
Cellulose Dynamics during Foliar Litter Decomposition in an Alpine Forest Meta-Ecosystem

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Experimental Design and Litterbag Incubation
2.3. Local-Scale Environmental Factors
2.4. Measurements of Litter Initial Traits and Cellulose Degradation
2.5. Statistical Analyses
3. Results
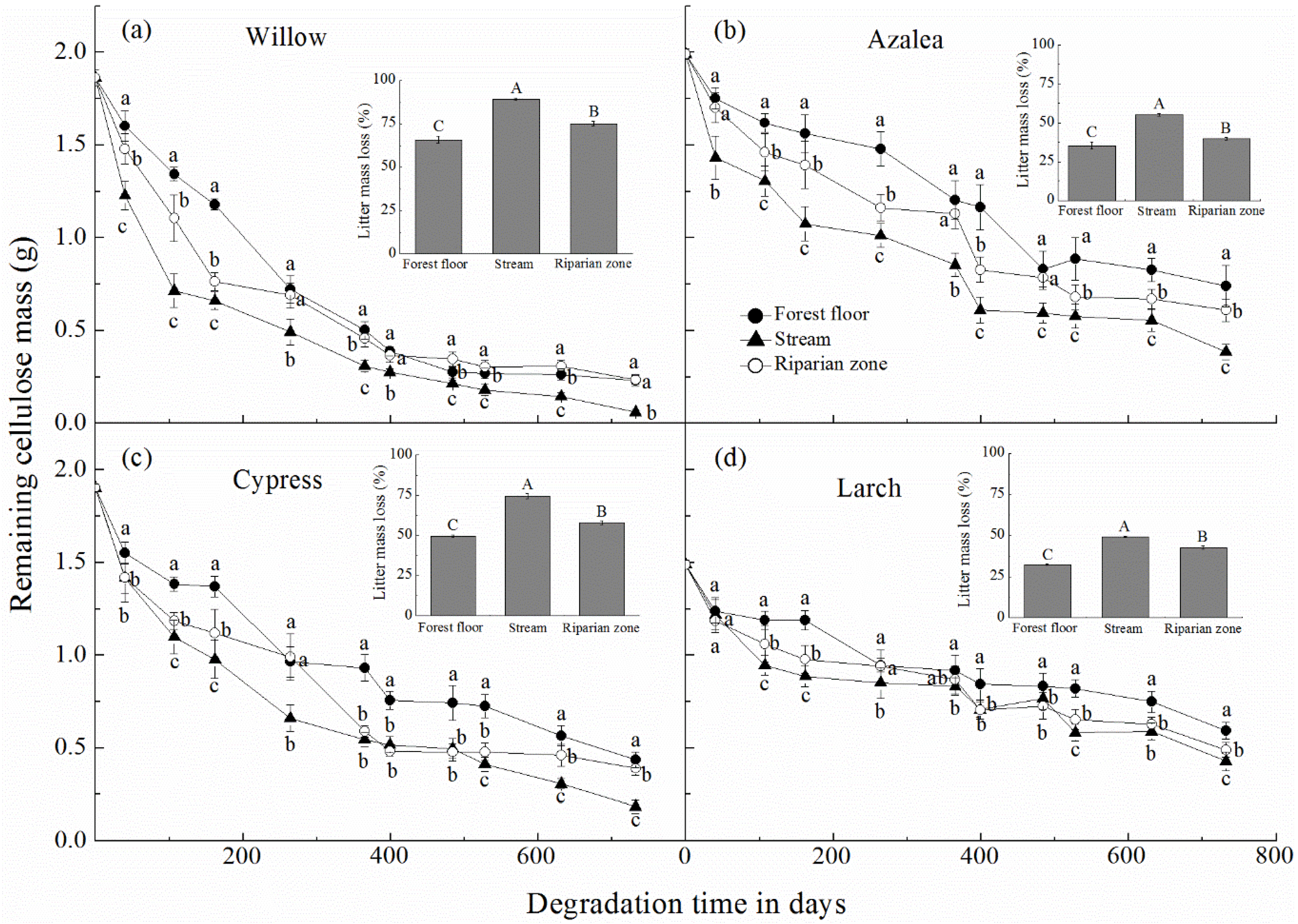
3.1. Litter Mass Loss and Cellulose Mass Remaining
3.2. Cellulose Concentration
3.3. Cellulose Degradation Rate
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- ACIA. Arctic Climate Impact Assessment-Scientific Report; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Berg, B.; McClaugherty, C. Plant Litter: Decomposition, Humus Formation, Carbon Sequestration, 3rd ed.; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Prescott, C. Decomposition and mineralization of nutrients from litter and humus. In Nutrient Acquisition by Plants: An Ecological Perspective; BassirRad, H., Ed.; Springer-Verlag: Berlin, Germany, 2005; pp. 125–141. [Google Scholar]
- Berg, B.; Laskowski, R. Litter decomposition: A guide to carbon and nutrient turnover. In Advances in Ecological Research; Elsevier: Amsterdam, The Netherlands, 2006. [Google Scholar]
- Fioretto, A.; Di Nardo, C.; Papa, S.; Fuggi, A. Lignin and cellulose degradation and nitrogen dynamics during decomposition of three leaf litter species in a mediterranean ecosystem. Soil Biol. Biochem. 2005, 37, 1083–1091. [Google Scholar] [CrossRef]
- Li, H.; Wu, F.; Yang, W.; Xu, L.; Ni, X.; He, J.; Tan, B.; Hu, Y. Effects of forest gaps on litter lignin and cellulose dynamics vary seasonally in an alpine forest. Forests 2016, 7. [Google Scholar] [CrossRef]
- Talbot, J.M.; Treseder, K.K. Interactions among lignin, cellulose, and nitrogen drive litter chemistry-decay relationships. Ecology 2012, 93, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Coûteaux, M.-M.; Bottner, P.; Berg, B. Litter decomposition, climate and liter quality. Trends Ecol. Evol. 1995, 10, 63–66. [Google Scholar] [CrossRef]
- Hättenschwiler, S.; Jørgensen, H.B. Carbon quality rather than stoichiometry controls litter decomposition in a tropical rain forest. J. Ecol. 2010, 98, 754–763. [Google Scholar] [CrossRef]
- Cornwell, W.K.; Cornelissen, J.H.C.; Amatangelo, K.; Dorrepaal, E.; Eviner, V.T.; Godoy, O.; Hobbie, S.E.; Hoorens, B.; Kurokawa, H.; Pérez-Harguindeguy, N.; et al. Plant species traits are the predominant control on litter decomposition rates within biomes worldwide. Ecol. Lett. 2008, 11, 1065–1071. [Google Scholar] [CrossRef] [PubMed]
- Makkonen, M.; Berg, M.P.; Handa, I.T.; Hättenschwiler, S.; van Ruijven, J.; van Bodegom, P.M.; Aerts, R. Highly consistent effects of plant litter identity and functional traits on decomposition across a latitudinal gradient. Ecol. Lett. 2012, 15, 1033–1041. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.A.; Berg, B.; Maynard, D.S.; Wieder, W.R.; Wood, S.A. Understanding the dominant controls on litter decomposition. J. Ecol. 2016, 104, 229–238. [Google Scholar] [CrossRef]
- Bradford, M.A.; Warren, R.J., II; Baldrian, P.; Crowther, T.W.; Maynard, D.S.; Oldfield, E.E.; Wieder, W.R.; Wood, S.A.; King, J.R. Climate fails to predict wood decomposition at regional scales. Nat. Clim. Chang. 2014, 4, 625–630. [Google Scholar] [CrossRef]
- Wardle, D.A.; Bardgett, R.D.; Klironomos, J.N.; Setälä, H.; van der Putten, W.H.; Wall, D.H. Ecological linkages between aboveground and belowground biota. Science 2004, 304, 1629–1633. [Google Scholar] [CrossRef] [PubMed]
- Boyero, L.; Pearson, R.G.; Gessner, M.O.; Barmuta, L.A.; Ferreira, V.; Graça, M.A.S.; Dudgeon, D.; Boulton, A.J.; Callisto, M.; Chauvet, E.; et al. A global experiment suggests climate warming will not accelerate litter decomposition in streams but might reduce carbon sequestration. Ecol. Lett. 2011, 14, 289–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Palacios, P.; Maestre, F.T.; Kattge, J.; Wall, D.H. Climate and litter quality differently modulate the effects of soil fauna on litter decomposition across biomes. Ecol.Lett. 2013, 16, 1045–1053. [Google Scholar] [CrossRef] [PubMed]
- Graça, M.A.; Ferreira, V.; Canhoto, C.; Encalada, A.C.; Guerrero-Bolaño, F.; Wantzen, K.M.; Boyero, L. A conceptual model of litter breakdown in low order streams. Int. Rev. Hydrobiol. 2015, 100, 1–12. [Google Scholar] [CrossRef]
- Wagener, S.M.; Oswood, M.W.; Schimel, J.P. Rivers and soils: Parallels in carbon and nutrient processing. BioScience 1998, 48, 104–108. [Google Scholar] [CrossRef]
- Loreau, M.; MOuquet, N.; Holt, R.D. Meta-ecosystems: A theoretical framework for a spatial ecosystem ecology. Ecol. Lett. 2003, 6, 673–679. [Google Scholar] [CrossRef]
- Battin, T.J.; Kaplan, L.A.; Findlay, S.; Hopkinson, C.S.; Marti, E.; Packman, A.I.; Newbold, J.D.; Sabater, F. Biophysical controls on organic carbon fluxes in fluvial networks. Nat. Geosci. 2008, 1, 95–100. [Google Scholar] [CrossRef]
- Battin, T.J.; Luyssaert, S.; Kaplan, L.A.; Aufdenkampe, A.K.; Richter, A.; Tranvik, L.J. The boundless carbon cycle. Nat. Geosci. 2009, 2, 598–600. [Google Scholar] [CrossRef]
- Gessner, M.O.; Swan, C.M.; Dang, C.K.; McKie, B.G.; Bardgett, R.D.; Wall, D.H.; Hättenschwiler, S. Diversity meets decomposition. Trends Ecol. Evol. 2010, 25, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Wallace, J.B.; Eggert, S.L.; Meyer, J.L.; Webster, J.R. Multiple trophic levels of a forest stream linked to terrestrial litter inputs. Science 1997, 277, 102–104. [Google Scholar] [CrossRef]
- Chapin, F.S., III; Matson, P.A.; Vitousek, P. Principles of Terrestrial Ecosystem Ecology, 2nd ed.; Springer: New York, NY, USA, 2011. [Google Scholar]
- Zhu, J.; He, X.; Wu, F.; Yang, W.; Tan, B. Decomposition of abies faxoniana litter varies with freeze–thaw stages and altitudes in subalpine/alpine forests of southwest China. Scand. J. For. Res. 2012, 27, 586–596. [Google Scholar] [CrossRef]
- Yue, K.; Yang, W.; Peng, Y.; Zhang, C.; Huang, C.; Wu, F. Chromium, cadmium, and lead dynamics during winter foliar litter decomposition in an alpine forest river. Arct. Antarct. Alp. Res. 2016, 48, 79–91. [Google Scholar] [CrossRef]
- Olsson, P.Q.; Sturm, M.; Racine, C.H.; Romanovsky, V.; Liston, G.E. Five stages of the alaskan arctic cold season with ecosystem implications. Arct. Antarct. Alp. Res. 2003, 35, 74–81. [Google Scholar] [CrossRef]
- Yue, K.; Yang, W.; Peng, C.; Peng, Y.; Zhang, C.; Huang, C.; Tan, Y.; Wu, F. Foliar litter decomposition in an alpine forest meta-ecosystem on the eastern Tibetan plateau. Sci. Total Environ. 2016, 566–567, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Peng, C.; Zhu, J.; Zhang, J.; Tan, B.; Yang, W. Impact of freezing and thawing dynamics on foliar litter carbon release in alpine/subalpine forests along an altitudinal gradient in the eastern Tibetan plateau. Biogeosciences 2014, 11, 6471–6481. [Google Scholar]
- Yue, K.; Yang, W.; Peng, Y.; Zhang, C.; Huang, C.; Xu, Z.; Tan, B.; Wu, F. Dynamics of multiple metallic elements during foliar litter decomposition in an alpine forest river. Ann. For. Sci. 2016, 73, 547–557. [Google Scholar] [CrossRef]
- American Public Health Association; American Water Works Association; Water Environment Federation. Standard Methods for the Examination of Water and Wastewater, 22nd ed.; American Public Health Association: Washington, DC, USA, 2012. [Google Scholar]
- He, W.; Wu, F.; Yang, W.; Tan, B.; Zhao, Y.; Wu, Q.; He, M. Lignin degradation in foliar litter of two shrub species from the gap center to the closed canopy in an alpine fir forest. Ecosystems 2016, 19, 115–128. [Google Scholar] [CrossRef]
- Lovett, G.M.; Arthur, M.A.; Weathers, K.C.; Fitzhugh, R.D.; Templer, P.H. Nitrogen addition increases carbon storage in soils, but not in trees, in an eastern US deciduous forest. Ecosystems 2013, 16, 980–1001. [Google Scholar] [CrossRef]
- Boyero, L.; Pearson, R.G.; Hui, C.; Gessner, M.O.; Pérez, J.; Alexandrou, M.A.; Graça, M.A.; Cardinale, B.J.; Albariño, R.J.; Arunachalam, M. Biotic and abiotic variables influencing plant litter breakdown in streams: A global study. Proc. R. Soc. B 2016, 283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cotrufo, M.F.; Wallenstein, M.D.; Boot, C.M.; Denef, K.; Paul, E. The microbial efficiency-matrix stabilization (mems) framework integrates plant litter decomposition with soil organic matter stabilization: Do labile plant inputs form stable soil organic matter? Glob. Chang. Biol. 2013, 19, 988–995. [Google Scholar] [CrossRef] [PubMed]
- Herman, J.; Moorhead, D.; Berg, B. The relationship between rates of lignin and cellulose decay in aboveground forest litter. Soil Biol. Biochem. 2008, 40, 2620–2626. [Google Scholar] [CrossRef]
- Klotzbücher, T.; Kaiser, K.; Guggenberger, G.; Gatzek, C.; Kalbitz, K. A new conceptual model for the fate of lignin in decomposing plant litter. Ecology 2011, 92, 1052–1062. [Google Scholar] [CrossRef] [PubMed]
- Halvorson, H.M.; Scott, E.E.; Entrekin, S.A.; Evans-White, M.A.; Scott, J.T. Light and dissolved phosphorus interactively affect microbial metabolism, stoichiometry and decomposition of leaf litter. Freshw. Biol. 2016, 61, 1006–1019. [Google Scholar] [CrossRef]
- García-Palacios, P.; McKie, B.G.; Handa, I.T.; Frainer, A.; Hättenschwiler, S. The importance of litter traits and decomposers for litter decomposition: A comparison of aquatic and terrestrial ecosystems within and across biomes. Funct. Ecol. 2016, 30, 819–829. [Google Scholar] [CrossRef]
- Nakajima, T.; Asaeda, T.; Fujino, T.; Nanda, A. Leaf litter decomposition in aquatic and terrestrial realms of a second-order forested stream system. J. Freshw. Ecol. 2006, 21, 259–263. [Google Scholar] [CrossRef]
- Berg, B.; Kjønaas, O.; Johansson, M.-B.; Erhagen, B.; Åkerblom, S. Late stage pine litter decomposition: Relationship to litter N, Mn, and acid unhydrolyzable residue (aur) concentrations and climatic factors. For. Ecol. Manag. 2015, 358, 41–47. [Google Scholar] [CrossRef]
- Dalias, P.; Anderson, J.; Bottner, P.; Coûteaux, M.-M. Long-term effects of temperature on carbon mineralisation processes. Soil Biol. Biochem. 2001, 33, 1049–1057. [Google Scholar] [CrossRef]
- Štursová, M.; Žifčáková, L.; Leigh, M.B.; Burgess, R.; Baldrian, P. Cellulose utilization in forest litter and soil: Identification of bacterial and fungal decomposers. FEMS Microbiol. Ecol. 2012, 80, 735–746. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Ecosystem | Variable | ||||||
|---|---|---|---|---|---|---|---|
| DAT (°C) | C (g/kg) | N (g/kg) | P (g/kg) | pH | |||
| Forest floor | 2.0 (5.2) | 126 (26) | 5.8 (1.1) | 1.2 (0.2) | 6.6 (0.1) | ||
| DAT (°C) | HCO3− (mg/L) | NH4+ (mg/L) | NO3− (mg/L) | PO43− (μg/L) | pH | FV (m/s) | |
| Stream | 5.1 (2.6) | 13.9 (1.96) | 0.10 (0.05) | 0.29 (0.07) | 7.85 (0.38) | 6.6 (0.4) | 0.53 (0.15) |
| Riparian zone | 4.8 (3.4) | 19.7 (1.33) | 0.04 (0.02) | 0.34 (0.08) | 7.84 (0.41) | 6.9 (0.3) | 0.05 (0.01) |
| Species | C (%) | N (%) | P (%) | Lignin (%) | Cellulose (%) | C/N | C/P | N/P | Lignin/N |
|---|---|---|---|---|---|---|---|---|---|
| Willow | 34.8 | 2.64 | 0.17 | 24.7 | 18.6 | 13.2 | 207 | 15.7 | 9.38 |
| (0.9) c | (0.15) a | (0.01) a | (1.3) d | (0.4) c | (0.8) d | (20) c | (1.7) a | (0.84) c | |
| Azalea | 38.6 | 0.69 | 0.10 | 29.8 | 19.9 | 57.2 | 375 | 6.75 | 44.3 |
| (1.0) b | (0.10) d | (0.02) d | (0.8) b | (0.3) a | (10.2) a | (54) a | (1.5) c | (8.3) a | |
| Cypress | 46.9 | 1.05 | 0.15 | 28.1 | 19.0 | 45.1 | 304 | 6.79 | 26.9 |
| (1.8) a | (0.06) c | (0.01) b | (0.8) c | (0.2) b | (3.9) b | (13) b | (0.7) c | (1.8) b | |
| Larch | 37.5 | 1.59 | 0.12 | 37.8 | 14.9 | 23.6 | 320 | 13.6 | 30.1 |
| (0.5) b | (0.11) b | (0.01) c | (0.9) a | (0.2) d | (1.8) c | (25) b | (0.8) b | (2.1) b |
| Factor | d.f. | F-Value | p-Value |
|---|---|---|---|
| Species | 3 | 23.439 | <0.001 |
| Ecosystem | 2 | 83.765 | <0.001 |
| Period | 9 | 1334.260 | <0.001 |
| Species × ecosystem | 6 | 5.932 | <0.001 |
| Species × period | 27 | 17.349 | <0.001 |
| Ecosystem × period | 27 | 19.603 | <0.001 |
| Species × ecosystem × period | 54 | 6.276 | <0.001 |
| Ecosystem | Regression Model | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Step 1 | Step 2 | Step 3 | Step 4 | |||||||
| a0 | a1X1 | R12 | a2X2 | R22 | a3X3 | R32 | a4X4 | R42 | ||
| Forest | ŷ = | 2.438 | +8.091 P | (0.687) | −0.028 lignin | (0.795) | +0.035 N/P | (0.866) | ||
| Stream | ŷ = | −1.895 | +6.927 P | (0.748) | +0.055 N/P | (0.899) | +0.173 cellulose | (0.922) | +0.019 C | (0.937) |
| Riparian zone | ŷ = | 2.164 | +3.466 P | (0.644) | −0.026 lignin | (0.771) | +0.050 N/P | (0.855) | +0.021 C | (0.881) |
| Species | DAT | C | N | P | pH | ||
|---|---|---|---|---|---|---|---|
| Forest floor | |||||||
| Willow | (−) 14.809 *** | 1.316 | (−) 19.318 *** | 0.636 | (+) 5.923 * | ||
| Azalea | (−) 10.701 ** | 3.301 | 3.583 | 1.073 | 2.196 | ||
| Cypress | (−) 9.890 ** | 0.909 | (−) 9.663 ** | (−) 8.960 ** | 2.474 | ||
| Larch | 3.596 | 0.464 | (−) 12.819 ** | (−) 17.690 *** | 3.391 | ||
| DAT | HCO3− | NH4+ | NO3− | PO43− | pH | FV | |
| Stream | |||||||
| Willow | 3.551 | 0.019 | 0.331 | (+) 5.567 * | 1.696 | 2.112 | 3.036 |
| Azalea | 1.784 | (+) 10.305 ** | (+) 8.100 ** | 1.846 | 0.026 | (+) 10.514 ** | 0.322 |
| Cypress | 2.638 | 0.837 | 1.794 | (+) 7.335 ** | 0.058 | 0.095 | 0.001 |
| Larch | 1.864 | 1.268 | 0.440 | 0.227 | 0.571 | (+) 5.835 * | 0.550 |
| Riparian zone | |||||||
| Willow | (−) 27.150 *** | 3.069 | (−) 13.888 *** | (+) 9.471 ** | 0.347 | 2.042 | (−) 14.576 *** |
| Azalea | (−) 34.288 *** | 0.424 | (−) 12.601 ** | 0.356 | 1.014 | (+) 7.886 ** | 2.420 |
| Cypress | (−) 16.155 *** | (+) 4.815 * | (−) 10.753 ** | 3.791 | 0.215 | 0.082 | (−) 12.630 ** |
| Larch | (−) 22.839 *** | 0.802 | (−) 10.324 ** | 1.305 | 0.007 | 2.010 | (−) 4.679 * |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yue, K.; Wu, F.; Yang, W.; Zhang, C.; Peng, Y.; Tan, B.; Xu, Z.; Huang, C. Cellulose Dynamics during Foliar Litter Decomposition in an Alpine Forest Meta-Ecosystem. Forests 2016, 7, 176. https://doi.org/10.3390/f7080176
Yue K, Wu F, Yang W, Zhang C, Peng Y, Tan B, Xu Z, Huang C. Cellulose Dynamics during Foliar Litter Decomposition in an Alpine Forest Meta-Ecosystem. Forests. 2016; 7(8):176. https://doi.org/10.3390/f7080176
Chicago/Turabian StyleYue, Kai, Fuzhong Wu, Wanqin Yang, Chuan Zhang, Yan Peng, Bo Tan, Zhenfeng Xu, and Chunping Huang. 2016. "Cellulose Dynamics during Foliar Litter Decomposition in an Alpine Forest Meta-Ecosystem" Forests 7, no. 8: 176. https://doi.org/10.3390/f7080176