
Recovery of Vegetation Cover and Soil after the Removal of Sheep in Socorro Island, Mexico

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Normalized Difference Vegetation Index (NDVI)
2.3. Field Assessment of Vegetation
2.4. Soil Compaction
2.5. Physical-Chemical Soil Parameters
3. Results
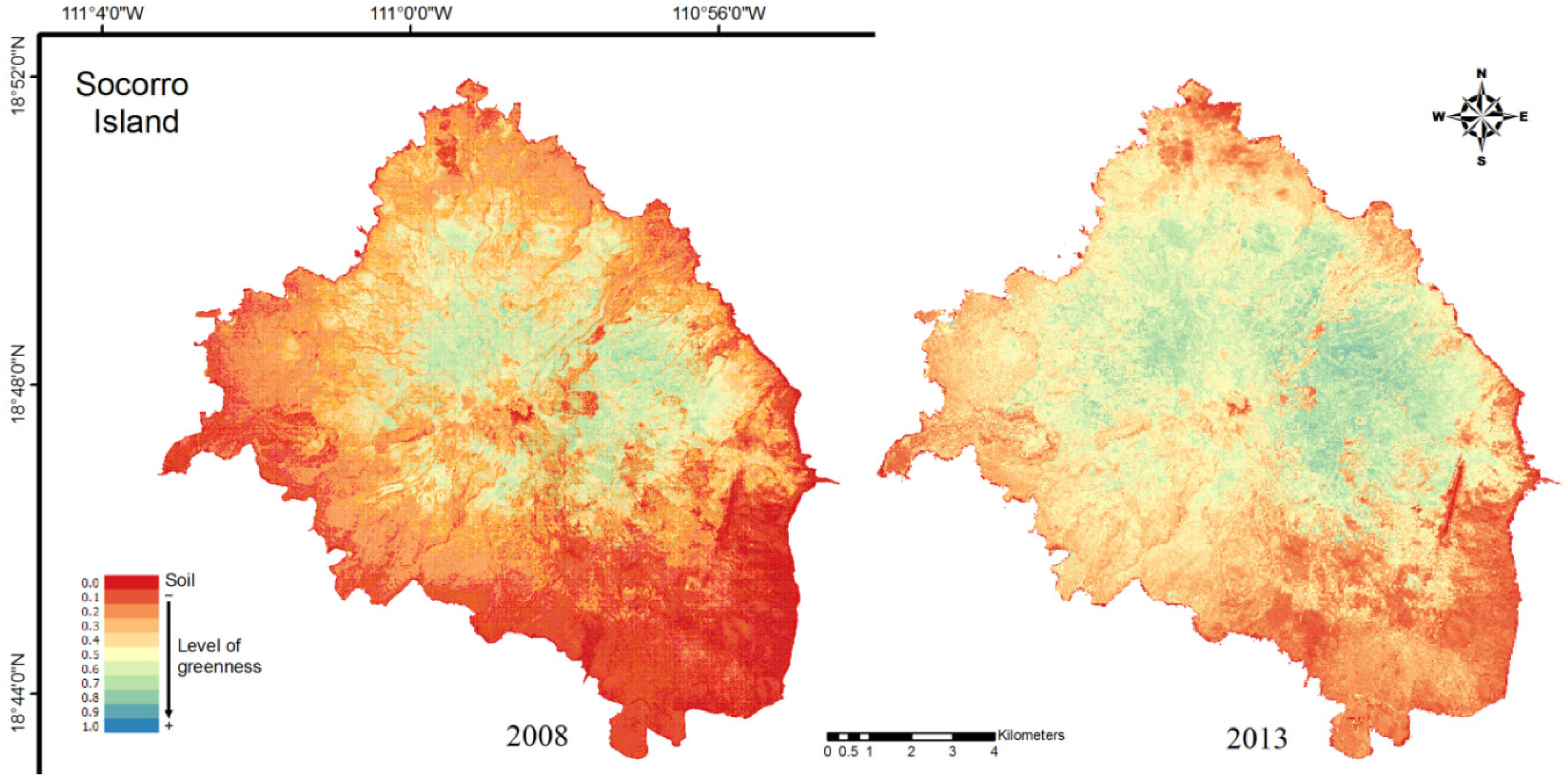
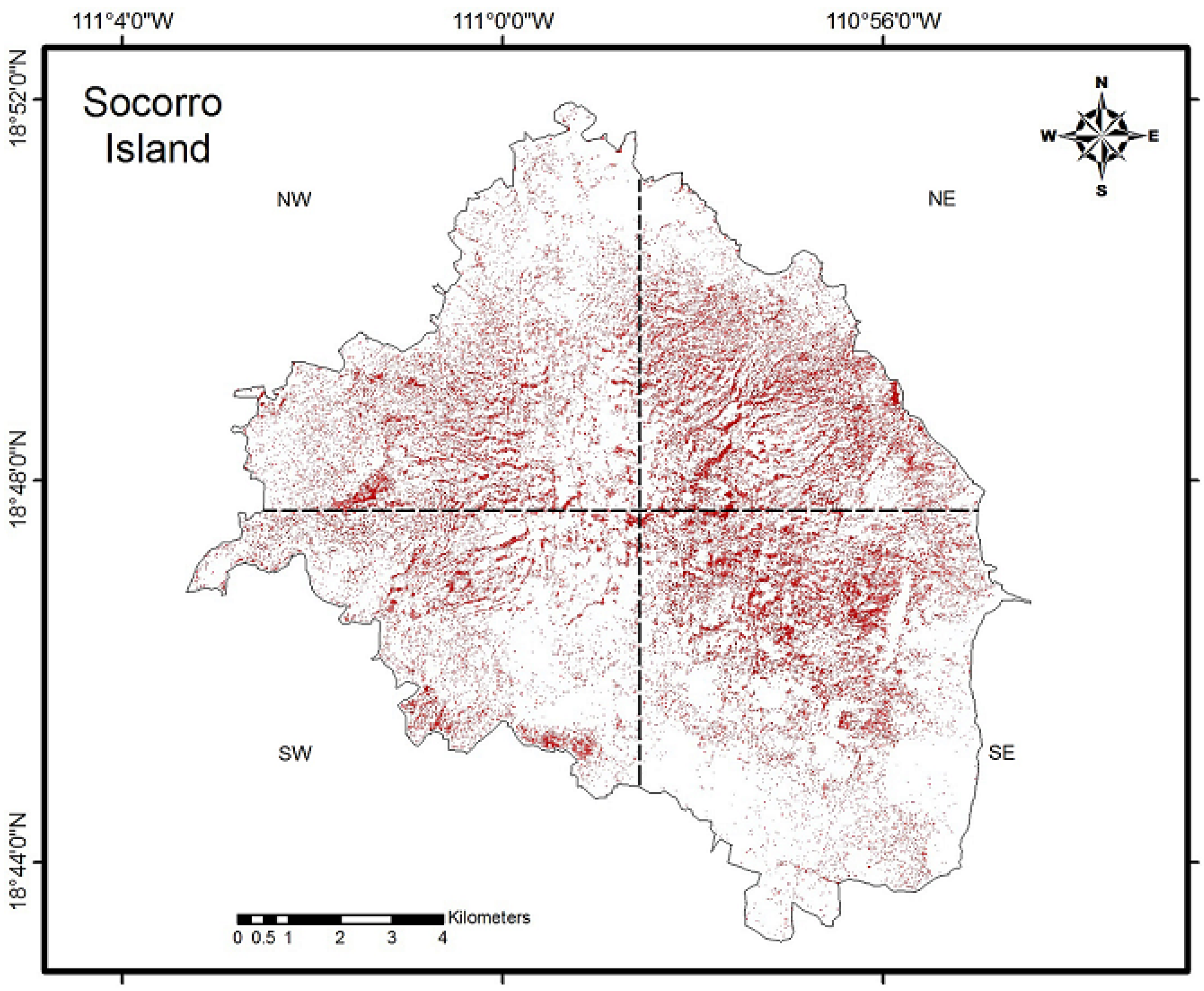
3.1. NDVI
3.2. Field Assessment of Vegetation
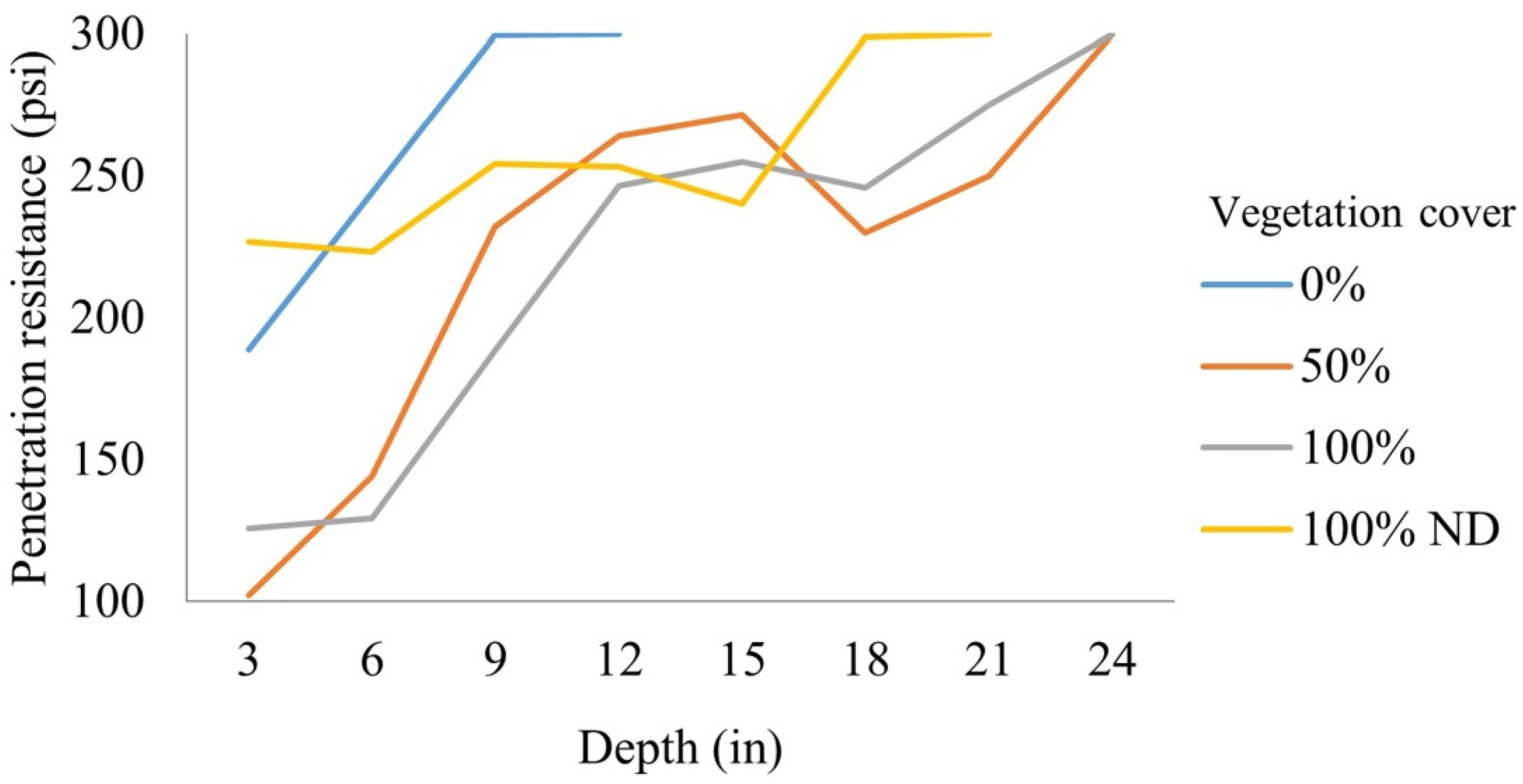
3.3. Soil Compaction
3.4. Physical-Chemical Soil Parameters
4. Discussion
4.1. NDVI and Field Assessment of Vegetation
4.2. Soil Recovery
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Mason, H.L. The edaphic factor in narrow endemism. II. The Geographic occurrence of plants of highly restricted patterns of distribution. Madroño 1946, 8, 241–257. [Google Scholar]
- Tilman, D.; May, R.M.; Lehman, C.L.; Nowak, M.A. Habitat destruction and the extinction debt. Nature 1994, 371, 65–66. [Google Scholar] [CrossRef]
- Coblentz, B.E. The effects of feral goat (Capra hircus) on islands ecosystems. Biol. Cons. 1978, 13, 279–286. [Google Scholar] [CrossRef]
- Eberhard, T. Introduced birds and mammals and their ecological effects. Swed. Wildl. Res. 1988, 13, 1–107. [Google Scholar]
- Rosenberg, D.K. The impact of introduced herbivores on the Galapagos rail (Laterallus spilonotus). Monogr. Syst. Bot. Missour.i Bot. Gard. 1990, 32, 169–178. [Google Scholar]
- Mooney, H.A.; Hobbs, R.J. Invasive Species in a Changing World; Island Press: Washington, DC, USA, 2000. [Google Scholar]
- Donlan, C.J.; Tershy, B.R.; Croll, D.A. Islands and introduced herbivores: Conservation action as ecosystem experimentation. J. Appl. Ecol. 2002, 39, 235–246. [Google Scholar] [CrossRef]
- Primack, R.B. Essentials of Conservation Biology; Sinauer Associates Press: Sunderland, MA, USA, 2002. [Google Scholar]
- Veitch, C.R.; Clout, M.N. Turning the Tide: The Eradication of Invasive Species; IUCN, SSC Invasive Species Specialist Group: Gland, Switzerland, 2002; pp. 374–380. [Google Scholar]
- Courchamp, F.; Chapuis, J.L.; Pascal, M. Mammal invaders on islands: Impact, control, and control impact. Biol. Rev. 2003, 78, 347–383. [Google Scholar] [CrossRef] [PubMed]
- Nogales, M.; Martín, A.; Tershy, B.R.; Donlan, C.J.; Veitch, D.; Puerta, N.; Wood, B.; Alonso, J. A Review of feral cat eradication on islands. Cons. Biol. 2004, 18, 310–319. [Google Scholar] [CrossRef]
- Campbell, K.; Donlan, C.J. Feral goat eradication on islands. Cons. Biol. 2005, 19, 1362–1374. [Google Scholar] [CrossRef]
- Pimentel, D.; Zúñiga, R.; Morrison, D. Update on the environmental and economic costs associated with alien invasive species in the United States. Ecol. Econ. 2005, 52, 273–288. [Google Scholar] [CrossRef]
- Aguirre-Muñoz, A.; Samaniego-Herrera, A.; Luna-Mendoza, L.; Ortiz-Alcaraz, A.; Rodríguez-Malagón, M.; Méndez-Sánchez, F.; Félix-Lizárraga, M.; Hernández-Montoya, J.; González-Gómez, R.; Torres-García, F.; et al. Island restoration in Mexico: Ecological outcomes after systematic eradications of invasive mammals. In Island Invasives: Eradication and Management; Veitch, C.R., Clout, M.N., Towns, D.R., Eds.; IUCN: Gland, Switzerland, 2011; pp. 250–258. [Google Scholar]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography. Vol 1; Princeton University Press: Princeton, NJ, USA, 1967. [Google Scholar]
- Álvarez-Castañeda, S.T.; Ortega-Rubio, A. Current status of rodents on islands of the Gulf of California. Biol. Cons. 2003, 109, 157–163. [Google Scholar] [CrossRef]
- Blumstein, D.T.; Daniel, J.C. The loss of anti-predator behavior following isolation on islands. Proc. R. Soc. B Biol. Sci. 2005, 272, 1663–1668. [Google Scholar] [CrossRef] [PubMed]
- Cortés-Calva, P.; Gallo-Reynoso, J.P.; Delgadillo-Rodríguez, J.; Lorenzo, C.; Álvarez-Castañeda, S.T. The effect of feral dogs and other alien species on native mammals of Isla de Cedros, Mexico. Nat. Areas J. 2013, 33, 466–473. [Google Scholar] [CrossRef]
- Cincotta, P.R.; Wisnewski, J.; Engelman, R. Human population in the biodiversity hotspot. Nature 2000, 404, 990–992. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, R.J.; Fernandez-Palacios, J.M. Island Biogeography; Oxford University Press: Oxford, UK, 2007. [Google Scholar]
- Donlan, C.J.; Croll, D.A.; Tershy, B.R. Islands, exotic herbivores, and invasive plants: Their roles in Coastal California restauration. Res. Ecol. 2003, 11, 524–530. [Google Scholar] [CrossRef]
- Álvarez-Cárdenas, S.; Castellanos, A.; Galina, P.; Ortega-Rubio, A.; Arnaud, G. Aspectos de la población y el hábitat del borrego doméstico (Ovis aries). In La Isla Socorro, Reserva de la Biosfera Archipiélago de Revillagigedo, México; Ortega Rubio, A., Castellanos, A., Eds.; Centro de Investigaciones Biológicas del Noroeste S.C.: La Paz, Mexico, 1994; pp. 301–317. [Google Scholar]
- Villa, B. La Isla Socorro. Vertebrados terrestres. Monogr. Inst. Geofís. 1960, 2, 203–2016. [Google Scholar]
- Walter, H.S.; Levin, G.A. Feral sheep on Socorro Island: Facilitators of alien plant colonization and ecosystem decay. Divers. Distrib. 2008, 14, 422–431. [Google Scholar] [CrossRef]
- Richards, A.F.; Brattstrom, B.H. Bibliography, cartography, discovery and exploration of the islas Revillagigedo. Proc. Calif. Acad. Sci. 1959, 29, 315–360. [Google Scholar]
- Veitch, C.R. The Eradication of Cats and Sheep from Socorro Island: A Report to the Socorro Island Project; Northern Region Technical Report Series No. 11; Department of Conservation: Auckland, New Zealand, 1989. [Google Scholar]
- Castellanos, A.; Arnaud, G.; Álvarez, S.; Rodríguez, R.; Galina, P. El borrego doméstico (Ovis aries) en isla Socorro, su situación actual e impacto en el suelo, la vegetación y la fauna nativa. In Proceedings of the XI Congreso Nacional de Zoología, Mérida, Mexico, 28–31 October 1991.
- Ortega, A.; Castellanos, A.; Arnaud, G.; Maya, Y.; Rodríguez, R.; León, J.L.; Cancino, J.; Jiménez, C.; Llinas, J.; Álvarez, S.; et al. Estudio de los Recursos Naturales de la Isla Socorro, Revillagigedo. Ciencia 1992, 43, 175–184. [Google Scholar]
- Castellanos, A.; Rodríguez-Estrella, R. Current status of the Socorro Mockingbird. Wilson Bull. 1993, 105, 167–171. [Google Scholar]
- Galina, P.; Álvarez, S.; Ortega Rubio, A. Aspectos ecológicos de la herpetofauna. In La Isla Socorro, Reserva de la Biosfera Archipiélago de Revillagigedo, México; Ortega Rubio, A., Castellanos, A., Eds.; Centro de Investigaciones Biológicas del Noroeste S.C.: La Paz, Mexico, 1994; pp. 185–197. [Google Scholar]
- Rodríguez-Estrella, R.; Rivera-Rodríguez, L.; Mata, E. Avifauna Terrestre. In La Isla Socorro, Reserva de la Biosfera Archipiélago de Revillagigedo, México; Ortega Rubio, A., Castellanos, A., Eds.; Centro de Investigaciones Biológicas del Noroeste S.C.: La Paz, Mexico, 1994; pp. 199–224. [Google Scholar]
- Maya-Delgado, Y.; Salinas-Zavala, F.; Troyo-Diéguez, E. Estado actual del suelo y propuestas para su conservación. In La Isla Socorro, Reserva de la Biosfera Archipiélago de Revillagigedo, México; Ortega Rubio, A., Castellanos, A., Eds.; Centro de Investigaciones Biológicas del Noroeste S.C.: La Paz, Mexico, 1994; pp. 63–75. [Google Scholar]
- Adema, E.O.; Babinec, F.J.; Peinemann, N. Pérdida de nutrientes por erosión hídrica en dos suelos del cardenal pampeano. Cienc. Suelo 2001, 19, 144–154. [Google Scholar]
- Peterson, G.; Allen, C.R.; Holling, C.S. Ecological resilience, biodiversity, and scale. Ecosystems 1998, 1, 6–18. [Google Scholar] [CrossRef]
- Zavaleta, E.S.; Hobbs, R.J.; Mooney, H.A. Viewing invasive species removal in a whole-ecosystem context. Trends Ecol. Evol. 2001, 16, 454–459. [Google Scholar] [CrossRef]
- Holl, K.D.; Aide, T.M. When and where to actively restore ecosystems? Forest Ecol. Manag. 2011, 261, 1558–1563. [Google Scholar] [CrossRef]
- Jones, H.P.; Schmitz, O.J. Rapid Recovery of Damaged Ecosystems. PLoS ONE 2009, 4, 53–56. [Google Scholar] [CrossRef] [PubMed]
- CONANP-SEMARNAT. Programa de Conservación y Manejo, Reserva de la Biósfera Archipiélago de Revillagigedo; Comisión Nacional de Áreas Naturales Protegidas: Mexico, D.F., Mexico, 2004. [Google Scholar]
- León de la Luz, J.L.; Breceda, A.; Coria, R.; Cancino, J. Asociaciones vegetales. In La Isla Socorro, Reserva de la Biosfera Archipiélago de Revillagigedo, México; Ortega Rubio, A., Castellanos, A., Eds.; Centro de Investigaciones Biológicas del Noroeste S.C.: La Paz, Mexico, 1994; pp. 115–141. [Google Scholar]
- Campa-Molina, M.A. Flora y potencial apícola de Isla Socorro, Archipiélago de las Revillagigedo. Master’s Thesis, Universidad de Guadalajara, Jalisco, Mexico, 1989. [Google Scholar]
- Levin, G.A.; Moran, R. The Vascular Flora of Isla Socorro, Mexico; San Diego Society of Natural History: San Diego, CA, USA, 1989. [Google Scholar]
- Hall, E.R. The Mammals of North America; Wiley Inter-Sciences Publications: New York, NY, USA, 1981; p. 191. [Google Scholar]
- Ortiz-Alcaraz, A.; Barredo-Barberena, J.M.; Aguirre-Muñoz, A.; Santos del Prado-Gasca, K. Acciones para la Recuperación del Ambiente en Isla Socorro, Archipiélago de Revillagigedo. Reporte Final; Grupo de Ecología y Conservación de Islas, A.C.-Instituto Nacional de Ecología: Mexico City, Mexico, 2011; p. 32. [Google Scholar]
- Valdez-Lazalde, J.; González-Guillen, M.; de los Santos-Posadas, H. Estimación de cobertura arbórea mediante imágenes satelitales multiespectrales de alta resolución. Agrociencia 2006, 40, 383–394. [Google Scholar]
- Cabral, F. Dinámica espaciotemporal de uso del suelo en la UMAFOR 12 del Estado de México mediante el uso de sistemas de información geográfica (1990–2007). Master’s Thesis, Universidad Autónoma Chapingo, Mexico, 2011. [Google Scholar]
- Mostacedo, B.; Fredericksen, T.S. Manual de Métodos Básicos de Muestreo y Análisis en Ecología Vegetal; Proyecto de Manejo Forestal Sostenible (BOLFOR): Santa Cruz, Bolivia, 2000. [Google Scholar]
- Ortiz-Alcaraz, A.; Aguirre-Muñoz, A.; Méndez-Sánchez, F.; Ortega-Rubio, A. Feral sheep eradication at Socorro Island, México: A mandatory step to ensure ecological restoration. Interciencia 2016, 41, 184–189. [Google Scholar]
- Jackson, M.L. Análisis químico de suelos, 4th ed.; Ediciones Omega S.A.: Barcelona, Spain, 1982; p. 652. [Google Scholar]
- Bates, R.G. Determination of pH. Theory and Practice; John Wiley and Sons: New York, NY, USA, 1964; pp. 11–12. [Google Scholar]
- Page, A.L.; Miller, R.H.; Keeney, D.R. Methods of Soil Analysis. Part 2, Chemical and Microbiological Properties, 2nd ed.; American Society of Agronomy, Inc. and Soil Science Society of America, Inc: Wisconsin, WI, USA, 1982; p. 1159. [Google Scholar]
- Pinzón, A.; Amézquita, E. Compactación de suelos por pisoteo de animales en pastoreo en el piedemonte amazónico de Colombia. Pasturas tropicales 1979, 13, 21–26. [Google Scholar]
- Riney, T. Study and management of large mammals; John Wiley & Sons Ltd.: Chichester, UK, 1982. [Google Scholar]
- González, M.; Nogues, E. Pisoteo animal y su efecto en la densidad aparente del suelo en un Haplustol franco arenoso bajo diferentes manejos. Rev. Divulg. Técnic. Agríc. Agroind. 2012, 31, 1–9. [Google Scholar]
- Bakker, E.S.; Ritchie, M.E.; Olff, H.; Milchunas, D.G.; Knops, J.M.H. Herbivore impact on grassland plant diversity depends on habitat productivity and herbivore size. Ecol. Lett. 2006, 9, 780–788. [Google Scholar] [CrossRef] [PubMed]
- Milchunas, D.G.; Sala, O.E.; Lauenroth, W.K. A generalized model of the effects of grazing by large herbivores on grassland community structure. Am. Nat. 1988, 132, 87–106. [Google Scholar] [CrossRef]
- Vibrans, H. Taller de identificación de Malezas. In Manejo de Malezas en México. Vol. 1 Maleza terrestre; Bojórquez-Bojórquez, G., Rosales-Robles, E., Zeta-Padilla, G., Eds.; Universidad Autónoma de Sinaloa: Culiacan, Mexico, 2011; pp. 293–308. [Google Scholar]
- Castellano, M.J.; Valone, T.J. Livestock, soil compaction and water infiltration rate: Evaluating a potential desertification recovery mechanism. J. Arid Environ. 2007, 71, 97–108. [Google Scholar] [CrossRef]
- Allington, G.R.H.; Valone, T.J. Reversal of desertification: The role of physical and chemical soil properties. J. Arid Environ. 2010, 74, 973–977. [Google Scholar] [CrossRef]
- Amézquita, E.; Chávez, L.F. La compactación del suelo y sus efectos en la productividad de los suelos. In Proceedings of the Congreso Costarricense de la Ciencia del Suelo, San José, Costa Rica, 9 April 1999.
- Sellés, G.; Fereyra, R.; Ruíz, R.; Ferreyra, R.; Ahumada, R. Compactación de Suelos y su Control: Estudio de Casos en el Valle de Aconcagua, Boletín INIA 234; Instituto de Investigaciones Agropecuarias: Santiago, Chile, 2012. [Google Scholar]
- USDA. Soil Survey Laboratory Methods Manual. Soil Survey Investigations Report No. 42. Version 3.0; Natural Resources Conservation Service: Washington, DC, USA, 1996. [Google Scholar]
- Hartmann, A.; Aßmus, B.; Kirchhof, G.; Schloter, M. Direct approaches for studying soil microbes. In Modern Soil Microbiology; van Elsas, J.D., Trevors, J.T., Wellington, E.M.H., Eds.; Marcel Dekker Inc.: New York, NY, USA, 1997; p. 712. [Google Scholar]
- Mediavilla, M.J. Meteorización química. Enseñanza de las Ciencias de la Tierra; Universidad de Girona: Girona, Spain, 2014; pp. 230–238. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Portions of the Island | 2008 | 2013 | Difference |
|---|---|---|---|
| Northwest (NW) | 0.443 | 0.501 | 0.058 |
| Northeast (NE) | 0.461 | 0.612 | 0.150 |
| Southwest (SW) | 0.394 | 0.499 | 0.105 |
| Southeast (SE) | 0.346 | 0.533 | 0.187 |
| Type of Habitat | No. of Plant Species | p | Vegetation Cover (%) | p | ||
|---|---|---|---|---|---|---|
| 2009 | 2013 | 2009 | 2013 | |||
| Forest | 8.86 (± 4.41) | 15.00 (± 2.31) | * | 29.29 (± 8.38) | 85.00 (± 9.24) | ** |
| Mixed scrub | 6.67 (± 2.73) | 13.67 (± 4.23) | * | 19.17 (± 2.99) | 72.50 (± 11.54) | ** |
| Eroded surface | 4.71 (3.45) | 8.29 (± 2.56) | ** | 14.29(± 4.27) | 50.29 (± 16.13) | ** |
| Species | Life Form | Zones within the Sampling Area | |||||
|---|---|---|---|---|---|---|---|
| Northeast | Southeast | ||||||
| ind ha−1 | Increase(%) | ind ha−1 | Increase(%) | ||||
| 2009 | 2013 | 2009 | 2013 | ||||
| Hyptis pectinata (L.) Poit. (x) | Shrub | 60 | 210 | 350 | 330 | 4860 | 1473 |
| Dodonaea viscosa Jacq. (n) | Shrub | 910 | 6190 | 680 | 140 | 1370 | 979 |
| Pteridium caudatum (L.) Maxon (n)* | Herbs | 6% | 58% | 967 | 11% | 43% | 391 |
| Nicotiana stocktonii Brandegee (e) | Herbs | 30 | 190 | 633 | 40 | 260 | 650 |
| Erigeron socorrensis Brandegee (e) | Herbs | 240 | 1420 | 592 | 80 | 350 | 437 |
| Mitracarpus hirtus (L.) DC. (x) | Herbs | 80 | 440 | 550 | 20 | 90 | 450 |
| Aristida spp. (x)^* | Herbs | 1% | 3% | 300 | 6% | 31% | 517 |
| Amaranthus palmeri S.Watson (n) | Sub-shrub | 220 | 900 | 409 | 100 | 180 | 180 |
| Psidium socorrense I.M.Johnst. (e) | Tree | 50 | 190 | 380 | 30 | 100 | 333 |
| Chamaesyce spp. (x)^ | Herbs | 70 | 160 | 229 | 100 | 370 | 370 |
| Cenchrus spp. (x)^ | Herbs | 40 | 110 | 275 | 160 | 530 | 331 |
| Guettarda insularis Brandegee (e) | Tree | 80 | 220 | 275 | 170 | 320 | 188 |
| Perityle socorrosensis Rose (e) | Herbs | 390 | 780 | 200 | 310 | 620 | 200 |
| Vegetation Cover | ||||
|---|---|---|---|---|
| 0% | 50% | 100% | 100% ND | |
| pH | 7.20 (0.16)a | 6.99 (0.28)b | 7.03 (0.16)a,b | 7.11 (0.22)a,b |
| Electrical conductivity (dS m−1) | 0.09 (0.03)a | 0.07 (0.03)a | 0.07 (0.02)b | 0.09 (0.03)a |
| Total nitrogen (%) | 0.20 (0.07)a | 0.45 (0.11)b | 0.48 (0.14)b | 0.51 (0.17)b |
| Organic carbon (%) | 1.53 (0.52)a | 3.60 (0.86)b | 3.22 (0.78)b | 3.27 (1.08)b |
| Phosphorus (meq 100g−1) | 0.01 (0.01)a | 0.03 (0.02)b | 0.03 (0.02)b | 0.03 (0.02)b |
| Calcium (meq 100g−1) | 0.03 (0.01)a | 0.08 (0.04)b | 0.05 (0.02)b | 0.09 (0.03)b |
| Magnesium (meq 100g−1) | 0.10 (0.04)a | 0.10 (0.03)a | 0.11 (0.03)a | 0.20 (0.10)b |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ortíz-Alcaraz, A.; Maya-Delgado, Y.; Cortés-Calva, P.; Aguirre-Muñoz, A.; Rojas-Mayoral, E.; Cordoba-Matson, M.V.; Ortega-Rubio, A. Recovery of Vegetation Cover and Soil after the Removal of Sheep in Socorro Island, Mexico. Forests 2016, 7, 91. https://doi.org/10.3390/f7040091
Ortíz-Alcaraz A, Maya-Delgado Y, Cortés-Calva P, Aguirre-Muñoz A, Rojas-Mayoral E, Cordoba-Matson MV, Ortega-Rubio A. Recovery of Vegetation Cover and Soil after the Removal of Sheep in Socorro Island, Mexico. Forests. 2016; 7(4):91. https://doi.org/10.3390/f7040091
Chicago/Turabian StyleOrtíz-Alcaraz, Antonio, Yolanda Maya-Delgado, Patricia Cortés-Calva, Alfonso Aguirre-Muñoz, Evaristo Rojas-Mayoral, Miguel V. Cordoba-Matson, and Alfredo Ortega-Rubio. 2016. "Recovery of Vegetation Cover and Soil after the Removal of Sheep in Socorro Island, Mexico" Forests 7, no. 4: 91. https://doi.org/10.3390/f7040091