Tree Species Identity Shapes Earthworm Communities

 , ,
, ,
Abstract
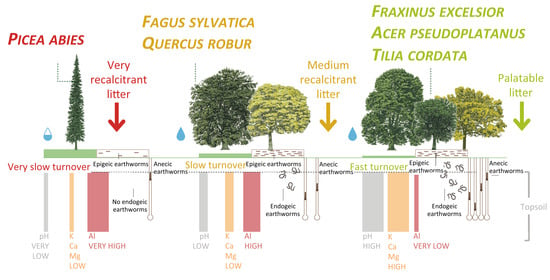
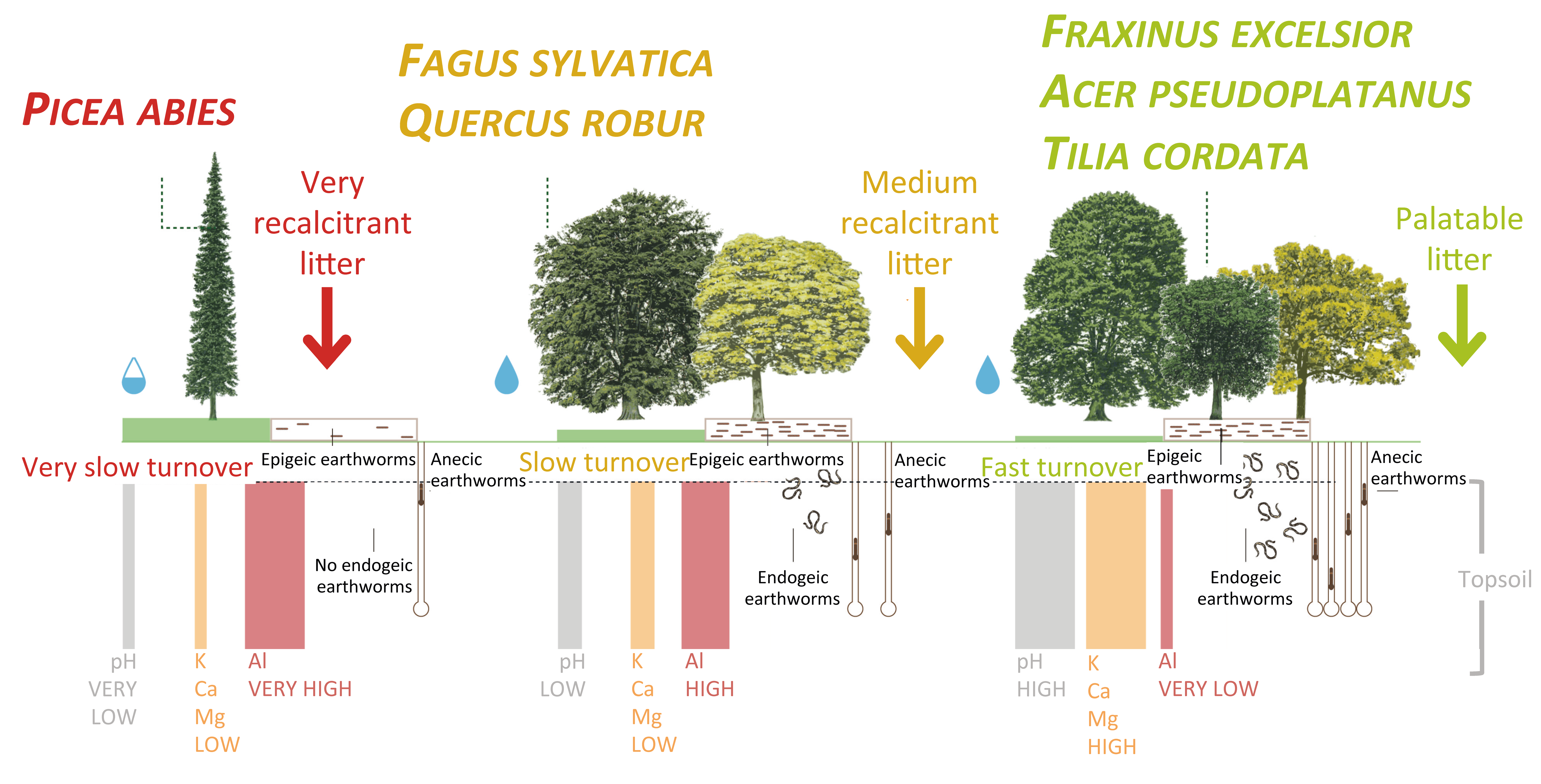
:
1. Introduction
2. Materials and Methods
2.1. Common Garden
2.2. Soil Sampling and Analyses
2.3. Sampling and Analyses of Litter and Forest Floor
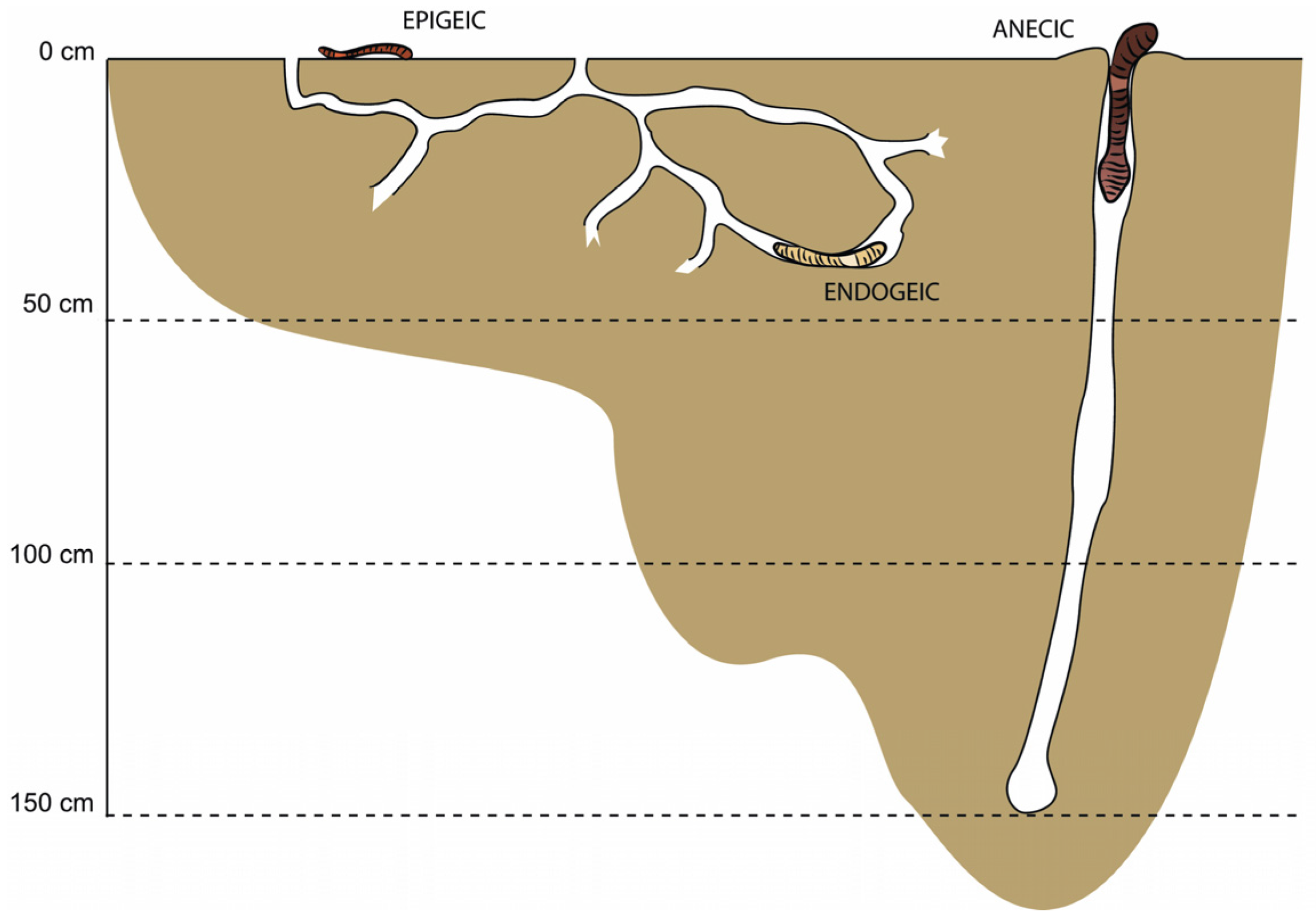
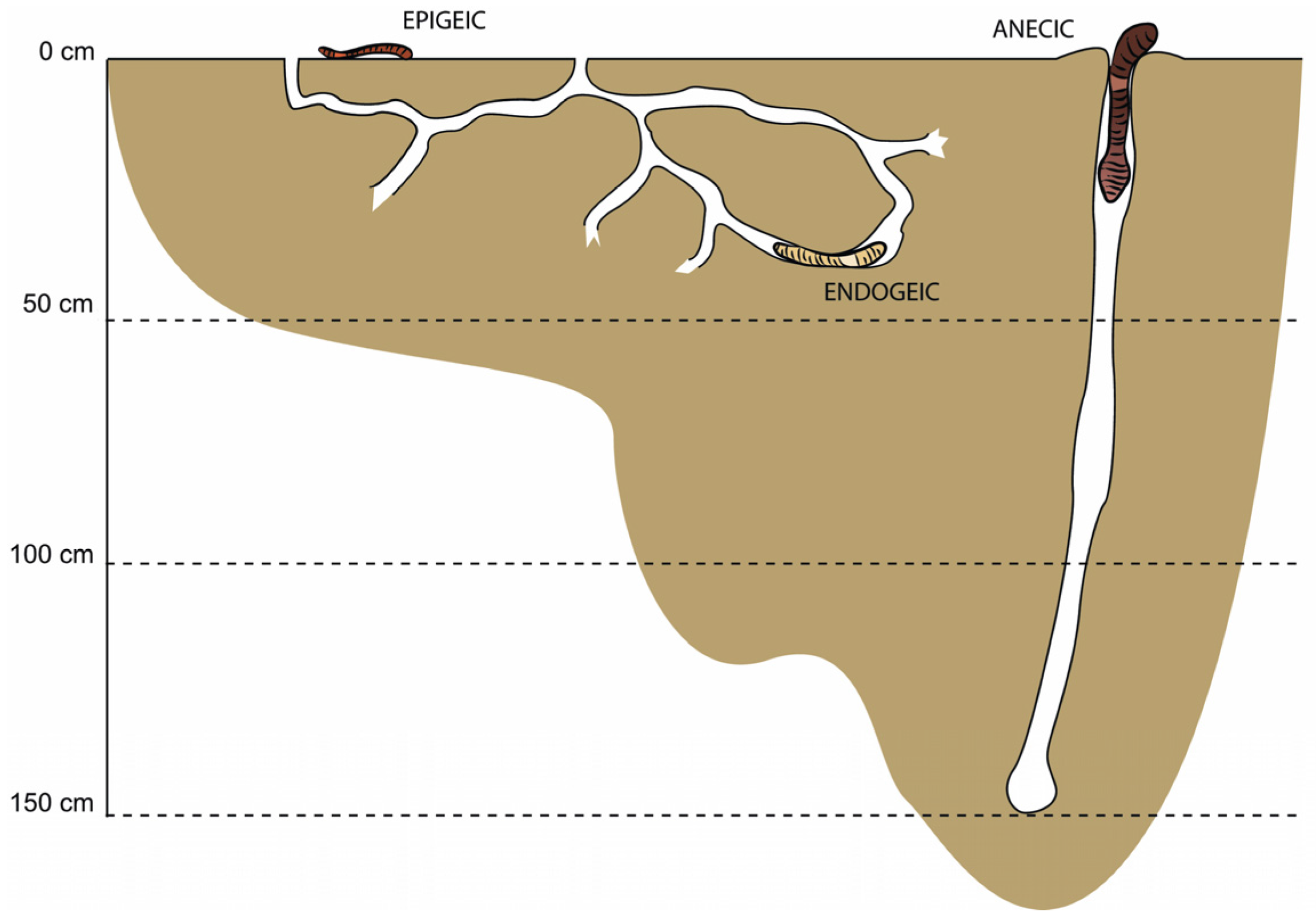
2.4. Earthworm Sampling and Identification
2.5. Calculations
2.5.1. Handling of Missing Data
2.5.2. Forest Floor Turnover Rate
2.6. Data Analyses
2.6.1. Tree Species Effect
2.6.2. Links between Litter Quality, Soil Quality and Earthworms
3. Results
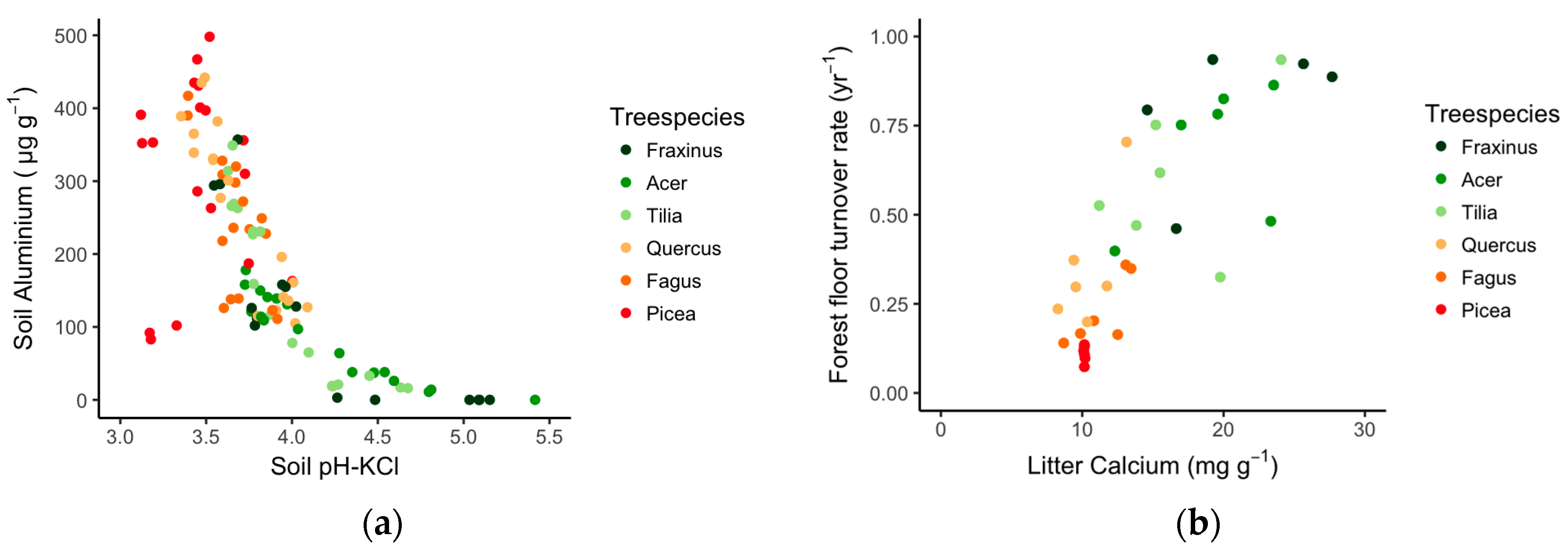
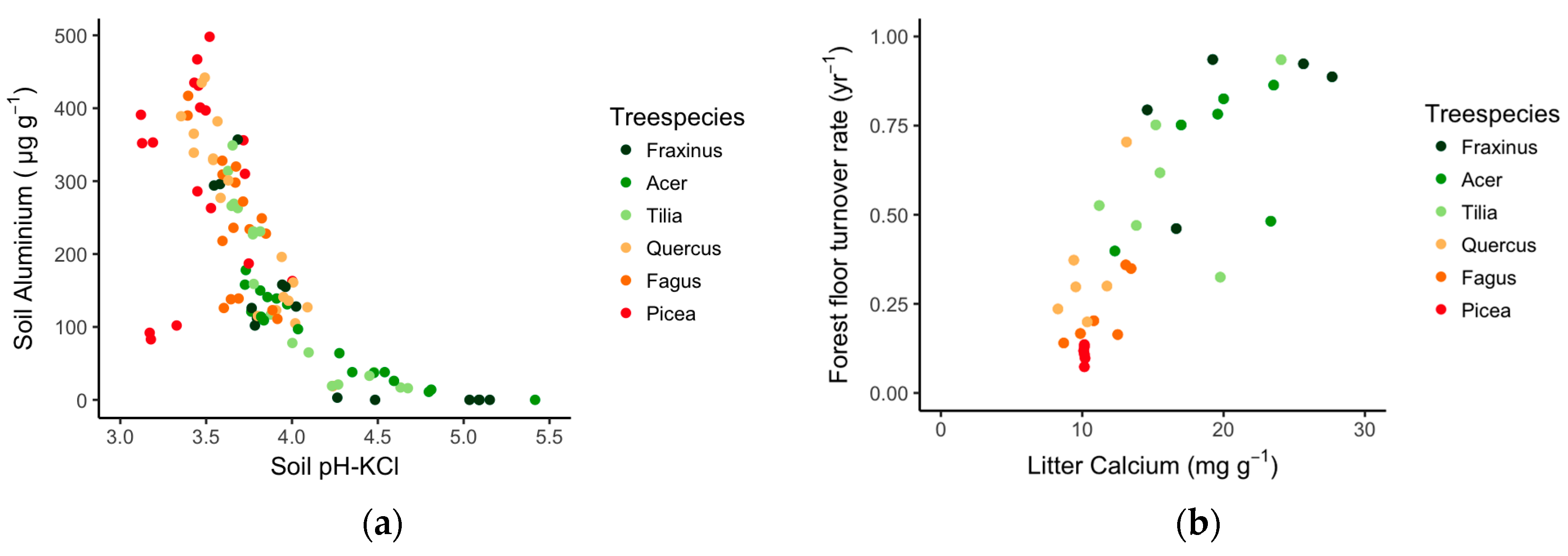
3.1. The Tree Species Effect on Soil Properties and Litter Quality
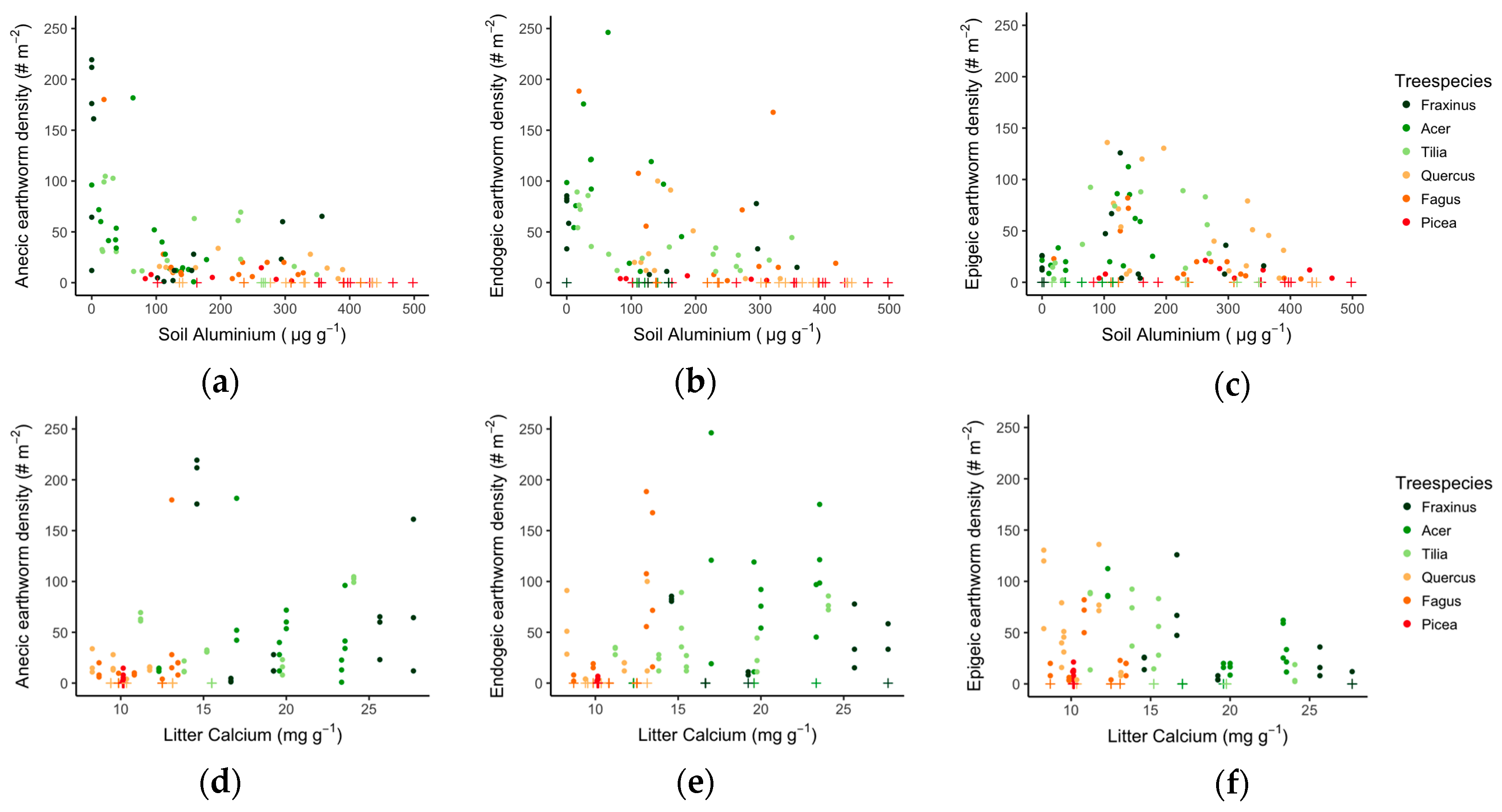
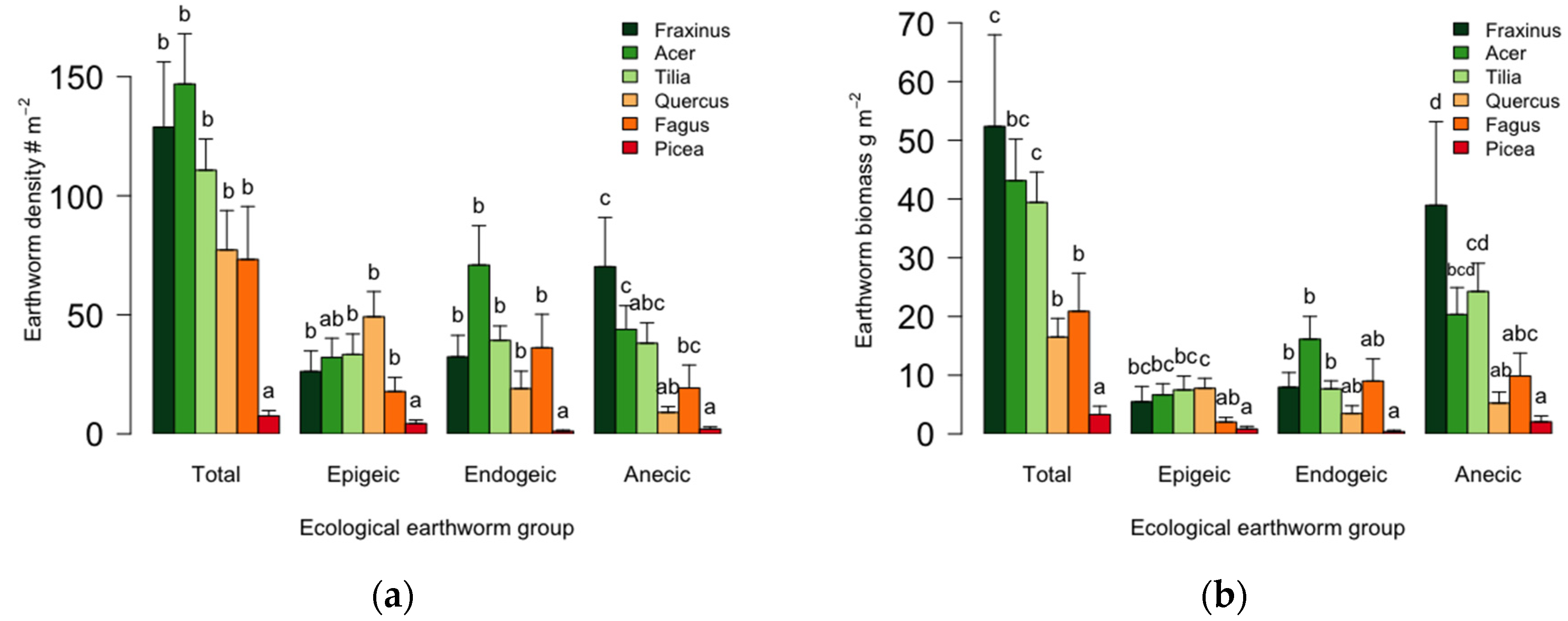
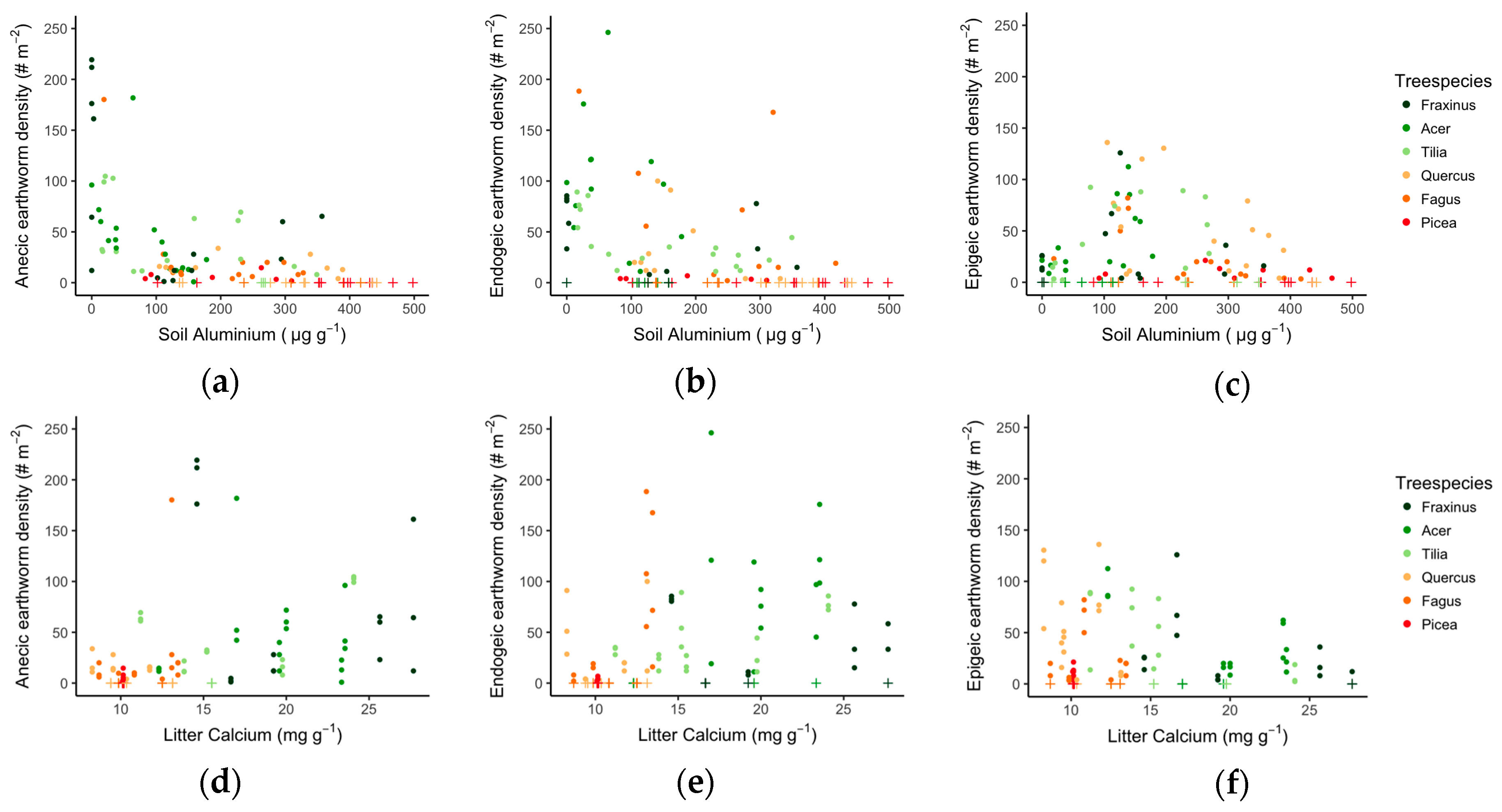
3.2. Tree Species Effect on Earthworm Populations
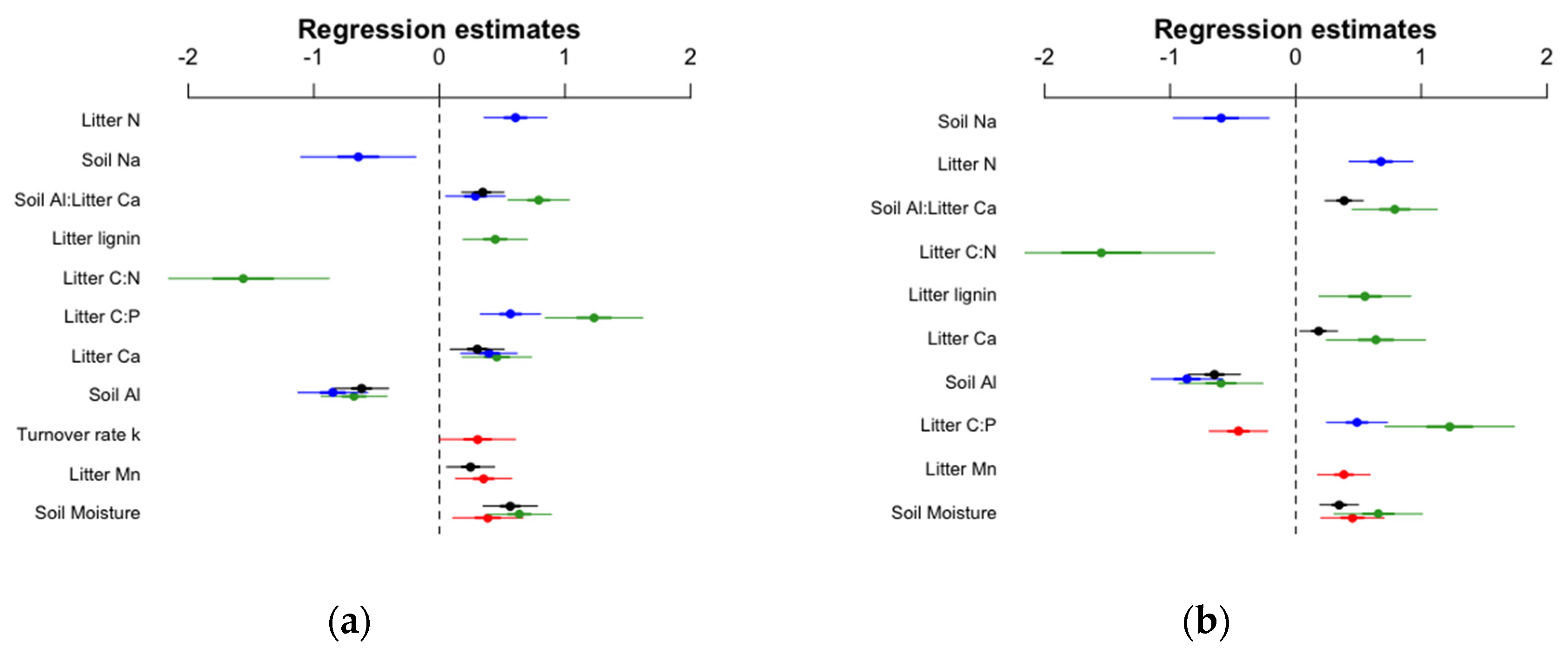
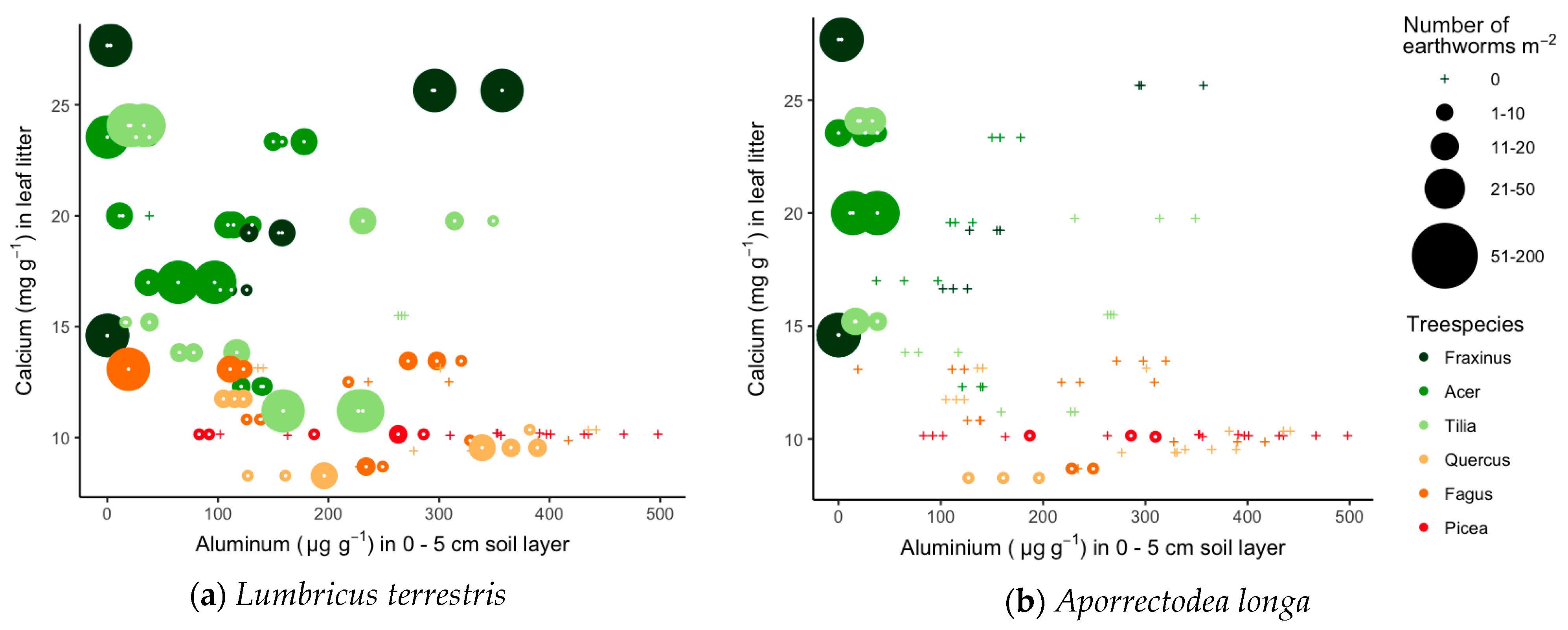
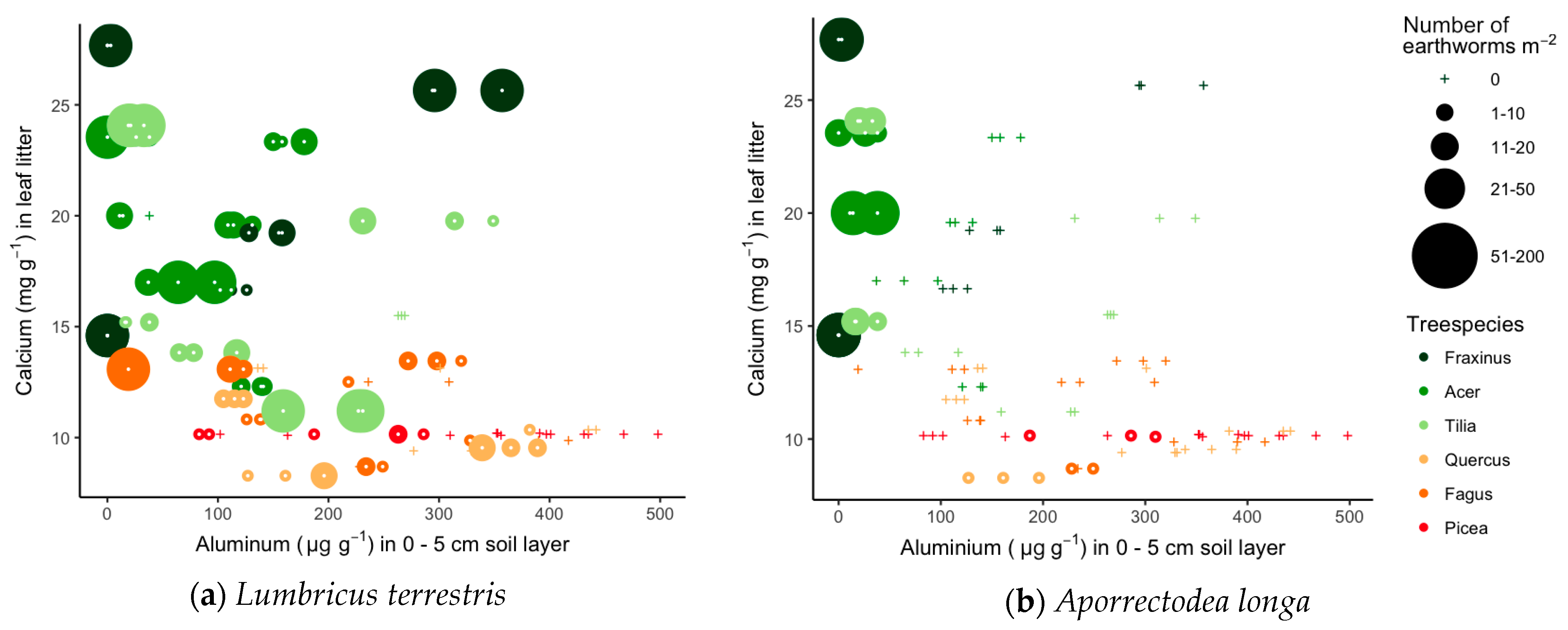
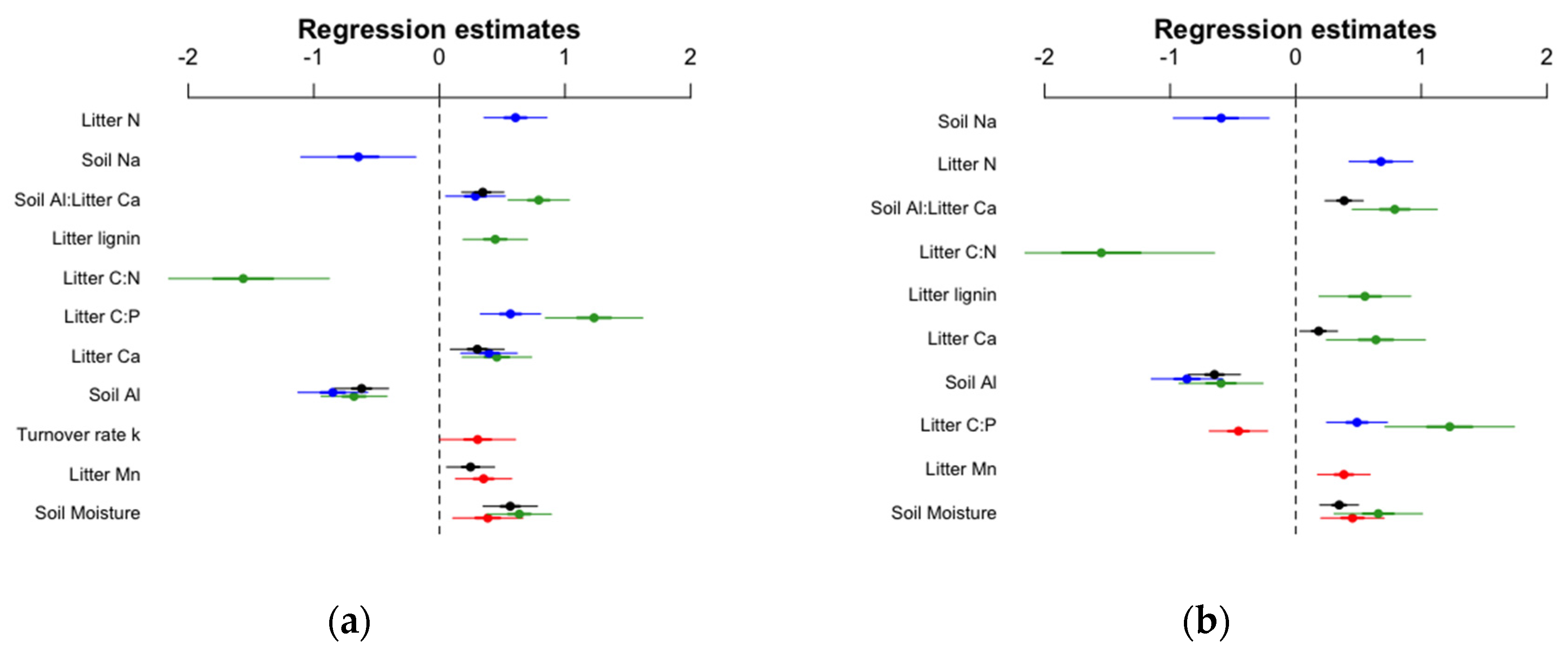
3.3. Links between Soil Quality, Litter Quality and Earthworm Communities
4. Discussion and Conclusions
Acknowledgments
Author Contributions
Conflicts of interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Longitude, Latitude |
|---|---|
| Kragelund | 56°10′N, 9°25′E |
| Mattrup | 55°57′N, 9°38′E |
| Odsherred | 55°50′N, 11°42′E |
| Vallø | 55°25′N, 12°03′E |
| Viemose | 55°01′N, 12°09′E |
| Wedellsborg | 55°24′N, 9°52′E |
Appendix B
| Response Variable | Predictor Variables in Model | AICc | ΔAICc | Weight |
|---|---|---|---|---|
| Earthworm density | ||||
| Total | null model | 1125 | 64 | <0.001 |
| Soil-Al | 1072 | 11 | 0.002 | |
| Soil-Al + Litter-Ca | 1074 | 13 | <0.001 | |
| Soil-Al + Litter-Ca + Soil-Al:Litter-Ca | 1064 | 4 | 0.11 | |
| Soil-Al + Litter-Ca + Soil-Al:Litter-Ca + Litter-Mn | 1063 | 2 | 0.25 | |
| Soil-Al + Litter-Ca + Soil-Al:Litter-Ca + Litter-Mn + Soil Moisture | 1061 | 0 | 0.64 | |
| Anecic | null model | 850 | 84 | <0.001 |
| Soil-Al | 787 | 21 | <0.001 | |
| Soil-Al + Litter-N | 786 | 20 | <0.001 | |
| Soil-Al + Litter-N + Litter-C:P | 778 | 13 | 0.0014 | |
| Soil-Al + Litter-N + Litter-C:P + Soil-Na | 770 | 5 | 0.08 | |
| Soil-Al + Litter-N + Litter-C:P + Soil-Na + Litter-Ca | 769 | 3 | 0.17 | |
| Soil-Al + Litter-N + Litter-C:P + Soil-Na + Litter-Ca + Soil-Al:Litter-Ca | 766 | 0 | 0.75 | |
| Endogeic | null model | 800 | 75 | <0.001 |
| Soil-Al | 788 | 63 | <0.001 | |
| Soil-Al +Soil-Al:Litter-Ca | 784 | 60 | <0.001 | |
| Soil-Al + Soil-Al:Litter-Ca + Litter-Ca | 781 | 57 | <0.001 | |
| Soil-Al + Soil-Al:Litter-Ca + Litter-Ca + Soil Moisture | 750 | 26 | <0.001 | |
| Soil-Al + Litter-Ca + Soil-Al:Litter-Ca + Soil Moisture + Litter-C:P | 740 | 14 | <0.001 | |
| Soil-Al + Litter-Ca + Soil-Al:Litter-Ca + Soil Moisture + Litter-C:P + Litter-C:N | 735 | 10 | 0.0058 | |
| Soil-Al + Litter-Ca + Soil-Al:Litter-Ca + Soil Moisture + Litter-C:P + Litter-C:N + Litter-lignin | 725 | 0 | 0.99 | |
| Epigeic | null model | 817 | 13 | 0.0012 |
| Soil Moisture | 810 | 6 | 0.036 | |
| Soil Moisture + Litter-Mn | 806 | 2 | 0.28 | |
| Soil Moisture + Litter-Mn + Forest floor turnover rate | 804 | 0 | 0.68 | |
| Earthworm biomass | ||||
| Total | null model | 902 | 79 | <0.001 |
| Litter-Ca | 874 | 52 | <0.001 | |
| Litter-Ca +Soil-Al:Litter-Ca | 869 | 47 | <0.001 | |
| Litter-Ca + Soil-Al:Litter-Ca + Soil-Al | 833 | 10 | 0.006 | |
| Litter-Ca + Soil-Al:Litter-Ca + Soil-Al + Soil Moisture | 822 | 0 | 0.99 | |
| Anecic | null model | 739 | 77 | <0.001 |
| Soil-Al | 683 | 20 | <0.001 | |
| Soil-Al + Litter-N | 676 | 13 | 0.0013 | |
| Soil-Al + Litter-N + Litter-C:P | 670 | 8 | 0.020 | |
| Soil-Al + Litter-N + Litter-C:P + Soil-Na | 662 | 0 | 0.98 | |
| Endogeic | null model | 561 | 47 | <0.001 |
| Soil-Al | 540 | 26 | <0.001 | |
| Soil-Al +Soil-Al:Litter-Ca | 532 | 19 | <0.001 | |
| Soil-Al + Soil-Al:Litter-Ca + Litter-Ca | 529 | 16 | <0.001 | |
| Soil-Al + Soil-Al:Litter-Ca + Litter-Ca + Soil Moisture | 525 | 12 | 0.0022 | |
| Soil-Al + Litter-Ca + Soil-Al:Litter-Ca + Soil Moisture + Litter-C:P | 517 | 4 | 0.10 | |
| Soil-Al + Litter-Ca + Soil-Al:Litter-Ca + Soil Moisture + Litter-C:P + Litter-C:N | 516 | 2 | 0.22 | |
| Soil-Al + Litter-Ca + Soil-Al:Litter-Ca + Soil Moisture + Litter-C:P + Litter-C:N + Litter-lignin | 514 | 0 | 0.67 | |
| Epigeic | null model | 509 | 27 | <0.001 |
| Soil Moisture | 498 | 16 | <0.001 | |
| Soil Moisture + Litter-Mn | 496 | 14 | 0.0012 | |
| Soil Moisture + Litter-Mn + Litter-C:P | 482 | 0 | 1.00 | |
Appendix C
| Tree Species | ||||||||
|---|---|---|---|---|---|---|---|---|
| Soil variables (15–30 cm) | f-value | p | Fraxinus | Acer | Tilia | Quercus | Fagus | Picea |
| pH-KCl | 275 | <0.001 | 4.2 ± 0.58 c | 4 ± 0.37 bc | 3.9 ± 0.28 ab | 3.8 ± 0.27 a | 3.8 ± 0.17 a | 3.7 ± 0.26 a |
| Base saturation (%) | 10 | <0.001 | 60 ± 36 bc | 60 ± 30 c | 43 ± 27 ab | 35 ± 28 a | 36 ± 26 a | 41 ± 32 a |
| K in BaCl2 (μg·K·g−1) | 28 | <0.001 | 38 ± 17 b | 54 ± 57 b | 43 ± 35 b | 46 ± 34 b | 36 ± 23 b | 28 ± 20 a |
| Na in BaCl2 (μg·Na·g−1) | 26 | <0.001 | 17 ± 19 a | 13 ± 9 a | 10 ± 5 a | 9 ± 7 a | 11 ± 6 a | 37 ± 47 b |
| Mg in BaCl2 (μg·Mg·g−1) | 16 | <0.001 | 77 ± 81 b | 61 ± 61 b | 33 ± 26 a | 38 ± 42 ab | 29 ± 25 a | 53 ± 45 ab |
| Ca in BaCl2 (μg·Ca·g−1) | 18 | <0.001 | 954 ± 1049 b | 659 ± 603 b | 375 ± 358 ab | 357 ± 433 a | 312 ± 288 a | 482 ± 481 a |
| Al in BaCl2 (μg·Al·g−1) | 57 | <0.001 | 133 ± 131 a | 136 ± 87 ab | 211 ± 118 bc | 262 ± 133 c | 232 ± 103 bc | 248 ± 136 bc |
| Tree Species | ||||||||
|---|---|---|---|---|---|---|---|---|
| Soil variables (15–30 cm) | f-value | p | Fraxinus | Acer | Tilia | Quercus | Fagus | Picea |
| pH-KCl | 236 | <0.001 | 4.4 ± 0.57 b | 4.2 ± 0.37 ab | 4.0 ± 0.31 a | 4.1 ± 0.4 ab | 4.1 ± 0.43 ab | 4.1 ± 0.37 ab |
| Base saturation (%) | 4.0 | <0.005 | 60 ± 38 | 55 ± 33 | 39 ± 33 | 46 ± 33 | 51 ± 35 | 51 ± 39 |
| K in BaCl2 (μg·K·g−1) | 13 | <0.001 | 29 ± 21 | 32 ± 38 | 29 ± 26 | 34 ± 29 | 27 ± 24 | 26 ± 22 |
| Na in BaCl2 (μg·Na·g−1) | 17 | <0.001 | 17 ± 20 a | 12 ± 7 a | 9,0 ± 5,4 a | 10 ± 9,5 a | 13 ± 8,7 a | 42 ± 58 b |
| Mg in BaCl2 (μg·Mg·g−1) | 8.7 | <0.001 | 76 ± 94 b | 47 ± 59 ab | 28 ± 33 a | 51 ± 60 ab | 43 ± 44 ab | 58 ± 55 ab |
| Ca in BaCl2 (μg·Ca·g−1) | 13 | <0.001 | 1109 ± 1252 b | 590 ± 694 ab | 339 ± 402 a | 522 ± 628 a | 527 ± 524 ab | 692 ± 733 ab |
| Al in BaCl2 (μg·Al·g−1) | 37 | <0.001 | 115 ± 113 | 128 ± 86 | 175 ± 105 | 175 ± 117 | 149 ± 97 | 149 ± 113 |
Appendix D
Appendix E
| Earthworm Species | Component A | Component B | IndVal | p-Value | |
|---|---|---|---|---|---|
| Association with tree species group: | |||||
| Earthworm density | |||||
| Fraxinus + Acer + Tilia | A. longa | 0.97 | 0.33 | 0.57 | 0.008 |
| Fraxinus + Acer + Tilia + Quercus | L. rubellus | 0.92 | 0.59 | 0.74 | 0.009 |
| Fraxinus + Acer + Tilia + Fagus | L. terrestris | 0.92 | 0.87 | 0.89 | 0.001 |
| Fraxinus + Acer + Tilia + Quercus + Fagus | A. caliginosa | 0.99 | 0.70 | 0.84 | 0.001 |
| D. octaedra | 0.98 | 0.53 | 0.72 | 0.006 | |
| Earthworm biomass | |||||
| Fraxinus + Acer + Tilia | A. longa | 0.98 | 0.33 | 0.57 | 0.014 |
| Fraxinus + Acer + Tilia + Quercus | L. rubellus | 0.92 | 0.59 | 0.74 | 0.006 |
| Fraxinus + Acer + Tilia + Quercus + Fagus | L. terrestris | 0.97 | 0.82 | 0.89 | 0.001 |
| A. caliginosa | 0.99 | 0.70 | 0.83 | 0.002 | |
| D. octaedra | 0.96 | 0.53 | 0.71 | 0.010 |
References
- Darwin, C. The Formation of Vegetable Mould Through the Action of Worms with Some Observations on Their Habits; John Murray: London, UK, 1881. [Google Scholar]
- Edwards, C.A. Earthworm Ecology, 2nd ed.; CRC Press: Columbus, OH, USA, 2004. [Google Scholar]
- Lee, K.E. Earthworms—Their Ecology and Relationships with Soils and Land Use; Academic Press: Sydney, Australia, 1985. [Google Scholar]
- Muys, B.; Lust, N. Inventory of the earthworm communities and the state of litter decomposition in the forests of Flanders (Belgium) and its implications for forest management. Soil Biol. Biochem. 1992, 24, 1677–1681. [Google Scholar] [CrossRef]
- Lavelle, P.; Bignell, D.; Lepage, M.; Wolters, V.; Roger, P.; Ineson, P.; Heal, O.W.; Dhillion, S. Soil function in a changing world: The role of invertebrate ecosystem engineers. Eur. J. Soil Biol. 1997, 33, 159–193. [Google Scholar]
- Lavelle, P.; Decaens, T.; Aubert, M.; Barot, S.; Blouin, M.; Bureau, F.; Margerie, P.; Mora, P.; Rossi, J.P. Soil invertebrates and ecosystem services. Eur. J. Soil Biol. 2006, 42, S3–S15. [Google Scholar] [CrossRef]
- Jones, C.G.; Lawton, J.H.; Shachak, M. Organisms as ecosystem engineers. Oikos 1994, 69, 373–386. [Google Scholar] [CrossRef]
- Dempsey, M.A.; Fisk, M.C.; Yavitt, J.B.; Fahey, T.J.; Balser, T.C. Exotic earthworms alter soil microbial community composition and function. Soil Biol. Biochem. 2013, 67, 263–270. [Google Scholar] [CrossRef]
- Hale, C.M.; Frelich, L.E.; Reich, P.B.; Pastor, J. Effects of European earthworm invasion on soil characteristics in Northern Hardwood Forests of Minnesota, USA. Ecosystems 2005, 8, 911–927. [Google Scholar] [CrossRef]
- Vesterdal, L.; Clarke, N.; Sigurdsson, B.D.; Gundersen, P. Do tree species influence soil carbon stocks in temperate and boreal forests? For. Ecol. Manage. 2013, 309, 4–18. [Google Scholar] [CrossRef]
- Frouz, J.; Livecková, M.; Albrechtová, J.; Chronáková, A.; Cajthaml, T.; Pizl, V.; Hánel, L.; Stary, J.; Baldrian, P.; Lhotáková, Z.; et al. Is the effect of trees on soil properties mediated by soil fauna ? A case study from post-mining sites. For. Ecol. Manag. 2013, 309, 87–95. [Google Scholar]
- Curry, J.P.; Schmidt, O. The feeding ecology of earthworms—A review. Pedobiologia (Jena) 2006, 50, 463–477. [Google Scholar] [CrossRef]
- Sheehan, C.; Kirwan, L.; Connolly, J.; Bolger, T. The effects of earthworm functional diversity on microbial biomass and the microbial community level physiological profile of soils. Eur. J. Soil Biol. 2008, 44, 65–70. [Google Scholar] [CrossRef]
- Bouché, M.B. Strategies lombriciennes. Ecol. Bull. 1977, 25, 122–132. [Google Scholar]
- Sims, R.W.; Gerard, B.M. Earthworms: Notes for the Identification of British Species, 4th ed.; The Linnean Society of London & The Estuarine & Coastal Sciences Association by Field Studies Council: Shrewsbury, UK, 1999. [Google Scholar]
- Haimi, J.; Huhta, V. Effects of earthworms on decomposition processes in raw humus forest soil—A microcosm study. Biol. Fertil. Soils 1990, 10, 178–183. [Google Scholar]
- Muys, B.; Granval, P. Earthworms as bio-indicators of forest site quality. Soil Biol. Biochem. 1997, 29, 323–328. [Google Scholar] [CrossRef]
- Haimi, J.; Einbork, M. Effects of endogeic earthworms on soil processes and plant growth in coniferous forest soil. Biol. Fertil. Soils 1992, 13, 6–10. [Google Scholar] [CrossRef]
- Zicsi, A.; Szlavecz, K.; Csuzdi, C. Leaf litter acceptance and cast deposition by peregrine and endemic European lumbricids (Oligochaeta: Lumbricidae). Pedobiologia (Jena) 2011, 54, 1–8. [Google Scholar] [CrossRef]
- Potthoff, M.; Asche, N.; Stein, B.; Muhs, A.; Beese, F. Earthworm communities in temperate beech wood forest soils affected by liming. Eur. J. Soil Biol. 2008, 44, 247–254. [Google Scholar] [CrossRef]
- Jänsch, S.; Steffens, L.; Höfer, H.; Horak, F.; Roß-nickoll, M.; Russell, D.; Toschki, A.; Römbke, J. State of knowledge of earthworm communities in German soils as a basis for biological soil quality assessment. Soil Org. 2013, 85, 215–233. [Google Scholar]
- Schelfhout, S. Tree Species Effect on Earthworm Communities in Danish and Flemish Forests. Master’s Thesis, Ghent University, Ghent, Belgium, June 2010. [Google Scholar]
- De Wandeler, H.; Sousa-Silva, R.; Ampoorter, E.; Bruelheide, H.; Carnol, M.; Dawud, S.M.; Dănilă, G.; Finer, L.; Hättenschwiler, S.; Hermy, M.; et al. Drivers of earthworm incidence and abundance across European forests. Soil Biol. Biochem. 2016, 99, 167–178. [Google Scholar] [CrossRef]
- Ponge, J.F.; Patzel, N.; Delhaye, L.; Devigne, E.; Levieux, C.; Beros, P.; Wittebroodt, R. Interactions between earthworms, litter and trees in an old-growth beech forest. Biol. Fertil. Soils 1999, 29, 360–370. [Google Scholar] [CrossRef]
- Muys, B.; Lust, N.; Granval, P. Effects of Grassland Afforestation with Different Tree Species on Earthworm Communities, Litter Decomposition and Nutrient Status. Soil Biol. Biochem. 1992, 24, 1459–1466. [Google Scholar] [CrossRef]
- Hobbie, S.E.; Reich, P.B.; Oleksyn, J.; Ogdahl, M.; Zytkowiak, R.; Hale, C.; Karolewski, P. Tree species effects on decomposition and forest floor dynamics in a common garden. Ecology 2006, 87, 2288–2297. [Google Scholar] [CrossRef]
- De Schrijver, A.; De Frenne, P.; Staelens, J.; Verstraeten, G.; Muys, B.; Vesterdal, L.; Wuyts, K.; Van Nevel, L.; Schelfhout, S.; Neve, S.; et al. Tree species traits cause divergence in soil acidification during four decades of postagricultural forest development. Glob. Chang. Biol. 2012, 18, 1127–1140. [Google Scholar] [CrossRef]
- Augusto, L.; De Schrijver, A.; Vesterdal, L.; Smolander, A.; Prescott, C.; Ranger, J. Influences of evergreen gymnosperm and deciduous angiosperm tree species on the functioning of temperate and boreal forests. Biol. Rev. 2015, 90, 444–466. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Hui, D.; Luo, Y.; Zhou, G. Rates of litter decomposition in terrestrial ecosystems: Global patterns and controlling factors. J. Plant Ecol. 2008, 1, 85–93. [Google Scholar] [CrossRef]
- Hobbie, S.E. Plant species effects on nutrient cycling: Revisiting litter feedbacks. Trends Ecol. Evol. 2015, 30, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Reich, P.B.; Oleksyn, J.; Modrzynski, J.; Mrozinski, P.; Hobbie, S.E.; Eissenstat, D.M.; Chorover, J.; Chadwick, O.A.; Hale, C.M.; Tjoelker, M.G. Linking litter calcium, earthworms and soil properties: A common garden test with 14 tree species. Ecol. Lett. 2005, 8, 811–818. [Google Scholar] [CrossRef]
- Hagen-Thorn, A.; Callesen, I.; Armolaitis, K.; Nihlgard, B. The impact of six European tree species on the chemistry of mineral topsoil in forest plantations on former agricultural land. For. Ecol. Manag. 2004, 195, 373–384. [Google Scholar] [CrossRef]
- Neirynck, J.; Mirtcheva, S.; Sioen, G.; Lust, N. Impact of Tilia platyphyllos Scop., Fraxinus excelsior L., Acer pseudoplatanus L., Quercus robur L. and Fagus sylvatica L. on earthworm biomass and physico-chemical properties of a loamy topsoil. For. Ecol. Manag. 2000, 133, 275–286. [Google Scholar] [CrossRef]
- Cesarz, S.; Craven, D.; Dietrich, C.; Eisenhauer, N. Effects of soil and leaf litter quality on the biomass of two endogeic earthworm species. Eur. J. Soil Biol. 2016, 77, 9–16. [Google Scholar] [CrossRef]
- Vesterdal, L.; Schmidt, I.K.; Callesen, I.; Nilsson, L.O.; Gundersen, P. Carbon and nitrogen in forest floor and mineral soil under six common European tree species. For. Ecol. Manag. 2008, 255, 35–48. [Google Scholar] [CrossRef]
- Vesterdal, L.; Elberling, B.; Christiansen, J.R.; Callesen, I.; Schmidt, I.K. Soil respiration and rates of soil carbon turnover differ among six common European tree species. For. Ecol. Manag. 2012, 264, 185–196. [Google Scholar] [CrossRef]
- Christiansen, J.R.; Vesterdal, L.; Callesen, I.; Elberling, B.; Schmidt, I.K.; Gundersen, P. Role of six European tree species and land-use legacy for nitrogen and water budgets in forests. Glob. Chang. Biol. 2010, 16, 2224–2240. [Google Scholar] [CrossRef]
- Zaborski, E.R. Allyl isothiocyanate: An alternative chemical expellant for sampling earthworms. Appl. Soil Ecol. 2003, 22, 87–95. [Google Scholar] [CrossRef]
- Valckx, J.; Govers, G.; Hermy, M.; Muys, B. Optimizing earthworm sampling in ecosystems. In Biology of Earthworms; Karaca, A., Ed.; Soil Biology; Springer: Berlin/Heidelberg, Germany, 2011; Volume 24, pp. 19–38. [Google Scholar]
- Bouché, M.B. Lombriciens de France: Ecologie et Systématique. INRA Ann. Zool. Ecol. Anim. 1972, 72, 1–671. [Google Scholar]
- Valckx, J.; Hermy, M.; Muys, B. Indirect gradient analysis at different spatial scales of prorated and non-prorated earthworm abundance and biomass data in temperate agro-ecosystems. Eur. J. Soil Biol. 2006, 42, 341–347. [Google Scholar] [CrossRef]
- Hansen, K.; Vesterdal, L.; Schmidt, I.K.; Gundersen, P.; Sevel, L.; Bastrup-Birk, A.; Pedersen, L.B.; Bille-Hansen, J. Litterfall and nutrient return in five tree species in a common garden experiment. For. Ecol. Manag. 2009, 257, 2133–2144. [Google Scholar] [CrossRef]
- Olson, J.S. Energy storage and the balance of producers and decomposers in ecological systems. Ecology 1963, 44, 322–331. [Google Scholar] [CrossRef]
- R-Core-Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Pinheiro, J.; Bates, D.; Debroy, S.; Sarkar, D.; R-Core-Team. nlme: Linear and Nonlinear Mixed Effects Models, R package version 3.1-128; 2016. Available online: https://CRAN.R-project.org/package=nlme (accessed on 1 December 2016).
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biometr. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed]
- Skaug, H.; Fournier, D.; Bolker, B.; Magnusson, A.; Nielsen, A. Generalized Linear Mixed Models using “AD Model Builder”, R package version 0.8.3.3; 2016. Available online: http://glmmadmb.r-forge.r-project.org/repos/ (accessed on 1 December 2016).
- Martin, T.G.; Wintle, B.A.; Rhodes, J.R.; Kuhnert, P.M.; Field, S.A.; Low-Choy, S.J.; Tyre, A.J.; Possingham, H.P. Zero tolerance ecology: Improving ecological inference by modelling the source of zero observations. Ecol. Lett. 2005, 8, 1235–1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewin, W.-C.; Freyhof, J.; Huckstorf, V.; Mehner, T.; Wolter, C. When no catches matter: Coping with zeros in environmental assessments. Ecol. Indic. 2010, 10, 572–583. [Google Scholar] [CrossRef]
- Sileshi, G. The excess-zero problem in soil animal count data and choice of appropriate models for statistical inference. Pedobiologia (Jena) 2008, 52, 1–17. [Google Scholar] [CrossRef]
- Van Gestel, C.A.M.; Hoogerwerf, G. Influence of soil pH on the toxicity of aluminium for Eisenia andrei (Oligochaeta: Lumbricidae) in an artificial soil substrate. Pedobiologia (Jena) 2001, 45, 385–395. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography (Cop.) 2013, 36, 027–046. [Google Scholar] [CrossRef]
- Hurvich, C.; Tsai, C.-L. Regression and time series model selection in small samples. Biometrika 1989, 76, 297–307. [Google Scholar] [CrossRef]
- Bohlen, P.J.; Groffman, P.M.; Fahey, T.J.; Fisk, M.C.; Suarez, E.; Pelletier, D.M.; Fahey, R.T. Ecosystem consequences of exotic earthworm invasion of north temperate forests. Ecosystems 2004, 7, 1–12. [Google Scholar] [CrossRef]
- Bowman, W.D.; Cleveland, C.C.; Halada, L.; Hresko, J. Negative impact of nitrogen deposition on soil buffering capacity. Nat. Geosci. 2008, 1, 767–770. [Google Scholar] [CrossRef]
- Blake, L.; Goulding, K.W.T.; Mott, C.J.B.; Johnston, A.E. Changes in soil chemistry accompanying acidification over more than 100 years under woodland and grass at Rothamsted Experimental Station, UK. Eur. J. Soil Sci. 1999, 50, 401–412. [Google Scholar] [CrossRef]
- Eggleton, P.; Inward, K.; Smith, J.; Jones, D.T.; Sherlock, E. A six year study of earthworm (Lumbricidae) populations in pasture woodland in southern England shows their responses to soil temperature and soil moisture. Soil Biol. Biochem. 2009, 41, 1857–1865. [Google Scholar] [CrossRef]
- Augusto, L.; Ranger, J.; Binkley, D.; Rothe, A. Impact of several common tree species of European temperate forests on soil fertility. Ann. For. Sci. 2002, 59, 233–253. [Google Scholar] [CrossRef]
- Owojori, O.J.; Reinecke, A.J.; Voua-Otomo, P.; Reinecke, S.A. Comparative study of the effects of salinity on life-cycle parameters of four soil-dwelling species (Folsomia candida, Enchytraeus doerjesi, Eisenia fetida and Aporrectodea caliginosa). Pedobiologia (Jena) 2009, 52, 351–360. [Google Scholar] [CrossRef]
- Decaëns, T.; Margerie, P.; Aubert, M.; Hedde, M.; Bureau, F. Assembly rules within earthworm communities in North-Western France-A regional analysis. Appl. Soil Ecol. 2008, 39, 321–335. [Google Scholar] [CrossRef]
- Millenium Ecosystem Assessment. Ecosystems and Human Well-Being: Biodiversity Synthesis; World Resources Institute: Washington DC, USA, 2005. [Google Scholar]
- European Environment Agency. Climate Change Impacts and Vulnerability in Europe 2016. An Indicator-Based Report; Publications Office of the European Union: Luxembourg, 2017. [Google Scholar]
- Sanderson, M.G.; Collins, W.J.; Johnson, C.E.; Derwent, R.G. Present and future acid deposition to ecosystems: The effect of climate change. Atmos. Environ. 2006, 40, 1275–1283. [Google Scholar] [CrossRef]
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- De Cáceres, M.; Legendre, P.; Moretti, M. Improving indicator species analysis by combining groups of sites. Oikos 2010, 119, 1674–1684. [Google Scholar] [CrossRef]
- De Cáceres, M.; Jansen, F. Indicspecies: Relationship between species and groups of sites, R package version 1.7.6; 2016. Available online: https://cran.r-project.org/package=indicspecies (accessed on 1 December 2016).
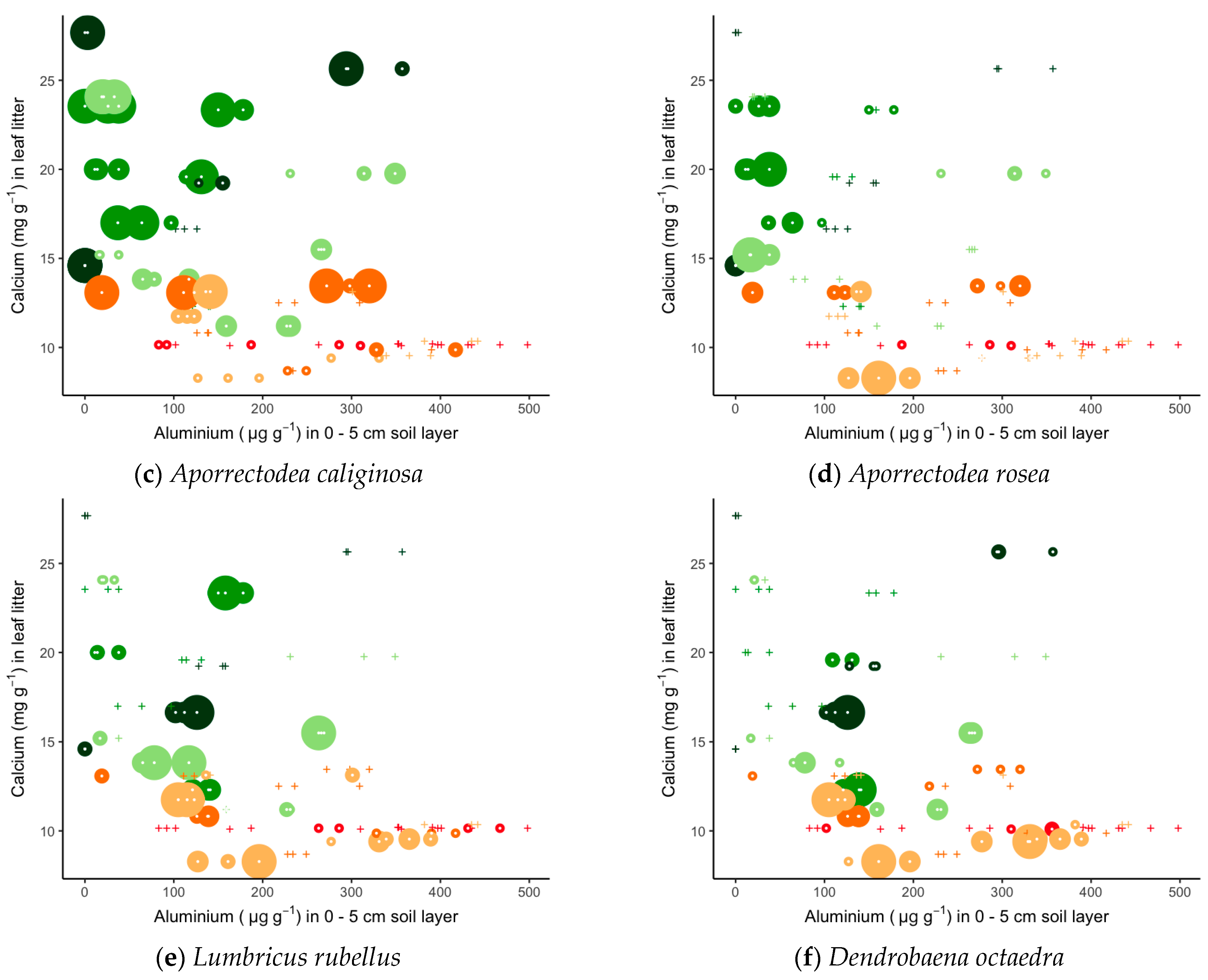
| Tree Species | ||||||||
|---|---|---|---|---|---|---|---|---|
| Soil variables (0–5 cm) | f-value | p | Fraxinus | Acer | Tilia | Quercus | Fagus | Picea |
| Moisture (%) | 1475 | <0.001 | 14 ± 5 c | 15 ± 4 c | 12 ± 3 b | 13 ± 4 bc | 12 ± 4 b | 9 ± 2 a |
| pH-KCl | 325 | <0.001 | 4.2 ± 0.6 c | 4.2 ± 0.5 c | 4.0 ± 0.4 c | 3.7 ± 0.3 b | 3.7 ± 0.2 b | 3.5 ± 0.2 a |
| Base saturation (%) | 108 | <0.001 | 73 ± 28 b | 78 ± 24 b | 71 ± 20 b | 49 ± 20 a | 49 ± 21 a | 41 ± 19 a |
| K+ in BaCl2 (μg·g−1) | 50 | <0.001 | 100 ± 88 bc | 114 ± 91 c | 91 ± 56 bc | 85 ± 57 bc | 67 ± 42 ab | 41 ± 22 a |
| Na+ in BaCl2 (μg·g−1) | 28 | <0.001 | 19 ± 16 a | 17 ± 11 a | 15 ± 8 a | 13 ± 7 a | 13 ± 7 a | 38 ± 48 b |
| Mg2+ in BaCl2 (μg·g−1) | 48 | <0.001 | 139 ± 106 c | 108 ± 72 bc | 81 ± 39 ab | 68 ± 53 a | 49 ± 32 a | 57 ± 41 a |
| Ca2+ in BaCl2 (μg·g−1) | 42 | <0.001 | 1241 ± 1020 c | 1050 ± 690 bc | 796 ± 437 ab | 481 ± 388 a | 446 ± 293 a | 467 ± 351 a |
| Al3+ in BaCl2 (μg·g−1) | 42 | <0.001 | 115 ± 121 a | 87 ± 58 a | 151 ± 118 a | 261 ± 121 bc | 231 ± 105 b | 309 ± 133 c |
| Tree Species | ||||||||
|---|---|---|---|---|---|---|---|---|
| f-value | p | Fraxinus | Acer | Tilia | Quercus | Fagus | Picea | |
| Litterfall | ||||||||
| Foliar mass (Mg·ha−1·year−1) | 127 | <0.001 | 2.7 ± 1.0 bc | 2.8 ± 0.39 c | 2.4 ± 0.68 ab | 2.6 ± 0.55 abc | 2.1 ± 0.34 a | 3.9 ± 0.46 d |
| C:N ratio | 151 | <0.001 | 25 ± 5.4 a | 27 ± 2.6 ab | 28 ± 3.5 ab | 32 ± 3.0 b | 42 ± 14 d | 36 ± 1.9 c |
| C:P ratio | 5200 | <0.001 | 358 ± 128 a | 477 ± 81 b | 407 ± 87 a | 415 ± 44 a | 575 ± 113 c | 473 ± 37 b |
| N (mg·g−1) | 250 | <0.001 | 19 ± 3.6 d | 17 ± 1.7 c | 18 ± 2.5 c | 16 ± 1.5 b | 13 ± 3.6 a | 13 ± 0.74 a |
| Ca (mg·g−1) | 124 | <0.001 | 21± 5.3 c | 19 ± 4.0 c | 17 ± 4.3 b | 10 ± 1.7 a | 11 ± 1.8 a | 10 ± 0.03 a |
| Mn (mg·g−1) | 1025 | <0.001 | 0.23 ± 0.27 a | 0.63 ± 0.33 b | 1.1 ± 0.52 c | 1.6 ± 0.50 d | 1.6 ± 0.55 d | 1.2 ± 0.04 c |
| P (mg·g−1) | 362 | <0.001 | 1.5 ± 0.56 d | 1.0 ± 0.19 ab | 1.3 ± 0.28 cd | 1.2 ± 0.13 bc | 0.87 ± 0.18 a | 1.0 ± 0.08 ab |
| Lignin (%) | 506 | <0.001 | 18 ± 2.8 a | 18 ± 3.3 a | 27 ± 4.1 c | 27 ± 2.6 bc | 29 ± 1.6 d | 25 ± 0.17 b |
| Forest floor | ||||||||
| Foliar mass (Mg·ha−1) | 39 | <0.001 | 0.57 ± 0.4 a | 1.7 ± 1.4 a | 1.7 ± 1.0 a | 6.0 ± 2.6 b | 8.1 ± 2.8 b | 37 ± 10 c |
| Foliar forest floor turnover rate (year−1) | 93 | <0.001 | 0.80 ± 0.18 e | 0.68 ± 0.18 de | 0.60 ± 0.20 d | 0.35 ± 0.17 c | 0.23 ± 0.09 b | 0.11 ± 0.02 a |
| Earthworm Species | Ecological Group | Sites | Tree Species |
|---|---|---|---|
| Allolobophoridella eiseni (Levinsen) | Epigeic | K | Pi |
| Dendrodrilus rubidus (Savigny) | Epigeic | K M O V W | Fr Ac Ti Qu Fa Pi |
| Dendrobaena octaedra (Savigny) | Epigeic | K M O V Vi W | Fr Ac Ti Qu Fa Pi |
| Eisenia fetida (Savigny) | Epigeic | Vi | Ac |
| Lumbricus castaneus (Savigny) | Epigeic | K M O V | Fr Ac Ti Qu Fa |
| Lumbricus festivus (Savigny) | Epigeic | W | Ac Ti Qu |
| Lumbricus rubellus (Hoffmeister) | Epigeic | K M O V W | Fr Ac Ti Qu Fa Pi |
| Lumbricus terrestris (Linnaeus) | Anecic | K M O V Vi W | Fr Ac Ti Qu Fa Pi |
| Aporrectodea longa (Ude) | Anecic | M W | Fr Ac Ti Qu Fa Pi |
| Aporrectodea caliginosa (Savigny) | Endogeic | K M O V Vi W | Fr Ac Ti Qu Fa Pi |
| Aporrectodea rosea (Savigny) | Endogeic | M O V Vi W | Fr Ac Ti Qu Fa Pi |
| Octolasion cyaneum (Savigny) | Endogeic | O Vi W | Fr Ac Ti Qu Fa |
| Response Variable | Predictor Variables in Optimal Model | df | AICc Optimal Model | AICc Null Model | R2 Optimal Model | R2 Null Model |
|---|---|---|---|---|---|---|
| Earthworm density | ||||||
| Total | Soil: Moisture, Al Litter: Ca, Mn Soil-Al:Litter-Ca | 9 | 1061 | 1125 | 0.48 | 0.14 |
| Anecic | Soil: Al, Na Litter: N, C:P ratio, Ca Soil-Al:Litter-Ca | 10 | 766 | 850 | 0.62 | 0.12 |
| Endogeic | Soil: Moisture, Al Litter: C:P ratio, C:N ratio, Ca, Lignin Soil-Al:Litter-Ca | 11 | 725 | 800 | 0.28 | 0.14 |
| Epigeic | Soil: Moisture Litter: Mn Forest floor turnover rate | 7 | 804 | 817 | 0.42 | 0.36 |
| Earthworm biomass | ||||||
| Total | Soil: Moisture, Al Litter: Ca Soil-Al:Litter-Ca | 7 | 821 | 902 | 0.53 | 0.12 |
| Anecic | Soil: Al, Na Litter: N, C:P ratio | 8 | 662 | 739 | 0.66 | 0.13 |
| Endogeic | Soil: Al, Moisture Litter: Ca, Lignin, C:P ratio, C:N ratio Soil-Al:Litter-Ca | 11 | 514 | 561 | 0.21 | 0.13 |
| Epigeic | Soil: Moisture Litter: Mn, C:P ratio | 7 | 482 | 509 | 0.53 | 0.40 |
| Predictor Variables | Total Density | Anecic Density | Endogeic Density | Epigeic Density | ||||
|---|---|---|---|---|---|---|---|---|
| z value | p value | z value | p value | z value | p value | z value | p value | |
| Soil Al | −5.7 | <0.001 | −6.0 | <0.001 | −5.1 | <0.001 | ||
| Soil Na | −2.8 | 0.006 | ||||||
| Soil Moisture | 5.1 | <0.001 | 4.9 | <0.001 | 2.7 | 0.007 | ||
| Litter Ca | 2.8 | 0.006 | 3.4 | <0.001 | 3.2 | 0.001 | ||
| Litter N | 4.7 | <0.001 | ||||||
| Litter Mn | 2.6 | 0.01 | 3.1 | 0.002 | ||||
| Litter lignin | 3.4 | <0.001 | ||||||
| Litter C:N ratio | −4.5 | <0.001 | ||||||
| Litter C:P ratio | 4.63 | <0.001 | 6.2 | <0.001 | ||||
| Forest floor turnover rate | 2.0 | 0.047 | ||||||
| Soil-Al:Litter-Ca | 4.0 | <0.001 | 2.38 | 0.018 | 6.4 | <0.001 | ||
| Total Biomass | Anecic Biomass | Endogeic Biomass | Epigeic Biomass | |||||
| z value | p value | z value | p value | z value | p value | z value | p value | |
| Soil Al | −6.2 | <0.001 | −6.0 | <0.001 | −3.5 | <0.001 | ||
| Soil Na | −3.1 | 0.002 | ||||||
| Soil Moisture | 4.4 | <0.001 | 3.7 | <0.001 | 3.5 | <0.001 | ||
| Litter Ca | 2.4 | 0.017 | 3.2 | 0.001 | ||||
| Litter N | 5.3 | <0.001 | ||||||
| Litter Mn | 3.6 | <0.001 | ||||||
| Litter lignin | 3.0 | 0.003 | ||||||
| Litter C:N ratio | −3.4 | <0.001 | ||||||
| Litter C:P ratio | 4.0 | <0.001 | 4.7 | <0.001 | -3.9 | <0.001 | ||
| Soil-Al:Litter-Ca | 5.0 | <0.001 | 4.6 | <0.001 | ||||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schelfhout, S.; Mertens, J.; Verheyen, K.; Vesterdal, L.; Baeten, L.; Muys, B.; De Schrijver, A. Tree Species Identity Shapes Earthworm Communities. Forests 2017, 8, 85. https://doi.org/10.3390/f8030085
Schelfhout S, Mertens J, Verheyen K, Vesterdal L, Baeten L, Muys B, De Schrijver A. Tree Species Identity Shapes Earthworm Communities. Forests. 2017; 8(3):85. https://doi.org/10.3390/f8030085
Chicago/Turabian StyleSchelfhout, Stephanie, Jan Mertens, Kris Verheyen, Lars Vesterdal, Lander Baeten, Bart Muys, and An De Schrijver. 2017. "Tree Species Identity Shapes Earthworm Communities" Forests 8, no. 3: 85. https://doi.org/10.3390/f8030085