1. Introduction
Glossy buckthorn (
Frangula alnus Mill., hereafter known as ‘buckthorn’), a tall shrub or small tree native to Europe, is one of over 20 non-native, woody plant species reported as invasive in forests of eastern North America [
1]. Moderately shade tolerant [
2,
3,
4], buckthorn can form a dense shrub layer that dominates the forest understory especially under canopies of eastern white pine (
Pinus strobus L.) [
2,
5,
6,
7,
8]. Buckthorn abundance in white pine stands is enhanced by logging, with resulting densities sometimes exceeding 60,000 stems·ha
−1 [
4]. Consequently, buckthorn can potentially inhibit native tree regeneration before logging occurs and especially so afterward.
As is true for most eastern North American woody invasive plants, relatively little is known about the competitive effects of buckthorn on other forest species. Correlative evidence suggests that buckthorn is associated with lower herb cover [
2,
9] and lower density, growth, and survival of tree seedlings and saplings [
2,
7]. For example, Fagan and Peart [
7] found that seedlings of economically important eastern white pine growing under buckthorn had lower growth and survival than those away from buckthorn. Despite the strong association of buckthorn with pine-dominated forests, only two studies [
8,
10] have reported the effects of experimental buckthorn removal on eastern white pine regeneration and, in both of these, white pine was one of many understory species examined. Neither found significant buckthorn effects, but pine abundance and samples sizes were low.
Buckthorn’s ability to alter forest structure has prompted managers to seek control methods. Potential controls include biological agents, mechanical methods, and herbicides. Despite a major effort, no promising buckthorn-specific biocontrol agent has been found [
11]. Thus, buckthorn control is presently limited primarily to chemical and mechanical means. One or two applications of herbicide cause buckthorn mortality [
12,
13], although time of year [
14] and soil moisture level [
15] may influence herbicide effectiveness.
Few peer-reviewed publications discuss mechanical control of buckthorn and none employ the scheme of timing and repetition we describe in this paper. Mechanical uprooting is possible and has been used to control other invasive shrubs [
16,
17] but it is labor and time intensive, difficult to apply to large areas, and often requires follow-up control [
18]. Single applications of stem girdling [
19] and flaming [
20,
21,
22] are reported to be ineffective for buckthorn, but have been tested under a limited range of conditions. Repeated cutting, mowing, or burning, however, seem to provide buckthorn control [
23,
24]. Indeed, repeated application of mechanical techniques, such as flaming, have been effective in controlling other invasive shrub species [
25,
26].
Effectiveness of mechanical control of buckthorn is limited by the production of butt or stump sprouts immediately after cutting [
12,
19,
27]. To minimize such sprouting, some authors emphasize the timing of control. Kays and Canham [
28] and Love and Anderson [
16] advocate cutting stems of shrubs when root reserves are low, thus reducing sprouting and maximizing the chance of plant death. Cutting buckthorn stems not only promotes sprouting, but is often followed by germination of buried dormant seed [
29].
We sought to determine the effect of mechanical removal of buckthorn on the abundance and growth of eastern white pine seedlings. Moreover, we sought to quantify these effects in association with a logging event. Although an obvious approach to minimizing buckthorn effects on pine regeneration would be to treat buckthorn at or just after the time of logging, the novel approach we report here involved treatment beginning two years prior to logging. We expected that repeated cutting or flaming (heat-killing) of buckthorn stems while they remained beneath a forest canopy in a competitive environment would more effectively reduce buckthorn and liberate pine seedlings than in the light- and nutrient-rich post-logging environment.
Our first objective was to test the hypotheses that destruction of buckthorn stems by cutting, or cutting and heat killing, prior to logging would (1) reduce post-logging density and height of buckthorn and (2) increase post-logging density and size of eastern white pine seedlings. After logging of the forest canopy, we found that buckthorn recovered to some degree within most of our treated areas, and sometimes formed dense, multi-individual patches with continuous cover of foliage. We refer to such patches as ‘buckthorn thickets’. Our second objective was to test the hypothesis that white pine seedlings in such thickets had reduced height, biomass, and growth relative to seedlings in non-thicket areas.
2. Materials and Methods
2.1. Study Area
The MacDonald Lot, Durham, New Hampshire, USA, is a 31.9 ha parcel owned and managed by the University of New Hampshire. In Durham, the average annual precipitation of 1169 mm, 10% of which occurs as snow, is evenly distributed over the year and mean annual temperature is 8.9 °C [
30]. In the area that we studied, soils were Buxton silt loam and Suffield silt loam, with limited Hollis–Charlton very rocky fine sandy loams [
31]. White-tailed deer (
Odocoileus virginianus) are abundant at this site but rarely browse either eastern white pine or buckthorn. Windstorms are occasional, with the February 2010 storm causing some windthrow and windsnap of pines on this property.
A 2006 inventory of the MacDonald Lot identified several stand types, one of which was old-field eastern white pine approximately 60 years old. Typical of old-field pine, many of these trees had either multiple tops, severe deformity in the first log, or unusually large lower branches. In such stands, a reasonable silvicultural approach is to clearcut the area with the intent of regenerating a higher quality pine stand. Although some white pine advanced regeneration was present on site, the understory was dominated by buckthorn (density given below). Clearcutting the stand without first treating the understory would likely have resulted in an increase of buckthorn [
4].
2.2. Study Design
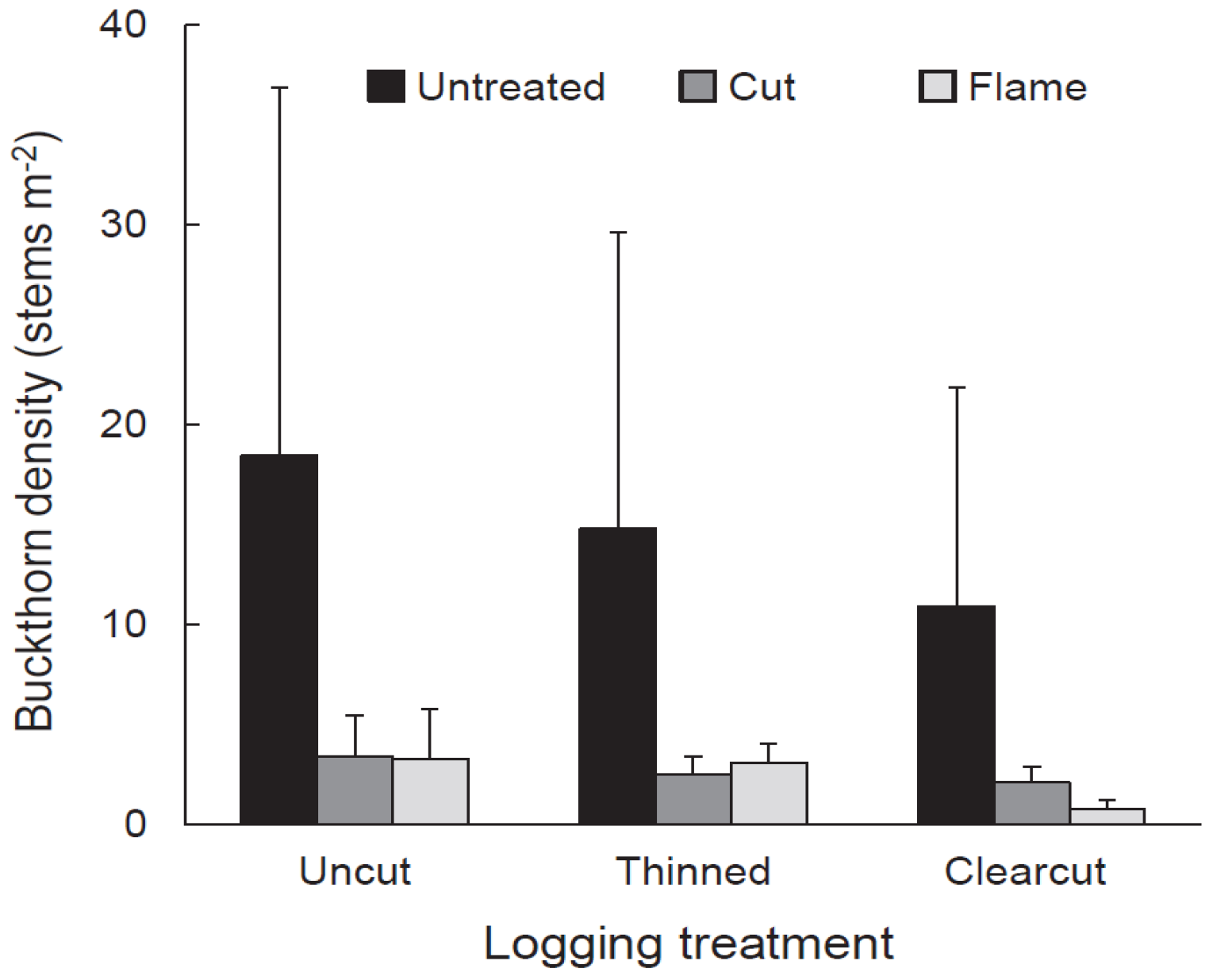
We selected nine 0.4 ha plots of forest with eastern white pine canopies and understories dominated by buckthorn. Three plots were randomly chosen to be clearcut, three to be thinned, and three to serve as uncut controls. Logging treatments were applied in the winter of 2009–2010. Each 0.4 ha plot was divided into three 0.13 ha sub-units (“subplots”), each of which received a different buckthorn treatment in 2008–2009, the two years before logging. The buckthorn treatments were (1) manually cutting buckthorn stems at the base (‘cut’ treatment), (2) initial cutting of stems followed by flaming of stem bases after initial cutting (‘flame’ treatment), and (3) untreated. Pre-treatment density of buckthorn, estimated using three 4 m2 circular quadrats per subplot, averaged 7.00 stems m−2 (s.d. = 4.07; n = 27; observations were means of three 4 m2 quadrats).
2.3. Treatments
Each of the understory buckthorn treatments was applied five times over two years, twice in 2008 (June and September) and three times in 2009 (June, July, September). The strategy was to destroy stems immediately after bud break in June when root reserves were low [
16,
28], and again late in the season to minimize overwintering reserves. Based on 2008 observations indicating greater than expected sprout growth by mid-summer, in 2009 a third application was added in mid-July. The first of the five applications, in June 2008, involved manual cutting of stems regardless of whether subplots were assigned for ‘cut’ or ‘flame’ treatments. This initial application consisted of, first, using Stihl FS 110R line trimmers (Stihl Inc., Virginia Beach, VA, USA) to remove the smallest stems (<3.2 mm basal diameter). Second, Stihl FS 450 brush saws and MS 361 chainsaws were used to cut larger stems. Large material, if cut first, would have hidden seedlings and interfered with the cutting of small stems using line trimmers. Larger material was cut into smaller pieces so as to make follow-up treatments as easy as possible. In the four subsequent applications (September 2008 and all three 2009 applications), the targets—newly emerging seedlings and stump sprouts (from surviving individuals)—were reduced with just two tools, Weed Dragon VT 2-23SVC flame guns (Flame Engineering, Inc., LaCrosse, KS, USA) for the flame treatments and Stihl FS 110R line trimmers for cut treatments. In all treated subplots, cutting and flaming was done as close to the buckthorn stem base as physically possible. All cut materials were left in place.
Tree harvest was conducted in the winter of 2009–2010 by a whole-tree harvesting contractor. All marked trees greater than 10.2 cm diameter at breast height were harvested. In clearcuts, all canopy trees were harvested. In thinned plots, less valuable canopy trees (deformed, injured) were harvested along with more valuable trees to create a uniform canopy cover. Thinning was intended to reduce standing live basal area (BA) to the same value in each plot, ca. 30 m2·ha−1, which required an 18%–29% reduction in BA from pre-treatment levels. Trees were taken off-site whole, using a grapple skidder.
2.4. Intensive Quadrat Sampling: 2012
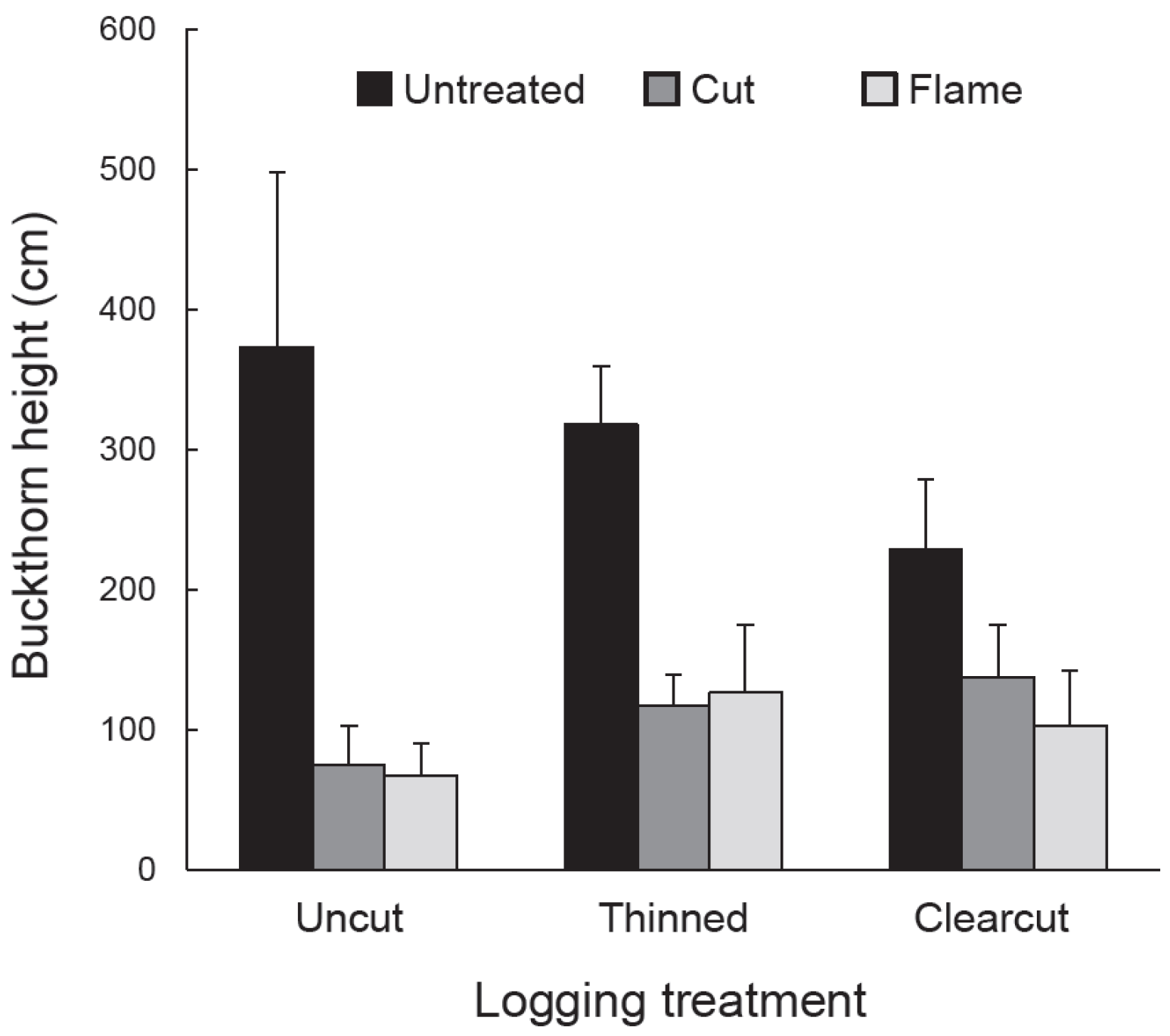
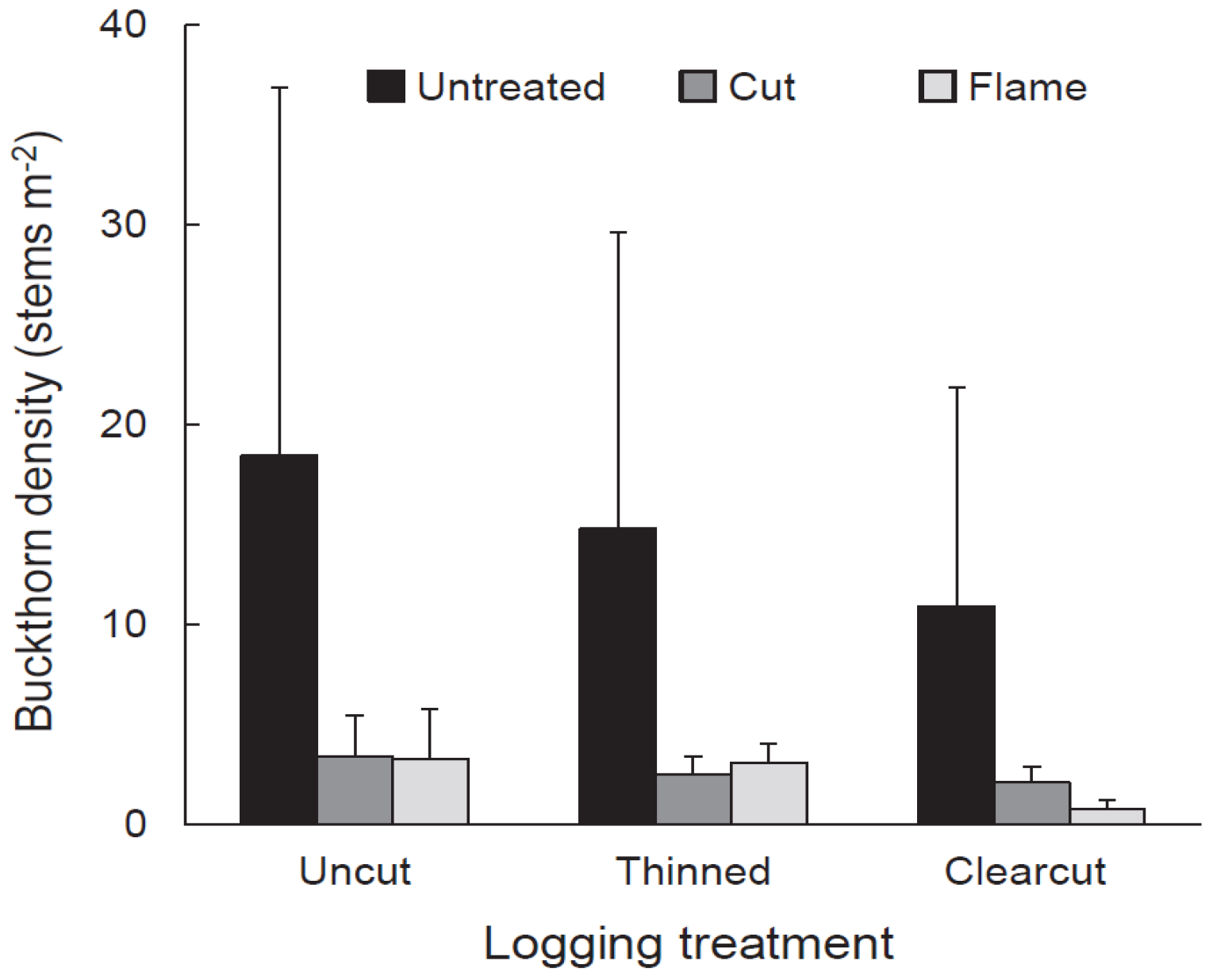
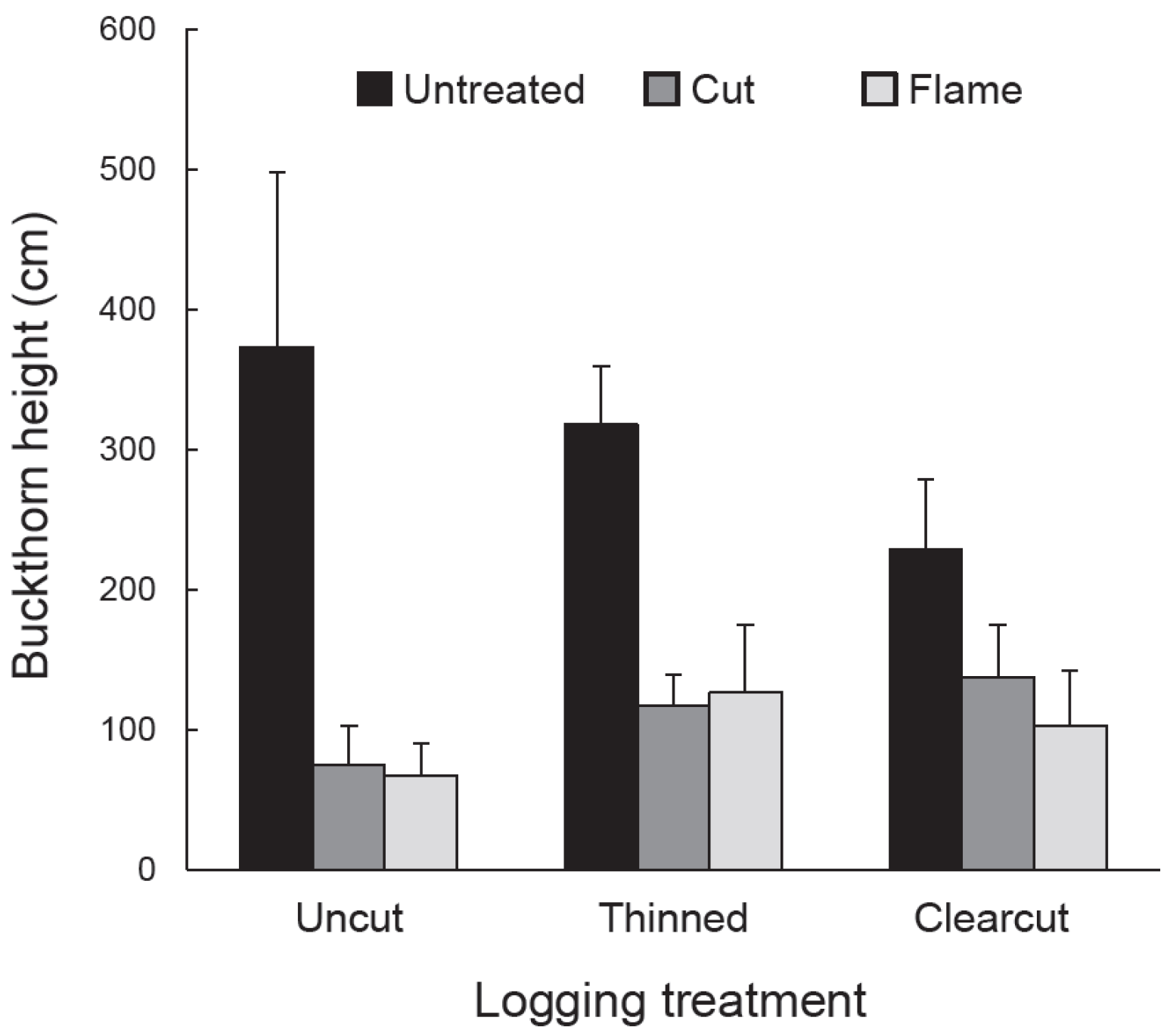
Vegetation response to logging and buckthorn removal was assessed using quadrat sampling in July–August 2012, the third growing season after logging. In each subplot, we randomly placed six, 2 m × 4 m (8 m2) quadrats and in each quadrat we tallied the number of buckthorn stems and the number of eastern white pine seedlings from the 2010 cohort. We also measured the height of the tallest eastern white pine seedling (2010 cohort) and tallest buckthorn stem within each quadrat. Density and height data from the six quadrats in each subplot were averaged to obtain single values to be used in statistical analysis. Independent variables were buckthorn density, height of tallest buckthorn, pine seedling density, and height of tallest pine seedling. We used a split-plot ANOVA in the statistical analysis program JMP Pro 12 (SAS Inc., Cary, NC, USA) to determine the response of each independent variable to the dependent variables: logging treatment, buckthorn treatment, and their interaction. All independent variables were loge-transformed to homogenize variances. Where appropriate, treatment and cell means were compared using Tukey HSD (honestly significant difference) in JMP Pro 12.
2.5. Buckthorn Origin
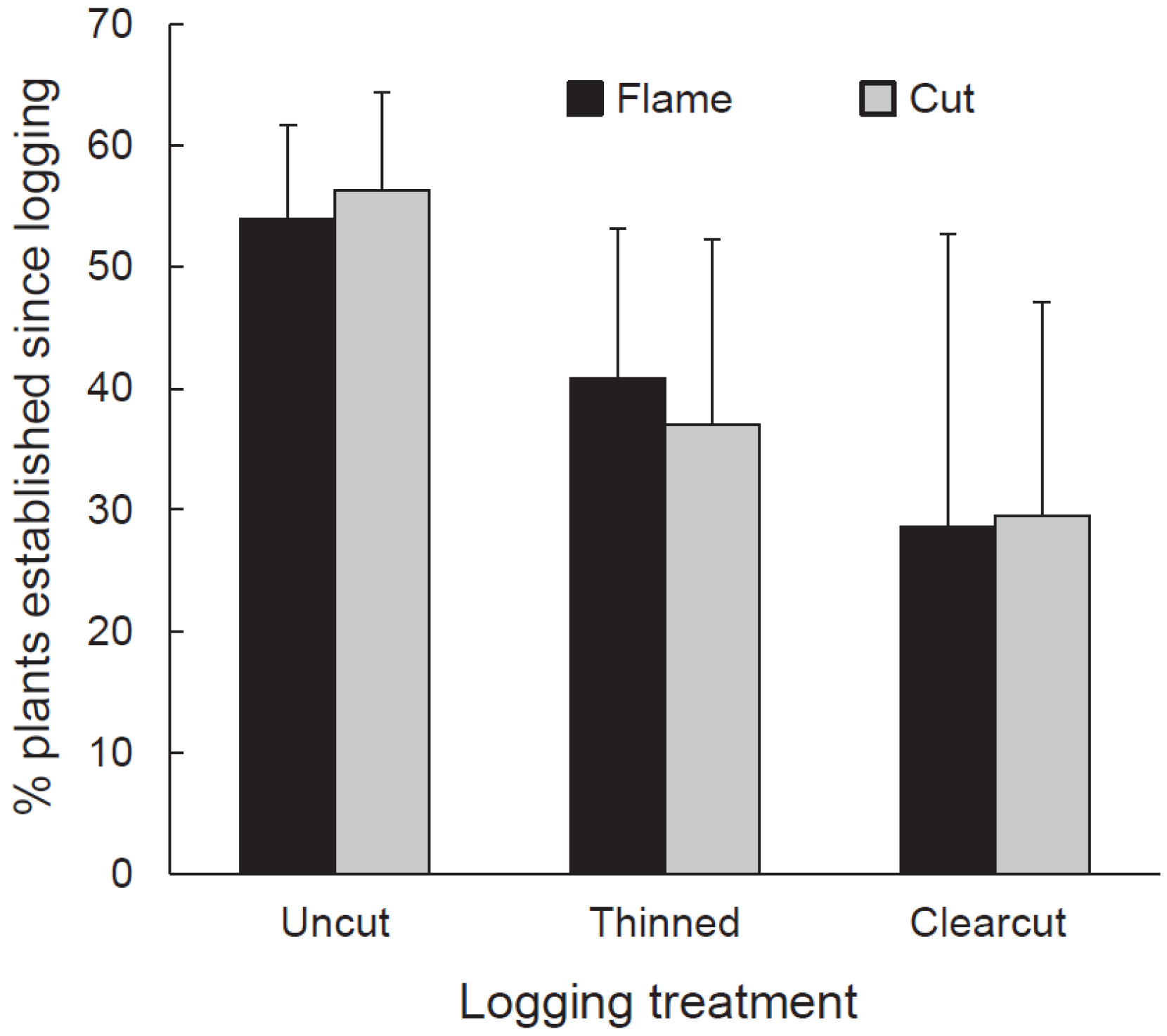
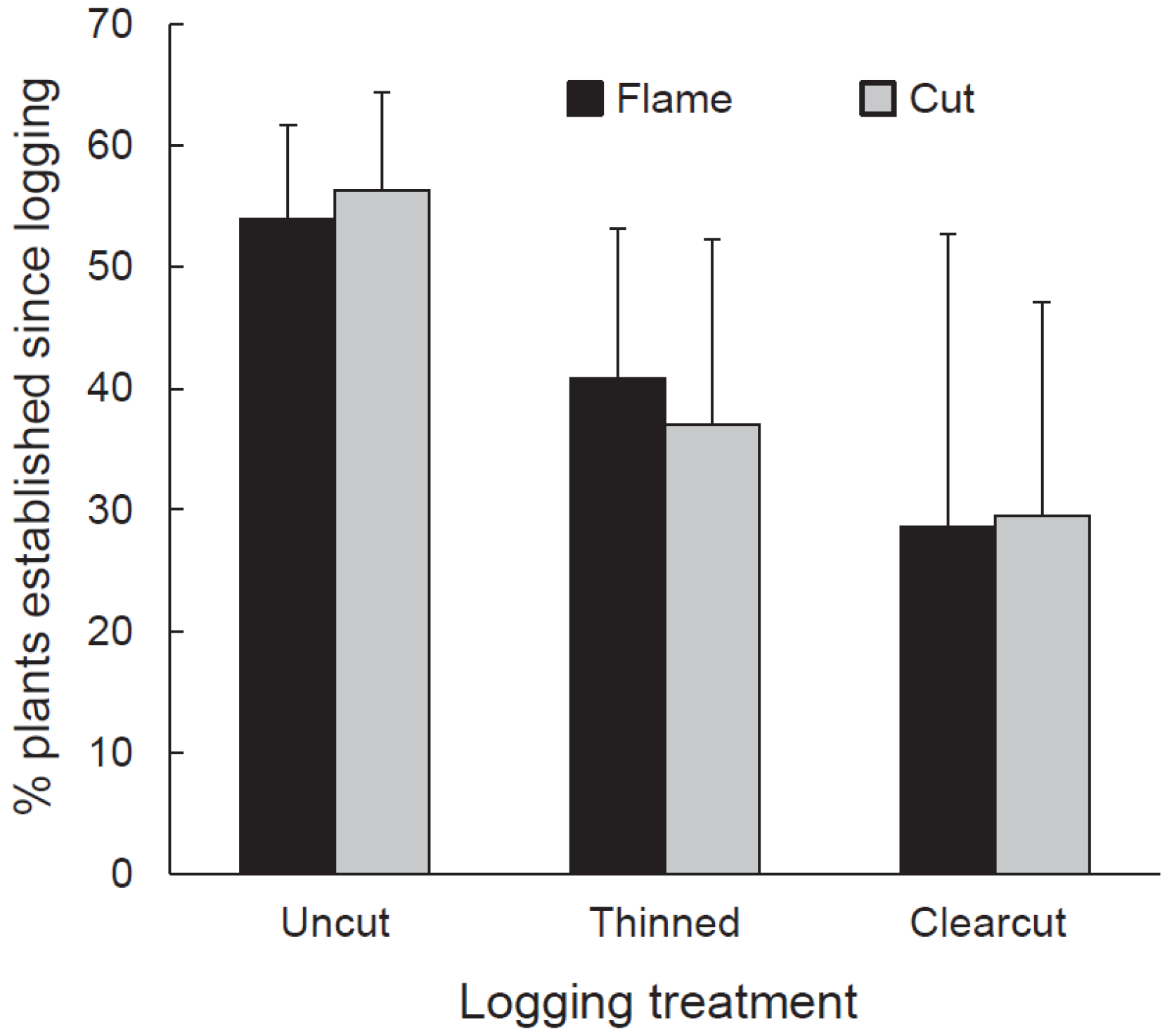
Our 2012 sampling revealed that buckthorn had recovered on some subplots where it had been treated with cutting or cutting and flaming. Between June 4 and 11, 2013, we sampled living buckthorn plants in the treated subplots (in which buckthorn was cut or flamed). We wanted to determine if these buckthorn individuals had established before or after the logging event of winter 2009–2010. In each subplot, we excavated 60 randomly selected buckthorn individuals. We counted wood rings at the stem base to estimate age and then examined each plant for evidence of previous cutting or burning. Plants establishing prior to 2010 would have survived at least some of the cutting or flaming treatments and so should have exhibited some evidence of stem damage. Based on plant age and evidence of cutting or flaming, we classified our selected buckthorn plants into those having established prior to and after logging treatments were imposed. The cut and cut-and-flamed subplots in one clearcut had insufficient buckthorn for sampling, so the number of subplots sampled was 16 (i.e., cut and cut-and-flamed subplots in all three thinned treatments, all three uncut treatments, and in two of the three clearcuts). For statistical analysis, the independent variable, proportion of buckthorn stems established after logging, was arcsine square root transformed and then tested against the dependent variables of logging, buckthorn removal, and their interaction using split-plot analysis of variance (ANOVA) in JMP Pro 12.
2.6. Intensive Quadrat Sampling: 2014
The 2012 quadrat study suggested that buckthorn regrowth in some of the treated subplots was sufficient to inhibit growth of regenerating white pine (see Results). Consequently, in July and August, 2014, we examined white pine seedling density and height in relation to buckthorn abundance in the two clearcut plots and the two thinned plots that had abundant recovering buckthorn. In the cut and cut/flamed subplots of these four plots, we delineated two habitats: buckthorn ‘thickets,’ characterized by buckthorn plants with continuous foliar cover and with foliage from adjacent buckthorn individuals separated by no more than 20 cm (horizontal distance), and ‘non-thicket areas’, characterized by non-continuous buckthorn cover and with foliage from adjacent buckthorn individuals separated by more than 20 cm. In each subplot, we used a compass and Distance Measuring Equipment (DME, Haglӧf Sweden, Längsele, Sweden) to estimate the area occupied by thicket and non-thicket. To estimate buckthorn and pine density and height in thicket and non-thicket areas of each subplot, we randomly located twenty 1 m × 1 m quadrats in buckthorn thickets and 20 more in non-thicket areas. In addition, we deployed 20 such quadrats to estimate buckthorn and pine performance in the untreated buckthorn subplots of each plot, which were generally dominated by buckthorn thickets.
In each quadrat, we tallied the number of buckthorn stems arising from the ground surface by height class (<1 m and ≥1 m tall). We also tallied the number of eastern white pine seedlings from the 2010 cohort, per quadrat, and determined the height of the tallest pine seedling per quadrat. These tallest pine seedlings were cut at the base and dried in an oven at 90 °C to a constant weight and then weighed to determine aboveground biomass. Maximum height of the buckthorn stems above each plot was estimated to the nearest 0.5 m.
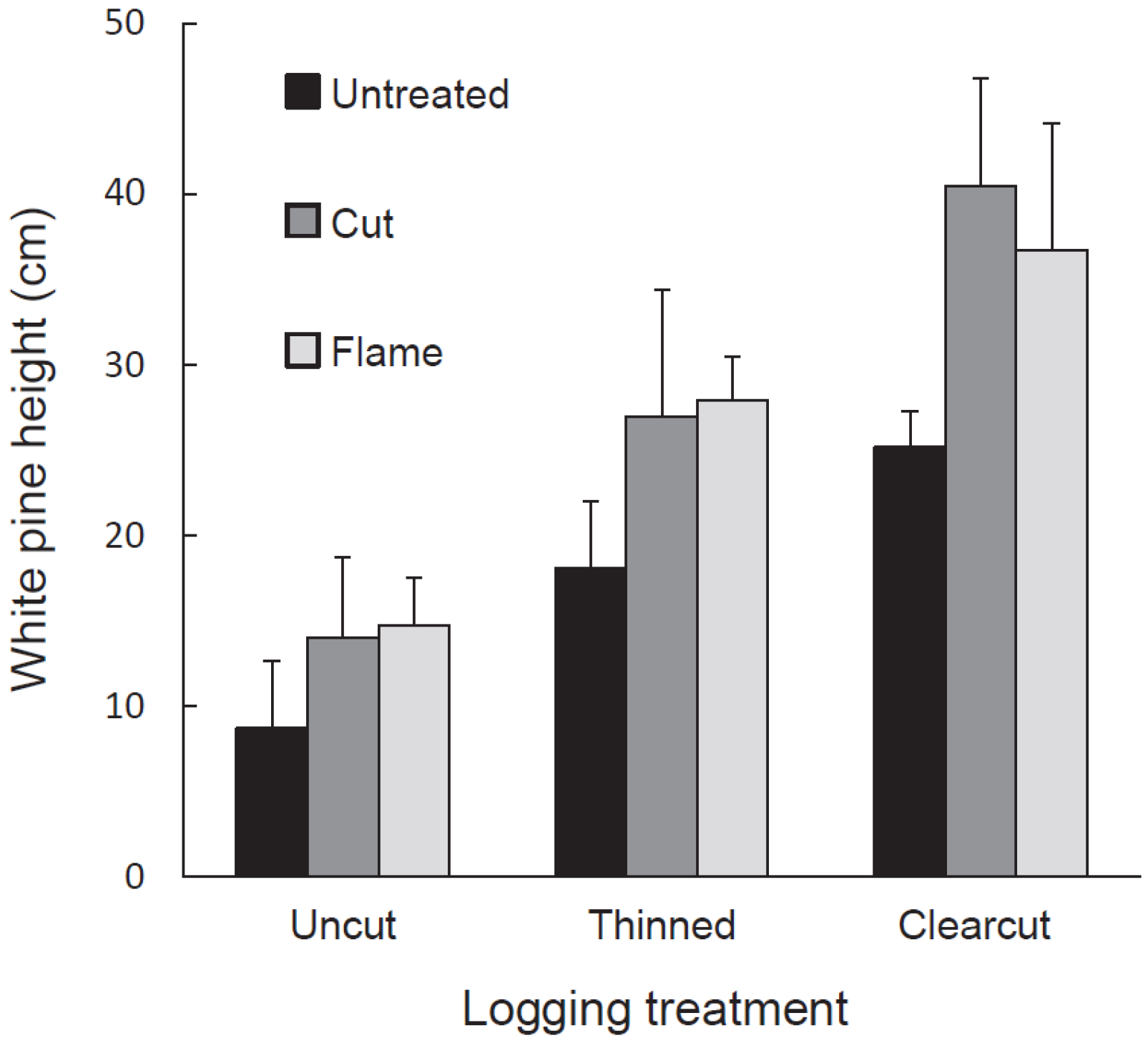
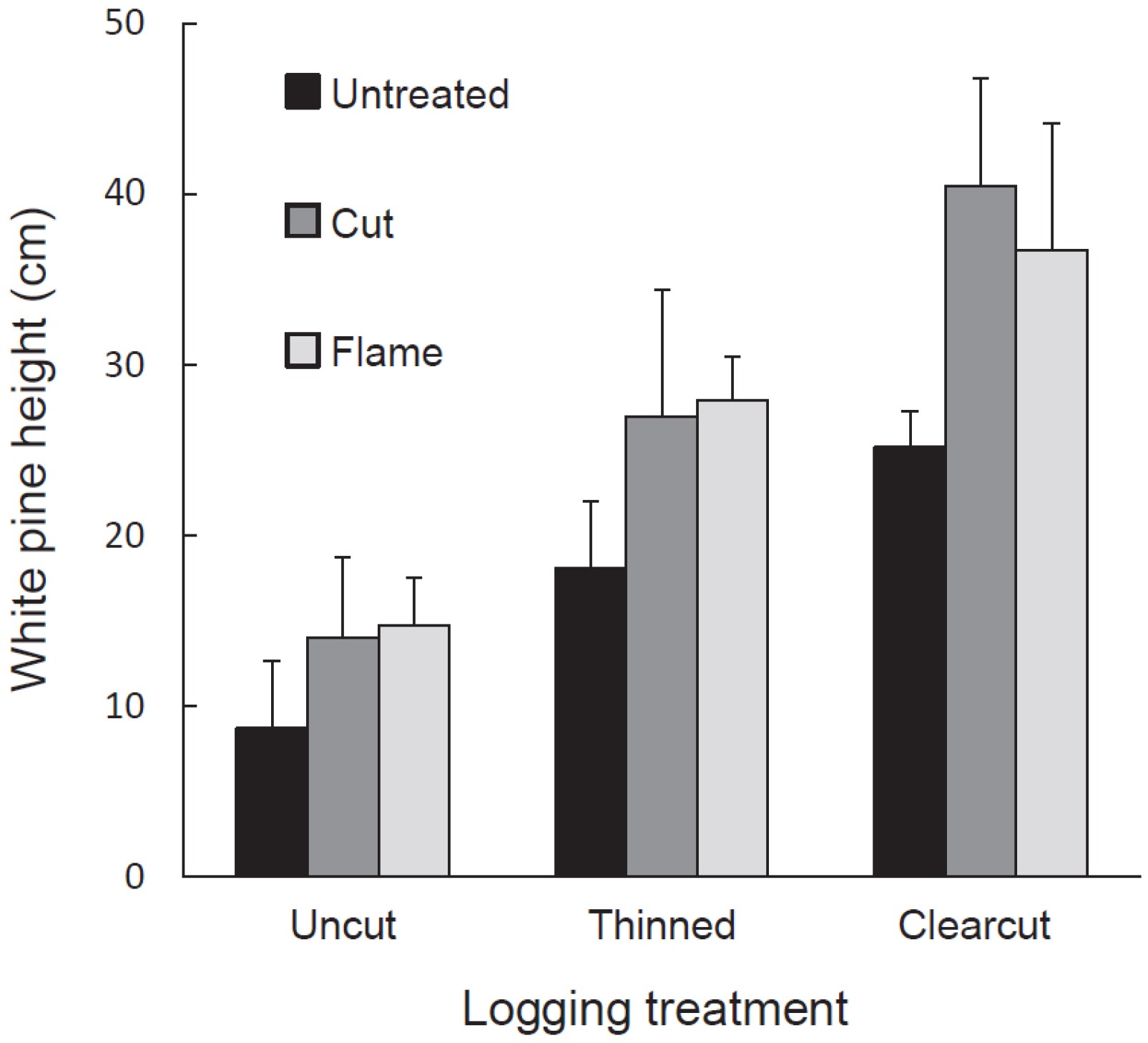
Pine density and, for the tallest white pine per plot, stem height, height growth in 2014, and aboveground biomass were designated as independent variables. We used a split-plot ANOVA in JMP Pro 12 to determine the response of each independent variable to the dependent variables: logging treatment, buckthorn treatment, and thicket presence. All independent variables were loge-transformed to homogenize variances. Treatment and cell means were compared using Tukey HSD in JMP Pro 12.
4. Discussion
4.1. Response of Pine to Buckthorn Removal
As expected given our treatments and study site, glossy buckthorn inhibited eastern white pine seedlings. Post-logging density and especially height of pine seedlings were greater in areas where buckthorn stems had been killed by cutting or burning than in untreated controls. Where recovering buckthorn formed thickets, pine seedlings showed lower annual increase in height and lower aboveground biomass compared to non-thicket areas. Our results thus suggest that pre-logging buckthorn reduction can be effective in releasing an economically important tree species.
Our findings are somewhat consistent with those of Fagan and Peart [
7] who found that, in canopy gaps, white pine seedling height growth over one growing season was lower in the presence of buckthorn. Under the closed white pine canopy, however, they found that pine seedling growth did not vary with buckthorn presence. Pine seedling survival in their study showed the reverse pattern, declining under buckthorn only when the pine canopy was intact. Our results differ only in that pine seedling growth and density were higher without buckthorn in both canopy openings and under a closed canopy. Differences in the results of these studies may be due to differences in time span (one vs. five growing seasons) or approach (e.g., correlative vs. experimental). Our results are also consistent with reports showing negative response of various other tree species to buckthorn presence [
2,
3,
8].
4.2. Buckthorn Recovery and Its Consequences for Pine
Our experimental buckthorn treatments neither completely eliminated buckthorn nor prevented it from re-establishing. In the fourth growing season after logging (2013), buckthorn was present in all cut and flamed subplots and, regardless of buckthorn treatment or logging regime, populations were roughly an even mix of plants that established from seed since logging and older plants that survived cutting or flaming and subsequent logging. Despite buckthorn persistence and recovery, buckthorn densities in cut and flamed areas in the fifth growing seasons after application of treatments were generally lower than untreated controls. Thus, treatment effects were not ephemeral.
Buckthorn influence was reduced sufficiently by cutting and flaming so that white pine seedlings were released, a response consistent with an extensive literature indicating that white pine grows better with little competition, e.g., [
32,
33]. The response of pine seedling density in 2012 to buckthorn removal, however, suggested a complex relationship to logging, with greatest pine response to buckthorn removal occurring in stands that were not logged. The less dramatic response of pine to buckthorn removal in logged stands may have resulted from damage to buckthorn or to potential pine establishment sites from slash deposition and skidder effects.
Although buckthorn removal resulted in taller white pine seedlings three years after logging, the tallest recovering buckthorn (per 8 m2 quadrat) on these same subplots was already twice the height, on average (>100 cm), of the tallest pine. Given this relationship, one could argue that pine release might be temporary, with pine becoming suppressed by the taller buckthorn. However, in buckthorn removal areas, buckthorn density was generally low, with patches of continuous buckthorn cover (thickets) 33% or less. Indeed, over most of the treated plots, buckthorn remained sparse even in the fifth year after logging.
Our 2014 data also suggest that the buckthorn thickets arising anew on buckthorn removal subplots affected pine differently than the thickets in untreated areas. Thickets on buckthorn removal plots seemed to be less resistant to pine seedling success than were the older, established thickets on the untreated subplots. Although white pine seedling density was similar in both thicket types, white pine biomass and annual increase in height were significantly greater in thickets on treated subplots than in those on untreated ones. New thickets on treated subplots may have allowed more pine growth for two reasons. First, in the untreated subplots, pine had to establish and grow under pre-existing buckthorn plants while, in treated areas, pine and buckthorn both established at the same time and the pine may thus have experienced less competition. Second, thickets in untreated subplots were large and continuous, whereas those in treated plots occurred in small patches whose edges received peripheral light.
There is little information available on the quantitative relationship between buckthorn density and the intensity of competition with white pine seedlings or those of other tree species. Based on buckthorn removal experiments conducted elsewhere in New Hampshire, Koning and Singleton [
8] found that buckthorn effects on understory woody plant growth were not detectable until buckthorn density exceeded 1.5–8.25 stems m
−2. Within buckthorn thickets in our study, stem densities averaged 15–16 m
−2 and so, assuming that the Koning and Singleton data apply, we might expect competitive effects on pine in these habitats. Outside of thickets, however, buckthorn density was ca. 3.6 stems m
−2, within the Koning and Singleton limit of detectable effects. Given that buckthorn thickets were limited in extent, as noted above, white pine has been able to grow with little competition from buckthorn in well over half the area treated.
Our experiment revealed strong treatment effects on both buckthorn and white pine density and growth despite low replication and high residual variation. Within-treatment (residual) variation may have been due to local heterogeneity in soils, slash deposition, and logging traffic. Although the canopy gaps created by a February, 2010, windstorm likely added to this variation, the large herd of white-tailed deer on site seemed to avoid browsing both buckthorn and pine.
4.3. Potential for Buckthorn Control
The two methods we used for killing buckthorn stems, (1) repeated manual cutting at the stem base and (2) a single manual cut followed by repeated flaming (heat) treatments of the stem, were similarly successful, not only in reducing buckthorn, but also in releasing eastern white pine seedlings. However, our second method was initiated with one round of stem cutting because using flame guns to kill the large buckthorn stems we encountered initially would have been prohibitive. Working with Japanese barberry, Ward et al. [
26], showed that the labor cost of initial stem destruction was five times greater for flaming than for cutting with a brush saw. Moreover, cutting and flaming differ greatly in utility, as flaming is limited to field conditions in which there is no chance of initiating wildfire whereas cutting is not so constrained.
The reduction in buckthorn density and height caused by our methods is difficult to compare with the rather limited literature available on buckthorn control. Although one or two mechanical treatments do not reduce buckthorn [
19,
20,
21], our results concur with reports that repeated cutting, mowing, or burning provide some level of buckthorn control [
24,
25,
26]. Herbicides (e.g., glyphosate) have been successful with buckthorn [
12,
13] and offer an alternative that will likely have lower costs than mechanical methods [
17,
26].
Ultimately, whether or not one chooses to manage buckthorn—and what control method one chooses should they decide to manage—depends on the (1) values identified by the landowner; (2) effectiveness of the method in realizing those values; and (3) cost of the method. Based on current costs and the return on eastern white pine timber in today’s market, it is questionable whether mechanical control would be cost effective if only timber values were considered. Inclusion of values associated with wildlife, biodiversity, and aesthetics, however, may influence a landowner’s perspective.
4.4. Future Research
Our study was limited to one forest type and one site with particular soil characteristics and climate. To assess the generality of our findings, the study should be repeated in other forest types and at sites with different limiting factors. There are no long-term studies of glossy buckthorn effects on pine and our report provides a limited time frame (five years) for evaluating plant responses to buckthorn and logging treatments. Extending this window will allow more definitive assessment of impacts and we plan to monitor our plots for several more years. Only widely replicated, long-term studies of invasive plant impact and control can inform development of best management practices required by land managers and, increasingly, by government, e.g., [
34]. Future research on buckthorn should also quantify the costs of control methods.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}