
Effects of Host Variability on the Spread of Invasive Forest Diseases


1
Swiss Federal Institute for Forest, Snow and Landscape Research WSL, Zuercherstrasse 111, CH-8903 Birmensdorf, Switzerland
2
Swedish University of Agricultural Sciences, Southern Swedish Forest Research Centre, Sundsvägen 3, SE-23053 Alnarp, Sweden
*
Author to whom correspondence should be addressed.
Forests 2017, 8(3), 80; https://doi.org/10.3390/f8030080
Submission received: 1 February 2017
/
Revised: 5 March 2017
/
Accepted: 11 March 2017
/
Published: 15 March 2017
(This article belongs to the Special Issue Forest Pathology and Plant Health)
Abstract
:Biological invasions, resulting from deliberate and unintentional species transfers of insects, fungal and oomycete organisms, are a major consequence of globalization and pose a significant threat to biodiversity. Limiting damage by non-indigenous forest pathogens requires an understanding of their current and potential distributions, factors affecting disease spread, and development of appropriate management measures. In this review, we synthesize innate characteristics of invading organisms (notably mating system, reproduction type, and dispersal mechanisms) and key factors of the host population (namely host diversity, host connectivity, and host susceptibility) that govern spread and impact of invasive forest pathogens at various scales post-introduction and establishment. We examine spread dynamics for well-known invasive forest pathogens, Hymenoscyphus fraxineus (T. Kowalski) Baral, Queloz, Hosoya, comb. nov., causing ash dieback in Europe, and Cryphonectria parasitica, (Murr.) Barr, causing chestnut blight in both North America and Europe, illustrating the importance of host variability (diversity, connectivity, susceptibility) in their invasion success. While alien pathogen entry has proven difficult to control, and new biological introductions are indeed inevitable, elucidating the key processes underlying host variability is crucial for scientists and managers aimed at developing effective strategies to prevent future movement of organisms and preserve intact ecosystems.
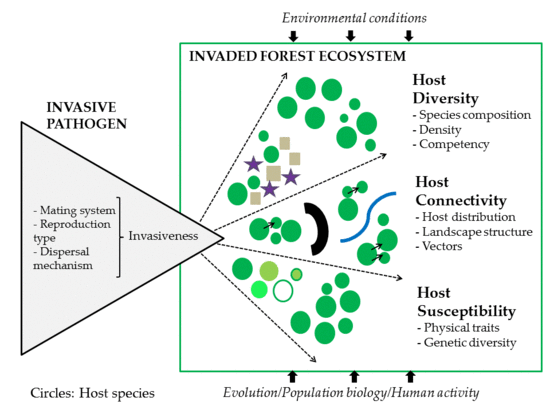
1. Introduction
Increased connectivity and globalization have greatly accelerated the frequency and magnitude of biological invasions around the globe by facilitating the long-distance movement of species into regions outside their historical distribution range. The current rate of non-indigenous species movement, resulting from human mediated intervention, is unprecedented [1], and has been the main accelerator driving the increase in novel encounters between host plants and pathogens, and the probability of invasive species emergence [2]. Biosecurity policies adopted by most countries for preventing new introductions (e.g., inspections at ports of entry, phytosanitary certification) can have positive effects, but, unless efforts are further strengthened and better coordinated internationally, biological invasions will inevitably continue [3].
Recent reviews have highlighted the escalating rate of exotic pathogen species introductions affecting forest trees in North America and Europe [4,5,6], mainly as a result of changes in trade practices surrounding the global movement of live plants and plant materials [7,8]. In some cases, alien forest pathogens (i.e., those that are nonnative, introduced from distant countries) have caused large-scale transformations of native ecosystems and changed the ecological dynamics through local and regional extinction of native host species. For example, the host-specialist pathogen Cryphonectria parasitica, the causal agent of chestnut blight, has dramatically reduced populations of Castanea dentata (Marsh.) Borkh. in North America where it had a defined role as a keystone species in ecosystem structure and function [9]. Generalist pathogens are thought to be better invaders than specialists due to their non-selective ability of seeking out new hosts in a new environment. For example, the oomycete plant pathogen Phytophthora ramorum (Werres, De Cock & Man in’t Veld), has a broad host range affecting both natural (forests) and semi-natural (urban green area) environments and hardy nursery stock. In western USA, P. ramorum has largely influenced the California oak woodlands landscape through diffuse impacts on a large number of host species [10,11], causing lethal stem infections on ‘canker hosts’ (mainly Notholithocarpus densiflorus (Hook. & Arn.) Manos, Cannon & S.H. Oh and Quercus spp.), and non-lethal foliar infections on ‘foliar hosts’ (e.g., Umbellularia californica (Hook. & Arn.) Nutt.).
In the strictest sense, ‘spread’ (also synonymous with the terms ‘dissemination’ and ‘dispersal’) refers to the movement of individuals either through random diffusion or directed dispersal such that they expand beyond the frontier of their geographic range [12]. The expansion phase is generally described by the change in range distance as a function of time [13]. Pathogens become ’invasive’ when they acquire a competitive advantage in a new area following the disappearance of natural obstacles to their proliferation including native antagonists, and successfully adapt to new conditions [14]. Most experimental studies (including modeling) on the spread of invasive pathogens focus on agricultural systems, humans, and animals. Given the importance of invasive pathogens currently causing global threats to biodiversity, a deeper understanding of spread mechanisms and dynamics of spread can improve the ability to predict and manage impacts of biological invasions affecting forests and urban landscapes. Though several recent reviews of invasive forest pathogens have highlighted human activity and international trade as major determinants of invasiveness [5,6], surprisingly few have focused specifically on factors driving disease spread of invasive pathogens across various scales from a local stand level to a broader landscape level.
The goal of this review is to discuss mechanisms underlying the invasion process for alien pathogens and disease spread, post-introduction and establishment. We present a framework for local, regional, and continental-scale factors governing the spread and impact of invasive forest pathogens. Specifically, we recapitulate characteristics and modes of pathogen reproduction, pathogen dispersal, and parasitic specialization, in relation to spread and epidemiology. We provide an overview of the main factors affecting variability in disease spread: host diversity, host connectivity, and host susceptibility and use one historical and one recent example of invasive pathogens that have significantly impacted forests to illustrate their invasion success and spread dynamics in that context. Finally, we discuss the way-forward in which a deeper understanding of the factors promoting disease spread across local and continental scales can help address the global ecological and societal challenges of managing novel disease epidemics.
1.1. Novel Environments, Novel Hosts
Biological invasions are a special type of range expansion [15] that can be defined by a series of discrete, consecutive phases which include: ‘Arrival’—single or multiple arrivals of a nonindigenous species at one or more points of entry into a new locale/environment, and ‘Establishment’—whereby arriving populations start to reproduce in situ, surpassing barriers of initial extinction (geographic, environment, reproductive, antagonists) [16,17,18].
After arrival and initial invasion of a new host (‘spillover’), production of transmission stages within the new host, and subsequent establishment, invasive forest pathogens can then ‘Spread’ expanding their range of occupied territory into new hospitable areas. This ability to spread is highly dependent on environmental suitability, resource availability, and the ability to adapt and naturalize [12]. Strong filtering exists between the different phases; successful invasion is a rare event such that only a small fraction of alien species survive to pass through and establish themselves in a novel environment. Both host availability (as a suitable ecological niche) and environmental conditions (extremes of temperature, moisture, and UV radiation) in the new location place strong limitations on a pathogen’s survival, its ability to reproduce and disperse, and subsequently spread [19,20]. These factors act as a strong selection filter leading to rapid adaptation to new environmental conditions and rapid evolution and exploitation of novel hosts [20]. Understanding novel forest pathogen introductions and the factors driving invasion success requires a deeper understanding of the invasion sequence that is conditioned by local or long-distance transport mechanisms from their native habitat to a novel environment, as well as environmental conditions and ecological factors determining an organism’s survival and reproducibility, and any population and community effects affecting their dynamics across a range of spatial and temporal scales (see Figure 1).
The most accurate records documenting the spread of forest pathogens can be derived from field mapping of invasion fronts over successive years from a defined geographic area, or from a time series of aerial photos. The kind and quality of data that is used as a measure of increasing abundance (presence/absence), the scale of mapping, and range size is important for extrapolating information about spread rates [21]. For example, ground surveys and geographic information system (GIS) mapping of Dutch elm-diseased and elm bark beetles-attacked trees on the geographically isolated island of Gotland allowed for tracking patterns of disease spread in relation to management strategies to reduce disease prevalence [22]. For many invading forest pathogens, the key to understanding dispersal is by measuring the human transport process through imported plant material via random checks in quarantine facilities from nurseries [23,24], or on vehicles [25]. For example, the spread of P. ramorum has been demonstrated using various approaches such as environmental niche models [26], risk assessment maps based on host distribution [27], landscape structure [28,29], multi-scale patterns of human activity [30], and trade networks [31]. When patterns of past spread are missing, future spread may be forecasted with simulations using either a mechanistic model or extrapolations (e.g., [32,33]).
1.2. Pathogen Invasiveness Affected by Species Traits
Certain species traits can favor invasiveness of pathogens and their spread and subsequent impact on forest ecosystems [34]. To some extent the plasticity of pathogen traits allows for some level of pre-adaption prior to an introduction. Janzen [35] described ‘ecological fitting’ as the interaction of an organism with its biotic and abiotic environment in a way that indicates a shared evolutionary history, when the organismal traits relevant to the interaction actually evolved elsewhere under different environmental conditions. Ecological fitting of novel forest pathogens depends in part on the ability of certain organismal traits to be co-opted for novel functions [36], and is largely influenced by phenotypic plasticity, correlated trait evolution, and phylogenetic conservatism [37].
Among the most important traits affecting spread ability for invasive forest pathogens is the mating system [38], namely for its ability to generate more virulent strains [34] and also to adapt to newly encountered host species in a new environment [20], and the type of reproduction (e.g., polycyclic pathogens complete their lifecycle multiple times throughout the growing season). Bazin et al. [39] showed that invasion dynamics of an introduced population are largely affected by the rate of asexuality. It is generally assumed that purely asexual organisms may exhibit lower invasion success compared to other organisms with, for example, mixed mating systems due to their inability to generate new sets of meiotic progeny which can rapidly adapt to the new host and environment [40]. In addition, asexual spores are dispersed mainly over short distances or at the plant level. Several examples, however, show that clonality does not necessarily reduce invasiveness. For instance, new populations of C. parasitica in Europe are frequently founded by one or a few genotypes (e.g., [41]). Similarly, Laurel wilt disease, which in the USA is threatening communities of native plant species in the family Lauraceae, is caused by the clonal ascomycete fungus Raffaelea lauricola T.C. Harr., Fraedrich & Aghayeva 2008 [42].
Other important species traits include: spore shape, which affects the release, transport, and deposition of inoculum (especially for aerial pathogens), and long-distance dispersal mechanisms (e.g., mito- or meiospores mediated by wind, running water, or vectors versus rain splash only) [34]. Other traits affecting spread, but perhaps of lesser importance than sexuality, spore shape, and dispersal mechanisms, include pluricellular spores which can facilitate survival in stressful environments, and abiotic niche characteristics such as the climate in the area of origin and the pathogen’s optimal temperature (as a proxy of climate-matching) [34].
Propagative spores are produced in many different ways, in for example, sporangia or simple or complex conidiophores in or on ascocarps and basidiocarps, by budding or fusion, within pycnidia, perithecia, or other various types [43]. The number of spores produced per unit area of infected leaf tissue can be enormous, (e.g., a relatively small apothecium can produce several million spores). The duration and periodicity of sporulation is as important as the number of spores produced [44]. Some pathogens produce an almost continuous crop of propagules, while others may have sporophores that bear several successive propagules (e.g., Phytophthora spp.). This capacity to produce a steady stream of infectious propagules over a prolonged period of time is advantageous to the pathogen [43]. Following initial primary infection on a suitable susceptible host plant, pathogens may undergo secondary spread to the same or new host plant species within the same location or to new hosts in another location. The transmission of a pathogen is highly dependent on its ability to produce numerous spores or infectious propagules under favorable environmental conditions that are dispersed and then deposited in a viable condition on a susceptible host plant under conditions conductive for infection. In many cases, pathogens also possess mechanisms to survive between periods of unfavorable environmental conditions (through formation of resting structures).
In nature, organisms can move or be transferred over short and large distances. Stratified dispersal, a two-scale dispersal process of combined short-distance, continuous dispersal, and discontinuous, long-range dispersal, is a major driver of spread dynamics [45]. Where long-distance dispersal events are normally rare, but facilitated by inadvertent human transport, they can cause much greater rates of spread than that which would normally occur with short-range dispersal since populations jump well ahead of the advancing invasion front [13]. Subsequently, isolated populations become established far from the moving population front, and will grow and eventually coalesce with the source population to significantly advance the population front.
Plant pathogens typically disseminate through direct transmission and indirect (passive) transmission (Table 1). With direct transmission, pathogen dispersal occurs intrinsically alongside seeds (germinative) and/or other plant parts (vegetative). Indirect transmission may be autonomous, by wind, water, insect, or mammalian vectors, and human-mediated. Many pathogens are dispersed by more than one mechanism. The importance of understanding these organismal traits and how they affect spread dynamics is essential for preventative and predictive actions. For example, during Pest Risk Assessment (PRA) the likelihood of pathogen spread within an importing country or region considers such factors like the dispersal potential as it relates to the pathogen’s reproductive potential (rated by the presence of multiple generations per year or growing season, and the relative number of offspring or propagules per generation), the pathogen’s inherent mobility (e.g., rapid movement), and external dispersal facilitation modes (e.g., the presence of natural barriers or enemies, and dissemination enhanced by wind, water, vectors, or human assistance).
2. Host Factors
2.1. Diversity
Most studies recognize that plant diversity can affect disease prevalence and spread through the direct effect of host density (plant species richness) on the transmission of plant pathogens and the role that plant diversity has in influencing host density through, for example, competition (e.g., [48,49]). Although most plant species are susceptible to infection and damage by one or more pathogens, some species or individuals may exhibit different degrees of susceptibility [50]. For example, within the genus Fraxinus, there is large variation between species in susceptibility to the ash dieback pathogen Hymenoscyphus fraxineus [51,52] (see Section 3.1 Case studies), and large genotypic effects in susceptibility among individuals of European ash (Fraxinus excelsior L.) [53]. Similarly, R. lauricola affects several members of the laurel family (Lauraceae) in the USA though lethal damage is most prominent on only a few native hosts including red bay (Persea borbonia (L.) Spreng. sensu stricto [42,54]).
In ecological communities, a high diversity of plant species usually contributes to maintaining the functional integrity of the ecosystem (’insurance hypothesis’; e.g., [55]). As different species show varying responses to a specific pathogen, a high species diversity will act as a sort of buffer, diluting the effects of the pathogen on the ecosystem (‘diversity-disease hypothesis’ or ‘dilution hypothesis’; [56,57]). When pathogen transmission is density dependent or where the host range is narrow, biodiversity can alter infection prevalence through a change in the absolute abundance of important hosts and any associated vectors [58]. For example, a non-host species may reduce the probability of encountering hosts and therefore lessening opportunities for healthy susceptible individuals to become infected [56]. Similarly, in the case of an environmentally transmitted disease, added species abundance can reduce the probability that contact between individuals lead to transmission, thereby leading to the idea of encounter reduction [56,59]. In contrast, if added species function as alternative sources of infection, or serve as a source for increased vector activity, disease prevalence may subsequently increase [56]. Thus, host biodiversity can largely influence disease spread through interspecific variability in suitable host species. At the same time, a mixture of species including non-hosts, competent hosts (i.e., with a high effectiveness of passing on infection), and less competent hosts, may intercept inoculum and also reduce the spread of a pathogen [48].
Monocultures of host species, despite a few exceptions, are highly susceptible to epidemics of invasive pathogens because of the lack of intraspecific host diversity [49,50]. Moreover, hosts in high quantity and density, as observed in monocultures, mean reduced distance to which inoculum must traverse to spread between plants, consequently increasing pathogen transmission [59]. This is particularly relevant for competent hosts on which the pathogen may readily produce inoculum. In humans, when transmission of a specialist pathogen is density-dependent, theory predicts a minimal density of the host population below which the pathogen becomes extinct (‘crowd disease’, [60]). In mixed communities, density-dependent disease dynamics confer an advantage to uncommon species which benefit from a lower enemy pressure (‘rare-species advantage’; [61]) and may, therefore, increase in incidence. Examples where host density has affected the spread of an invasive forest pathogen include the beech bark disease involving the exotic beech scale insect Cryptococcus fagisuga Lind. and the exotic fungus Neonectria coccinea var. faginata (Pers.) Fr. in North America. Morin et al. [62] showed that about 100 years after its first detection the disease had invaded most regions where American beech (Fagus grandifolia Ehrh.) is a dominant component of stands, but not the regions where the host occurs at low densities. Noteworthy, a study conducted in northern Maine showed that the disease epidemic is also influenced by climatic conditions (e.g., winter temperatures) which affect the survival of the insect vector [63]. A host-density dependent disease dynamic was also revealed using a model by Hatala et al. [64] for the invasive white pine blister rust Cronartium ribicola J.C. Fisch in the forests of the Greater Yellowstone Ecosystem. In general, these examples among others suggest that the risk of disease is lower if the competent host for the invasive pathogen comprises a small fraction of the overall diverse host community.
The above discussed points may, however, not strictly apply to invasive pathogens infecting a wide range of host species (i.e., polyphagous or generalist pathogens). In such case, the “insurance hypothesis” can fail and hosts driving the epidemics seem to be decisive. Weste et al. [65] showed that in the major types of forest and woodland of the Grampians (Western Australia) the activity of the generalist root pathogen Phytophthora cinnamomi Rands. in certain cases resulted in important changes in species composition and community structure. The high functional diversity of the local ecosystems could not prevent the spread of this invasive oomycete. In mixed forests of coastal California, the spread of P. ramorum, the causal agent of Sudden Oak Death, is driven by foliar hosts, on which this polyphagous pathogen sporulates [66]. These so-called ‘reservoir hosts’ showing only cryptic or asymptomatic infections but acting as a source of inoculum for other hosts may be difficult to predict and can also play a major role in the epidemic of invasive pathogens. This, for example, is also the case for the invasive Rhododendron ponticum and P. ramorum and P. kernoviae in the UK [67], and for non-beech hardwood hosts (e.g., Acer rubrum L.) and bark beech disease in North America [68].
2.2. Connectivity
The spatial structure of the host population influences the spread of invasive pathogens both at the local (forest stand) and regional (landscape) scale [69,70]. In a forest stand, the epidemic spread rates are driven by the contacts between infected and healthy hosts or between vectors and infected/healthy hosts [71]. Both types of contact strongly depend on host density, which, in mixed forests, may vary significantly. Frequently, hosts show a clumped (aggregated) distribution, with a ‘patch phase’ (i.e., higher than average host density) and a ‘gap phase’ (i.e., lower than average host density) [59]. For pathogens spreading via active vectors (e.g., insects), the spatial heterogeneity of hosts can hinder the advance of the vector and/or physically separate the pathogen from the vector and consequently limit the spread of the disease [72,73]. Similarly, if the host heterogeneity scales at distances over which a pathogen is transmitted, a spatial variation in host density may constrain the spread of pathogens that rely on passive vectors (e.g., wind, water).
At the landscape scale, invasive pathogens generally first colonize areas with continuous forests and then, eventually, isolated (scattered) forest stands or trees (e.g., chestnut blight, see Section 3.2 Case studies). A scattered distribution of hosts does not always allow them to escape infection. For instance, although butternut (Juglans cinerea L.) in North America usually occurs as scattered individuals or in small groups in deciduous and mixed forests, it could not escape infection by the invasive canker pathogen Ophiognomonia clavigignenti-juglandacearum (N.B. Nair, Kostichka & J.E. Kuntze) Broders & Boland, which was dispersed over longer distances by beetle vectors and infected seeds [74]. Similarly, a fragmented distribution of some white pines (e.g., Pinus strobiformis Engelm.) in western North America could only retard, but not prevent their infection by the white pine blister rust fungus C. ribicola [75]. When hosts are scattered, landscape connectivity (i.e., how the landscape structure facilitates or impedes the disease spread among patches [76]) plays an important role for disease spread. Such connectivity is strongly related to landscape structure, which, as shown by Real and Biek [77] for rabies, can present two possibilities; namely individuals aggregated over a uniform landscape or individuals assorted by the environment into different spatial locations. This second possibility most likely applies to trees whose distribution is mainly shaped by environmental features [78]. Although most studies on the influence of landscape heterogeneity on disease dynamics have focused on agricultural systems, frequently using simulation models (e.g., [76]), in recent years the interest in how landscape features affect the spread of forest pathogens has increased considerably (landscape pathology; [70]). Several investigations have dealt with the spread of P. ramorum in coastal California. For example, Condeso and Meentemeyer [28] showed that the effect of forest fragmentation on disease severity is scale-dependent. In another study, Ellis et al. [79] demonstrated that environmental variables were relatively more important than landscape connectivity in shaping the spatial pattern of Sudden Oak Death. Filipe et al. [29] found that host-free barriers would contain the spread of P. ramorum for a significantly long time only if combined with additional buffers (e.g., topographic conditions).
In a landscape still not completely colonized by an invasive pathogen, the connectivity may be increased by additional introductions, which also have the potential to introduce new genotypes [80,81], or by insect vectors that may actively or passively spread the pathogen over long distances. Fusarium circinatum Nirenberg and O’Donnell, the causal agent of pitch canker, is a potentially dangerous invasive pathogen in pine (Pinus sp.) forests in Europe. Möykkynen et al. [82] modeled the rate of spread of F. circinatum as a function of several factors, among which included host distribution and flight distance of insect vectors. Their model showed that because of the short distance at which spores are dispersed and the fragmentation of pine forests, unless there will be new introductions, the pathogen will most likely not spread to northern Europe. In a broader sense, one of the main reasons for the increasing number of invasive pathogens which are spread around the planet is the high global connectivity through transportation and trade networks [5,6,7].
2.3. Susceptibility
Intraspecific variability in host susceptibility is a main source of heterogeneity controlling the trajectory of a disease epidemic [83]. The susceptibility of a host to a specific pathogen’s transmission, maintenance, and proliferation is influenced by physical host traits (e.g., size, morphology), external factors (e.g., availability of nutrients, local microclimate, topography), and, of course, the genetic host background [49,84]. Tree size, which often correlates with tree age, may show a contradictory effect on host susceptibility. Frequently, the impact of invasive pathogens is greater on larger trees, probably because of the larger contact area available for the pathogen and its vectors and/or traits correlated with age (e.g., vigor; [57,85]). A study by Jules et al. [25] indicated that in Oregon and California large Port Orford cedars (Chamaecyparis lawsoniana (A. Murray) Parl.) located in close proximity of streams and roads were more likely to be killed by the invasive oomycete Phytophthora lateralis Tucker & Milbrath. According to that study, high susceptibility was due to the larger root systems and the position of the trees which allowed the roots to reach the water and, thus, be a good target for waterborne zoospores. On the other hand, in Europe the impact of oak (Quercus sp.) powdery mildew caused mainly by the invasive fungus Erysiphe alphitoides Griffon & Maublanc seems to be more severe on seedlings, particularly in natural regeneration, than on adult trees [86]. Age-related susceptibility has also been documented for Ophiostoma novo-ulmi (Brasier) causing Dutch elm disease on Ulmus spp., whereby younger trees, possessing smaller diameter vessels, are generally more tolerant [87].
Host population genetic diversity appears to play an important role in buffering populations against disease epidemics [88]. In fact, empirical observations and modeling studies indicate that in a population of a host species intraspecific genetic diversity represents the best insurance against pathogens [89]. According to Garrett and Mundt [90], a mixture of host genotypes reduces a disease in three ways: first, the presence of less susceptible/resistant genotypes dilutes the inoculum; second, less susceptible/resistant genotypes represent a physical barrier for inoculum deposition; and third, the potential for induced host resistance is increased. Monocultures of genetically similar or identical (clones) trees are usually highly susceptible to invasive pathogens (the so-called ’monoculture effect’ earlier mentioned; [91,92]). Numerous examples worldwide support this general assumption, including plantations of Pinus radiata D. Don in the Southern hemisphere affected by needle blight (Dothistroma septosporum (Dorog.) Morelet) [93] or Eucalyptus spp. plantations in south-east Asia which are susceptible to leaf, bud, and shoot blight caused by Teratosphaeria destructans (M.J. Wingf. & Crous) [94]. Increased host susceptibility to infectious parasites may also be due to reduced individual-level and population-level genetic heterozygosity, which may increase the occurrence of inbreeding [95,96].
Usually, in the initial phase of an invasive pathogen epidemic, either resistance in the host population is completely lacking or resistant host genotypes are at too low of a frequency to reduce the effects of the pathogen [97,98]. Successively, the continued exposure to the pathogen will select for less susceptible host genotypes, which will increase in frequency. However, if the impact of an invasive pathogen is extreme with large-scale mortality within a relatively short-time frame, host resistance may never evolve due to rapid elimination of the host species. In forest trees, large differences in generation times may also be a disadvantage for developing resistance to invasive pathogens. As stated by Aegerter and Gordon [97], for such a directional selection to be successful, young individuals in a population have to be challenged by a pathogen after the stage where the physiological mechanisms for resistance can be operative. The same authors showed that in Pinus radiata different mechanisms of resistance against Fusarium circinatum are active in seedlings and adult trees. Still, benefits deriving from a genetically diverse host population may also depend on the genetic diversity of the pathogen population [99].
3. Case Studies
Following initial introduction and establishment, spread rates for alien forest pathogens are typically slower than alien insect pests due in part to dispersal mechanisms. The following two examples of invasive forest pathogens serve to illustrate their spread post-introduction and establishment and their effects on local ecosystems.
3.1. Ash Dieback
The invasion of the ash dieback pathogen Hymenoscyphus fraxineus (Table 2) into Europe and its subsequent spatial spread to most European countries throughout the natural distribution range of native Fraxinus species can been characterized by relatively slow spread dispersal by airborne spores, coupled with some few pulsed events involving long-distance establishment via anthropogenic means within or ahead of the invasion boundary namely through—then, unknowing or naïve movement of nursery stock from commercial nurseries within or outside already-infested areas [100,101]. Ash dieback has had variable recognizable consequences, partially driven by both the known and still unknown or unrecognized importance of the species from a forestry or nature conservations perspective. Mitchell et al. [102] suggest that the disease could have wide-ranging ecological implications particularly for obligate-associated organisms to ash and the indirect effect of the disease on nutrient cycling in woodland ecosystems. Baseline information on tree species contributions to ecosystem functions is necessary in order to determine actual short-term impacts on light penetration, nitrogen cycling, and primary production, long-term impacts involving interactions with other abiotic or biotic stress factors, and any compensatory effects of other tree species in the post-epidemic/decline phase.
Spread rates for ash dieback have been difficult to accurately track due to the lag time in reporting disease presence in various countries. However, where good records have been kept, mean spread rates have ranged between 30 and up to 75 km per year [103]. Regular monitoring of disease presence at the onset of its introduction to Norway initially gave an annual mean spread rate of 30 km, but then it increased rapidly in subsequent years averaging more than 50 km per year [104]. In Sweden, the disease was observed already in 2001 in only a few places and within just a few years, the occurrence of ash dieback was reported on trees of all age classes throughout the natural distribution range of F. excelsior in that country [105]. By 2010, ash was added to the Swedish Red-List with vulnerable status, and recently elevated to critically endangered status considered to be at high risk of extinction in the wild.
Some factors contributing to variable rates of disease spread among countries may be due to large differences in natural geographic barriers that may limit natural dispersal, the length of growing (and hence sporulation) seasons, fragmented distribution and density of the host species throughout Europe, between and within-season fluctuations in optimal climate conditions, and stratified dispersal also involving the import or transfer of diseased plants which resulted in large jumps ahead of the advancing infection front. Within the nearly 20 years since damage was first reported in Lithuania and Poland, the pathogen has spread throughout most of western and eastern Europe where native Fraxinus spp. are growing. Within this region, the large majority of native ash species (especially F. excelsior) are highly susceptible, and some non-native species planted within the zone of infestation also exhibit moderate to high susceptibility. Fortunately, a small proportion (<5%) of the natural population has shown better tolerance to the disease, which offers the potential to revitalize and restore forest and urban landscapes through breeding for resistance [53].
3.2. Chestnut Blight
Chestnut blight caused by the fungus Cryphonectria parasitica (Table 2) is most likely the best-known example of an invasive forest disease in Europe and North America. After its first discovery, the pathogen rapidly colonized the whole geographic range of the susceptible host species in the two continents. In eastern North America, the fungus spread at a rate of more than 30 km per year throughout the approx. 800,000 km2 of native American chestnut forests [106]. In some stands, C. dentata accounted for more than 50% of the basal area of standing trees and was the canopy forming species [107]. Within 50 years, this species was confined to the understory, with significant ecological and economic consequences [108]. This dramatic course of the epidemic was favored by the high susceptibility of American chestnut to the introduced pathogen [109]. Nevertheless, it is still not clear whether the low genetic diversity of C. dentata compared to that of the congeneric species is the result or also a cause of the species decline due to chestnut blight [110]. Recent population genetic analyses [111] showed that the initial introduction of C. parasitica into North America occurred from the main Japanese island Honshu. Later on, the fungus was also introduced from other Japanese regions, China, and Korea. Introductions into Europe occurred both from North America (into Italy) and Asia (into south-western France) [111]. Although European chestnut is slightly less susceptible to C. parasitica than the American chestnut, the spread of the pathogen on the European continent was also rapid. Thirty years after its first detection, the disease was reported in the main chestnut growing areas of Europe and to date chestnut blight is widespread throughout most of the distribution range of C. sativa [112,113]. The presence of important geographic barriers and the sometimes scattered distribution of chestnut stands could only slow down, but not stop the spread of the pathogen. For instance, in Switzerland chestnut blight was first observed south of the Alps in 1948 [41]. About forty years later, despite the potential barrier represented by the Alps and the adoption of quarantine regulations, the disease also appeared in the scattered stands north of the Alps. The host connectivity in Europe has likely increased through several introduction events targeting different regions, which unintentionally propagated C. parasitica via infected plant material.
Due to the unexpected appearance and spread of natural hypovirulence within the C. parasitica population, however, consequences of chestnut blight in Europe were less dramatic than in North America. European chestnut stands, in spite of a high disease incidence, have survived the epidemics and are still successfully fulfilling their important ecological and cultural functions. Nevertheless, differences in the success of biocontrol by hypovirulence exist among chestnut-growing regions, which may be explained by several factors, including the diversity of the local C. parasitica population in terms of vegetative compatibility (vc) types, the subtype of the occurring hypovirus, the presence of adequate vectors, management practices, and variable environmental conditions [113,114].
In eastern North America, biological control has failed nearly completely due to the high vc type diversity of the local C. parasitica populations and the high susceptibility of C. dentata to the pathogen [114]. To increase the chance of establishment of the hypovirus, transgenic C. parasitica strains which transmit the hypovirus also to the sexual spores (ascospores), as well as super hypovirus donor strains, have been recently used [115,116]. The success of these fungal strains in the field, however, still needs to be demonstrated. From the host side, blight-resistant chestnut trees have been obtained by backcrossing the resistance to C. parasitica of the Chinese chestnut into the genome of the American chestnut [117]. Recently, a chestnut tree with an increased resistance to chestnut blight was also created using a transgenic approach [118].
4. “The Way Forward”
The study of invasive species epidemiology will continue to be important in the future as new introductions associated with anthropogenic activity and novel plant-organismal interactions will inevitably continue to cause disease epidemics around the globe. Despite recent advances in our understanding of pathways for biological introductions and factors contributing to emerging invasive diseases, much remains to be learned. Here, we mention some future challenges and research priorities which are strongly related to the spread of invasive forest pathogens.
4.1. Host Diversity Threshold
As mentioned above, diversity of plant species can strongly influence the spread of invasive forest pathogens by modifying the relative abundance of host species. In particular, a high species diversity negatively affects the spread of pathogens with a limited host range or whose transmission is density dependent. Thus, we could imagine that in a forest a threshold of tree species diversity may exist, above which an invasive pathogen may not be able to become established and spread. Determining the diversity threshold of a given pathosystem toward specific invasive pathogens would be of great value for estimating the potential vulnerability of ecosystems. An eventual high vulnerability could be reduced by introducing new, possibly non-host or less susceptible, tree species. Although this approach may be difficult to apply to already existing natural forests, it could be adopted when establishing new artificial forests. As shown by studies conducted with agricultural crops (e.g., [73,130]), not only plant diversity but also the spatial organization of the host species may influence the spread of pathogens. Therefore, when introducing additional plant diversity, host spatial heterogeneity should also be created.
A similar ‘diversity threshold’ approach could also target the intraspecific genetic diversity of the host. Although it is known that a high genetic variation within a specific host represents the best insurance against pathogens, only a few studies have examined the rate of spatial spread of invasive forest pathogens as a function of the genetic diversity of the host population. In particular, almost no information is available about how much host genetic diversity is needed for the effects to be realized, specifically for the spread of a pathogen to be reduced and less susceptible host genotypes to emerge.
4.2. Spread Dynamics
Predicting the spread dynamics (e.g., when the invader will arrive at a specific location, from where it will arrive) of an invasive pathogen after its establishment in a new area is of importance for developing and applying effective management strategies. In recent years, network theory has started to be used to model the epidemiology of plant pests and diseases (e.g., [131]). For example, Ferrari et al. [132] developed a dynamic network model for analyzing the spread of the invasive phloem-boring pest Adelges tsugae Annand (hemlock woolly adelgid) in the eastern United States. They showed that this kind of model, which allows connections to change through time, can provide valuable information about the spatio-temporal dynamics of invasion processes. Harwood et al. [31] developed a simulation model of the P. ramorum epidemic development in the UK which included information on the spatial distribution of hosts and a realistic network of plant trade. For several reasons (e.g., heterogeneity of the landscape, patchiness of host distribution, and environmental conditions; [131]), however, network theory is still poorly applied to invasive pathogens in forest ecosystems [133]. In the future, approaches combining network analysis with landscape genetic analyses [134], monitoring data (e.g., host distribution in the target region), and realistic network data (e.g., plant trade) should become a standard tool for predicting (and reconstructing) the spread of invasive plant diseases.
4.3. How to Slow the Spread
The eradication of invasive forest pathogens post-establishment is difficult and has been rarely achieved [135]. For example, in the case of ash dieback, photographic evidence suggests the disease likely had reached an epidemic level by the time conspicuous symptoms were formally documented and had been noticed by foresters in the mid-1990s. Therefore, slowing the spread may be a more realistic objective. A premise for such a goal to be reached is the early detection of a pathogen by optimizing the monitoring strategies (for a modeling approach, see e.g., [136]). Once an invasive pathogen is detected, the ability to actually slow down its spread and the type of management strategy to be adopted will vary between pathogen species due to differences in their infection biology. However, any attempts to minimize the spread and impact of invasive pathogens must ideally consider all factors that influence their dispersal, population growth, or a combination of both. For example, long distance dispersal events (frequently mediated by human activity) bringing a pathogen ahead of the actual advancing front of the epidemic, even if difficult to predict, are an important determinant of spread rates. Hence, even smaller efforts aimed at minimizing long-distance dispersal can greatly reduce pathogen spread [137]. Among possible approaches to be considered, potential strategies might include optimizing pathogen detection/diagnostics and biosurveillance monitoring [138], reducing landscape connectivity and susceptibility by adopting adequate land-use strategies (e.g., promoting increased diversity in mixed plantations instead of monocultures), or, in particular for soil borne pathogens, careful management of routine operations, recreation activities, and machinery traffic in and around known infested areas.
4.4. Evolution of Invasive Forest Pathogens
Invading populations are faced with new environmental conditions to which they have to respond rapidly [139,140]. In the case of invasive forest pathogens, changes in the selective regime (e.g., increased selection for adapted genotypes) compared to the native range, combined with intrinsic characteristics (e.g., genetic diversity) of the invading population may influence the dynamic of an epidemic. It is easy to predict that adaptations in response to invasions will also affect the dispersal ability of a pathogen, thereby introducing uncertainty in the prediction of the rate of its range expansion. Additional uncertainty may be provided by the ability of some pathogens to overcome the so-called ’host species barrier‘ and infect a new host (‘host jump’; [141]), which could accelerate the spread and amplify the consequences of an invasion. Similarly, hybridization events of an established invasive pathogen with native or other similar invasive species [46] could originate new, rapidly-spreading invasive species. Unfortunately, both host jumps and hybridization events of invasive species are particularly difficult to predict. Of importance is also the fact that invading pathogen populations can also induce evolutionary changes in their host populations. For example, virulent host genotypes may select for less susceptible host genotypes. Last, but not least, the coevolution of an invasive pathogen with its new host will also be influenced by the environmental conditions.
5. Conclusions
Understanding the host variability as it is affected by host diversity, host connectivity, and host genetics (Figure 1) will improve our ability to predict invasion success and potential consequences to local ecosystem health and services—such as carbon sequestration, nutrient cycling—on which humans rely. Quite often data related to host species characteristics under various local driver values (e.g., gradients of temperature, moisture, elevation, human activities) is either lacking altogether, is fragmented, or has emerged slowly because sampling is too local and often not adequate enough to apply across a landscape-level [2].
Further complicating our ability to predict and manage disease epidemics is ‘episodic temporal variation’, where the success of a new invader only coincides with, for example, a disturbance, or major rainfall event [21]. In addition, changing land-use patterns and changes in climate can and will continue to influence the range expansion of native species affecting the spread and impact of introduced non-native species [2]. Continued efforts on the avoidance of new introductions and recognizing the human factor affecting the emergence of pests and diseases, interdisciplinary research on emerging invasive diseases, and creative strategies for the mitigation of plant disease impacts in natural and semi-natural ecosystems, is warranted [6,142].
Acknowledgments
Funding was provided by The Swedish Research Council FORMAS, Carl Tryggers Stiftelse för Vetenskaplig Forskning and Kungl. Skogs- och Lantbruksakademien.
Author Contributions
M.C. and S.P. contributed equally to this work.
Conflicts of Interest
The authors declare no conflict of interest. The funding sponsors had no role in the design or writing of this review manuscript.
References
- Ricciardi, A. Are Modern Biological Invasions an Unprecedented Form of Global Change? Conserv. Biol. 2007, 21, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Crowl, T.A.; Cris, T.O.; Parmenter, R.R.; Belovsky, G.; Lugo, A.E. The spread of invasive species and infectious disease as drivers of ecosystem change. Front Ecol. Environ. 2008, 6, 238–246. [Google Scholar] [CrossRef]
- Lovett, G.M.; Weiss, M.; Liebhold, A.M.; Holmes, T.P.; Leung, B.; Lambert, K.F.; Orwig, D.A.; Campbell, F.T.; Rosenthal, J.; McCullough, D.G.; et al. Nonnative forest insects and pathogens in the United States: Impacts and policy options. Ecol. Appl. 2016, 26, 1437–1455. [Google Scholar] [CrossRef] [PubMed]
- Aukema, J.E.; McCullough, D.G.; Von Holle, B.; Liebhold, A.J.; Britton, K.O.; Frankel, S.J. Historical accumulation of non-indigenous forest pests in the continental United States. BioScience 2010, 60, 886–897. [Google Scholar] [CrossRef]
- Fisher, M.C.; Henk, D.A.; Briggs, C.; Brownstein, J.S.; Madoff, L.; McCraw, S.L.; Gurr, S. Emerging fungal threats to animal, plant and ecosystem health. Nature 2012, 484, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Santini, A.; Ghelardini, L.; de Pace, C.; Desprez-Loustau, M.L.; Capretti, P.; Chandelier, A.; Cech, T.; Chira, D.; Diamandis, S.; Gaitniekis, T.; et al. Biogeographical, patterns and determinants of invasion by forest pathogens in Europe. New Phytol. 2013, 197, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Liebhold, A.M.; Brockerhoff, E.G.; Garrett, L.J.; Parke, J.L.; Britton, K.O. Live plant imports: The major pathway for forest insect and pathogen invasions of the US. Front. Ecol. Environ. 2012, 10, 135–143. [Google Scholar] [CrossRef]
- Ghelardini, L.; Pepori, A.L.; Luchi, N.; Capretti, P.; Santini, A. Drivers of emerging fungal diseases of forest trees. For. Ecol. Manag. 2016, 381, 235–246. [Google Scholar] [CrossRef]
- Anagnostakis, S.L. Chestnut blight: The classical problem of an introduced pathogen. Mycologia 1987, 79, 23–37. [Google Scholar] [CrossRef]
- Hansen, E.M.; Kanaskie, A.; Prospero, S.; McWilliams, M.; Goheen, E.M.; Osterbauer, N.; Reeser, P.; Sutton, W. Epidemiology of Phytophthora ramorum in Oregon tanoak forests. Can. J. For. Res. 2008, 38, 1133–1143. [Google Scholar] [CrossRef]
- Rizzo, D.M.; Garbelotto, M. Sudden oak death: Endangering California and Oregon forest ecosystems. Front. Ecol. Environ. 2003, 1, 197–204. [Google Scholar] [CrossRef]
- Hui, C.; Krug, R.M.; Richardson, D.M. Modelling spread in invasion ecology: A synthesis. In Fifty Years of Invasion Ecology: The Legacy of Charles Elton, 1st ed.; Richardson, D.M., Ed.; Wiley-Blackwell: Oxford, UK, 2011; pp. 329–343. [Google Scholar]
- Shigesada, N.; Kawasaki, K.; Takeda, Y. Modelling stratified diffusion in biological invasions. Am. Nat. 1995, 146, 229–251. [Google Scholar] [CrossRef]
- Valéry, L.; Fritz, H.; Lefeuvre, J.C.; Simberloff, D. In search of a real definition of the biological invasion phenomenon itself. Biol. Invasions 2008, 10, 1345–1351. [Google Scholar] [CrossRef]
- Wilson, J.R.U.; Dormontt, E.E.; Prentis, P.J.; Lowe, A.J.; Richardson, D.M. Biogeographic concepts define invasion biology. TRENDS Ecol. Evol. 2009, 24, 586. [Google Scholar] [CrossRef]
- Lockwood, J.L.; Hoopes, M.F.; Marchetti, M.P. Invasion Ecology, 2nd ed.; Wiley Blackwell: West Sussex, UK, 2013. [Google Scholar]
- Smith, C.S.; Lonsdale, W.M.; Fortune, J. When to ignore advice: Invasion predictions and decision theory. Biol. Invasions 1999, 1, 89–96. [Google Scholar] [CrossRef]
- Mack, R.N.; Simberloff, D.; Lonsdale, W.M.; Evans, H.; Clout, M.; Bazzaz, F. Biotic invasions: Causes, epidemiology, global consequences, and control. Ecol. Appl. 2000, 10, 689–710. [Google Scholar] [CrossRef]
- Aylor, D.E. Spread of plant disease on a continental scale: Role of aerial dispersal of pathogens. Ecology 2003, 84, 1989–1997. [Google Scholar] [CrossRef]
- Parker, I.M.; Gilbert, G.S. The Evolutionary Ecology of Novel Plant-Pathogen Interactions. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 675–700. [Google Scholar] [CrossRef]
- Hastings, A.; Cuddington, K.; Davies, K.F.; Dugaw, C.J.; Elmendorf, S.; Freestone, A.; Harrison, S.; Holland, M.; Lambrinos, J.; Malvadkar, U.; et al. The spatial spread of invasions: New developments in theory and evidence. Ecol. Lett. 2005, 8, 91–101. [Google Scholar] [CrossRef]
- Menkis, A.; Östbrant, I.-L.; Wågström, K.; Vasaitis, R. Dutch elm disease on the island of Gotland: Monitoring disease vector and combat measures. Scand. J. For. Res. 2016, 31, 237–241. [Google Scholar] [CrossRef]
- Goss, E.M.; Larsen, M.; Vercauteren, A.; Werres, S.; Heungens, K.; Grünwald, N.J. Phytophthora ramorum in Canada: Evidence for migration within North America and from Europe. Phytopathology 2011, 101, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Goss, E.M.; Larsen, M.M.; Chastagner, G.A.; Givens, D.R.; Grünwald, N.J. Population genetic analysis infers migration pathways of Phytophthora ramorum in US nurseries. PLoS Pathol. 2009, 5, e1000583. [Google Scholar] [CrossRef] [PubMed]
- Jules, E.W.S.; Kauffman, M.J.; Ritts, W.D.; Carrol, A.L. Spread of an invasive pathogen over a variable landscape: A nonnative root rot on Port Orford cedar. Ecology 2002, 83, 3167–3181. [Google Scholar] [CrossRef]
- Kelly, M.; Gueo, Q.; Liu, D.; Shaari, D. Modelling the risk for a new invasive forest disease in the United States: An evaluation of five environmental niche models. Comput. Environ. Urban 2007, 31, 689–710. [Google Scholar] [CrossRef]
- Meentemeyer, R.; Rizzo, D.; Mark, W.; Lotz, E. Mapping the risk of establishment and spread of sudden oak death in California. For. Ecol. Manag. 2004, 200, 195–214. [Google Scholar] [CrossRef]
- Condeso, T.E.; Meentemeyer, R.K. Effects of landscape heterogeneity on the emerging forest disease sudden oak death. J. Ecol. 2007, 95, 364–375. [Google Scholar] [CrossRef]
- Filipe, J.A.N.; Cobb, R.C.; Meentemeyer, R.K.; Lee, C.A.; Valachovic, Y.S.; Cook, A.R.; Rizzo, D.M.; Gilligan, C.A. Landscape Epidemiology and Control of Pathogens with Cryptic and Long-Distance Dispersal: Sudden Oak Death in Northern Californian Forests. PLoS Comput. Biol. 2012, 8, e1002328. [Google Scholar] [CrossRef] [PubMed]
- Cushman, J.H.; Meentemeyer, R.K. Multi-scale patterns of human activity and the incidence of an exotic forest pathogen. J. Ecol. 2008, 96, 766–776. [Google Scholar] [CrossRef]
- Harwood, T.D.; Xu, X.; Pautasso, M.; Jeger, M.J.; Shaw, M.W. Epidemiological risk assessment using linked network and grid based modelling: Phytophthora ramorum and Phytophthora kernoviae in the UK. J. Ecol. Model. 2009, 220, 3353–3361. [Google Scholar] [CrossRef]
- Gilbert, M.; Liebhold, A. Comparing methods for measuring the rate of spread of invading populations. Ecography 2010, 33, 809–817. [Google Scholar] [CrossRef]
- Tisseuil, C.; Gryspeirt, A.; Lancelot, R.; Pioz, M.; Liebhold, A.; Gilbert, M. Evaluating methods to quantify spatial variation in the velocity of biological invasions. Ecography 2016, 39, 409–418. [Google Scholar] [CrossRef]
- Philibert, A.; Desprez-Loustau, M.-L.; Fabre, B.; Frey, P.; Halkett, F.; Husson, C.; Lung-Escarmant, B.; Marçais, B.; Robin, C.; Vacher, C.; et al. Predicting invasion success of forest pathogenic fungi from species traits. J. Appl. Ecol. 2011, 48, 1381–1390. [Google Scholar] [CrossRef]
- Janzen, D.H. On Ecological Fitting. Oikos 1985, 45, 308–310. [Google Scholar] [CrossRef]
- Brooks, D.R.; McLennan, D.A. The Nature of Diversity: An Evolutionary Voyage of Discovery; University of Chicago Press: Chicago, IL, USA, 2002. [Google Scholar]
- Agosta, S.J.; Klemens, J.A. Ecological fitting by phenotypically flexible genotypes: Implications for species associations, community assembly and evolution. Ecol. Lett. 2008, 11, 1123–1134. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.H.D.; Burdon, J.J. Mating systems and colonizing success in plants. In Colonization, Succession and Stability; Cary, A.J., Crawley, M.J., Edwards, P.J., Eds.; Blackwell Scientific Publications: Oxford, UK, 1987; pp. 115–131. [Google Scholar]
- Bazin, E.; Mathe-Hubert, H.; Facon, B.; Carlier, J.; Ravigné, V. The effect of mating system on invasiveness: Some genetic load may be advantageous when invading new environments. Biol. Invasions 2014, 16, 875–886. [Google Scholar] [CrossRef] [Green Version]
- McDonald, B.A.; Linde, C. Pathogen population genetics, evolutionary potential, and durable resistance. Annu. Rev. Phytopathol. 2002, 40, 349–379. [Google Scholar] [CrossRef] [PubMed]
- Prospero, S.; Rigling, D. Invasion genetics of the chestnut blight fungus Cryphonectria parasitica in Switzerland. Phytopathology 2012, 102, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Pisani, C.; Ploetz, R.C.; Stover, E.; Ritenour, M.; Scully, B. Laurel Wilt in Avocado: Review of an Emerging Disease. Int. J. Plant Biol. Res. 2015, 3, 1043. [Google Scholar]
- Mehrotra, R.S.; Aggarwal, A. Dispersal of plant pathogens. In Plant Pathology, 2nd ed.; Tata McGraw-Hill Publishing Co. Ltd.: New Delhi, India, 2005; pp. 199–211. [Google Scholar]
- Mehrotra, R.S.; Aggarwal, A. Dispersal of plant pathogens. In Fundamentals of Plant Pathology; Tata McGraw-Hill Publishing Co. Ltd.: New Delhi, India, 2013; Chapter 9. [Google Scholar]
- Hengeveld, R. Dynamics of Biological Invasions; Chapman & Hall: London, UK, 1989. [Google Scholar]
- Gladieux, P.; Feurtey, A.; Hood, M.E.; Snirc, A.; Clavel, J.; Dutech, C.; Roy, M.; Giraud, T. The population biology of fungal invasions. Mol. Ecol. 2015, 24, 1969–1986. [Google Scholar] [CrossRef] [PubMed]
- Pyšek, P.; Jarošik, V.; Hulme, P.E.; Kühn, I.; Wild, J.; Arianoutsou, M.; Bacher, S.; Chiron, F.; Didziulis, V.; Essl, F.; et al. Disentangling the role of environmental and human pressures on biological invasions. Proc. Natl. Acad. Sci. USA 2010, 107, 12157–12162. [Google Scholar]
- Keesing, F.; Belden, L.K.; Daszak, P.; Dobson, A.; Harvell, C.D.; Holt, R.D.; Hudson, P.; Jolles, A.; Jones, K.E.; Mitchell, C.E.; et al. Impacts of biodiversity on the emergence and transmission of infectious diseases. Nature 2010, 468, 647–652. [Google Scholar] [CrossRef] [PubMed]
- Ostfeld, R.S.; Keesing, F. Effects of Host Diversity on Infectious Disease. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 157–182. [Google Scholar] [CrossRef]
- Pautasso, M.; Holdenrieder, O.; Stenlid, J. Susceptibility to Fungal Pathogens of Forests Differing in Tree Diversity. In Forest Diversity and Function: Temperate and Boreal Systems, Ecological Studies; Scherer-Lorenzen, M., Körner, C., Schulze, E.-D., Eds.; Springer: New York, NY, USA, 2005; Volume 176, pp. 263–289. [Google Scholar]
- Cleary, M.; Nguyen, D.; Marčiulynienė, D.; Berlin, A.; Vasaitis, R.; Stenlid, J. Friend or foe? Biological and ecological traits of the European ash dieback pathogen Hymenoscyphus fraxineus in its native environment. Sci. Rep. 2016, 6, 21895. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, L.R.; McKinney, L.V.; Hietala, A.M.; Kjær, E.D. The susceptibility of Asian, European and North American Fraxinus species to the ash dieback pathogen Hymenoscyphus fraxineus reflects their phylogenetic history. Eur. J. Forest. Res. 2016. [Google Scholar] [CrossRef]
- McKinney, L.V.; Neilsen, L.R.; Collinge, D.B.; Thomsen, I.M.; Hansen, J.K.; Kjær, E.J. The ash dieback crisis: Genetic variation in resistance can prove a long-term solution. Plant Pathol. 2014, 63, 485–499. [Google Scholar] [CrossRef]
- Hughes, M.A.; Smith, J.A.; Ploetz, R.C.; Kendra, P.E.; Mayfield, A.E., III; Hanula, J.L.; Hulcr, J.; Stelinski, L.L.; Cameron, S.; Riggins, J.J.; et al. Recovery plan for laurel wilt on redbay and other forest species caused by Raffaelea lauricola and disseminated by Xyleborus glabratus. Plant Health Prog. 2015. [Google Scholar] [CrossRef]
- Mitchell, C.E.; Tilman, D.; Groth, J.V. Effects of grassland plant species diversity, abundance, and composition on foliar fungal disease. Ecology 2002, 83, 1713–1726. [Google Scholar] [CrossRef]
- Keesing, F.; Holt, R.D.; Ostfel, R.S. Effects of species diversity on disease risk. Ecol. Lett. 2006, 9, 485–498. [Google Scholar] [CrossRef] [PubMed]
- Haas, S.E.; Cushman, J.H.; Dillon, W.W.; Rank, N.E.; Rizzo, D.M.; Meentemeyer, R.K. Effects of individual, community, and landscape drivers on the dynamics of a wildland forest epidemic. Ecology 2016, 97, 649–660. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, C.; Jolles, A.; Seabloom, E.W.; Power, A.G.; Mitchell, C.E.; Borer, E.T. Non-random biodiversity loss underlies predictable increases in viral disease, prevalence. J. R. Soc. Interface 2014, 11, 20130947. [Google Scholar] [CrossRef] [PubMed]
- Burdon, J.J.; Chilvers, G.A. Host density as a factor in plant disease ecology. Annu. Rev. Phytopathol. 1982, 20, 143–166. [Google Scholar] [CrossRef]
- Wolfe, N.D.; Dunavan, C.P.; Diamond, J. Origins of major human infectious diseases. Nature 2007, 447, 279–283. [Google Scholar] [CrossRef] [PubMed]
- Parker, I.M.; Saunders, S.; Bontrager, M.; Weitz, A.P.; Hendricks, R.; Magarey, R.; Suiter, K.; Gilbert, G.S. Phylogenetic structure and host abundance drive disease pressure in communities. Nature 2015, 520, 542–544. [Google Scholar] [CrossRef] [PubMed]
- Morin, R.S.; Liebhold, A.M.; Tobin, P.C.; Gottschalk, K.W.; Luzader, E. Spread of beech bark disease in the eastern United States and its relationship to regional forest composition. Can. J. For. Res. 2007, 37, 726–736. [Google Scholar] [CrossRef]
- Kasson, M.T.; Livingston, W.H. Relationships among beech bark disease, climate, radial growth response and mortality of American beech in northern Maine, USA. For. Pathol. 2012, 42, 199–212. [Google Scholar] [CrossRef]
- Hatala, J.A.; Dietze, M.C.; Crabtree, R.L.; Kendall, K.; Six, D.; Moorcroft, P.R. An ecosystem-scale model for the spread of a host-specific forest pathogen in the Greater Yellowstone Ecosystem. Ecol. Appl. 2011, 21, 1138–1153. [Google Scholar] [CrossRef] [PubMed]
- Weste, G.; Brown, K.; Kennedy, J.; Walshe, T. Phytophthora cinnamomi infestation—A 24 year study of vegetation change in forests and woodlands of the Grampians, Western Victoria. Aust. J. Bot. 2002, 50, 247–274. [Google Scholar] [CrossRef]
- Davidson, J.M.; Patterson, H.A.; Wickland, A.C.; Fichtner, E.J.; Rizzo, D.M. Forest type influences transmission of Phytophthora ramorum in California oak woodlands. Phytopathology 2001, 101, 492–501. [Google Scholar] [CrossRef] [PubMed]
- Purse, B.V.; Graeser, P.; Searle, K.; Edwards, C.; Harris, C. Challenges in predicting invasive reservoir hosts of emerging pathogens: Mapping Rhododendron ponticum as a foliar host for Phytophthora ramorum and Phytophthora kernoviae in the UK. Biol. Invasions 2013, 15, 529–545. [Google Scholar] [CrossRef]
- Kasson, M.T.; Livingston, W.H. Spatial distribution of Neonectria species associated with beech bark disease in northern Maine. Mycologia 2009, 101, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Park, A.W.; Gubbins, S.; Gilligan, C.A. Extinction times for closed epidemics: The effects of host spatial structure. Ecol. Lett. 2002, 5, 747–755. [Google Scholar] [CrossRef]
- Holdenrieder, O.; Pautasso, M.; Weisberg, P.J.; Lonsdale, D. Tree diseases and landscape processes: The challenge of landscape pathology. TRENDS Ecol. Evol. 2004, 19, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Dwyer, G. On the spatial spread of insect viruses: Theory and experiment. Ecology 1992, 73, 479–494. [Google Scholar] [CrossRef]
- Burdon, J.J.; Jarosz, A.M.; Kirby, G.C. Pattern and patchiness in plant-pathogen interactions: Causes and consequences. Ann. Rev. Ecol. Syst. 1989, 20, 119–136. [Google Scholar] [CrossRef]
- Caraco, T.; Duryea, M.C.; Glavanakov, S.; Maniatty, W.; Szymanski, B.K. Host Spatial Heterogeneity and the Spread of Vector-Borne Infection. Theor. Popul. Biol. 2001, 59, 185–206. [Google Scholar] [CrossRef] [PubMed]
- Broders, K.; Boraks, A.; Barbison, L.; Brown, J.; Boland, G.J. Recent insights into the pandemic disease butternut canker caused by the invasive pathogen Ophiognomonia clavigignenti-juglandacearum. For. Pathol. 2015, 45, 1–8. [Google Scholar]
- Brar, S.; Tsui, C.K.M.; Dhillon, B.; Bergeron, M.-J.; Joly, D.L.; Zambino, P.J.; El-Kassaby, Y.A.; Hamelin, R.C. Colonization History, Host Distribution, Anthropogenic Influence and Landscape Features Shape Populations of White Pine Blister Rust, an Invasive Alien Tree Pathogen. PLos ONE 2015, 10, e0127916. [Google Scholar] [CrossRef] [PubMed]
- Papaïx, J.; Touzeau, S.; Monod, H.; Lannou, C. Can epidemic control be achieved by altering landscape connectivity in agricultural systems? Ecol. Model. 2014, 284, 35–47. [Google Scholar] [CrossRef]
- Real, L.A.; Biek, R. Spatial dynamics and genetics of infectious diseases on heterogeneous landscapes. J. R. Soc. Interface 2007, 4, 935–948. [Google Scholar] [CrossRef] [PubMed]
- Castello, J.D.; Leopold, D.J.; Smallidge, P.J. Pathogens patterns, and processes in forest ecosystems. Bioscience 1995, 45, 16–24. [Google Scholar] [CrossRef]
- Ellis, A.M.; Váklavík, T.; Meentemeyer, R.K. When is connectivity important? A case study of the spatial pattern of sudden oak death. Oikos 2010, 119, 485–493. [Google Scholar] [CrossRef]
- Garnas, J.R.; Auger-Rozenberg, M.-A.; Roques, A.; Bertelsmeier, C.; Wingfield, M.J.; Saccaggi, D.L.; Roy, H.E.; Slippers, B. Complex patterns of global spread in invasive insects: Eco-evolutionary and management consequences. Biol. Invasions 2016, 18, 935–952. [Google Scholar] [CrossRef]
- Dutech, C.; Fabreguettes, O.; Capdevielle, X.; Robin, C. Multiple introductions of divergent genetic lineages in an invasive fungal pathogen, Cryphonectria parasitica, in France. Heredity 2010, 105, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Möykkynen, T.; Capretti, P.; Pukkala, T. Modelling the potential spread of Fusarium circinatum, the causal agent of pitch canker in Europe. Ann. For. Sci. 2015, 72, 169–181. [Google Scholar] [CrossRef]
- Jules, E.S.; Carroll, A.L.; Garcia, A.M.; Steenbock, C.M.; Kauffman, M.J. Host heterogeneity influences the impact of a non-native disease invasion on populations of a foundation tree species. Ecosphere 2014, 5, 1–17. [Google Scholar] [CrossRef]
- Jarosz, A.M.; Burdon, J.J. The effect of small scale environmental changes on disease incidence and severity in a natural plant pathogen interaction. Oecologia 1988, 75, 78–81. [Google Scholar] [CrossRef]
- Smith, J.P.; Hoffman, J.T. Site and stand characteristics related to white pine blister rust in high-elevation forests of Southern Idaho and Western Wyoming. West. N. Am. Nat. 2001, 61, 409–416. [Google Scholar]
- Marçais, B.; Desprez-Loustau, M.-L. European oak powdery mildew: Impact on trees, effects of environmental factors, and potential effects of climate change. Ann. For. Sci. 2014, 71, 633–642. [Google Scholar] [CrossRef]
- Solla, A.; Martín, J.A.; Ouellette, G.B.; Gil, L. Influence of plant age on symptom development in Ulmus minor following inoculation by Ophiostoma novo-ulmi. Plant Dis. 2005, 89, 1035–1040. [Google Scholar] [CrossRef]
- King, K.C.; Lively, C.M. Does genetic diversity limit disease spread in natural host populations? Heredity 2012, 109, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Burdon, R.D. Genetic diversity and disease resistance: Some considerations for research, breeding, and deployment. Can. J. For. Res. 2001, 31, 596–606. [Google Scholar] [CrossRef]
- Garrett, K.A.; Mundt, C.C. Epidemiology in mixed populations. Phytopathology 1999, 89, 984–990. [Google Scholar] [CrossRef] [PubMed]
- Pilet, F.; Chacon, G.; Forbes, G.A.; Adrivon, D. Protection of susceptible potato cultivars against late blight in mixtures increases with decreasing disease pressure. Phytopathology 2006, 96, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Chen, H.; Fan, J.; Wang, Y.; Li, Y.; Chen, J.; Fan, J.X.; Yang, S.; Hu, L.; Leung, H.; et al. Genetic diversity and disease control in rice. Nature 2000, 406, 718–722. [Google Scholar] [CrossRef] [PubMed]
- Barnes, I.; Wingfield, M.J.; Carbone, I.; Kirisits, T.; Wingfield, B.D. Population structure and diversity of an invasive pine needle pathogen reflects anthropogenic activity. Ecol. Evol. 2014, 4, 3642–3661. [Google Scholar] [CrossRef] [PubMed]
- Andjic, V.; Dell, B.; Barber, P.; Hardy, G.; Wingfield, M.; Burgess, T. Plants for planting; indirect evidence for the movement of a serious forest pathogen, Teratosphaeria destructans, in Asia. Eur. J. Plant Pathol. 2011, 131, 49–58. [Google Scholar] [CrossRef]
- Dwyer, G.; Elkinton, J.S.; Buonaccorsi, J.P. Host Heterogeneity in susceptibility and disease dynamics: Tests of a mathematical model. Am. Nat. 1997, 150, 685–707. [Google Scholar] [CrossRef] [PubMed]
- Spielman, D.; Brook, B.W.; Briscoe, D.A.; Frankham, R. Does inbreeding and loss of genetic diversity decrease disease resistance. Conserv. Genet. 2004, 5, 439–448. [Google Scholar] [CrossRef]
- Aegerter, B.J.; Gordon, T.R. Rates of pitch canker induced seedling mortality among Pinus radiate families varying in levels of genetic resistance to Gibberella circinata (anamorph Fusarium circinatum). For. Ecol. Manag. 2006, 235, 14–17. [Google Scholar] [CrossRef]
- Sniezko, R.A. Resistance breeding against nonnative pathogens in forest trees—Current successes in North America. Can. J. Plant Pathol. 2006, 28, S270–S279. [Google Scholar] [CrossRef]
- Ganz, H.H.; Ebert, D. Benefits of host genetic diversity for resistance to infection depend on parasite diversity. Ecology 2010, 91, 1263–1268. [Google Scholar] [CrossRef] [PubMed]
- Freer-Smith, P.H.; Webber, J.F. Tree pests and diseases: The threat to biodiversity and the delivery of ecosystem services. Biodivers. Conserv. 2015, 1–15. [Google Scholar] [CrossRef]
- Heuch, J. What lessons need to be learnt from the outbreak of Ash Dieback Disease, Chalara fraxinea in the United Kingdom? Arboric. J. 2014, 36, 32–44. [Google Scholar] [CrossRef]
- Mitchell, R.J.; Beaton, J.K.; Bellamy, P.E.; Broome, A.; Chetcuti, J.; Eaton, S.; Ellis, C.J.; Gimona, A.; Harmer, R.; Hester, A.J.; et al. Ash dieback in the UK: A review of the ecological and conservation implications and potential management options. Biol. Conserv. 2014, 175, 95–109. [Google Scholar] [CrossRef]
- Gross, A.; Holdenrieder, O.; Pautasso, M.; Queloz, V.; Sieber, T.N. Hymenoscyphus pseudoalbidus, the causal agent of European ash dieback. Mol. Plant Pathol. 2014, 15, 5–21. [Google Scholar] [CrossRef] [PubMed]
- Børja, I.; Timmermann, V.; Hietala, A.M.; Tollefsrud, M.M.; Nagy, N.E.; Vivian-Smith, A.; Cross, H.; Sønstebø, H.J.; Myking, T.; Solheim, H. Ash dieback in Norway—Current situation. In Dieback of European Ash (Fraxinus spp.)—Consequences and Guidelines for Sustainable Management; Vasaitis, R., Enderle, R., Eds.; SLU Service/Repro: Uppsala, Sweden, 2017; pp. 166–175. [Google Scholar]
- Cleary, M.; Nguyen, D.; Stener, L.-G.; Stenlid, J.; Skovsgaard, J.-P. Ash and ash dieback in Sweden: A review of disease history, current status, pathogen and host dynamics, host tolerance and management options in forests and landscapes. In Dieback of European Ash (Fraxinus spp.)—Consequences and Guidelines for Sustainable Management; Vasaitis, R., Enderle, R., Eds.; SLU Service/Repro: Uppsala, Sweden, 2017; pp. 195–208. [Google Scholar]
- Roane, M.K.; Griffin, G.J.; Elkins, J.R. Chestnut Blight, Other Endothial Diseases, and the Genus Endothia; American Phytopathological Society: St. Paul, MN, USA, 1986. [Google Scholar]
- Braun, E.L. Deciduous Forests of Eastern North America; Hafner: New York, NY, USA, 1950. [Google Scholar]
- Jacobs, D.F. Toward development of silvical strategies for forest restoration of American chestnut (Castanea dentata) using blight-resistant hybrids. Biol. Conserv. 2007, 137, 497–506. [Google Scholar] [CrossRef]
- Graves, A.H. Relative blight resistance in species and hybrids of Castanea. Phytopathology 1950, 40, 1125–1131. [Google Scholar]
- Dane, F.; Lang, P.; Huang, H.; Fu, Y. Intercontinental genetic divergence of Castanea species in eastern Asia and eastern North America. Heredity 2003, 91, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Dutech, C.; Barres, B.; Bridier, J.; Robin, C.; Milgroom, M.G.; Ravigne, V. The chestnut blight fungus world tour: Successive introduction events from diverse origins in an invasive plant fungal pathogen. Mol. Ecol. 2012, 21, 3931–3946. [Google Scholar] [CrossRef] [PubMed]
- Heiniger, U.; Rigling, D. Biological control of chestnut blight in Europe. Annu. Rev. Phytopathol. 1994, 32, 581–599. [Google Scholar] [CrossRef]
- Rigling, D.; Prospero, S. Cryphonectria parasitica, the causal agent of chestnut blight: Invasion history, population biology and disease control. Mol. Plant Pathol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Milgroom, M.G.; Cortesi, P. Biological control of chestnut blight with hypovirulence: A critical analysis. Annu. Rev. Phytopathol. 2004, 42, 311–338. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.S.; Choi, G.H.; Nuss, D.L. Mitotic stability and nuclear inheritance of integrated viral cDNA in engineered hypovirulent strains of the chestnut blight fungus. EMBO J. 1993, 12, 2991–2998. [Google Scholar] [PubMed]
- Zhang, D.X.; Nuss, D.L. Engineering super mycovirus donor strains of chestnut blight fungus by systematic disruption of multilocus vic genes. Proc. Natl. Acad. Sci. USA 2016, 113, 2062–2067. [Google Scholar] [CrossRef] [PubMed]
- Diskin, M.; Steiner, K.C.; Hebard, F.V. Recovery of American chestnut characteristics following hybridization and backcross breeding to restore blight-ravaged Castanea dentata. For. Ecol. Manag. 2006, 223, 439–447. [Google Scholar] [CrossRef]
- Newhouse, A.E.; Polin-McGuigan, L.D.; Baier, K.A.; Valletta, K.E.; Rottmann, W.H.; Tschaplinski, T.J.; Maynard, C.A.; Powell, W.A. Transgenic American chestnuts show enhanced blight resistance and transmit the trait to T1 progeny. Plant Sci. 2014, 228, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Hosoya, T.; Otani, Y.; Furuya, K. Materials for the fungus flora of Japan. Trans. Mycol. Soc. Jpn. 1993, 34, 429–432. [Google Scholar]
- Drenkhan, R.; Hanson, M. New host species for Chalara fraxinea. New Dis. Rep. 2010, 22, 16. [Google Scholar] [CrossRef]
- Cleary, M.R.; Arhipova, N.; Gaitnieks, T.; Stenlid, J.; Vasaitis, R. Natural infection of Fraxinus excelsior seeds by Chalara fraxinea. For. Pathol. 2013, 43, 83–85. [Google Scholar]
- Luchi, N.; Montecchio, L.; Santini, A. Situation with ash in Italy: Stand characteristics, health condition, ongoing work and research needs. In Interim Report from Chalara fraxinea, FRAXBACK Meeting in Vilnius, 13–14 November 2012; Mainprize, N., Hendry, S., Weir, J., Eds.; Forestry Commission: Bristol, UK, 2012; pp. 25–26. [Google Scholar]
- Zhao, Y.-J.; Hosoya, T.; Baral, H.-O.; Hosaka, K.; Kakishima, M. Hymenoscyphus pseudoalbidus, the correct name for Lambertella albida reported from Japan. MycoTaxon 2013, 122, 25–41. [Google Scholar] [CrossRef]
- Chandelier, A.; Gerarts, F.; San Martin, G.; Herman, M.; Delahaye, L. Temporal evolution of collar lesions associated with ash dieback and the occurrence of Armillaria in Belgian forests. For. Pathol. 2016. [Google Scholar] [CrossRef]
- Chandelier, A.; Helson, M.; Dvorak, M.; Gischer, F. Detection and quantification of airborne inoculum of Hymenoscyphus pseudoalbidus using real-time PCR assays. Plant Pathol. 2014, 63, 1296–1305. [Google Scholar] [CrossRef]
- Zheng, H.-D.; Zhuang, W.-Y. Hymenoscyphus albidoides sp. nov. and H. pseudoalbidus from China. Mycol. Prog. 2014, 13, 625–638. [Google Scholar] [CrossRef]
- Han, J.-G.; Shrestha, B.; Hosoya, T.; Lee, K.-H.; Sung, G.-H.; Shin, H.-D. First report of the ash dieback pathogen Hymensocyphus fraxineus in Korea. Mycobiology 2014, 42, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Kirisits, T.; Schwanda, K. First definite report of natural infection of Fraxinus ornus by Hymenoscyphus fraxineus. For. Pathol. 2015, 45, 430–432. [Google Scholar]
- Marçais, B.; Husson, C.; Godart, L.; Caël, O. Influence of site and stand factors on Hymenoscyphus fraxineus-induced basal lesion. Plant Pathol. 2016. [Google Scholar] [CrossRef]
- Park, A.W.; Gubbins, S.; Gilligan, C.A. Invasion persistence of plant parasites in a spatially structured host population. Oikos 2001, 94, 162–174. [Google Scholar] [CrossRef]
- Jeger, M.J.; Pautasso, M.; Holdenrieder, O.; Shaw, M.K. Modelling disease spread and control in networks: Implications for plant sciences. New Phytol. 2007, 114, 279–297. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, J.R.; Preisser, E.L.; Fitzpatrick, M.C. Modeling the spread of invasive species using dynamic network models. Biol. Invasions 2014, 16, 949–960. [Google Scholar] [CrossRef]
- Moslonka-Lefebvre, M.; Finley, A.; Dorigatti, I.; Dehnen-Schmutz, K.; Harwood, T.; Jeger, M.J.; Xu, X.; Holdenrieder, O.; Pautasso, M. Networks in plant epidemiology: From genes to landscapes, countries, and continents. Phytopathology 2011, 101, 392–403. [Google Scholar] [CrossRef] [PubMed]
- Biek, R.; Real, L.A. The landscape genetics of infectious disease emergence and spread. Mol. Ecol. 2010, 19, 3515–3531. [Google Scholar] [CrossRef] [PubMed]
- Desprez-Loustau, M.-L. The alien fungi of Europe. In Handbook of Alien Species in Europe, DAISIE. Invading Nature, Series in Invasion Ecology; Drake, J.A., Ed.; Springer: Berlin, Germany, 2009; Volume 3, pp. 15–28. [Google Scholar]
- Horie, T.; Haight, R.G.; Homans, F.R.; Venette, R.C. Optimal strategies for the surveillance and control of forest pathogens: A case study with oak wilt. Ecol. Econ. 2013, 86, 78–85. [Google Scholar] [CrossRef]
- Liebhold, A.M.; Tobin, P.C. Exploiting the Achilles Heels of Pest Invasions: Allee Effects, Stratified Dispersal and Management of Forest Insect Establishment and Spread. N. Zeal. J. For. Sci. 2010, 40, S25–S33. [Google Scholar]
- Homans, F.; Horie, T. Optimal detection strategies for an established invasive pest. Ecol. Econ. 2011, 70, 1129–1138. [Google Scholar] [CrossRef]
- Hänfling, B.; Kollmann, J. An evolutionary perspective of biological invasions. TRENDS Ecol. Evol. 2002, 17, 545–546. [Google Scholar] [CrossRef]
- Lee, C.E. Evolutionary genetics of invasive species. TRENDS Ecol. Evol. 2002, 17, 386–391. [Google Scholar] [CrossRef]
- Woolhouse, M.E.J.; Haydon, D.T.; Antia, R. Emerging pathogens: The epidemiology and evolution of species jumps TRENDS Ecol. Evol. 2005, 20, 238–244. [Google Scholar]
- Desprez-Loustau, M.-L.; Courtecuisse, R.; Robin, C.; Husson, C.; Moreau, P.-A.; Blancard, D.; Selosse, M.-A.; Lung-Escarmant, B.; Piou, D.; Sache, I. Species diversity and drivers of spread of alien fungi (sensu lato) in Europe with a particular focus on France. Biol. Invasions 2010, 12, 157–172. [Google Scholar] [CrossRef]
Figure 1.
Local, regional, and continental-scale factors governing the spread and impact of invasive forest pathogens post-introduction and establishment are affected by three main factors: host diversity, host connectivity, and host susceptibility. Following arrival and establishment, invasiveness is inherently affected by organismal traits (mating system, reproduction type, and dispersal mechanisms). Host diversity is mainly affected by plant species richness (density/composition) creating a dilution effect of pathogen impacts on the ecosystem. Non-hosts, competent hosts, and less competent hosts will have variable effects on their ability to intercept inoculum and subsequently reduce pathogen spread. Host connectivity is largely influenced by the distribution of available host species; more or less aggregated. Spatial heterogeneity of hosts becomes important for vector-induced pathogens. Variations in landscape structure (topography, natural geographic/environmental barriers, forest fragmentation) will largely influence spread dynamics on the landscape level. Host susceptibility to invasive pathogens is influenced by physical traits (size, age, morphology), the random presence of other (potentially antagonistic) organisms, environmental and site factors, and host genetic background. Intraspecific genetic diversity (mixtures of host genotypes) offers the best insurance against invasive pathogens through a dilution effect on inoculum production/deposition and the likelihood that some hosts will possess effective mechanisms to resist or minimize damage caused by invasives. Evolutionary and environmental factors, as well as continuous pressures caused by human activity will influence spread dynamics over time. Understanding how host variability is affected by host diversity, connectivity, and susceptibility will improve our ability to predict disease spread on the landscape and potential consequences to ecosystem services.
Figure 1.
Local, regional, and continental-scale factors governing the spread and impact of invasive forest pathogens post-introduction and establishment are affected by three main factors: host diversity, host connectivity, and host susceptibility. Following arrival and establishment, invasiveness is inherently affected by organismal traits (mating system, reproduction type, and dispersal mechanisms). Host diversity is mainly affected by plant species richness (density/composition) creating a dilution effect of pathogen impacts on the ecosystem. Non-hosts, competent hosts, and less competent hosts will have variable effects on their ability to intercept inoculum and subsequently reduce pathogen spread. Host connectivity is largely influenced by the distribution of available host species; more or less aggregated. Spatial heterogeneity of hosts becomes important for vector-induced pathogens. Variations in landscape structure (topography, natural geographic/environmental barriers, forest fragmentation) will largely influence spread dynamics on the landscape level. Host susceptibility to invasive pathogens is influenced by physical traits (size, age, morphology), the random presence of other (potentially antagonistic) organisms, environmental and site factors, and host genetic background. Intraspecific genetic diversity (mixtures of host genotypes) offers the best insurance against invasive pathogens through a dilution effect on inoculum production/deposition and the likelihood that some hosts will possess effective mechanisms to resist or minimize damage caused by invasives. Evolutionary and environmental factors, as well as continuous pressures caused by human activity will influence spread dynamics over time. Understanding how host variability is affected by host diversity, connectivity, and susceptibility will improve our ability to predict disease spread on the landscape and potential consequences to ecosystem services.


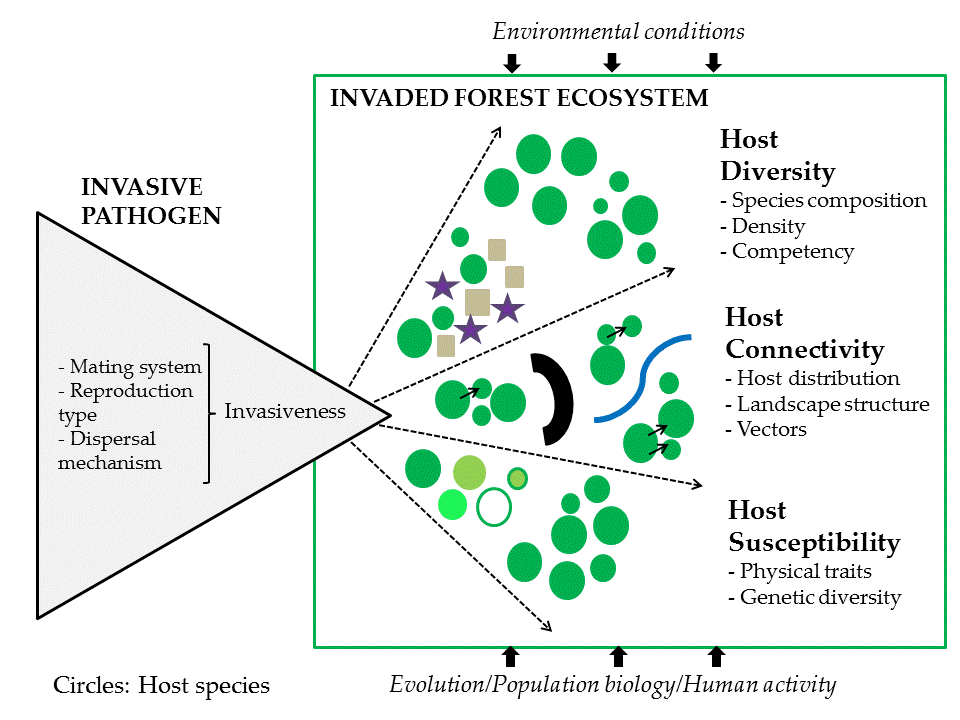{kind=link}
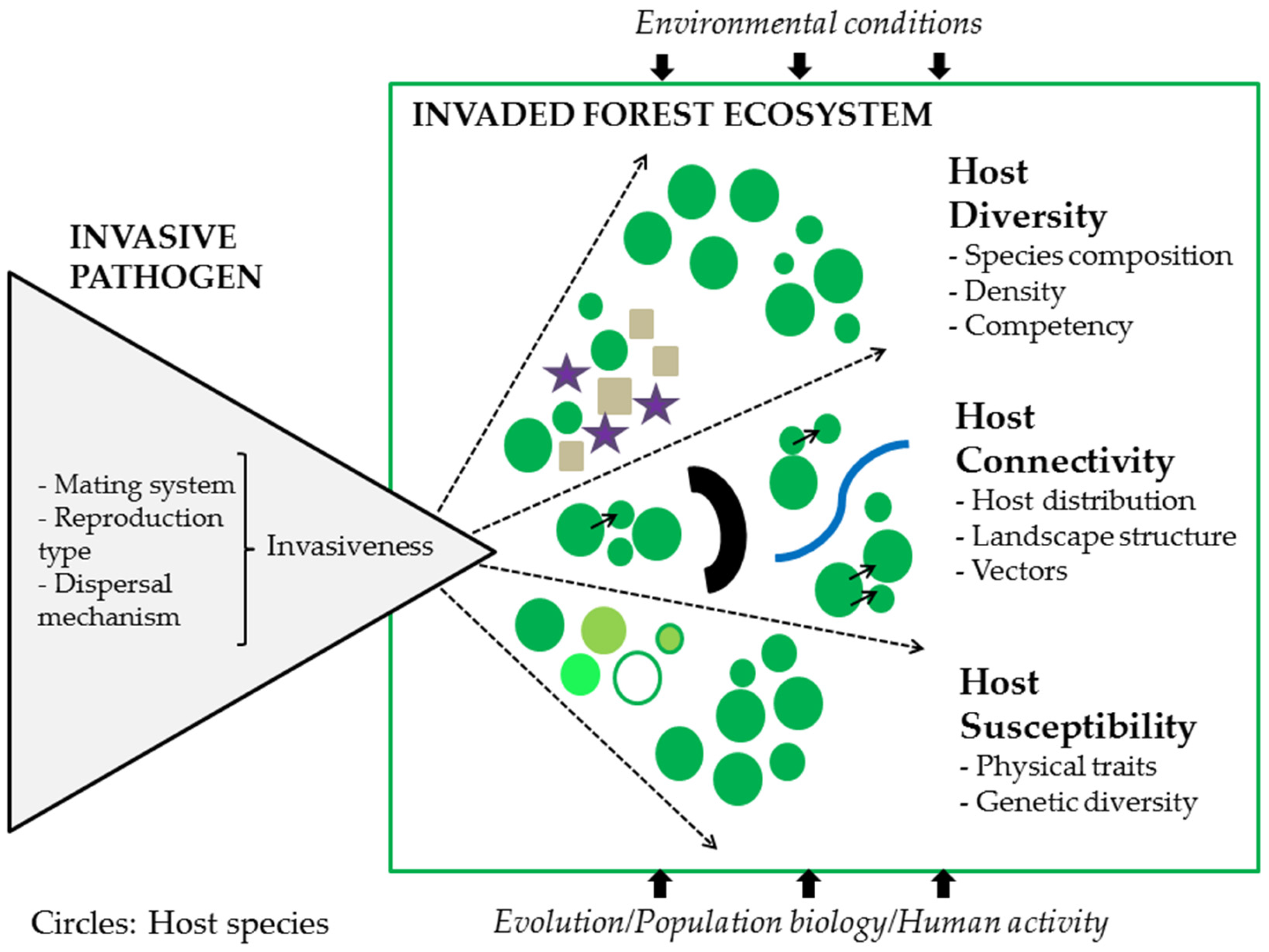{kind=link}
| Mechanism | Description |
|---|---|
| Autonomous transmission | This dispersal mechanism is characterized by continuous and persistent growth of hyphal strands that can migrate independently through the soil from plant to plant, quite characteristic of soil fungi (e.g., Armillaria spp.). Dispersal can range from a few cm to several meters per year. |
| Wind | Most fungal pathogens that produce spores or conidia externally on host surfaces are easily carried by wind currents (e.g., downy and powdery mildews, rust fungi). Fungal spores behave as inert particles, with terminal velocities ranging from about 0.05 to 2.5 cm per second, with the larger spores falling more rapidly than smaller ones. Turbulence redistributes spores and affects their progressive dilution with increasing distance from its source [43]. With normal wind and turbulence conditions, spores can travel large distances (from several hundred meters to kilometers) |
| Water | Except in the case of streams or rivers that may carry inoculum, water is usually less effective than wind for long-distance dissemination. Rain splash or splatter during heavy rains can locally distribute inoculum on or around the same or neighboring plants. Similarly, rain or irrigation water that moves either through the soil or on the soil surface can disseminate pathogen propagules. |
| Vectors | Some plant pathogens cannot be directly transferred from one plant to another and require a completely unrelated species to act as a vector. Many insects have piercing and sucking mouthparts that penetrate the plant surface and facilitate the transmission and inoculation of host plants (e.g., Dutch elm disease caused by Ophiostoma ulmi or O. novo-ulmi growing within the egg galleries of Scolytus bark beetles, contaminating emerging adults; Laurel wilt caused by Raffaelea lauricola transmitted by species of ambrosia beetles). Thus, vector-transmitted pathogens are usually transferred to the host with great efficiency and play a major role in the infection lifecycle. Some insects cause wounding of plant tissue through which plant pathogens can enter secondarily. Other vectors of pathogens may include nematodes or mammals that may transmit diseases both externally and internally. Most vectors of forest pathogens are usually, but not always, insects, and are sometimes referred to as alternate hosts or as having ‘hitchhiking’ dispersal. Hitchhiking dispersal is favored by typical fungal features such as inconspicuousness, and the production of numerous small propagules [46]. For vector-dependent fungi, if no alternate host exists, the infection cycle is broken. |
| Human | Plant diseases are often dispersed through human-mediated, extra-range dispersal typically through transportation of infected propagative material (e.g., seed, nursery stock, timber, plant products, or soil), or through mishandling or contamination of healthy plants or plant parts during cultivation practices. Organismal spread may be complicated by multiple introductions (genotypes) from multiple sources to multiple locations [7,47]. |
Table 2.
Main characteristics of the two invasive forest pathogens causing ash dieback (Hymenoscyphus fraxineus) and chestnut blight (Cryphonectria parasitica).
| Common Name | Ash Dieback 1 | Chestnut Blight 2 |
|---|---|---|
| Causal agent | Hymenoscyphus fraxineus (T. Kowalski) Baral, Queloz, Hosoya, comb. nov. (Family Helotiaceae, Order Helotiales, Class Leotiomycetes) | Cryphonectria parasitica (Murr.) Barr (Family Cryphonetriaceae, Order Diaporthales, Class Sordariomycetes) |
| Host species | Major: genus Fraxinus, in particular European ash (F. excelsior), Narrow-leafed ash (F. angustifolia); Black ash (F. nigra); Green ash (F. pennsylvanica); White ash (F. americana); Tianshan ash (F. sogdiana), and Blue ash (F. quadrangulata) Minor 3: Manna ash (F. ornus); Chinese ash (F. chinensis), Manchurian ash (F. mandschurica), Texas ash (F. albicans), Oregon ash (F. latifolia), Spaeth’s ash (F. platypoda), Pumpkin ash (F. profunda), and Velvet ash (F. veluntina). | Major: genus Castanea, in particular American chestnut (C. dentata), European chestnut (C. sativa), Japanese chestnut (C. crenata), and Chinese chestnut (C. mollissima); Minor (incidental): Quercus spp., Acer spp., Carpinus betulus, Castanea pumila |
| Symptoms | Necrotic lesions on leaflets expanding preferentially along leaf veins, and on leaf rachises; leaf wilting; premature leaf abscission; necrosis of buds, perennial necrotic lesions (cankers) on the bark of twigs, branches, and stem; brown discoloration of xylem; dieback of crown; prolific formation of epicormics shoots; basal stem necrosis | Perennial necrotic lesions (cankers) on the bark of above-ground woody parts (stem, branches) of host plants. The plant part distal to the infection point may wilt. |
| Spread mechanism | Sexual spores (ascospores) for short- (local), medium-, and potentially long-distance spread; Over long distances via movement of latently infected plants or plant material | Over short distances mainly via splash dispersed asexual spores (conidia); Over long distances via sexual ascospores or latently infected plants or plant material. |
| Mating system | Random mating, heterothallic, outcrossing | Mixed with outcrossing and self-fertilization occurring at variable frequencies. |
| Native range | Eastern Asia (China, Korea, Japan, Far East Russia) | Eastern Asia (China, Japan, Korea). |
| Invaded range | Europe | North America, Europe. |
| First detection | Early 1990s (Lithuania, Poland) | 1904 (North America), 1938 (Europe). |
| Introduction pathway | Primary pathway for introduction through nursery stock of latently infected plants (e.g., Manchurian ash F. mandshurica) for planting | Most likely infected plants for planting (probably Castanea crenata). |
| Primary dispersal pathway | Wind-dispersed spores (seasonal—between June and September); movement of infected plants or plant material. | Spores (spontaneous dispersal), infected plant material (e.g., plants for planting, wood with bark). |
| Mean dispersal rates | 30 km per year (Norway) 50–60 km per year (Italy) 75 km per year (from east Poland to Switzerland) | In North America, 37 km per year. In Europe (Italy), 29 km per year. |
| Control | Breeding for resistance through traditional screening and selection of disease tolerant genotypes from wild populations. Chemical and biological treatments have been tried on an experimental basis with varied efficacy. | North America: breeding for resistance by backcrossing blight-resistant Chinese chestnut into the American chestnut genome; Transgenic fungal strains and chestnut trees. Europe: biocontrol by a mycovirus in the family Hypoviridae (Cryphonectria hypovirus 1, CHV-1) which reduces both virulence and sporulation of the infected fungal strain (phenomenon called hypovirulence); Hybrids between C. sativa and C. crenata. |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Prospero, S.; Cleary, M. Effects of Host Variability on the Spread of Invasive Forest Diseases. Forests 2017, 8, 80. https://doi.org/10.3390/f8030080
AMA Style
Prospero S, Cleary M. Effects of Host Variability on the Spread of Invasive Forest Diseases. Forests. 2017; 8(3):80. https://doi.org/10.3390/f8030080
Chicago/Turabian StyleProspero, Simone, and Michelle Cleary. 2017. "Effects of Host Variability on the Spread of Invasive Forest Diseases" Forests 8, no. 3: 80. https://doi.org/10.3390/f8030080
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.