
Aboveground Biomass and Carbon in a South African Mistbelt Forest and the Relationships with Tree Species Diversity and Forest Structures

 ,
,
Abstract
:1. Introduction
- (1)
- Develop three multispecies AGB equations and compare their ability to accurately estimate AGB at the tree level; to do so, we first determined whether wood density and AGB varied among study species. We next tested whether the inclusion of tree height and wood density in biomass equations reduced the estimation error.
- (2)
- Estimate the total amount of biomass and carbon stored in the aboveground compartment in Mistbelt forests; we used the best multispecies AGB equation and the forest inventory data to upscale AGB from the tree to the stand level; we next applied the carbon fraction commonly used in natural forests.
- (3)
- Understand the aboveground carbon (AGC) variation in relation to the forest tree species diversity (richness) and stand characteristics. We assumed that selection effects and dominance patterns are the main drivers of carbon variation.
2. Materials and Methods
2.1. Study Area
2.2. Forest Inventory and Biomass Data
2.3. Assessing the Effect of Species on Wood Density and AGB
2.4. Multispecies DBH-Height and Biomass Allometric Models
2.5. Quantifying AGB and AGC at the Stand Level
2.6. Assessing the Structural Variables Influencing AGC
3. Results
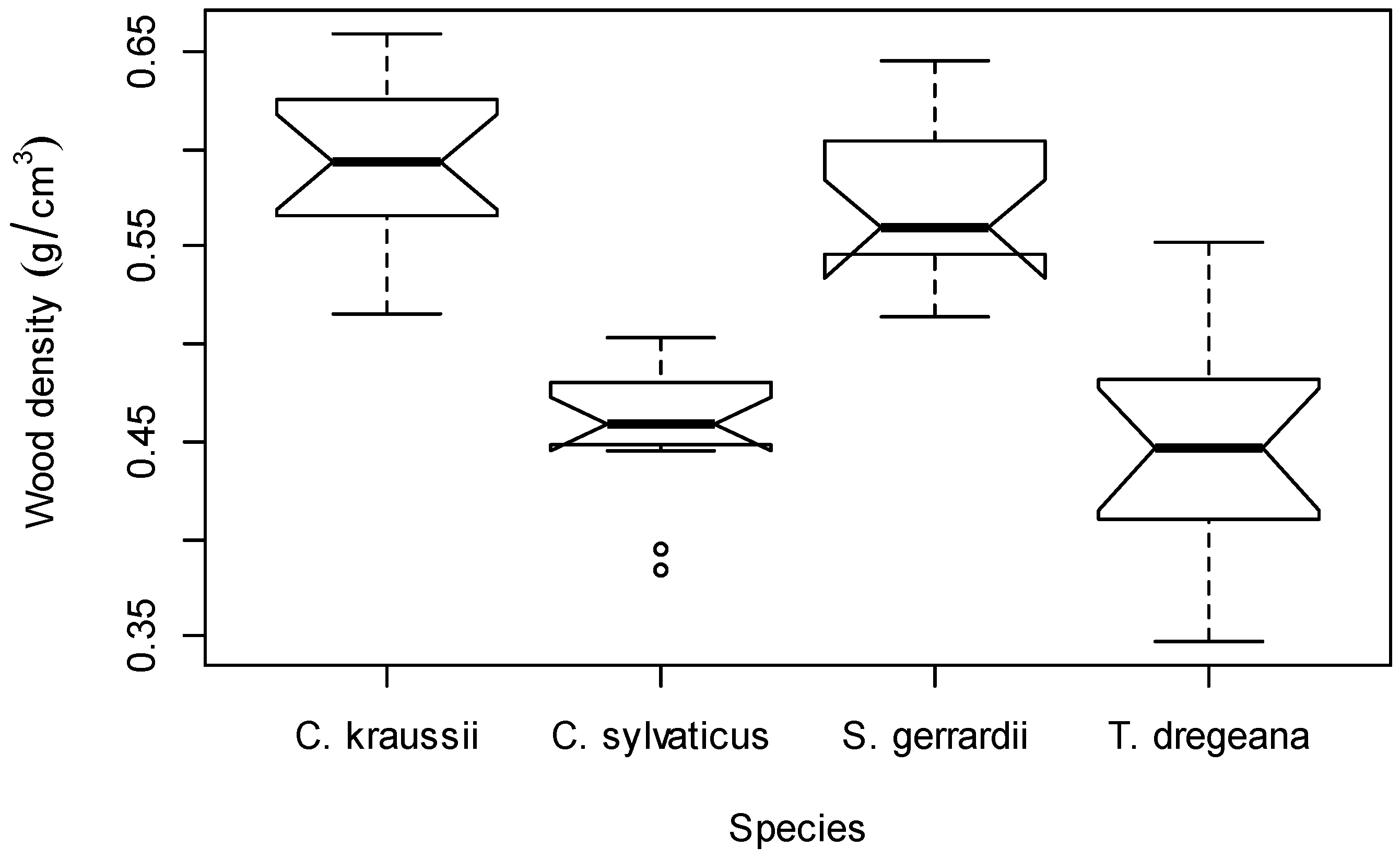
3.1. Effect of Species on Wood Density and AGB
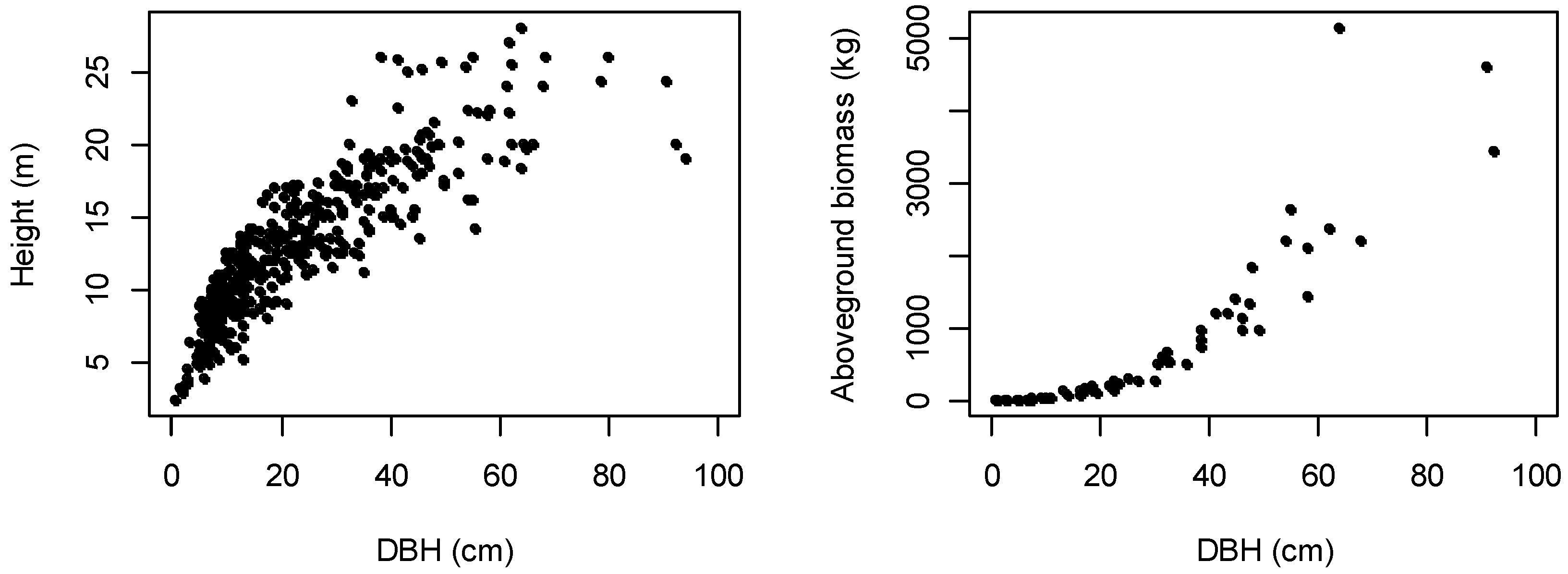
3.2. Multispecies DBH-Height and AGB Allometric Models
3.3. AGB and AGC Stocks at the Stand Level
3.4. Factors Influencing AGC Stocks
4. Discussion
4.1. Effect of Species on Wood Density and AGB
4.2. Uncertainties in the Multispecies AGB Equations
4.3. Predictors for Multispecies AGB Equations
4.4. AGB and AGC Stocks at the Stand Level
4.5. Implications for Landscape Management and Further Perspectives
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AGB | Aboveground biomass |
| AGC | Aboveground carbon |
| DBH | Diameter at breast height |
| H | Total height |
References
- Balvanera, P.; Kremen, C.; Martinez-Ramos, M. Applying community structure analysis to ecosystem function: Examples from pollination and carbon storage. Ecol. Appl. 2005, 15, 360–375. [Google Scholar] [CrossRef]
- Díaz, S.; Cabido, M. Vive la différence: Plant functional diversity matters to ecosystem processes. Trends Ecol. Evol. 2001, 16, 646–655. [Google Scholar] [CrossRef]
- Ruiz-Jaen, M.C.; Potvin, C. Tree diversity explains variation in ecosystem function in a neotropical forest in Panama. Biotropica 2010, 42, 638–646. [Google Scholar] [CrossRef]
- Goulson, D. Foraging strategies of insects for gathering nectar and pollen, and implications for plant ecology and evolution. Perspect. Plant Ecol. Evol. Syst. 1999, 2, 185–209. [Google Scholar] [CrossRef]
- Pywell, R.F.; Warman, E.A.; Carvell, C.; Sparks, T.H.; Dicks, L.V.; Bennett, D.; Wright, A.; Critchley, C.N.R.; Sherwood, A. Providing foraging resources for bumblebees in intensively farmed landscapes. Biol. Conserv. 2005, 121, 479–494. [Google Scholar] [CrossRef]
- Jha, S.; Kremen, C. Resource diversity and landscape-level homogeneity drive native bee foraging. Proc. Natl. Acad. Sci. USA 2013, 110, 555–558. [Google Scholar] [CrossRef] [PubMed]
- Saatchi, S.S.; Harris, N.L.; Brown, S.; Lefsky, M.; Mitchard, E.T.A.; Salas, W.; Zutta, B.R.; Buermann, W.; Lewis, S.L.; Hagen, S.; et al. Benchmark map of forest carbon stocks in tropical regions across three continents. Proc. Natl. Acad. Sci. USA 2011, 108, 9899–9904. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–994. [Google Scholar] [CrossRef] [PubMed]
- Beer, C.; Reichstein, M.; Tomelleri, E.; Ciais, P.; Jung, M.; Carvalhais, N.; Rödenbeck, C.; Arain, M.A.; Baldocchi, D.; Bonan, G.B.; et al. Terrestrial gross carbon dioxide Uptake: Global Distribution and Covariation with Climate. Science 2010, 329, 834–839. [Google Scholar] [CrossRef] [PubMed]
- MEA (Millenium Ecosystem Assessment). Ecosystems and Human Well-Being: Current State and Trends; Island Press: Washington, DC, USA, 2005; p. 47. [Google Scholar]
- Chave, J.; Andalo, C.; Brown, S.; Cairns, M.A.; Chambers, J.Q.; Eamus, D.; Fölster, H.; Fromard, F.; Higuchi, N.; Kira, T.; et al. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 2005, 145, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Chave, J.; Réjou-Méchain, M.; Búrquez, A.; Chidumayo, E.; Colgan, M.S.; Delitti, W.B.C.; Duque, A.; Eid, T.; Fearnside, P.M.; Goodman, R.C.; et al. Improved allometric models to estimate the aboveground biomass of tropical trees. Glob. Chang. Biol. 2014, 20, 3177–3190. [Google Scholar] [CrossRef] [PubMed]
- Mucina, L.; Rutherford, M.C. The Vegetation of South Africa, Lesotho and Swaziland; Mucina, L., Rutherford, M.C., Eds.; South African National Biodiversity Institute: Pretoria, South Africa, 2006. [Google Scholar]
- Geldenhuys, C.J. Tropical Secondary Forest Management in Africa: Reality and Perspectives; Food and Agriculture Organization: Rome, Italy, 2002. [Google Scholar]
- Jose, S.; Bardhan, S. Agroforestry for biomass production and carbon sequestration: An overview. Agrofor. Syst. 2012, 86, 105–111. [Google Scholar] [CrossRef]
- Dovey, S.B. Estimating biomass and macronutrient content of some commercially important plantation species in South Africa. South. For. 2009, 71, 245–251. [Google Scholar] [CrossRef]
- Phiri, D.; Ackerman, P.; Wessels, B.; du Toit, B.; Johansson, M.; Säll, H.; Lundqvist, S.-O.; Seifert, T. Biomass equations for selected drought-tolerant eucalypts in South Africa. South. For. 2015, 77, 255–262. [Google Scholar] [CrossRef]
- Du Toit, B. Effects of site management on growth, biomass partitioning and light use efficiency in a young stand of Eucalyptus grandis in South Africa. For. Ecol. Manag. 2008, 255, 2324–2336. [Google Scholar] [CrossRef]
- Colgan, M.S.; Asner, G.P.; Swemmer, T. Harvesting tree biomass at the stand level to assess the accuracy of field and airborne biomass estimation in savannas. Ecol. Appl. 2013, 23, 1170–1184. [Google Scholar] [CrossRef] [PubMed]
- Lung, M.; Espira, A. The influence of stand variables and human use on biomass and carbon stocks of a transitional African forest: Implications for forest carbon projects. For. Ecol. Manag. 2015, 351, 36–46. [Google Scholar] [CrossRef]
- Magalhães, T.M.; Seifert, T. Estimation of tree biomass, carbon stocks, and error propagation in Mecrusse Woodlands. Open J. For. 2015, 5, 471–488. [Google Scholar] [CrossRef]
- Seifert, T.; Seifert, S. Modelling and simulation of tree biomass. In Bioenergy from Wood; Seifert, T., Ed.; Springer Netherlands: Dordrecht, The Netherlands, 2014; pp. 43–65. [Google Scholar]
- Kunneke, A.; van Aardt, J.; Roberts, W.; Seifert, T. Localisation of biomass potentials. In Bioenergy from Wood; Seifert, T., Ed.; Springer Netherlands: Dordrecht, The Netherlands, 2014; pp. 11–41. [Google Scholar]
- Picard, N.; Rutishauser, E.; Ploton, P.; Ngomanda, A.; Henry, M. Should tree biomass allometry be restricted to power models? For. Ecol. Manag. 2015, 353, 156–163. [Google Scholar] [CrossRef]
- Day, M.; Baldauf, C.; Rutishauser, E.; Sunderland, T.C.H. Relationships between tree species diversity and above-ground biomass in Central African rainforests: Implications for REDD. Environ. Conserv. 2014, 41, 64–72. [Google Scholar] [CrossRef]
- Sharma, C.M.; Baduni, N.P.; Gairola, S.; Ghildiyal, S.K.; Suyal, S. Tree diversity and carbon stocks of some major forest types of Garhwal Himalaya, India. For. Ecol. Manag. 2010, 260, 2170–2179. [Google Scholar] [CrossRef]
- Poorter, L.; van der Sande, M.T.; Thompson, J.; Arets, E.J.M.M.; Alarcón, A.; Álvarez-Sánchez, J.; Ascarrunz, N.; Balvanera, P.; Barajas-Guzmán, G.; Boit, A.; et al. Diversity enhances carbon storage in tropical forests. Glob. Ecol. Biogeogr. 2015, 24, 1314–1328. [Google Scholar] [CrossRef]
- Ruiz-Jaen, M.C.; Potvin, C. Can we predict carbon stocks in tropical ecosystems from tree diversity? Comparing species and functional diversity in a plantation and a natural forest. New Phytol. 2011, 189, 978–987. [Google Scholar] [CrossRef] [PubMed]
- Cavanaugh, K.C.; Gosnell, J.S.; Davis, S.L.; Ahumada, J.; Boundja, P.; Clark, D.B.; Mugerwa, B.; Jansen, P.A.; O’Brien, T.G.; Rovero, F.; et al. Carbon storage in tropical forests correlates with taxonomic diversity and functional dominance on a global scale. Glob. Ecol. Biogeogr. 2014, 23, 563–573. [Google Scholar] [CrossRef]
- White, F. The Vegetation Map of Africa; Unesco: Paris, France, 1983. [Google Scholar]
- Cooper, K.H. The Conservation Status of Indigenous Forests in Transvaal, Natal, and O.F.S., South Africa; Wildlife Society of South Africa, Conservation Division: Durban, South Africa, 1985. [Google Scholar]
- Van Laar, A.; Akça, A. Forest Mensuration; Springer Netherlands: Dordrecht, The Netherlands, 2007. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer New York: New York, NY, USA, 2009. [Google Scholar]
- Sileshi, G.W. A critical review of forest biomass estimation models, common mistakes and corrective measures. For. Ecol. Manag. 2014, 329, 237–254. [Google Scholar] [CrossRef]
- Fayolle, A.; Doucet, J.-L.; Gillet, J.-F.; Bourland, N.; Lejeune, P. Tree allometry in Central Africa: Testing the validity of pantropical multi-species allometric equations for estimating biomass and carbon stocks. For. Ecol. Manag. 2013, 305, 29–37. [Google Scholar] [CrossRef]
- Dumont, C.; Mentré, F.; Gaynor, C.; Brendel, K.; Gesson, C.; Chenel, M. Optimal sampling times for a drug and its metabolite using SIMCYP(®) simulations as prior information. Clin. Pharmacokinet. 2013, 52, 43–57. [Google Scholar] [CrossRef] [PubMed]
- McCune, B.; Grace, J. Analysis of Ecological Communities; MjM Software Design: Gleneden Beach, OR, USA, 2002. [Google Scholar]
- Baskerville, G.L. Use of logarithmic regression in the estimation of plant biomass. Can. J. For. Res. 1972, 2, 49–53. [Google Scholar] [CrossRef]
- Zanne, A.E.; Lopez-Gonzalez, G.; Coomes, D.A.; Ilic, J.; Jansen, S.; Lewis, S.L.; Miller, R.B.; Swenson, N.G.; Wiemann, M.C.; Chave, J. Data from: Towards a worldwide wood economics spectrum. Dryad Digit. Repos. 2009. [Google Scholar] [CrossRef]
- Chave, J.; Coomes, D.; Jansen, S.; Lewis, S.L.; Swenson, N.G.; Zanne, A.E. Towards a worldwide wood economics spectrum. Ecol. Lett. 2009, 12, 351–366. [Google Scholar] [CrossRef] [PubMed]
- Stegen, J.C.; Swenson, N.G.; Valencia, R.; Enquist, B.J.; Thompson, J. Above-ground forest biomass is not consistently related to wood density in tropical forests. Glob. Ecol Biogeogr. 2009, 18, 617–625. [Google Scholar] [CrossRef]
- Reque, J.; Bravo, F. Identifying forest structure types using National Forest Inventory Data: The case of sessile oak forest in the Cantabrian range. Investig. Agrar. Sist. Recur. For. 2008, 17, 105–113. [Google Scholar] [CrossRef]
- Mensah, S.; Houehanou, T.D.; Sogbohossou, E.A.; Assogbadjo, A.E.; Glèlè Kakaï, R. Effect of human disturbance and climatic variability on the population structure of Afzelia africana Sm. ex pers. (Fabaceae–Caesalpinioideae) at country broad-scale (Bénin, West Africa). S. Afr. J. Bot. 2014, 95, 165–173. [Google Scholar] [CrossRef]
- Curtis, J.T.; McIntosh, R.P. An upland forest continuum in the prairie-forest border region of Wisconsin. Ecology 1951, 32, 476–496. [Google Scholar] [CrossRef]
- Chave, J.; Muller-Landau, H.C.; Baker, T.R.; Easdale, T.A.; ter Steege, H.; Webb, C.O. Regional and phylogenetic variation of wood density across 2456 neotropical tree species. Ecol. Appl. 2006, 16, 2356–2367. [Google Scholar] [CrossRef]
- Wright, S.J.; Kitajima, K.; Kraft, N.J.B.; Reich, P.B.; Wright, I.J.; Bunker, D.E.; Condit, R.; Dalling, J.W.; Davies, S.J.; Díaz, S.; et al. Functional traits and the growth-mortality trade-off in tropical trees. Ecology 2010, 91, 3664–3674. [Google Scholar] [CrossRef] [PubMed]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef] [PubMed]
- Mensah, S.; Glèlè Kakaï, R.; Seifert, T. Patterns of biomass allocation between foliage and woody structure: The effects of tree size and specific functional traits. Ann. For. Res. 2016, in press. [Google Scholar]
- Ebuy, J.; Lokombe, J.P.; Ponette, Q.; Sonwa, D.; Picard, N. Allometric equation for predicting aboveground biomass of three tree species. J. Trop. For. Sci. 2011, 23, 125–132. [Google Scholar]
- Deans, J.D.; Moran, J.; Grace, J. Biomass relationships for tree species in regenerating semi-deciduous tropical moist forest in Cameroon. For. Ecol. Manag. 1996, 88, 215–225. [Google Scholar] [CrossRef]
- Henry, M.; Besnard, A.; Asante, W.A.; Eshun, J.; Adu-Bredu, S.; Valentini, R.; Bernoux, M.; Saint-André, L. Wood density, phytomass variations within and among trees, and allometric equations in a tropical rainforest of Africa. For. Ecol. Manag. 2010, 260, 1375–1388. [Google Scholar] [CrossRef]
- Segura, M.; Kanninen, M. Allometric models for tree volume and total aboveground biomass in a tropical humid forest in Costa Rica. Biotropica 2005, 37, 2–8. [Google Scholar] [CrossRef]
- Ngomanda, A.; Engone Obiang, N.L.; Lebamba, J.; Moundounga Mavouroulou, Q.; Gomat, H.; Mankou, G.S.; Loumeto, J.; Midoko Iponga, D.; Kossi Ditsouga, F.; Zinga Koumba, R.; et al. Site-specific versus pantropical allometric equations: Which option to estimate the biomass of a moist central African forest? For. Ecol. Manag. 2014, 312, 1–9. [Google Scholar] [CrossRef]
- Henry, M.; Picard, N.; Trotta, C.; Manlay, R.J.; Valentini, R.; Bernoux, M.; Saint-André, L. Estimating tree biomass of sub-Saharan African forests: A review of available allometric equations. Silva Fenn. 2011, 45, 477–569. [Google Scholar] [CrossRef]
- Garber, S.M.; Temesgen, H.; Monleon, V.J.; Hann, D.W. Effects of height imputation strategies on stand volume estimation. Can. J. For. Res. 2009, 39, 681–690. [Google Scholar] [CrossRef]
- Temesgen, H.; Affleck, D.; Poudel, K.; Gray, A.; Sessions, J. A review of the challenges and opportunities in estimating above ground forest biomass using tree-level models. Scand. J. For. Res. 2015, 30, 326–335. [Google Scholar] [CrossRef]
- Falster, D.S. Sapling strength and safety: The importance of wood density in tropical forests. New Phytol. 2006, 171, 237–239. [Google Scholar] [CrossRef] [PubMed]
- Preece, N.D.; Lawes, M.J.; Rossman, A.K.; Curran, T.J.; van Oosterzee, P. Modelling the growth of young rainforest trees for biomass estimates and carbon sequestration accounting. For. Ecol. Manag. 2015, 351, 57–66. [Google Scholar] [CrossRef]
- Suzuki, E. Diversity in specific gravity and water content of wood among Bornean tropical rainforest trees. Ecol. Res. 1999, 14, 211–224. [Google Scholar] [CrossRef]
- Houghton, R.A.; Lawrence, K.T.; Hackler, J.L.; Brown, S. The spatial distribution of forest biomass in the Brazilian Amazon: A comparison of estimates. Glob. Chang. Biol. 2001, 7, 731–746. [Google Scholar] [CrossRef]
- Lewis, S.L.; Sonké, B.; Sunderland, T.; Begne, S.K.; Lopez-Gonzalez, G.; van der Heijden, G.M.F.; Phillips, O.L.; Affum-Baffoe, K.; Baker, T.R.; Banin, L.; et al. Above-ground biomass and structure of 260 African tropical forests. Philos. Trans. R. Soc. B 2013, 368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, R.; Ensslin, A.; Rutten, G.; Fischer, M.; Schellenberger Costa, D.; Kleyer, M.; Hemp, A.; Paulick, S.; Huth, A. Simulating carbon stocks and fluxes of an African Tropical Montane Forest with an Individual-Based Forest Model. PLoS ONE 2015, 10, e0123300. [Google Scholar] [CrossRef] [PubMed]
- Baccini, A.; Laporte, N.; Goetz, S.J.; Sun, M.; Dong, H. A first map of tropical Africa’s above-ground biomass derived from satellite imagery. Environ. Res. Lett. 2008, 3. [Google Scholar] [CrossRef]
- Geldenhuys, C.J. Native forest regeneration in pine and eucalypt plantations in Northern Province, South Africa. For. Ecol. Manag. 1997, 99, 101–115. [Google Scholar] [CrossRef]
- Wu, X.; Wang, X.; Tang, Z.; Shen, Z.; Zheng, C.; Xia, X.; Fang, J. The relationship between species richness and biomass changes from boreal to subtropical forests in China. Ecography 2015, 38, 602–613. [Google Scholar] [CrossRef]
- Ruiz-Benito, P.; Gómez-Aparicio, L.; Paquette, A.; Messier, C.; Kattge, J.; Zavala, M.A. Diversity increases carbon storage and tree productivity in Spanish forests. Glob. Ecol. Biogeogr. 2014, 23, 311–322. [Google Scholar] [CrossRef]
- Tilman, D.; Knops, J.; Wedin, D.; Reich, P.; Ritchie, M.; Siemann, E. The influence of functional diversity and composition on ecosystem processes. Science 1997, 277, 1300–1302. [Google Scholar] [CrossRef]
- Chave, J.; Condit, R.; Lao, S.; Caspersen, J.P.; Foster, R.B.; Hubbell, S.P. Spatial and temporal variation of biomass in a tropical forest: Results from a large census plot in Panama. J. Ecol. 2003, 91, 240–252. [Google Scholar] [CrossRef]
- Slik, J.W.F.; Aiba, S.I.; Brearley, F.Q.; Cannon, C.H.; Forshed, O.; Kitayama, K.; Nagamasu, H.; Nilus, R.; Payne, J.; Paoli, G.; et al. Environmental correlates of tree biomass, basal area, wood specific gravity and stem density gradients in Borneo’s tropical forests. Glob. Ecol. Biogeogr. 2010, 19, 50–60. [Google Scholar] [CrossRef]
- Seifert, T.; Seifert, S.; Seydack, A.; Durheim, G.; von Gadow, K. Competition effects in an afrotemperate forest. For. Ecosyst. 2014, 1. [Google Scholar] [CrossRef]
- Hernández-Barrios, J.C.; Anten, N.P.R.; Martínez-Ramos, M. Sustainable harvesting of non-timber forest products based on ecological and economic criteria. J. Appl. Ecol. 2014, 52, 389–401. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Number of Trees | DBH (cm) | Height (m) | Wood Density (g/cm3) | AGB (Kg) |
|---|---|---|---|---|---|
| C. kraussii | 16 | 1.5–91.0 | 3.1–24.2 | 0.51–0.66 | 0.26–4590.19 |
| C. sylvaticus | 14 | 4.8–64.0 | 5.4–28.0 | 0.38–0.50 | 4.17–5127.94 |
| S. gerrardii | 15 | 0.7–92.5 | 2.3–22.1 | 0.51–0.65 | 0.17–3423.33 |
| T. dregeana | 14 | 2.8–62.0 | 4.4–27.0 | 0.35–0.55 | 0.82–2357.97 |
| Estimate | Std. Error | t Value | Pr(>|t|) | ||
|---|---|---|---|---|---|
| (Intercept) | −1.963 | 0.252 | −7.791 | 0.000 | |
| Tree size | ln (DBH) | 2.365 | 0.079 | 30.036 | <0.001 |
| Species | C. sylvaticus | −0.958 | 0.439 | −2.183 | 0.034 |
| S. gerrardii | 0.403 | 0.322 | 1.254 | 0.215 | |
| T. dregeana | −0.465 | 0.402 | −1.155 | 0.253 | |
| Tree size: Species | ln (DBH): C. sylvaticus | 0.309 | 0.135 | 2.288 | 0.026 |
| ln (DBH): S. gerrardii | −0.132 | 0.101 | −1.305 | 0.198 | |
| ln (DBH): T. dregeana | 0.081 | 0.125 | 0.644 | 0.523 |
| Models | Predictors | Parameter | Estimate | SE | p | R2 | VIF | RSE | RMSE | MAD | AIC | CF | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Height | Equation (2) | Intercept | ln (β0) | 1.01 | 0.03 | <0.001 | 83.81 | - | 0.181 | - | - | - | 1.016 |
| DBH | β1 | 0.51 | 0.01 | <0.001 | |||||||||
| AGB | Equation (3) | Intercept | ln (β0) | −1.89 | 0.25 | <0.001 | 97.60 | - | 0.304 | 0.296 | 6.78 | 30.490 | 1.047 |
| Wood density | β1 | 0.37 | 0.26 | 0.159 | 1.005 | ||||||||
| DBH | β2 | 2.41 | 0.05 | <0.001 | 1.005 | ||||||||
| Equation (4) | Intercept | ln (β0) | −2.84 | 0.27 | <0.001 | 98.44 | 0.245 | 0.236 | 4.75 | 7.230 | 1.030 | ||
| Wood density | β1 | 0.75 | 0.22 | 0.001 | 1.115 | ||||||||
| DBH | β2 | 1.81 | 0.12 | <0.001 | 8.105 | ||||||||
| Height | β3 | 1.14 | 0.21 | <0.001 | 8.086 | ||||||||
| Equation (5) | Intercept | ln (β0) | −2.69 | 0.21 | <0.001 | 98.45 | 0.244 | 0.237 | 3.32 | 6.074 | 1.030 | ||
| Wood density | β1 | 0.69 | 0.21 | 0.002 | 1.002 | ||||||||
| DBH2 × Height | β2 | 0.95 | 0.02 | <0.001 | 1.002 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mensah, S.; Veldtman, R.; Du Toit, B.; Glèlè Kakaï, R.; Seifert, T. Aboveground Biomass and Carbon in a South African Mistbelt Forest and the Relationships with Tree Species Diversity and Forest Structures. Forests 2016, 7, 79. https://doi.org/10.3390/f7040079
Mensah S, Veldtman R, Du Toit B, Glèlè Kakaï R, Seifert T. Aboveground Biomass and Carbon in a South African Mistbelt Forest and the Relationships with Tree Species Diversity and Forest Structures. Forests. 2016; 7(4):79. https://doi.org/10.3390/f7040079
Chicago/Turabian StyleMensah, Sylvanus, Ruan Veldtman, Ben Du Toit, Romain Glèlè Kakaï, and Thomas Seifert. 2016. "Aboveground Biomass and Carbon in a South African Mistbelt Forest and the Relationships with Tree Species Diversity and Forest Structures" Forests 7, no. 4: 79. https://doi.org/10.3390/f7040079