
Analysing Atmospheric Processes and Climatic Drivers of Tree Defoliation to Determine Forest Vulnerability to Climate Warming

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
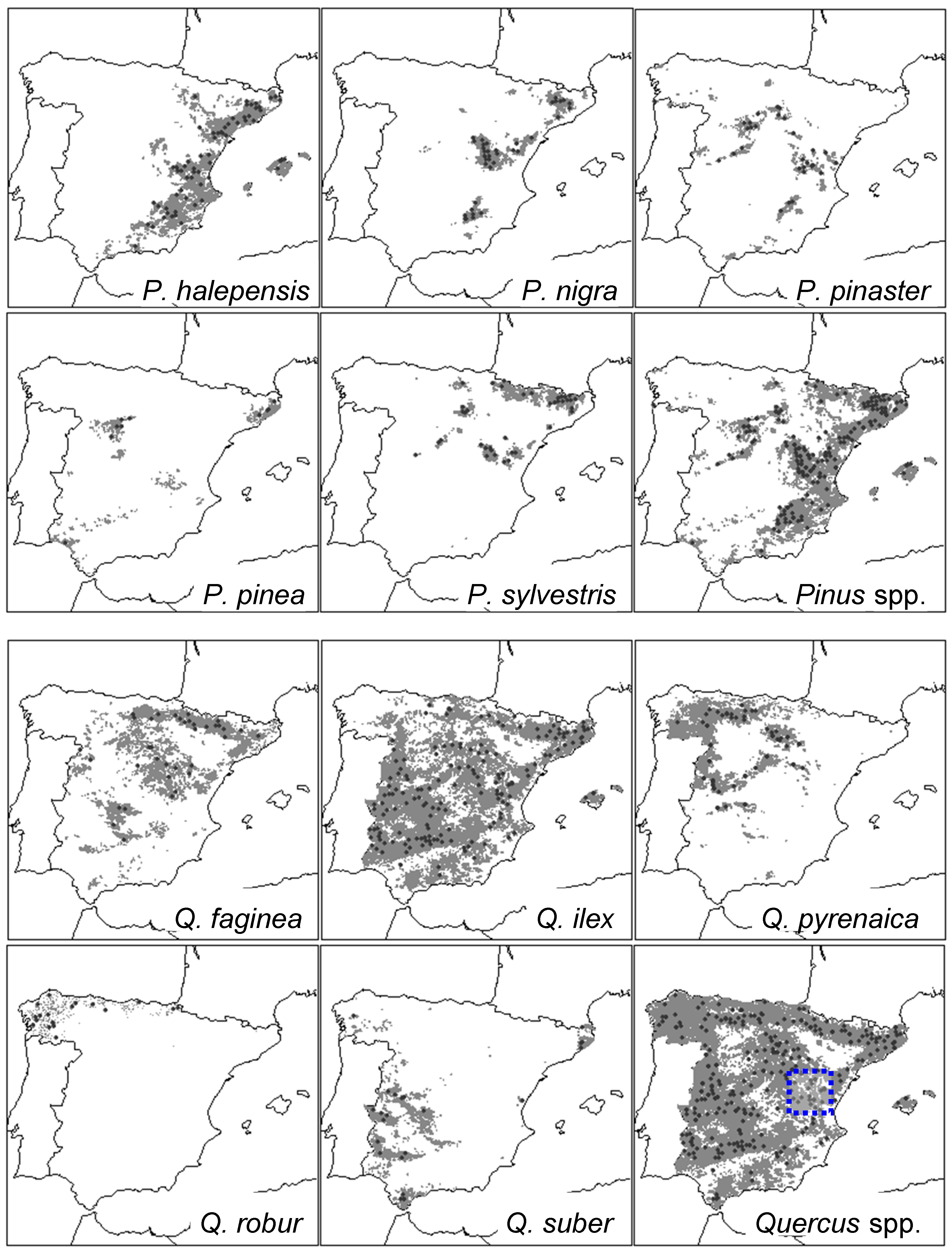
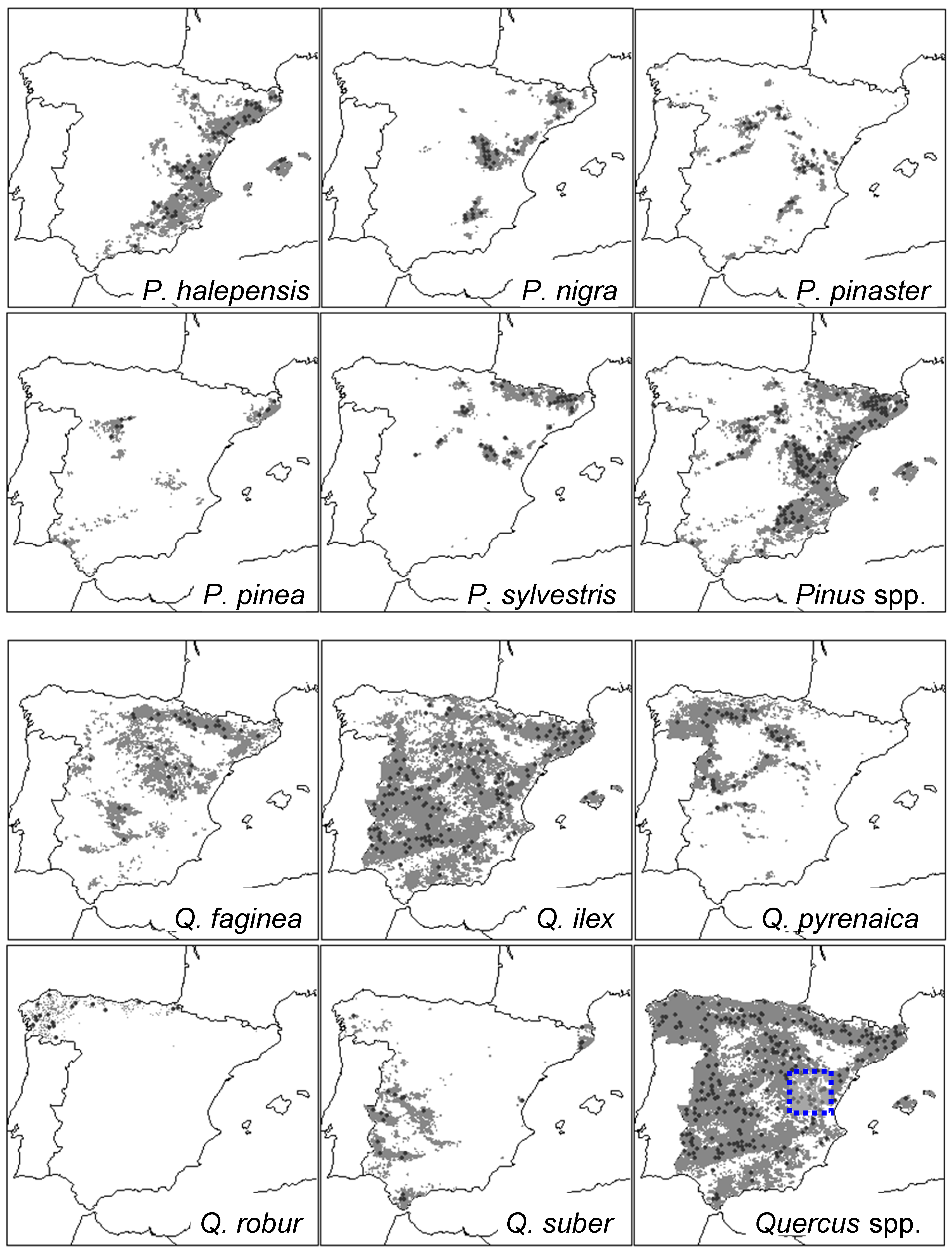
2.1. Study Area and Tree Species
2.2. Defoliation Data
2.3. Radial Growth Data
2.4. Climate Data, Atmospheric Processes, and SPEI Drought Index
2.5. Statistical Analyses
3. Results
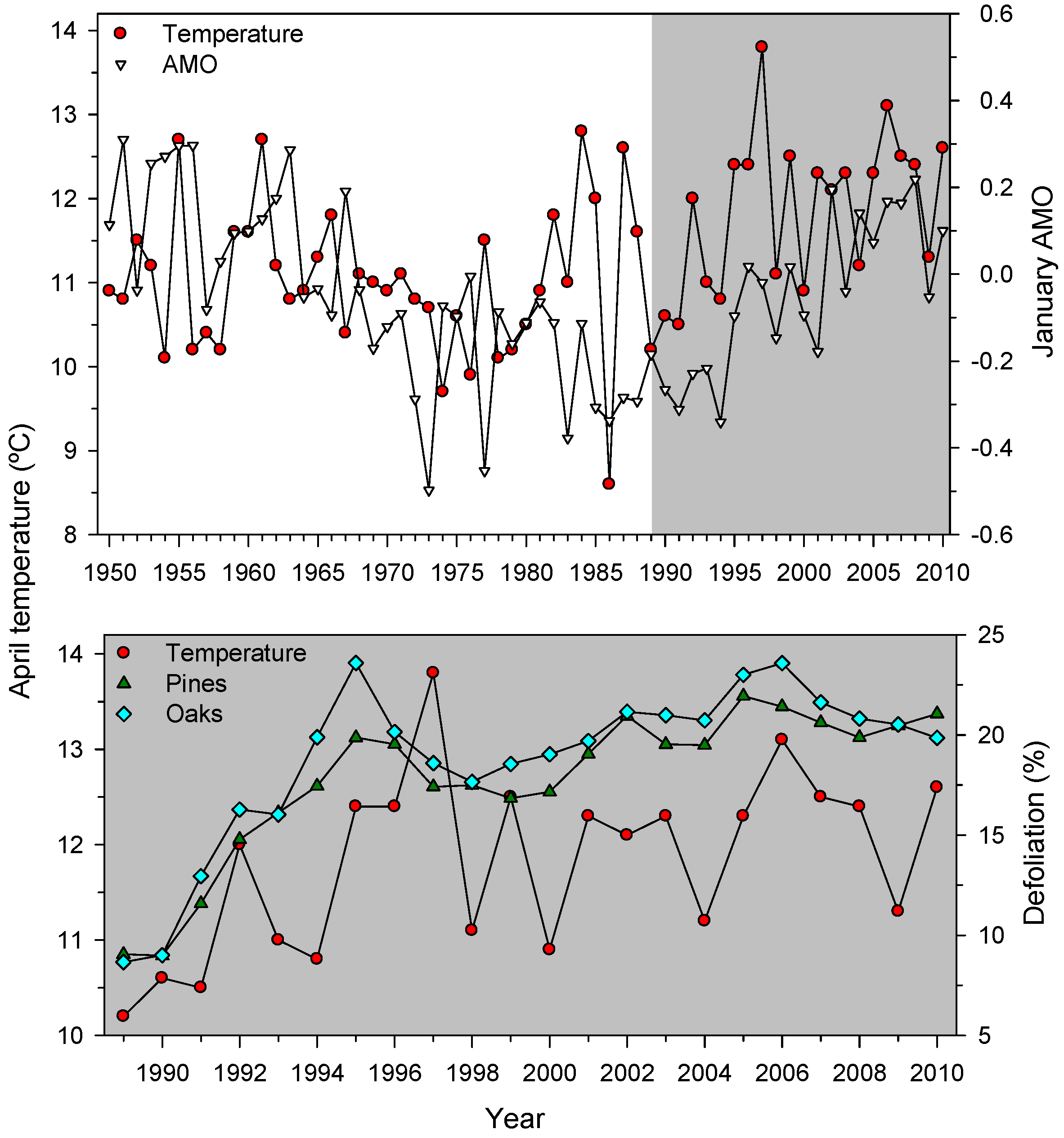
3.1. Patterns and Trends of Defoliation Data
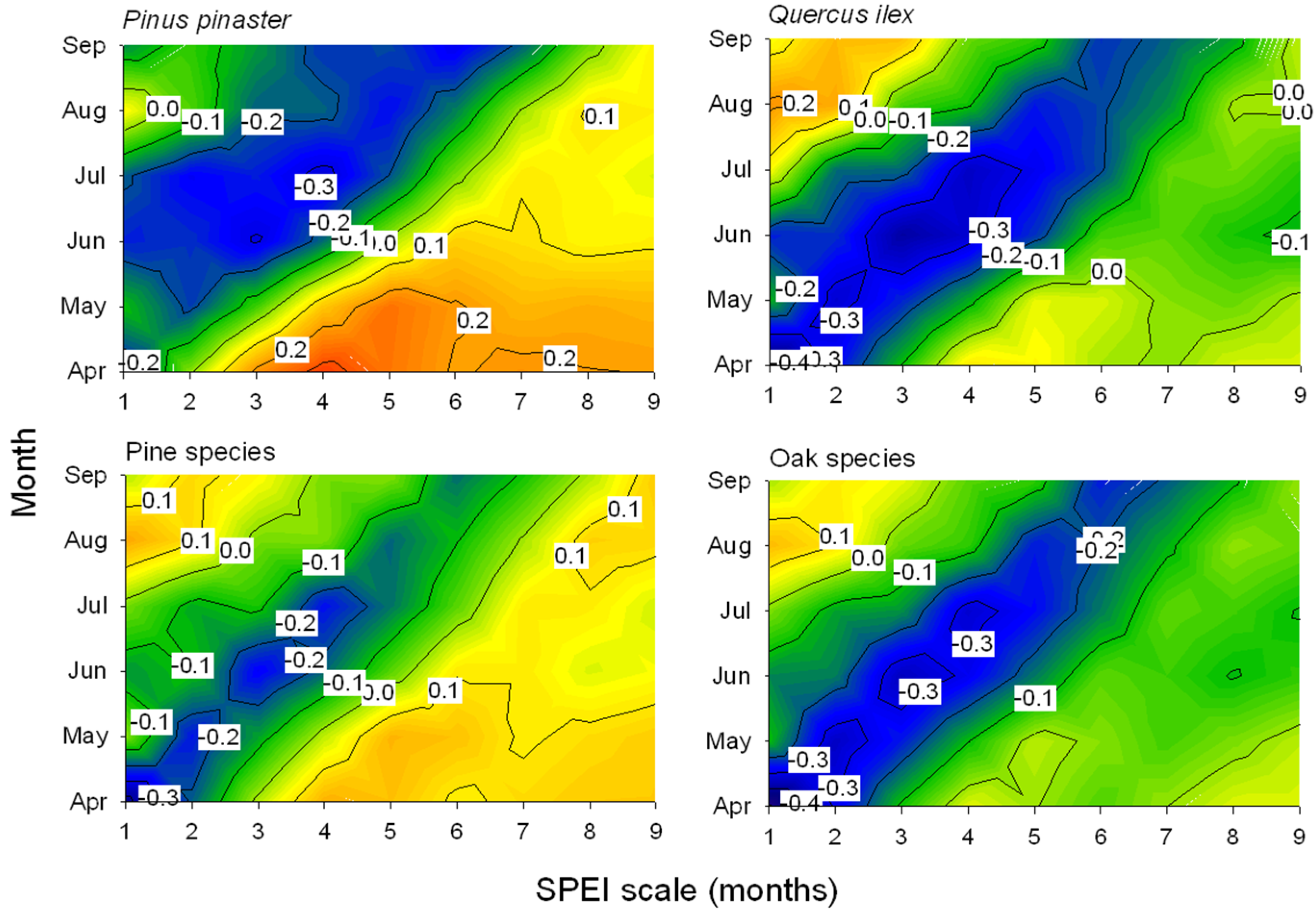
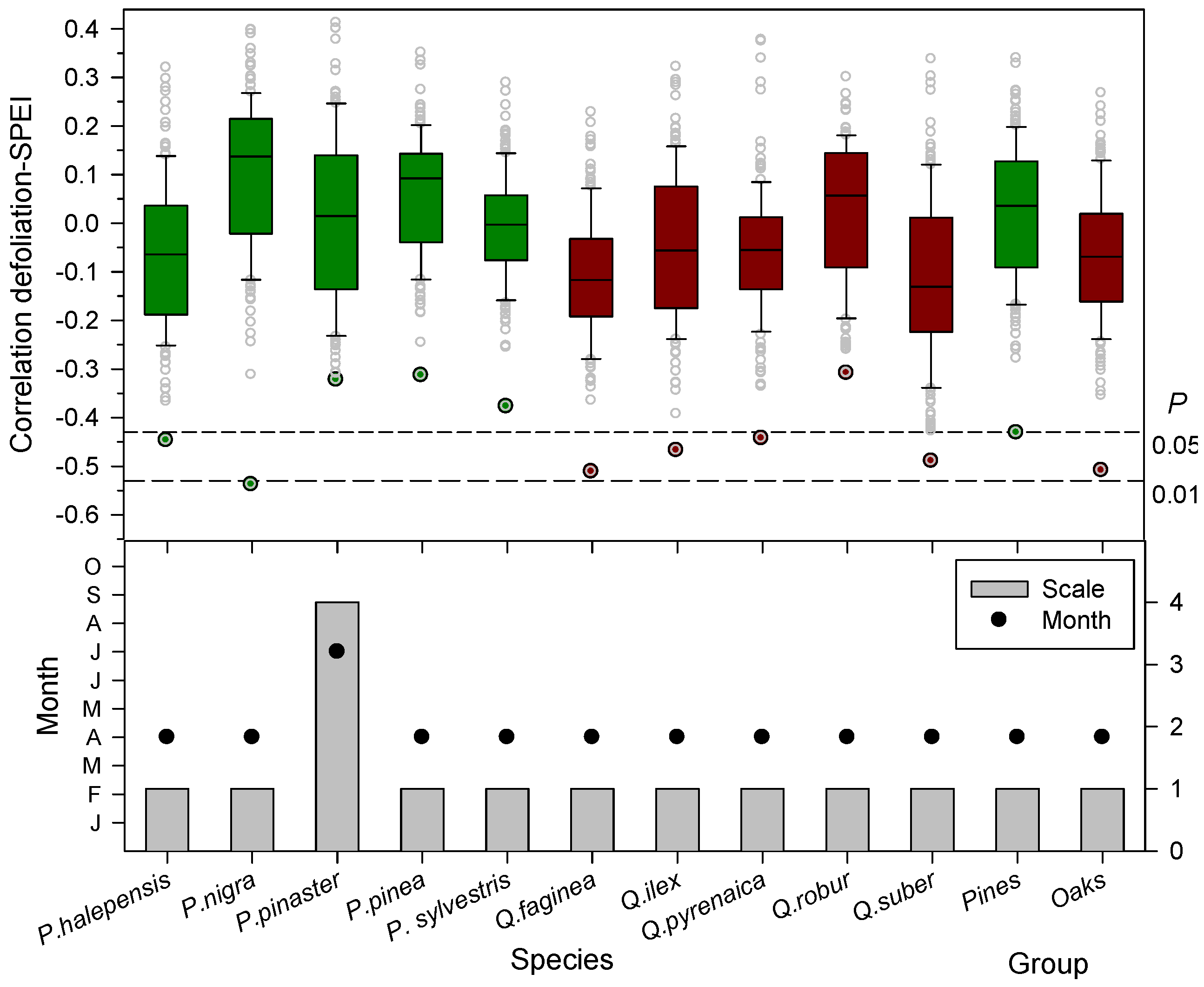
3.2. Associations between Defoliation, Climate and Drought
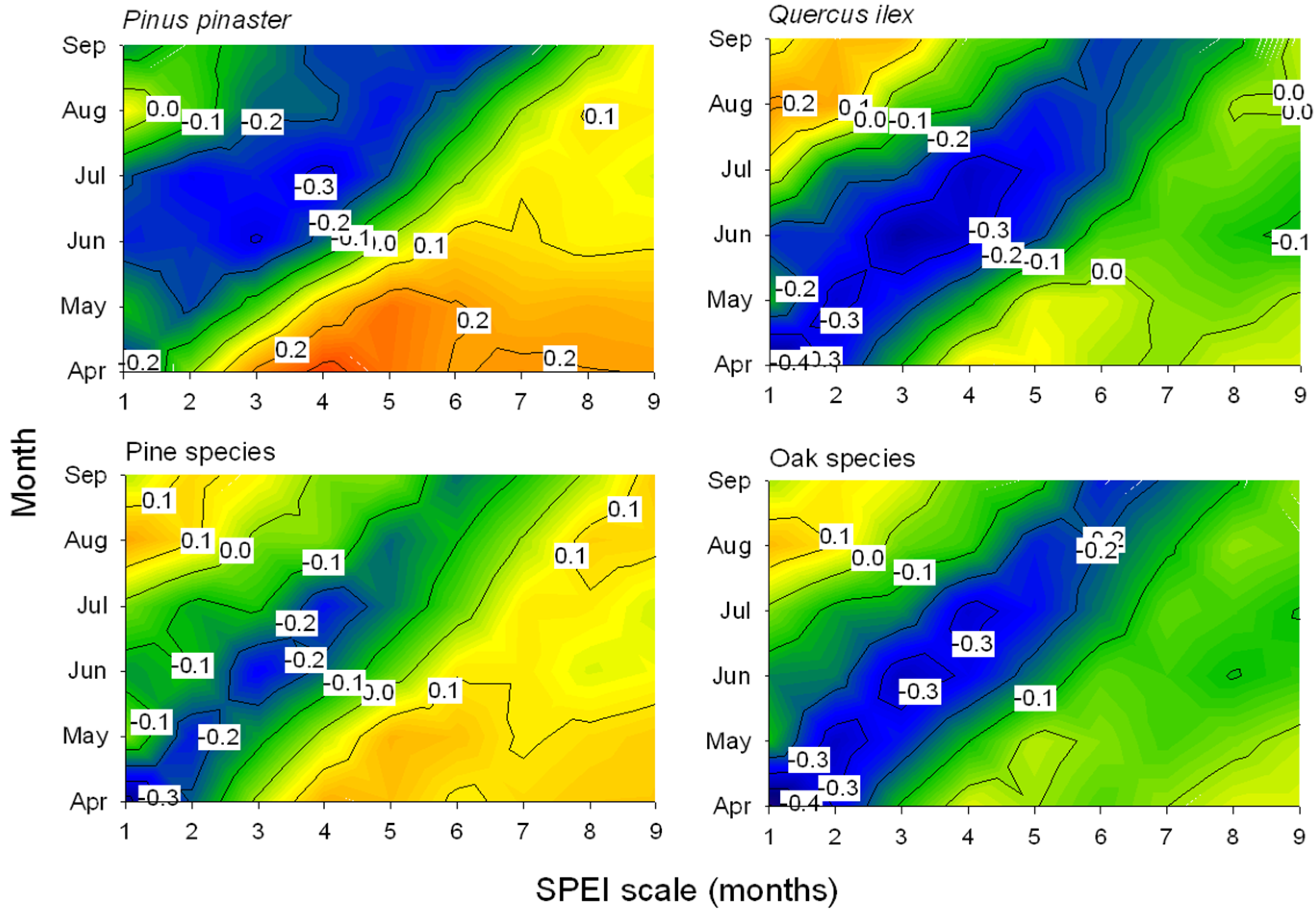
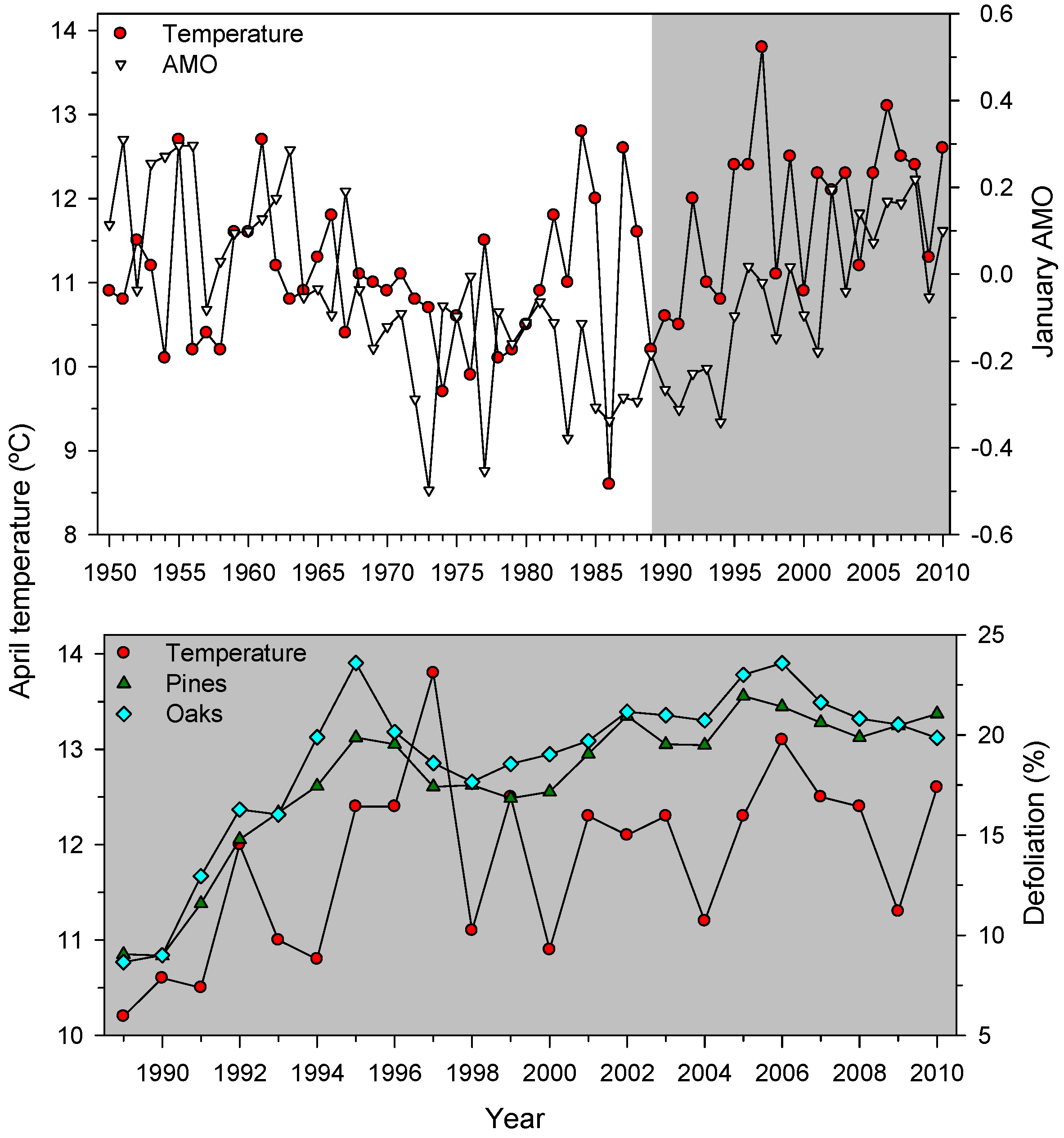
3.3. Associations between Defoliation and Atmospheric Processes
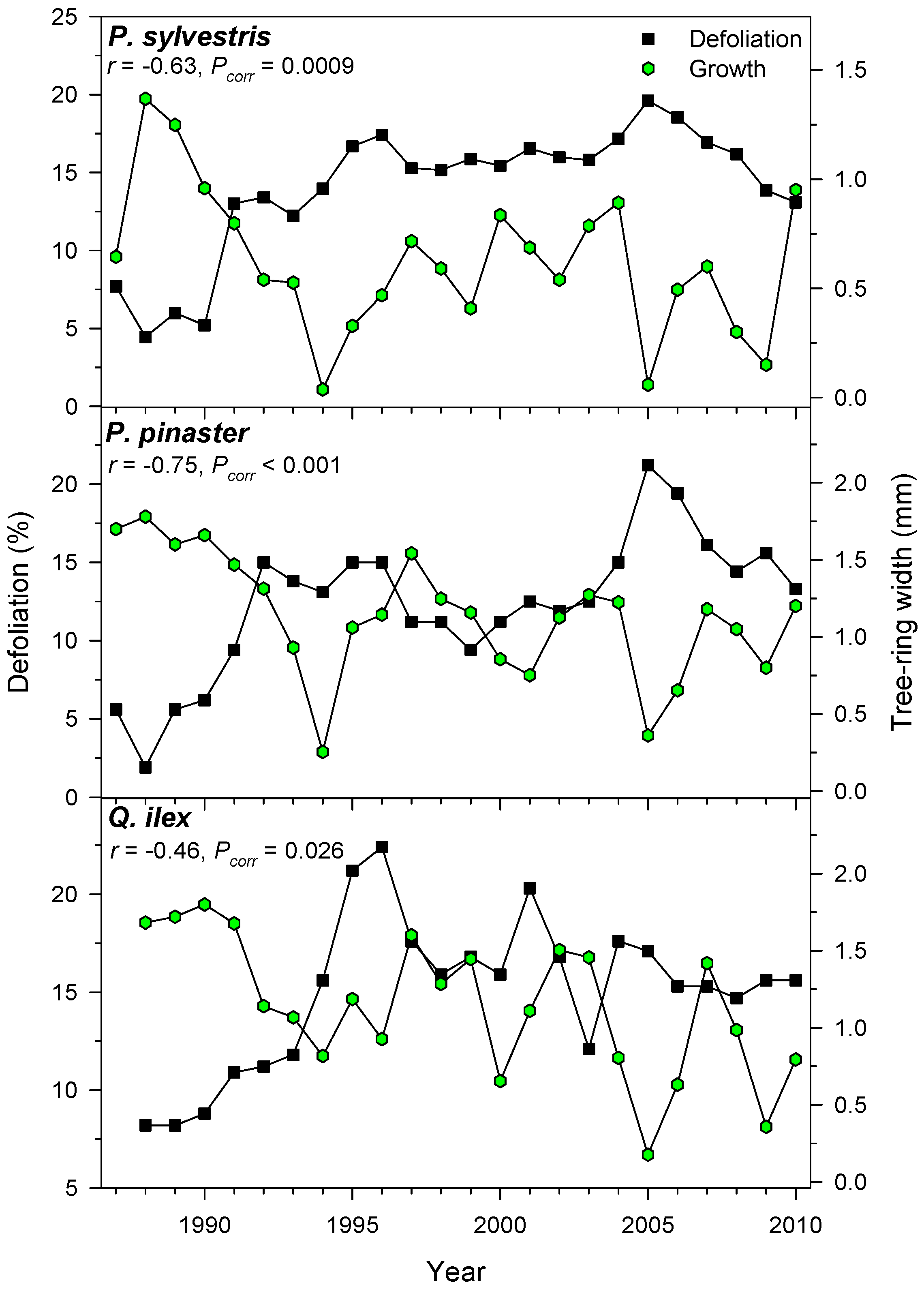
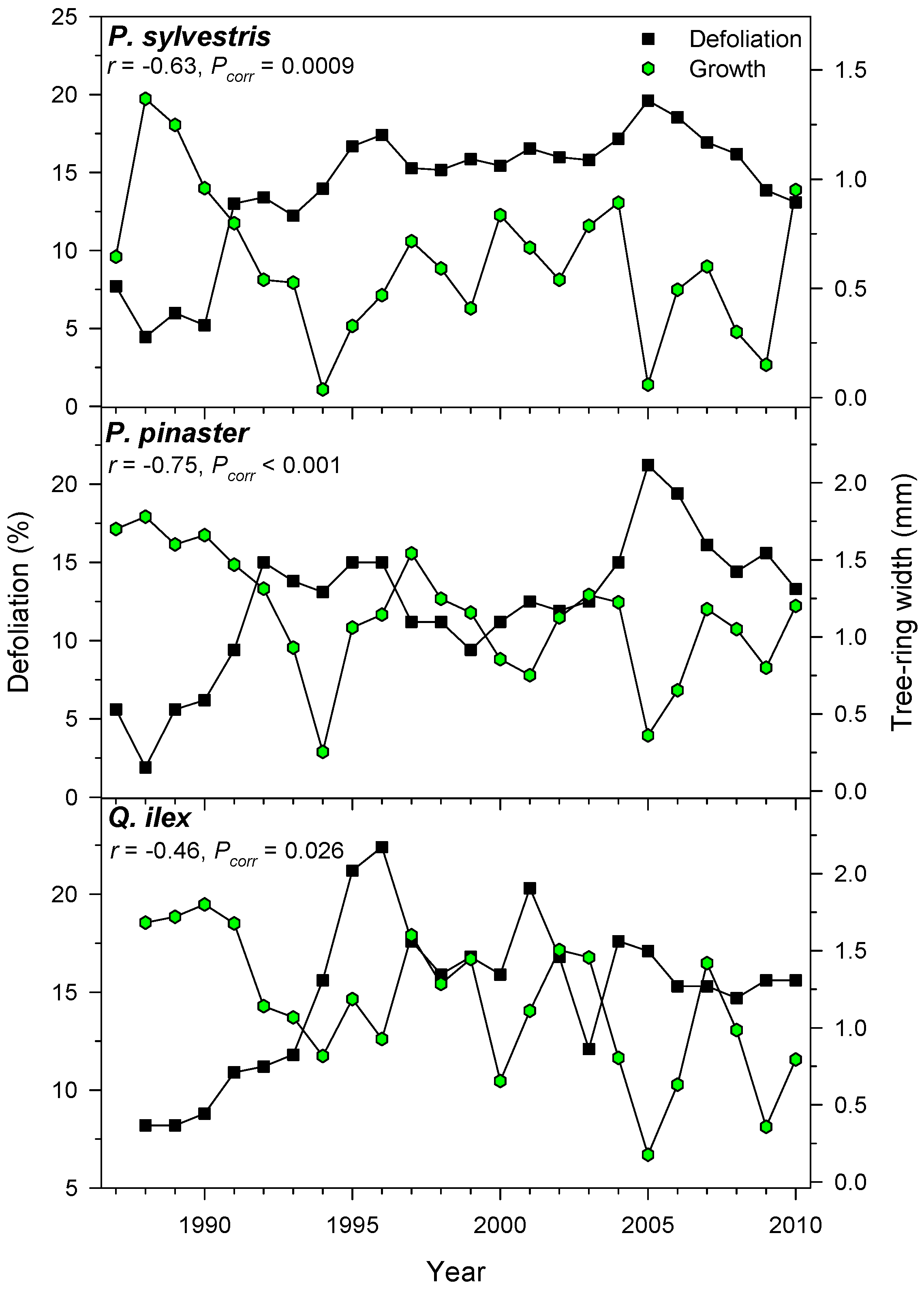
3.4. Associations between Defoliation and Tree Growth
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Intergovernmental Panel on Climate Change. Managing the Risks of Extreme Events and Disasters to Advance Climate Change Adaptation; Field, C.B., Barros, V., Stocker, T.F., Qin, D., Dokken, D.J., Ebi, K.L., Mastrandrea, M.D., Mach, K.J., Plattner, G.-K., Allen, S.K., et al., Eds.; A Special Report of Working Groups I and II of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2012; pp. 1–582. [Google Scholar]
- Yao, Y.; Luo, Y.; Huang, J.; Zhao, Z. Comparison of monthly temperature extremes simulated by CMIP3 and CMIP5 models. J. Clim. 2013, 26, 7692–7707. [Google Scholar] [CrossRef]
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogée, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, Chr.; Carrara, A.; et al. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature 2005, 437, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Bréda, N.; Huc, R.; Granier, A.; Dreyer, E. Temperate forest trees and stands under severe drought: A review of ecophysiological responses, adaptation processes and long-term consequences. Ann. For. Sci. 2006, 63, 625–644. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 2015, 6, 129. [Google Scholar] [CrossRef]
- Dobbertin, M. Tree growth as indicator of tree vitality and of tree reaction to environmental stress: A review. Eur. J. For. Res. 2005, 124, 319–333. [Google Scholar] [CrossRef]
- Teskey, R.; Wertin, T.; Bauweraerts, I.; Ameye, M.; McGuire, M.A.; Steppe, K. Responses of tree species to heat waves and extreme heat events. Plant. Cell Environ. 2015, 38, 1699–1712. [Google Scholar] [CrossRef] [PubMed]
- Filewod, B.; Thomas, S.C. Impacts of a spring heat wave on canopy processes in a northern hardwood forest. Glob. Chang. Biol. 2014, 20, 360–371. [Google Scholar] [CrossRef] [PubMed]
- Camarero, J.J.; Franquesa, M.; Sangüesa-Barreda, G. Timing of drought triggers distinct growth responses in holm oak: Implications to predict warming-induced forest defoliation and growth decline. Forests 2015, 6, 1576–1597. [Google Scholar] [CrossRef]
- Heide, O.M. High autumn temperature delays spring bud burst in boreal trees, counterbalancing the effect of climatic warming. Tree Physiol. 2003, 23, 931–936. [Google Scholar] [CrossRef] [PubMed]
- Solberg, S. Summer drought: A driver for crown condition and mortality of Norway spruce in Norway. For. Pathol. 2004, 34, 93–104. [Google Scholar] [CrossRef]
- Seidling, W. Signals of summer drought in crown condition data from the German Level I network. Eur. J. For. Res. 2007, 126, 529–544. [Google Scholar] [CrossRef]
- Carnicer, J.; Coll, M.; Ninyerola, M.; Pons, X.; Sánchez, G.; Peñuelas, J. Widespread crown condition decline, food web disruption, and amplified tree mortality with increased climate change-type drought. Proc. Natl. Acad. Sci. USA 2011, 108, 1474–1478. [Google Scholar] [CrossRef] [PubMed]
- Seidling, W.; Ziche, D.; Beck, W. Climate responses and interrelations of stem increment and crown transparency in Norway spruce, Scots pine, and common beech. For. Ecol. Manag. 2012, 284, 196–204. [Google Scholar] [CrossRef]
- Ferretti, M.; Nicolas, M.; Bacaro, G.; Brunialti, G.; Calderisi, M.; Croisé, L.; Frati, L.; Lanier, M.; Maccherini, S.; Santi, E.; et al. Plot-scale modelling to detect size, extent, and correlates of changes in tree defoliation in French high forests. For. Ecol. Manag. 2014, 311, 56–69. [Google Scholar] [CrossRef]
- De la Cruz, A.C.; Gil, P.M.; Fernandez-Cancio, A.; Minaya, M.; Navarro-Cerrillo, R.M.; Sanchez-Salguero, R.; Grau, J.M. Defoliation triggered by climate induced effects in Spanish ICP Forests monitoring plots. For. Ecol. Manag. 2014, 331, 245–255. [Google Scholar] [CrossRef]
- Solberg, S.; Aamlid, D.; Tveito, O.E.; Lystad, S. Increased needle fall and defoliation in Norway spruce induced by warm and dry weather. Boreal Environ. Res. 2015, 20, 335–349. [Google Scholar]
- Drobyshev, I.; Linderson, H.; Sonesson, K. Relationship between crown condition and tree diameter growth in Southern Swedish oaks. Environ. Monit. Assess. 2007, 128, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Salguero, R.; Camarero, J.J.; Dobbertin, M.; Fernández-Cancio, A.; Vilà-Cabrera, A.; Manzanedo, R.D.; Zavala, M.A.; Navarro-Cerillo, R.M. Contrasting vulnerability and resilience to drought-induced decline of densely planted vs. natural rear-edge Pinus nigra forests. For. Ecol. Manag. 2013, 310, 956–967. [Google Scholar] [CrossRef]
- Camarero, J.J. Direct and Indirect Effects of the North Atlantic Oscillation on Tree Growth and Forest Decline in Northeastern Spain. In Hydrological, Socioeconomic and Ecological Impacts of the North Atlantic Oscillation in the Mediterranean Region; Vicente-Serrano, S.M., Trigo, R.M., Eds.; Advances in Global Change Research; Springer: Berlin/Heidelberg, Germany, 2011; Volume 46, pp. 129–152. [Google Scholar]
- Gordo, O.; Sanz, J.J. Impact of climate change on plant phenology in Mediterranean ecosystems. Glob. Chang. Biol. 2010, 16, 1082–1106. [Google Scholar] [CrossRef]
- Peñuelas, J.; Lloret, F.; Montoya, R. Severe drought effects on Mediterranean woody flora in Spain. For. Sci. 2001, 47, 214–218. [Google Scholar]
- Lloret, F.; Siscart, D.; Dalmases, C. Canopy recovery after drought dieback in holm-oak Mediterranean forests of Catalonia (NE Spain). Glob. Chang. Biol. 2004, 10, 2092–2099. [Google Scholar] [CrossRef]
- Sánchez-Salguero, R.; Navarro-Cerillo, R.M.; Camarero, J.J.; Fernández-Cancio, A. Selective drought-induced decline of pine species in southeastern Spain. Clim. Chang. 2012, 113, 767–785. [Google Scholar] [CrossRef]
- Camarero, J.J.; Gazol, A.; Sangüesa-Barreda, G.; Oliva, J.; Vicente-Serrano, S.M. To die or not to die: Early-warning signals of dieback in response to a severe drought. J. Ecol. 2015, 103, 44–57. [Google Scholar] [CrossRef]
- Camarero, J.J.; Sangüesa-Barreda, G.; Vergarechea, M. Prior height, growth, and wood anatomy differently predispose to drought-induced dieback in two Mediterranean oak species. Ann. For. Sci. 2016, 73, 341–351. [Google Scholar] [CrossRef]
- ICP Forests. International Co-Operative Programme on Assessment and Monitoring of Air Pollution Effects on Forests Operating under the UNECE Convention on Long-Range Transboundary Air Pollution, 2016. Available online: http://icp-forests.net/ (accessed on 12 October 2016).
- Blanco-Castro, E.; Casado González, M.A.; Costa Tenorio, M. Los Bosques Ibéricos: Una Interpretación Geobotánica; GeoPlaneta: Barcelona, Spain, 1997. [Google Scholar]
- Ninyerola, M.; Pons, X.; Roure, J.M. Monthly precipitation mapping of the Iberian peninsula using spatial interpolation tools implemented in a geographic information system. Theor. Appl. Climatol. 2006, 89, 195–209. [Google Scholar] [CrossRef]
- Eichhorn, J.; Roskams, P.; Ferretti, M.; Mues, V.; Szepesi, A.; Durrant, D. Visual Assessment of Crown Condition and Damaging Agents. Manual Part IV, In Manual on Methods and Criteria for Harmonized Sampling, Assessment, Monitoring and Analysis of the Effects of Air Pollution on Forests. UNECE ICP Forests Programme Coordinating Centre, Hamburg, 2010. Available online: http://www.icp-forests.org/Manual.htm (accessed on 12 October 2016).
- Camarero, J.J.; Gazol, A.; Tardif, J.C.; Conciatori, F. Attributing forest responses to global-change drivers: Limited evidence of a CO2-fertilization effect in Iberian pine growth. J. Biogeogr. 2015, 42, 2220–2233. [Google Scholar] [CrossRef]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 1983, 43, 69–78. [Google Scholar]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I.A. Multi-scalar drought index sensitive to global warming: The Standardized Precipitation Evapotranspiration Index—SPEI. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef]
- Beguería, S.; Latorre, B.; Reig, F.; Vicente-Serrano, S.M. SPEI Global Drought Monitor. Available online: http://sac.csic.es/spei/map/maps.html (accessed on 12 October 2016).
- Enfield, D.B.; Mestas-Nunez, A.M.; Trimble, P.J. The Atlantic Multidecadal Oscillation and its relationship to rainfall and river flows in the continental US. Geophys. Res. Lett. 2001, 28, 2077–2080. [Google Scholar] [CrossRef]
- Trenberth, K.; Shea, D.J. Atlantic hurricanes and natural variability in 2005. Geophys. Res. Lett. 2006, 33, L12704. [Google Scholar] [CrossRef]
- Hurrell, J. Decadal trends in North Atlantic Oscillation and relationship to regional temperature and precipitation. Science 1995, 269, 676–679. [Google Scholar] [CrossRef] [PubMed]
- Sáenz, J.; Rodríguez-Puebla, C.; Fernández, J.; Zubillaga, J. Interpretation of interannual winter temperature variations over southwestern Europe. J. Geophys. Res. 2001, 106, 20641–20651. [Google Scholar] [CrossRef]
- Rodó, X.; Baert, E.; Comin, F. Variations in seasonal rainfall in Southern Europe during the present century: Relationships with the North Atlantic Oscillation and the El Niño-Southern Oscillation. Clim. Dyn. 1997, 13, 275–284. [Google Scholar] [CrossRef]
- ESRL-NOAA. Earth System Research Laboratory. Available online: http://www.esrl.noaa.gov/psd/ (accessed on 12 October 2016).
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S; Springer: New York, NY, USA, 2002. [Google Scholar]
- Mudelsee, M. Estimating Pearson’s correlation coefficient with bootstrap confidence interval from serial dependent time series. Math. Geol. 2002, 35, 651–665. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Ogaya, R.; Barbeta, A.; Basnou, C.; Penuelas, J. Satellite data as indicators of tree biomass growth and forest dieback in a Mediterranean holm oak forest. Ann. For. Sci. 2015, 72, 135–144. [Google Scholar] [CrossRef]
- Tyree, M.T.; Cochard, H.; Cruizat, P.; Sinclair, B.; Ameglio, T. Drought-induced leaf shedding in walnut: Evidence for vulnerability segmentation. Plant Cell Environ. 1993, 16, 879–882. [Google Scholar] [CrossRef]
- Montserrat-Martí, G.; Camarero, J.J.; Palacio, S.; Pérez-Rontomé, C.; Milla, R.; Albuixech, J.; Maestro, M. Summer-drought constrains the phenology and growth of two co-existing Mediterranean oaks with contrasting leaf habit: Implications for their persistence and reproduction. Trees Struct. Funct. 2009, 23, 787–799. [Google Scholar] [CrossRef]
- Corcuera, L.; Camarero, J.J.; Gil-Pelegrín, E. Effects of a severe drought on Quercus ilex radial growth and xylem anatomy. Trees Struct. Funct. 2004, 18, 83–92. [Google Scholar]
- Sánchez-Salguero, R.; Camarero, J.J.; Hevia, A.; Madrigal-González, J.; Linares, J.C.; Ballesteros-Canovas, J.A.; Sánchez-Miranda, A.; Alfaro-Sánchez, R.; Sangüesa-Barreda, S.; Galván, J.D.; et al. What drives growth of Scots pine in continental Mediterranean climates: Drought, low temperaturas or both? Agric. For. Meteorol. 2015, 206, 151–162. [Google Scholar] [CrossRef]
- Camarero, J.J.; Olano, J.M.; Parras, A. Plastic bimodal xylogenesis in conifers from continental Mediterranean climates. New Phytol. 2010, 185, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Peñuelas, J.; Filella, I.; Comas, P. Changed plant and animal life cycles from 1952 to 2000 in the Mediterranean region. Glob. Chang. Biol. 2002, 8, 531–544. [Google Scholar] [CrossRef]
- Gordo, O.; Sanz, J.J. Long-term temporal changes of plant phenology in the Western Mediterranean. Glob. Chang. Biol. 2009, 15, 1930–1948. [Google Scholar] [CrossRef]
- Fischer, R.; Barbosa, P.; Bastrup-Birk, A.; Becher, G.; Dobbertin, M.; Ferretti, M.; Flot, J.L.; Garcia-Fernandez, P.; Gerosa, G.; Gilbert, J.-M.; et al. The Conditions of Forests in Europe; 2004 Executive Report; United Nations Economic Commission for Europe (UNECE): Geneva, Switzerland, 2004; pp. 1–52. [Google Scholar]
- Solberg, S. Crown condition and growth relationships within stands of Picea abies. Scand. J. For. Res. 1999, 14, 320–327. [Google Scholar] [CrossRef]
- Mencuccini, M.; Grace, J. Climate influences the leaf area/sapwood area ratio in Scots pine. Tree Physiol. 1995, 15, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Sander, C.; Eckstein, D. Foliation of spruce in the Giant Mts. and its coherence with growth and climate over the last 100 years. Ann. For. Sci. 2001, 58, 155–164. [Google Scholar] [CrossRef]
- Michel, A.; Seidling, W.; Lorenz, M.; Becher, G. (Eds.) Forest Condition in Europe: 2013 Technical Report of ICP Forests; Report under the UNECE Convention on Long-Range Transboundary Air Pollution (CLRTAP); Thünen Working Paper 19; Johann Heinrich von Thünen-Institut: Eberswalde/Hamburg, Germany, 2014; p. 134.
- Shestakova, T.A.; Gutiérrez, E.; Kirdyanov, A.V.; Camarero, J.J.; Génova, M.; Knorre, A.A.; Linares, J.C.; Resco de Dios, V.; Sánchez-Salguero, R.; Voltas, J. Forests synchronize their growth in contrasting Eurasian regions in response to climate warming. Proc. Natl. Acad. Sci. USA 2016, 113, 662–667. [Google Scholar] [CrossRef] [PubMed]
- Hódar, J.A.; Castro, J.; Zamora, R. Pine processionary caterpillar Thaumetopoea pityocampa as a new threat for relict Mediterranean Scots pine forests under climatic warming. Biol. Conserv. 2003, 110, 123–129. [Google Scholar] [CrossRef]
- Sanz, M.J.; Calatayud, V.; Sánchez-Peña, G. Ozone concentrations measured by passive sampling at the intensive monitoring plots of South Western Europe. Environ. Pollut. 2007, 145, 620–628. [Google Scholar] [CrossRef] [PubMed]
- Molina, J.; Sánchez, G.; Prieto, M.; Soriano, A.; García, P.; Minaya, M.; de la Cruz, A.C. Caracterización de la Deposición Atmosférica en Varios Ecosistemas Forestales Españoles a Través de la Red CE de Nivel II, 6th ed.; Spanish Forest Congress: Vitoria-Gasteiz, Spain, 2013. [Google Scholar]
- Innes, J.L. Forest Health: Its Assessment and Status; Commonwealth Agricultural Bureau: Wallingford, CT, USA, 1993. [Google Scholar]
- Ferretti, M. Potential and limitations of visual indices of tree condition. Chemosphere 1998, 36, 1031–1036. [Google Scholar] [CrossRef]
- Johnson, J.; Jacob, M. Monitoring the effects of air pollution on forest condition in Europe: Is crown defoliation an adequate indicator? iForest 2010, 3, 86–88. [Google Scholar] [CrossRef]
- Cramer, H.H. On the predisposition to disorders of Middle European forests. Planzenshutz-Nachr. Bayer 1984, 2, 97–207. [Google Scholar]
- Phillips, O.L.; Baker, T.R.; Arroyo, L.; Higuchi, N.; Killeen, T.J.; Laurance, W.F.; Lewis, S.L.; Lloyd, J.; Malhi, Y.; Monteagudo, A.; et al. Pattern and process in Amazon tree turnover, 1976–2001. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2004, 359, 381–407. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.P.; Allen, C.D.; Macalady, A.K.; Griffin, D.; Woodhouse, C.A.; Meko, D.M.; Swetnam, T.W.; Rauscher, S.A.; Seager, R.; Grissino-Mayer, H.D.; et al. Temperature as a potent driver of regional forest drought stress and tree mortality. Nat. Clim. Chang. 2013, 3, 292–297. [Google Scholar] [CrossRef]
- Luo, Y.; Chen, H.Y.H. Climate change-associated tree mortality increases without decreasing water availability. Ecol. Lett. 2015, 18, 1207–1215. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tree Species or Group | Elevation (m) | Slope (°) | No. Plots (No. Trees) | Mean Annual Temperature (°C) | Total Annual Precipitation (mm) | DBH (cm) * |
|---|---|---|---|---|---|---|
| Pinus halepensis | 675 ± 334 | 24 ± 17 | 118 (3193) | 14.2 ± 1.7 | 502 ± 133 | 24.5 ± 10.4 |
| Pinus nigra | 1022 ± 316 | 23 ± 18 | 83 (1863) | 11.7 ± 1.4 | 610 ± 131 | 24.2 ± 9.9 |
| Pinus pinaster | 671 ± 395 | 16 ± 15 | 99 (3092) | 13.2 ± 1.6 | 840 ± 433 | 30.4 ± 11.4 |
| Pinus pinea | 574 ± 293 | 13 ± 11 | 34 (750) | 14.4 ± 2.2 | 551 ± 118 | 27.2 ± 8.9 |
| Pinus sylvestris | 1250 ± 324 | 31 ± 20 | 84 (2126) | 9.6 ± 1.9 | 825 ± 266 | 26.1 ± 10.2 |
| Quercus faginea | 873 ± 287 | 27 ± 19 | 56 (643) | 12.3 ± 1.9 | 610 ± 150 | 21.5 ± 11.6 |
| Quercus ilex | 722 ± 300 | 20 ± 18 | 213 (4465) | 13.8 ± 2.2 | 584 ± 129 | 26.2 ± 14.3 |
| Quercus pyrenaica | 978 ± 232 | 18 ± 17 | 55 (1366) | 11.1 ± 2.0 | 749 ± 194 | 21.2 ± 10.3 |
| Quercus robur | 455 ± 285 | 32 ± 26 | 36 (545) | 12.6 ± 1.5 | 1225 ± 243 | 31.2 ± 19.6 |
| Quercus suber | 425 ± 235 | 19 ± 15 | 42 (716) | 15.6 ± 1.3 | 661 ± 158 | 35.6 ± 19.5 |
| Oaks | 720 ± 325 | 23 ± 19 | 402 (7735) | 13.1 ± 1.8 | 765 ± 175 | 26.1 ± 14.9 |
| Pines | 837 ± 413 | 22 ± 18 | 418 (11024) | 12.6 ± 1.9 | 666 ± 216 | 26.4 ± 10.7 |
| Tree Species | Site (Code) | Latitude (N) | Longitude (W) | Elevation (m) | No. Trees (No. Cores) | Mean Tree-Ring Width (mm) | Defoliation (%) |
|---|---|---|---|---|---|---|---|
| P. pinaster | Valle de Cabra (VC) | 40°18′ | 0°47′ | 1165 | 15 (30) | 1.25 ± 0.60 | 11.8 ± 4.7 |
| P. sylvestris | Alcalá de la Selva (AL) | 40°21′ | 0°46′ | 1350 | 22 (44) | 0.65 ± 0.33 | 21.9 ± 13.9 |
| Q. ilex | Mora de Rubielos (MR) | 40°16′ | 0°48′ | 1050 | 15 (26) | 1.22 ± 0.63 | 15.0 ± 3.9 |
| Tree Species or Group | Maximum Defoliation (Year) | Mean ± SD Defoliation (%) | Coefficient of Variation | First-Order Autocorrelation | Trend, Kendall τ (p) |
|---|---|---|---|---|---|
| Pinus halepensis | 24.2 (2005) | 18.3 ± 4.4 | 0.24 | 0.74 | 0.61 (<0.001) |
| Pinus nigra | 21.0 (1996) | 17.7 ± 2.8 | 0.16 | 0.54 | 0.43 (0.005) |
| Pinus pinaster | 19.9 (2005) | 14.8 ± 3.3 | 0.22 | 0.86 | 0.73 (0.0001) |
| Pinus pinea | 28.6 (2010) | 17.9 ± 5.3 | 0.29 | 0.68 | 0.72 (0.0001) |
| Pinus sylvestris | 19.0 (2006) | 15.4 ± 3.2 | 0.21 | 0.78 | 0.49 (0.001) |
| Quercus faginea | 29.5 (1995) | 20.7 ± 4.9 | 0.24 | 0.65 | 0.46 (0.003) |
| Quercus ilex | 24.2 (1995) | 19.2 ± 4.1 | 0.21 | 0.75 | 0.47 (0.002) |
| Quercus pyrenaica | 22.1 (2006) | 17.6 ± 3.7 | 0.21 | 0.67 | 0.64 (<0.001) |
| Quercus robur | 23.0 (2004) | 18.4 ± 3.3 | 0.18 | 0.61 | 0.38 (0.012) |
| Quercus suber | 32.1 (1995) | 21.6 ± 6.0 | 0.28 | 0.70 | 0.45 (0.004) |
| Oaks | 24.7 (1995) | 19.5 ± 4.1 | 0.21 | 0.74 | 0.72 (0.0001) |
| Pines | 24.2 (2005) | 16.8 ± 3.5 | 0.21 | 0.78 | 0.72 (0.0001) |
| Tree Species or Group | P. sylvestris | P. nigra | P. pinaster | P. pinea | P. halepensis | Q. robur | Q. pyrenaica | Q. faginea | Q. ilex | Q. suber | Pines |
|---|---|---|---|---|---|---|---|---|---|---|---|
| P. sylvestris | |||||||||||
| P. nigra | 0.65 | ||||||||||
| P. pinaster | 0.87 | 0.77 | |||||||||
| P. pinea | 0.82 | 0.64 | 0.92 | ||||||||
| P. halepensis | 0.76 | 0.72 | 0.90 | 0.89 | |||||||
| Q. robur | 0.67 | 0.67 | 0.62 | 0.56 | 0.48 | ||||||
| Q. pyrenaica | 0.81 | 0.56 | 0.87 | 0.59 | 0.90 | 0.40 | |||||
| Q. faginea | 0.87 | 0.61 | 0.81 | 0.80 | 0.83 | 0.43 | 0.82 | ||||
| Q. ilex | 0.68 | 0.70 | 0.77 | 0.76 | 0.92 | 0.45 | 0.78 | 0.80 | |||
| Q. suber | 0.75 | 0.58 | 0.78 | 0.77 | 0.88 | 0.31 | 0.78 | 0.81 | 0.83 | ||
| Pines | 0.76 | 0.72 | 0.90 | 0.89 | 0.95 | 0.48 | 0.89 | 0.86 | 0.92 | 0.88 | |
| Oaks | 0.85 | 0.69 | 0.85 | 0.84 | 0.94 | 0.61 | 0.86 | 0.97 | 0.90 | 0.84 | 0.94 |
| Tree Species or Group | Selected Model of Tree Defoliation | t Ratio of First-Selected Variable (p) | R2 Adj |
|---|---|---|---|
| Pinus halepensis | −53.60 + 3.88 TApr + 1.50 TJun − 0.25 PJul | TApr, 5.67 (<0.0001) | 0.64 |
| Pinus nigra | −19.95 + 2.98 TApr + 0.65 TJun − 0.03 PNovt−1 | TApr, 5.60 (<0.0001) | 0.61 |
| Pinus pinaster | −42.18 + 2.66 TApr + 1.43 TJun − 0.19 PJul | TApr, 5.45 (<0.0001) | 0.67 |
| Pinus pinea | −56.31 + 2.74 TApr + 1.22 TJun − 0.37 PJul | TApr, 5.46 (<0.0001) | 0.56 |
| Pinus sylvestris | −29.59 + 1.98 TApr + 1.07 TJun | TApr, 3.50 (0.003) | 0.42 |
| Quercus faginea | −43.51 + 2.71 TApr + 1.31 TJun | TApr, 4.82 (0.0001) | 0.40 |
| Quercus ilex | −27.14 + 3.90 TApr + 1.69 TJun − 1.25 TSept−1 | TApr, 6.02 (<0.0001) | 0.71 |
| Quercus pyrenaica | −0.31 + 2.46 TApr + 1.46 TMar − 0.14 PAug | TApr, 4.52 (0.0003) | 0.52 |
| Quercus robur | 7.45 + 1.54 TApr + 1.21 TJun − 1.39 TJul | TApr, 2.43 (0.0256) | 0.35 |
| Quercus suber | −23.73 + 2.91 TApr + 2.37 TJun − 2.51 TSept−1 | TApr, 5.42 (<0.0001) | 0.63 |
| Oaks | −21.97 + 3.07 TApr + 1.67 TJun − 0.04 PNovt−1 | TApr, 5.70 (<0.0001) | 0.69 |
| Pines | −40.82 + 3.23 TApr + 1.14 TJun − 0.21 PJul | TApr, 5.92 (<0.0001) | 0.66 |
| Tree Species or Group | Selected Model of Tree Defoliation | t Ratio of First-Selected Variable (p) | R2Adj |
|---|---|---|---|
| Pinus halepensis | 14.62 + 15.46 AMOApr | AMOApr, 5.08 (<0.0001) | 0.54 |
| Pinus nigra | 17.05 + 5.22 AMOJan − 1.11 SOIMay − 0.73 NAOJan | AMOJan, 5.22 (0.0007) | 0.74 |
| Pinus pinaster | 13.63 + 7.94 AMOJan + 1.40 AMOSept−1 − 0.57 NAOFeb | AMOJan, 4.84 (<0.0001) | 0.82 |
| Pinus pinea | 15.42 + 14.70 AMOApr − 0.84 NAOFeb | AMOApr, 4.03 (0.0007) | 0.61 |
| Pinus sylvestris | 14.34 + 7.32 AMOJan − 0.94 SOIMay − 0.47 NAOFeb | AMOJan, 4.37 (0.0004) | 0.67 |
| Quercus faginea | 17.82 + 13.22 AMOJan − 0.58 SOIMay | AMOJan, 5.04 (0.0005) | 0.64 |
| Quercus ilex | 15.38 + 14.62 AMOApr − 0.95 SOIMay | AMOApr, 5.77 (<0.0001) | 0.65 |
| Quercus pyrenaica | 16.55 + 8.04 AMOJan − 0.99 SOIMay − 0.65 NAOFeb | AMOJan, 4.22 (0.0005) | 0.68 |
| Quercus robur | 16.85 + 9.05 AMOJan − 1.16 SOIApr − 0.86 NAOMar | AMOJan, 5.93 (<0.0001) | 0.75 |
| Quercus suber | 17.33 + 17.50 AMOApr − 0.67 NAOApr | AMOApr, 3.52 (0.0023) | 0.40 |
| Oaks | 15.79 + 13.68 AMOApr − 1.30 SOIMay | AMOApr, 5.58 (<0.0001) | 0.66 |
| Pines | 16.28 + 8.18 AMOJan − 0.74 NAOFeb − 0.64 NAOApr | AMOJan, 4.76 (0.0002) | 0.74 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Salguero, R.; Camarero, J.J.; Grau, J.M.; De la Cruz, A.C.; Gil, P.M.; Minaya, M.; Fernández-Cancio, Á. Analysing Atmospheric Processes and Climatic Drivers of Tree Defoliation to Determine Forest Vulnerability to Climate Warming. Forests 2017, 8, 13. https://doi.org/10.3390/f8010013
Sánchez-Salguero R, Camarero JJ, Grau JM, De la Cruz AC, Gil PM, Minaya M, Fernández-Cancio Á. Analysing Atmospheric Processes and Climatic Drivers of Tree Defoliation to Determine Forest Vulnerability to Climate Warming. Forests. 2017; 8(1):13. https://doi.org/10.3390/f8010013
Chicago/Turabian StyleSánchez-Salguero, Raúl, J. Julio Camarero, José M. Grau, Ana C. De la Cruz, Paula M. Gil, Mayte Minaya, and Ángel Fernández-Cancio. 2017. "Analysing Atmospheric Processes and Climatic Drivers of Tree Defoliation to Determine Forest Vulnerability to Climate Warming" Forests 8, no. 1: 13. https://doi.org/10.3390/f8010013
APA StyleSánchez-Salguero, R., Camarero, J. J., Grau, J. M., De la Cruz, A. C., Gil, P. M., Minaya, M., & Fernández-Cancio, Á. (2017). Analysing Atmospheric Processes and Climatic Drivers of Tree Defoliation to Determine Forest Vulnerability to Climate Warming. Forests, 8(1), 13. https://doi.org/10.3390/f8010013