
Changes in Structure and Diversity of Woody Plants in a Secondary Mixed Pine-Oak Forest in the Sierra Madre del Sur of Mexico

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sample Sites
2.3. Vegetation Sampling
2.4. Statistical Analysis
3. Results
3.1. Floristic Composition
3.2. Species Richness, Abundance, and Diversity
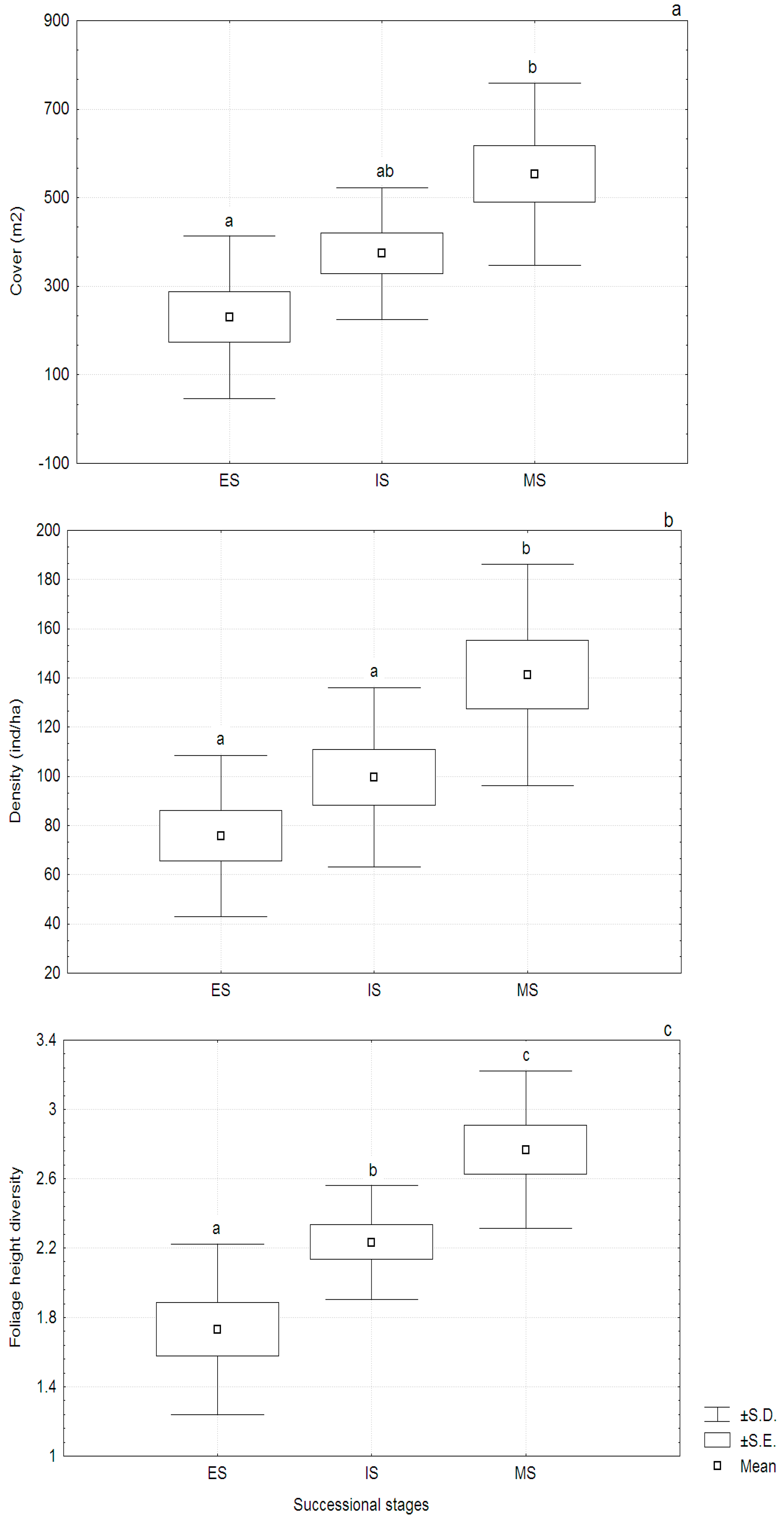
3.3. Vegetation Structure
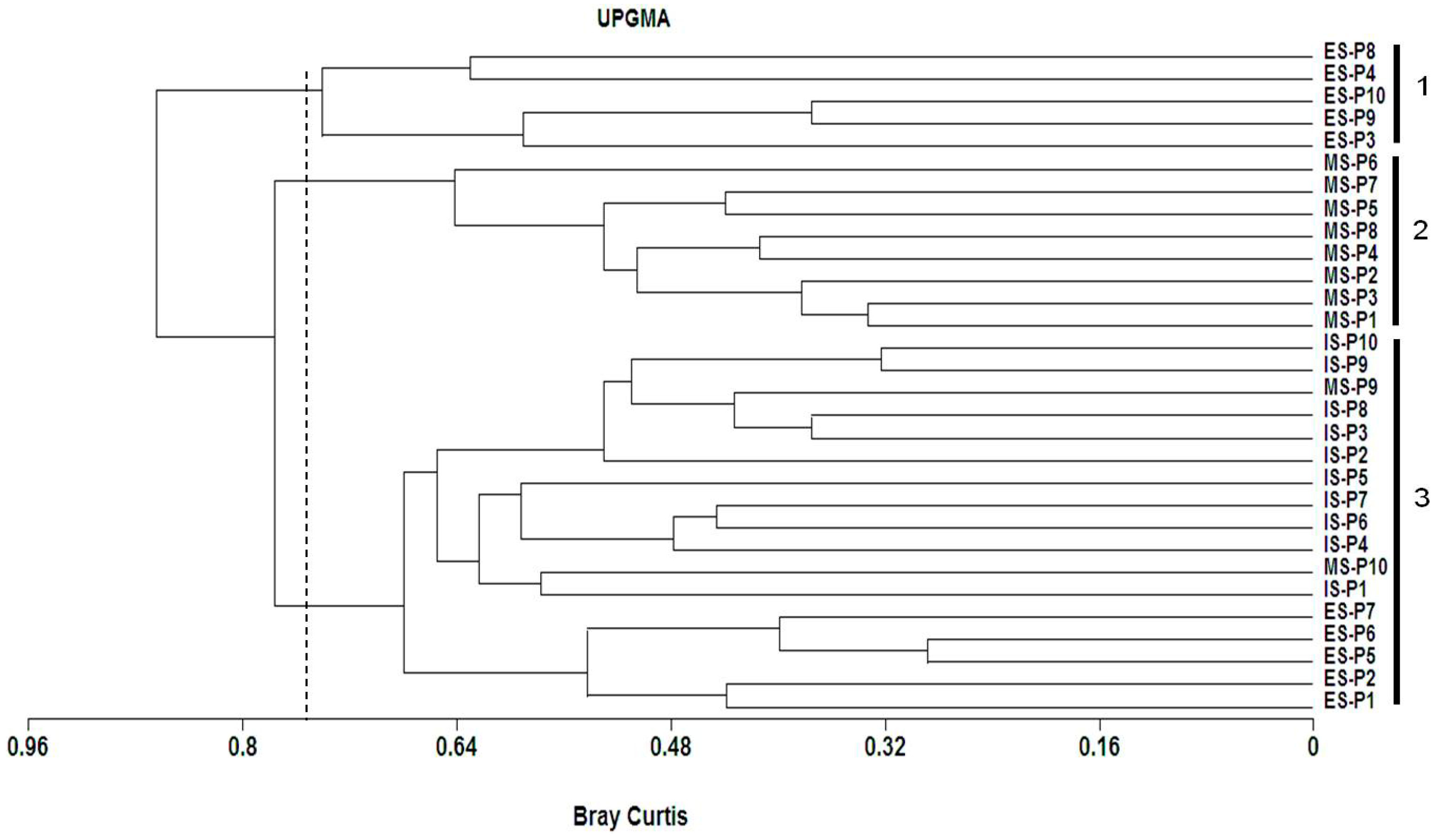
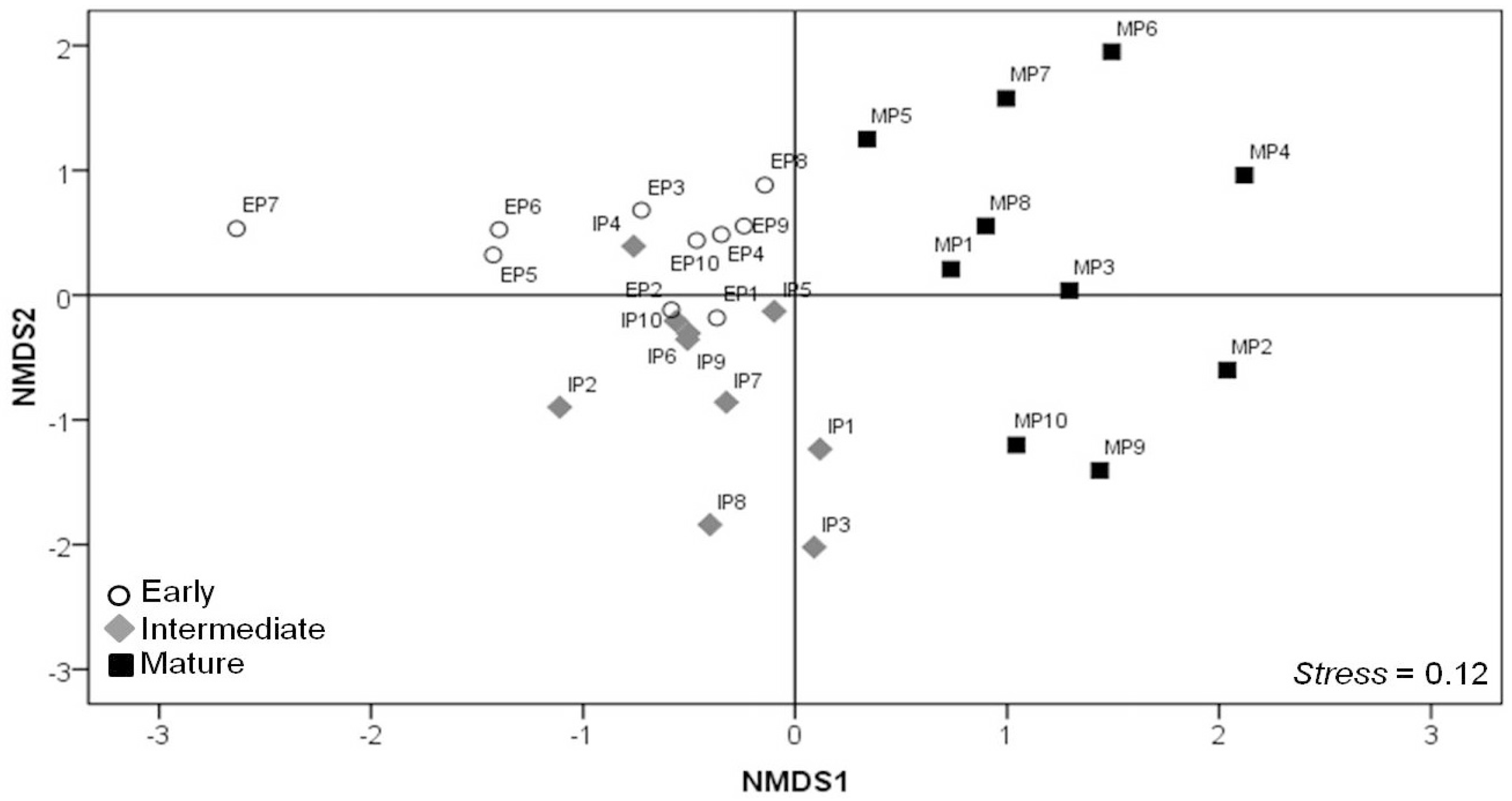
3.4. Spatial Distribution of the Plant Species
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix 1

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Species | Life Form | ES | IS | MS |
|---|---|---|---|---|---|
| Anacardiaceae | Rhus spp. L. 1753 | Tree | x | ||
| Actinocheita potentillifolia (Turcz.) Bullock 1937 | Shrub | x | |||
| Apocynaceae | Vallesia glabra (Cav.) Link 1821 | Shrub | x | ||
| Arecaceae | Brahea dulcis (Kunth) Mart. 1838 | Tree | x | x | |
| Asteraceae | Brickellia spp. | Shrub | x | x | |
| Baccharis heterophylla Kunth 1820 | Shrub | x | |||
| Betulaceae | Carpinus caroliniana Walter 1788 | Tree | x | ||
| Alnus acuminata Kunth 1817 | Tree | x | x | ||
| Burseraceae | Bursera cuneata (Schltdl.) Engl. 1880 | Tree | x | ||
| Cactaceae | Opuntia sp. Mill. 1754 | Shrub | x | ||
| Clethraceae | Clethra mexicana DC. 1839 | Tree | x | x | |
| Compositae | Vernonia alamanii DC.1836 | Shrub | x | ||
| Montanoa grandiflora DC. 1836 | Shrub | x | x | ||
| Piqueria trinervia Cav. 1794 | Shrub | x | |||
| Coreopsis mutica DC. 1836 | Shrub | x | x | x | |
| Cupressaceae | Cupressus lindleyi Klotzsch ex Endl. 1847 | Tree | x | x | |
| Cupressus lusitanica Mill. 1768 | Tree | x | |||
| Juniperus flaccida Schltdl. 1838 | Tree | x | |||
| Ericaceae | Arbutus xalapensis Kunth 1818 | Tree | x | x | x |
| Arctostaphylos discolor (Hook.) DC.1839 | Shrub | x | x | ||
| Fabaceae | Leucaena leucocephala (Lam.) de Wit 1961 | Tree | x | ||
| Eysenhardtia polystachia (Ortega) Sarg.1892 | Shrub | x | |||
| Senna multiglandulosa (Jacq.) H.S. Irwin & Barneby 1982 | Shrub | x | x | ||
| Mimosa tenuiflora Benth 1846 | Tree | x | |||
| Acacia farnesiana (L.) Willd. 1806 | Tree | x | x | x | |
| Acacia pennatula (Schltdl. & Cham.) Benth. 1842 | Tree | x | x | ||
| Dalea lutea (Cav.) Willd. 1802 | Shrub | x | |||
| Calliandra grandiflora (L'Hér.) Benth. 1840 | Shrub | x | x | ||
| Fagaceae | Quercus acutifolia Née 1801 | Tree | x | x | |
| Quercus candicans Née 1801 | Tree | x | |||
| Quercus castanea Née 1801 | Tree | x | |||
| Quercus crassifolia Benth. 1840 | Tree | x | |||
| Quercus liebmannii Oerst. ex Trel. 1924 | Tree | x | x | x | |
| Quercus magnoliifolia Née 1801 | Tree | x | |||
| Quercus spp. L. 1753 | Tree | x | |||
| Quercus elliptica Née 1801 | Tree | x | x | ||
| Quercus martinezii C.H. Mull.1954 | Tree | x | x | ||
| Quercus glaucescens Bonpl. 1809 | Tree | x | x | ||
| Flacourtiaceae | Xylosma flexuosum (Kunth) Hemsley 1879 | Shrub | x | ||
| Labiatae | Salvia sessei Benth. 1833 | Shrub | x | ||
| Litsea glaucescens Kunth 1817 | Shrub | x | x | ||
| Myricaceae | Myrica cerifera L. 1753 | Shrub | x | ||
| Pinaceae | Pinus lawsonii Roezl ex Gordon 1862 | Tree | x | x | x |
| Pinus pringlei Shaw 1905 | Tree | x | x | x | |
| Pinus oocarpa Schiede ex Schltdl. 1838 | Tree | x | x | ||
| Pinus pseudostrobus Lindl. 1839 | Tree | x | x | x | |
| Pinus montezumae Gordon & Glend. 1858 | Tree | x | x | ||
| Rosaceae | Crataegus pubescens (C. Presl) C. Presl 1826 | Tree | x | ||
| Prunus serotina Ehrh. 1783 | Tree | x | |||
| Pyrus communis L. 1753 | Tree | x | |||
| Cercocarpus fothergilloides Kunth 1823 | Tree | x | x | ||
| Rubiaceae | Randia echinocarpa DC. 1830 | Tree | x | ||
| Solanaceae | Datura candida Voigt 1845 | Shrub | x |
References
- Nave, L.E.; Vance, E.D.; Swanston, C.W.; Curtis, P.S. Harvest impacts on soil carbon storage in temperate forest. For. Ecol. Manag. 2010, 259, 857–866. [Google Scholar] [CrossRef]
- Price, N.W.; Gratzer, G.; Alemayehu-Duguma, L.; Kohler, T.; Maselli, D.; Romero, R. Mountain Forests in a Changing World—Realizing Values, Addressing Challenges; Food and Agriculture Organization of the United Nations-Swiss Agency for Development and Cooperation: Roma, Italy, 2011. [Google Scholar]
- Seppelt, R.; Dormann, C.F.; Eppink, F.V.; Lautenbach, S.; Schmidt, S. A quantitative review of ecosystem service studies: Approaches, shortcomings and the road ahead. J. Appl. Ecol. 2011, 48, 630–636. [Google Scholar] [CrossRef]
- González-Espinosa, M.; Ramírez-Marcial, N.; Galindo-Jaimes, L. Secondary succession in montane pine-oak forests of Chiapas, Mexico. In Ecology and Conservation of Neotropical Montane Oak Forests, Ecological Studies 185; Kapelle, M., Ed.; Springer-Verlag: Heidelberg/Berlin, Germany, 2006; pp. 209–221. [Google Scholar]
- Fregoso, A.; Velázquez, A.; Bocco, G.; Cortéz, G. El enfoque de paisaje en el manejo forestal de la comunidad indígena de Nuevo San Juan Parangaricutiro, Michoacán, México. Investig. Geogr. 2001, 46, 58–77. [Google Scholar]
- Petruccelli, C.A.; Sakulich, J.; Harley, G.L.; Grissino-Mayer, H.G. Structure and dynamics of an old-growth pine-oak community in the southern Appalachian mountains, Georgia, USA. Southeast. Geogr. 2014, 54, 161–182. [Google Scholar] [CrossRef]
- González-Espinosa, M.; Quintana-Ascensio, P.F.; Ramírez-Marcial, N.; Gaytan-Guzmán, P. Secondary succession in disturbed Pinus-Quercus forests in the Highlands of Chiapas, Mexico. J. Veg. Sci. 1991, 2, 351–360. [Google Scholar] [CrossRef]
- Rigling, A.; Bigler, C.; Eilmann, B.; Feldmeyer-Christe, E.; Gimmi, U.; Ginzler, C.; Graf, U.; Mayer, P.; Vacchiano, G.; Weber, P.; et al. Driving factors of a vegetation shift from Scots pine to pubescent oak in dry Alpine forest. Glob. Change. Biol. 2013, 19, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Wang, D.; Shi, X.; Yi, X.; Huang, Q.; Hu, Y. Effects of environmental factors on tree seedling regeneration in a pine-oak mixed forest in the Qinling Mountains, China. J. Mt. Sci. 2013, 10, 845–853. [Google Scholar] [CrossRef]
- Ochoa-Gaona, S.; González-Espinosa, M. Land-use and deforestation in the highlands of Chiapas, Mexico. Appl. Geogr. 2000, 20, 17–42. [Google Scholar] [CrossRef]
- Hao, Z.; Zhang, J.; Song, B.; Ye, J.; Li, B. Vertical structure and spatial associations of dominant tree species in an old-growth temperate forest. For. Ecol. Manag. 2007, 252, 1–11. [Google Scholar] [CrossRef]
- Álvarez-Yépiz, J.C.; Martínez-Yrizar, A.; Burquez, A.; Lindquist, C. Variation in vegetation structure and soil properties related to land use history of old-growth and secondary tropical dry forest in northwest Mexico. For. Ecol. Manag. 2008, 256, 355–366. [Google Scholar] [CrossRef]
- Wilms, J.J.A.; Kapelle, M. Frugivorous birds, hábitat preference and seed dispersal in a fragmented Costa Rican montane oak forest landscape. In Ecology and Conservation of Neotropical Montane Oak Forest, Ecological Studies 185; Kappelle, M., Ed.; Springer-Verlag: Heidelberg/Berlin, Germany, 2006; pp. 309–324. [Google Scholar]
- Rey-Benayas, J.M.; Newton, A.C.; Díaz, A.; Bullock, J.M. Enhancement of biodiversity and ecosystem services by ecological restoration: A meta-analysis. Science 2009, 325, 1121–1124. [Google Scholar] [CrossRef] [PubMed]
- Prach, K.; Walker, L.R. Four opportunities for studies of ecological succession. Trends. Ecol. Evol. 2011, 26, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Vander Wall, S.B. On the relative contributions of wind vs. animals to seed dispersal of four Sierra Nevada pines. Ecology 2008, 89, 1837–1849. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wang, L.; Baiketuerhan, Y.; Zhang, C.; Zhao, X.; von Gadow, K. Seed dispersal and seedling recruitment of trees at different successional stages in a temperate forest in northeastern China. J. Plant Ecol. 2013, 7, 337–346. [Google Scholar] [CrossRef]
- Madeira, B.G.; Espírito-Santo, M.M.; Neto, S.D.; Nunes, Y.R.F.; Sánchez-Azofeita, G.A.; Wilson, G.; Quesada, M. Changes in tree and liana communities along a successional gradient in a tropical dry forest in south-eastern Brazil. Plant. Ecol. 2009, 201, 291–304. [Google Scholar] [CrossRef]
- Almazán-Núñez, R.C.; Arizmendi, M.C.; Eguiarte, L.E.; Corcuera, P. Changes in composition, diversity and structture of woody plants in successional stages of tropical dry forest in southwest Mexico. Rev. Mex. Biodivers. 2012, 83, 1096–1109. [Google Scholar]
- Galindo-Jaimes, L.; González-Espinosa, M.; Quintana-Ascencio, P.; García-Barrios, L. Tree composition and structure in disturbed stands with varying dominance by Pinus spp. in the highlands of Chiapas, Mexico. Plant. Ecol. 2002, 162, 59–272. [Google Scholar] [CrossRef]
- Zavala, M.A.; Galindo-Jaimes, L.; González-Espinosa, M. Models of regional and local stand composition and dynamics of pine-oak forests in the central highlands of Chiapas (Mexico): Theoretical and management implications. In Biodiversity Loss and Conservation in Fragmented Forest Landscapes: The Forests of Montane Mexico and Temperate South America; Newton, A.C., Ed.; CAB International: Wallingford, UK, 2007; pp. 223–243. [Google Scholar]
- Bonfil, C.; Soberón, J. Quercus rugosa seedling dynamics in relation to its re-introduction in a disturbed Mexican landscape. Appl. Veg. Sci. 1999, 2, 189–200. [Google Scholar] [CrossRef]
- Kato, J.; Hayashi, I. The determination and prediction of pine to oak forest succession in Sugadaira, Central Japan. Korean. J. Ecol. 2003, 26, 155–163. [Google Scholar]
- García-Barrios, L.E.; González-Espinosa, M. Change in oak to pine dominance in secondary forests may reduce shifting agriculture yields: Experimental evidence from Chiapas, Mexico. Agr. Ecosyst. Environ. 2004, 102, 389–401. [Google Scholar] [CrossRef]
- Puhlick, J.J.; Laughlin, D.C.; Moore, M. Factors influencing ponderosa pine regeneration in the southwestern USA. For. Ecol. Manag. 2012, 264, 10–19. [Google Scholar] [CrossRef]
- Rodríguez-Trejo, D.A.; Fulé, P.Z. Fire ecology of Mexican pines and a fire management proposal. Int. J. Wildland Fire 2003, 12, 23–37. [Google Scholar] [CrossRef]
- Kobe, R.K. Light gradient partitioning among tropical tree species through differential seedling mortality and growth. Ecology 1999, 80, 187–201. [Google Scholar] [CrossRef]
- Lohbeck, M.; Poorter, L.; Lebrija-Trejos, E.; Martínez-Ramos, M.; Meave, J.A.; Paz, H.; Pérez-García, E.A.; Rómero-Pérez, I.E.; Tauro, A.; Bongers, F. Successional changes in functional composition contrast for dry and wet tropical forest. Ecology 2013, 94, 1211–1216. [Google Scholar] [CrossRef] [PubMed]
- Fulé, P.Z.; Convington, W.W. Changing fire regimes in Mexican pine forest. Ecological and management implications. J. Forest. 1996, 4, 33–38. [Google Scholar]
- Rodríguez-Trejo, D.A.; Catalina, H.; Ortega, V. Ecología del fuego en bosques de Pinus hartwegii. In Incendios Forestales en México. Métodos de Evaluación; Villers-Ruiz, L., López-Blanco, J., Eds.; Centro de Ciencias de la Atmósfera, UNAM: México City, Mexico, 2004; pp. 103–120. [Google Scholar]
- González-Tagle, M.A.; Schwendenmann, L.; Jiménez-Pérez, J.; Schulz, R. Forest structure and woody plant species composition along a fire chronosequence in mixed pine oak forest in the Sierra Madre Oriental, northeast Mexico. For. Ecol. Manag. 2008, 256, 161–167. [Google Scholar] [CrossRef]
- Lafon, C.W.; Waldron, J.D.; Cairns, D.M.; Tchakerian, D.M.; Coulson, R.N.; Klepzig, K.D. Modeling the effects of fire on the long-term dynamics and restoration of yellow pine and oak forest in the southern Appalachian mountains. Restor. Ecol. 2007, 15, 400–411. [Google Scholar] [CrossRef]
- Marañón-Jiménez, S.; Castro, J.; Querejeta, J.I.; Fernández-Ondoño, E.; Allen, C. Post-fire wood management alters stress, growth, and performance of pine regeneration in a Mediterranean ecosystem. For. Ecol. Manag. 2013, 308, 231–239. [Google Scholar] [CrossRef]
- Cervantes, G.V.; Arriaga, V.; Carabias, J. La problemática socioambiental e institucional de la reforestación en la región de La Montaña, Guerrero, México. Bol. Soc. Bot. Mex. 1996, 59, 67–80. [Google Scholar]
- Figueroa-Rangel, B.L.; Olvera-Vargas, M. Dinámica de la composición de especies en bosques de Quercus crassipes H. & B. en Cerro Grande, sierra de Manantlán, México. Agrociencia 2000, 34, 91–98. [Google Scholar]
- Fernández, I.; Carrasco, B.; Cabaneiro, A. Evolution of soil organic matter composition and edaphic carbon effluxes following oak forest clearing for pasture: Climate change implications. Eur. J. For. Res. 2011, 131, 1681–1693. [Google Scholar] [CrossRef]
- Egler, F.E. Vegetation science concepts I. Initial floristic composition—A factor in old-field vegetation development. Vegetation 1954, 4, 412–417. [Google Scholar] [CrossRef]
- Norden, N.; Chazdon, R.L.; Chao, A.; Jiang, Y.H.; Vilchez-Alvarado, B. Resilience of tropical rain forests: Tree community reassembly in secondary forests. Ecol. Lett. 2009, 12, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Luna, I.; Llorente, J. Historia Natural Del Parque Ecológico Estatal Omiltemi, Chilpancingo, Guerrero, México; CONABIO-UNAM: México City, Mexico, 1993. [Google Scholar]
- García, E. Modificaciones al Sistema de Clasificación Climática de Köppen; Universidad Autónoma de Chapingo: México City, Mexico, 2004. [Google Scholar]
- Muller-Dombois, D.; Ellenberg, H. Aims and Methods of Vegetation Ecology; John Wiley and Sons: New York, NY, USA, 1974. [Google Scholar]
- Montaña, C.; Ezcurra, E. Simple instrument for quick measurement of crown projections. J. Forest. 1980, 78, 699. [Google Scholar]
- Stevens, P.F. Angiosperm Phylogeny Website. Version 9. 2001. Available online: http://www.mobot.org/MOBOT/research/APweb/ (accessed on 7 October 2014).
- W3Trópicos. Missouri Botanical Garden’s VAST Nomenclatural Database and Associated Authority Files. 2014. Available online: http://www.tropicos.org (accessed on 10 November 2014).
- Hortal, J.; Borges, P.A.; Gaspar, C. Evaluating the performance of species richness estimators: Sensitivity to sample grain size. J. Anim. Ecol. 2006, 75, 274–287. [Google Scholar] [CrossRef] [PubMed]
- González-Oreja, J.A.; de la Fuente-Díaz, A.A.; Hernández-Santín, L.; Buzo-Franco, D.; Bonache-Regidor, C. Evaluación de estimadores no paramétricos de la riqueza de especies. Un ejemplo con aves en áreas verdes de la ciudad de Puebla, México. Anim. Biodivers. Conserv. 2010, 33, 31–45. [Google Scholar]
- Colwell, R.K. EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples; Version 8; Robert K. Colwell: Storrs, CT, USA, 2006; Available online: http://viceroy.eeb.uconn.edu/estimates (accessed on 20 June 2014).
- Brower, J.E.; Zar, J.H.; von Ende, C.N. Field and Laboratory Methods for General Ecology, 4th ed.; WCB-McGraw-Hill: New York, NY, USA, 1998. [Google Scholar]
- Kovach, W.L. MVSP: A Multivariate Statistical Package for Windows. Version 3.13r; Kovach Computing Services: Pentraeth, Wales, UK, 2009. [Google Scholar]
- Cheng, C. Statistical approaches on discriminating spatial variation of species diversity. Bot. Bull. Acad. Sin. 2004, 45, 339–346. [Google Scholar]
- Kruskal, J.B. Multidimensional scaling by optimizing goodness of fit to a nonmetric hypothesis. Psychometrika 1964, 29, 1–27. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-parametric multivariate analysis of changes in community structure. Aus. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Ramírez-Marcial, N.; González-Espinosa, M.; García-Moya, E. Establecimiento de Pinus spp. y Quercus spp. en matorrales y pastizales de Los Altos de Chiapas. Agrociencia 1996, 30, 249–257. [Google Scholar]
- Ochoa-Gaona, S.; González-Espinosa, M.; Meave, J.A.; Sorani, V. Effect of forest fragmentation on the woody flora of the highlands of Chiapas, Mexico. Biodivers. Conserv. 2004, 13, 867–884. [Google Scholar] [CrossRef]
- Castro, J.; Gómez, J.M.; García, D.; Zamora, R.; Hódar, J.A. Seed predation and dispersal in relict scots pine forests in southern Spain. Plant. Ecol. 1999, 145, 115–123. [Google Scholar] [CrossRef]
- Kreutz, A.; Aakala, T.; Grenfell, R.; Kuuluvainen, T. Spatial tree community structure in three stands across a forest succession gradient in northern boreal Fennoscandia. Silva. Fenn. 2015, 49, 1–17. [Google Scholar] [CrossRef]
- Martins, A.M.; Engel, V.L. Soil seed banks in tropical forests fragments with different disturbance histories in southeastern Brazil. Ecol. Eng. 2007, 31, 265–291. [Google Scholar] [CrossRef]
- Peña-Claros, M.; De Boo, H. The effect of forest successional stage on seed removal of tropical rain forest tree species. J. Trop. Ecol. 2002, 18, 261–274. [Google Scholar] [CrossRef]
- Olano, J.M.; Caballero, I.; Laskurain, N.A.; Loidi, J.; Escudero, A. Seed bank spatial pattern in a temperate secondary forest. J. Veg. Sci. 2002, 13, 775–784. [Google Scholar] [CrossRef]
- Onaindia, M.; Dominguez, I.; Albizu, I.; Garbisu, C.; Amezaga, I. Vegetation diversity and vertical structure ad indicators of forest disturbance. For. Ecol. Manag. 2004, 195, 341–354. [Google Scholar] [CrossRef]
- Bongers, F.; Poorter, L.; Hawthorne, W.D.; Sheil, D. The intermediate disturbance hypothesis applies to tropical forests, but disturbance contributes little to tree diversity. Ecol. Lett. 2009, 12, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ayerde-Lozada, D.A.; López-Mata, L. Estructura poblacional y parámetros demográficos de Juniperus flaccida Schltdl. Madera y Bosques 2006, 12, 65–76. [Google Scholar]
- Connell, J.H.; Slatyer, R.O. Mechanisms of succession in natural communities and their role in community stability and organization. Am. Nat. 1977, 111, 1119–1144. [Google Scholar] [CrossRef]
- Martínez-Orea, Y.; Orozco-Segovia, A.; Castillo-Argüero, A.; Collazo-Ortega, M.; Zavala-Hurtado, J.A. Seed rain as a source of propagules for natural regeneration in a temperate forest in Mexico City. J. Torrey Bot. Soc. 2014, 141, 135–150. [Google Scholar] [CrossRef]
- Marañón, T.; Zamora, R.; Villar, R.; Zavala, M.A.; Quero, J.L.; Pérez-Ramos, I.; Mendoza, I.; Castro, J. Regeneration of tree species and restoration under contrasted Mediterranean habitats: Fields and glasshouse experiments. Int. J. Ecol. Environ. Sci. 2004, 30, 187–196. [Google Scholar]
- Eilmann, B.; Zweifel, R.; Buchmann, N.; Graf, E.; Rigling, A. Drought alters timing, quantity, and quality of wood formation in Scots pine. J. Exp. Bot. 2011, 62, 2763–2771. [Google Scholar] [CrossRef] [PubMed]
- Sosa-Pérez, G.; Rodríguez-Trejo, D.A. Efecto de la calidad de planta en la supervivencia y crecimiento de Pinus patula en un área quemada. Rev. Chapingo Ser. Cienc. Forest Ambient. 2003, 9, 35–43. [Google Scholar]
- Rzedowski, J. Vegetación de México; Limusa: México City, Mexico, 1978. [Google Scholar]
- Quintana-Ascencio, P.F.; Ramírez-Marcial, N.; González-Espinosa, M.; Martínez-Icó, M. Sapling survival and growth of conifer and broad-leaved trees in successional highland habitats in Mexico. Appl. Veg. Sci. 2004, 7, 81–88. [Google Scholar] [CrossRef]
- Veblen, T.T. Forest preservation in Highland Guatemala. Geogr. Rev. 1978, 68, 417–434. [Google Scholar] [CrossRef]
- Tansley, A.G. The use and abuse of vegetational concepts and terms. Ecology 1935, 16, 284–307. [Google Scholar] [CrossRef]






| Successional Stage | Shrub Species Number | Tree Species Number | Observed Richness | Expected Richness (Chao2) | Chao2 % | Abundance of Shrubs | Abundance of Trees |
|---|---|---|---|---|---|---|---|
| Early | 8 | 14 | 22 | 25 | 88.0 | 102 | 117 |
| Intermediate | 10 | 17 | 27 | 31 | 87.1 | 97 | 226 |
| Mature | 9 | 26 | 35 | 42 | 83.3 | 65 | 428 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almazán-Núñez, R.C.; Corcuera, P.; Parra-Juárez, L.; Jiménez-Hernández, J.; Charre, G.M. Changes in Structure and Diversity of Woody Plants in a Secondary Mixed Pine-Oak Forest in the Sierra Madre del Sur of Mexico. Forests 2016, 7, 90. https://doi.org/10.3390/f7040090
Almazán-Núñez RC, Corcuera P, Parra-Juárez L, Jiménez-Hernández J, Charre GM. Changes in Structure and Diversity of Woody Plants in a Secondary Mixed Pine-Oak Forest in the Sierra Madre del Sur of Mexico. Forests. 2016; 7(4):90. https://doi.org/10.3390/f7040090
Chicago/Turabian StyleAlmazán-Núñez, Roberto Carlos, Pablo Corcuera, Liliana Parra-Juárez, Javier Jiménez-Hernández, and Grégory Michäel Charre. 2016. "Changes in Structure and Diversity of Woody Plants in a Secondary Mixed Pine-Oak Forest in the Sierra Madre del Sur of Mexico" Forests 7, no. 4: 90. https://doi.org/10.3390/f7040090
APA StyleAlmazán-Núñez, R. C., Corcuera, P., Parra-Juárez, L., Jiménez-Hernández, J., & Charre, G. M. (2016). Changes in Structure and Diversity of Woody Plants in a Secondary Mixed Pine-Oak Forest in the Sierra Madre del Sur of Mexico. Forests, 7(4), 90. https://doi.org/10.3390/f7040090