a three-year-old genetic selection field trial. Biomass 1990, 21, 101–114. [Google Scholar] [CrossRef]
- Ares, A. Changes through time in traits of poplar clones in selection trials. New For. 2002, 23, 105–119. [Google Scholar] [CrossRef]
- Zavitkovski, J. Small plots with unplanted plot border can distort data in biomass production studies. Can J. For. Res. 1981, 11, 9–12. [Google Scholar] [CrossRef]
- Schreiner, E. Rating Poplars for Melampsora Leaf Rust Infection; Forest Research Notes NE-90; Department of Agriculture, Forest Service: Minneapolis, MN, USA, 1959. [Google Scholar]
- Avery, T.E.; Burkhart, H.E. Forest Measurements, 5th ed.; Waveland Press, Inc.: Lon Grove, IL, USA, 2002. [Google Scholar]
- Martín-García, J.; Merino, A.; Diez, J.J. Relating visual crown conditions to nutritional status and site quality in monoclonal poplar plantations (Populus × euramericana). Eur. J. For. Res. 2012, 131, 1185–1198. [Google Scholar] [CrossRef]
- DeBell, D.S.; Singleton, R.; Harrington, C.A.; Gartner, B.L. Wood density and fiber length in young Populus stems: Relation to clone, age, growth rate, and pruning. Wood Fiber Sci. 2002, 34, 529–539. [Google Scholar]
- Morhart, C.; Sheppard, J.; Seidl, F.; Spiecker, H. Influence of different tillage systems and weed treatments in the establishment year on the final biomass production of short rotation coppice poplar. Forests 2013, 4, 849–867. [Google Scholar] [CrossRef]
- Ceulemans, R.; Pontailler, J.Y.; Mau, F.; Guittet, J. Leaf allometry in young poplar stands: Reliability of leaf area index estimation, site and clone effects. Biomass Bioenergy 1993, 4, 315–321. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Source of Variation | Height (m) | DBH (cm) | Volume Index (dm3) | LAI | Melampsora Rust Resistance Index | Foliar N (%) | Chlorophyll Content (mg·g−1) |
|---|---|---|---|---|---|---|---|
| p-Value | p-Value | p-Value | p-Value | p-Value | p-Value | p-Value | |
| Soil Tillage | 0.026 | 0.020 | 0.044 | 0.040 | 0.089 | 0.30 | 0.027 |
| Taxa | <0.0001 | <0.0001 | 0.0003 | 0.0013 | <0.0001 | 0.27 | 0.023 |
| Taxa(Genotype) | 0.0002 | 0.0005 | 0.0030 | 0.017 | <0.0001 | 0.060 | 0.19 |
| Soil Tillage × Taxa(Genotype) | 0.86 | 0.80 | 1.0 | 0.96 | 0.55 | 0.36 | 0.79 |
| Taxa | Genotype | Foliar N Mean (1 SD) | Nitrogen Rank | Chlorophyll Content Mean (1 SD) | Chlorophyll Rank | Volume Index Mean (1 SD) | Volume Rank |
|---|---|---|---|---|---|---|---|
| (%) | (mg·g−1) | (dm3) | |||||
| DD | 140 | 2.87% (0.34) | 3 BC | 0.034 (0.005) | 2 A | 34 (20) | 1 A |
| DD | 176 | 2.81% (0.42) | 6 C | 0.029 (0.003) | 4 A | 32 (24) | 4 ABC |
| DD | 356 | 2.68% (0.23) | 9 C | 0.022 (0.005) | 1 A | 30 (21) | 3 AB |
| DD | 373 | 2.82% (0.33) | 4 C | 0.032 (0.010) | 5 A | 19 (20) | 7 CDE |
| DM | 230 | 2.76% (0.33) | 8 C | 0.024 (0.007) | 6 A | 24 (18) | 5 ABC |
| TD | 185 | 2.81% (0.18) | 5 C | 0.030 (0.008) | 10 A | 33 (20) | 2 AB |
| TD | 187 | 3.22% (0.16) | 1 A | 0.024 (0.004) | 9 A | 19 (20) | 10 E |
| TD | 188 | 3.12% (0.44) | 2 AB | 0.025 (0.004) | 8 A | 29 (20) | 9 ED |
| TD | 229 | 2.64% (0.11) | 10 C | 0.028 (0.008) | 7 A | 20 (12) | 6 BCD |
| TD | 339 | 2.80% (0.18) | 7 C | 0.029 (0.008) | 3 A | 12 (10) | 8 ED |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shifflett, S.D.; Hazel, D.W.; Guthrie Nichols, E. Sub-Soiling and Genotype Selection Improves Populus Productivity Grown on a North Carolina Sandy Soil. Forests 2016, 7, 74. https://doi.org/10.3390/f7040074
Shifflett SD, Hazel DW, Guthrie Nichols E. Sub-Soiling and Genotype Selection Improves Populus Productivity Grown on a North Carolina Sandy Soil. Forests. 2016; 7(4):74. https://doi.org/10.3390/f7040074
Chicago/Turabian StyleShifflett, Shawn Dayson, Dennis W. Hazel, and Elizabeth Guthrie Nichols. 2016. "Sub-Soiling and Genotype Selection Improves Populus Productivity Grown on a North Carolina Sandy Soil" Forests 7, no. 4: 74. https://doi.org/10.3390/f7040074