
Impacts of Beech Bark Disease and Climate Change on American Beech

Department of Biology, Green Mountain College, Poultney, VT 05764, USA
*
Author to whom correspondence should be addressed.
Forests 2017, 8(5), 155; https://doi.org/10.3390/f8050155
Submission received: 14 March 2017
/
Revised: 25 April 2017
/
Accepted: 28 April 2017
/
Published: 3 May 2017
(This article belongs to the Special Issue Climate Effects on Forest Tree/Stand Predisposition to Biotic Disturbance)
Abstract
:American beech (Fagus grandifolia Ehrh.) is a dominant component of forest tree cover over a large portion of eastern North America and this deciduous, mast-bearing tree species plays a key role in these forest ecosystems. Beech bark disease (BBD) is a scale insect-fungus complex that has caused the decline and death of afflicted beech trees. This disease has become a common feature in North American forest landscapes. Resistance to BBD is at the level of the beech scale (Cryptococcus fagisuga Lind.). Beech scale attack predisposes the tree to subsequent infection by Neonectria fungi. The impact of this tree disease has been shown to be significant, particularly in beech dominated forests. Scale-free trees (resistant to BBD) have been reported to range from only 1% to 3% in infested stands, with estimates ranging from 80–95% for overall infestation (for all beech within the current North American range). In addition to BBD, overall beech health will be directly impacted by climate change, if one specifically considers the expected fluctuations in precipitation leading to both drought periods and flooding. Beech is particularly sensitive to both extremes and is less resilient than other broad leaf tree species. Although the increase in global temperatures will likely shift the current range of the American beech, milder winters and less snowpack will favor propagation and survival of the beech scale. This review aims to present the current outlook for American beech in light of climate change. The natural history of the American beech and the onslaught of BBD during the last century will be covered, followed by the potential effects of a changing climate on BBD-infested forests. Evidence from models of European beech (Fagus sylvatica) and other host-pathogen systems will supplement data directly gathered to evaluate BBD in American beech. We present the case that although climate change is likely to be a confounding factor in the continued loss of American beech, increasing our understanding of possible mechanisms of resistance and environmental factors that may influence susceptibility of American beech to BBD can inform proactive management strategies.
1. Introduction
This literature review aims to provide the reader with sufficient background to appreciate the role of the American beech (Fagus grandifolia Ehrh.) in forest ecosystems of North America, the pathology and impact of beech bark disease (BBD) on this species, and the potential effects of climate change on the overall health of both beech and the forests they populate.
In exploring the natural history of American beech, the broad scale of this geographic range will be presented and explained. Beech provides forest cover in different forest cover types. The implications and role of this shade-tolerant species in forest structure will also be discussed. American beech regularly produce large seed crops (every two to eight years) that provide an important food source for small mammals and birds as well as deer and bear.
Described as “an old disease in a new place” [1], BBD in American beech is the result of an inadvertent introduction of a European pathogen to North America. The current health status of American beech, BBD infestation rates, and prognosis for beech will be systematically presented with data from stand analyses in both the United States and Canada, while also providing a historical context.
The potential impacts of climate change on American beech are first considered in the absence of BBD, taking advantage of available studies focused on both American and European beech (Fagus sylvatica). This review focuses primarily on impacts of increased temperature as well as drought. Following general consideration, the impacts of climate change on beech will be revisited in light of intensive infestation by BBD. A brief but comprehensive overview of the possible mechanisms of resistance and environmental factors that that are most likely to influence susceptibility of American beech to BBD will then be discussed.
In addition to lessons and projections from studies in American and European Beech on the impacts of climate change on BBD-infested American beech, additional correlations will be drawn to and from other pathogen-host models, in particular hemlock and the hemlock wooly adelgid, and white pine and the mountain pine beetle. Current plans and projections for beech forest management in light of both BBD and climate change, including propagation of resistant beech, grafting efforts and reintroduction efforts will also be presented.
2. Natural History of the American Beech
American beech is a monoecious, flowering, late successional tree species endemic to eastern North America. The only native member of the Fagus genus in the Americas, American beech is among the most widely distributed hardwoods on the North American continent (Figure 1) [2]. Its range extends from Nova Scotia to the northern edge of Florida and to the eastern border of Wisconsin, the western border of Mississippi and into some parts of Texas and Oklahoma [3,4,5]. Researchers have also documented that American beech persists, albeit very rarely, in a few small, unique microclimates in the cloud forests of Mexico (Fagus grandfolia var. mexicana) [6]. Additionally, though predominantly an inland species, American beech is also found in coastal regions and may infrequently become a locally dominant species in previously fire-damaged forests along the southern New England coast [7]. Though the importance of winter temperatures has long been emphasized as a key explanatory tool for the northern limit of tree species in North America [8,9], research regarding the climatic controls on worldwide beech distribution has suggested that the northern and southern limit of American beech may be equally well explained by low growing season temperatures and high summer temperatures, respectively [2,10]. Researchers have noted that, whereas there are no known American beech stands in Newfoundland, there are some stands found in southern Quebec. Winter temperatures at sites in Newfoundland at similar latitudes to the beech range in Quebec are higher than in Quebec, whereas average growing season temperatures are much lower. Thus, it is possible that average temperature of the growing season, considered in conjunction with average coldest winter temperatures, is an important determining factor for American beech’s northern limit.
Likely due in part to the broad scale of this geographic range, American beech is associated with a wide variety of other trees. In New England, American beech is a dominant feature of forest cover in three different forest cover types, Beech-Sugar Maple, Sugar Maple-Beech-Yellow Birch, and Red Spruce-Sugar Maple-Beech [11]. Notable other associated trees and forest cover types include Balsam Fir forest types in New England, Michigan, Wisconsin and Minnesota, Oak-Hickory forest types across some of the northern central United States and southern magnolias in the southeastern United States [4,12,13].
Specific soil types certainly favor European beech (as would be expected for American beech). In European beech, calcareous soils seem to counteract the potentially negative effect of certain pollutants [14]. Decline of overall forest health due to soil acidification and intense agricultural fertilizer continues in both Europe and North America. As noted by Tubbs and Houston, American beech seems able to tolerate a soil pH ranging from 4.0 to 6.0 and multiple specific soil types, namely those of the orders Alfisol, Oxisol, and Spodisol, though soils with a large humus layer appear to be favored [4,15]. In line with its large geographic distribution, American beech grows in a range of soil conditions.
Throughout the range of environments that are home to American beech, there are two consistent characteristics exhibited by American beech. Firstly, they are typically classified as “very tolerant” of shade [15] and, as their Latin name implies, are shade creators. In beech dominated (or co-dominated) forests, this results in a distinctively shaded forest floor, creating an inability for less shade tolerant saplings to establish themselves. It is for this reason that some American beech forests have a very minimal understory other than beech or similarly shade tolerant species. It is not uncommon for beech dominated forests to have substantial beech sprouting that appears to be even more robust under adverse conditions [16] including beech bark disease [17,18,19]. The beech understory effectively outcompetes other less shade tolerant trees. The second ubiquitous characteristic of American beech is that it produces abundant mast that is utilized as food by a variety of species, a factor that is especially important in northern hardwood forests where they tend to be the only trees that produce edible nuts [4]. Species today that rely on the beech mast include black bear, marten, deer, porcupine, raccoons, squirrels and blue jays [20]. In the past, beech mast served as the principal food source for the passenger pigeon, and indeed some researchers and naturalists have speculated that widespread clearing of beech-dominated forests may have been a contributing factor in the decline of the now-extinct species [21,22].
Additionally, Garneau et al. found a contribution of abiotic (rainfall, temperature, land management practices) and soil factors on small mammal diversity in beech-dominated forests. They have stressed the importance of long term (versus short term) analyses to better grasp the intermingling roles of beech mast, forest structure and various biotic and abiotic factors [23].
3. Beech Bark Disease
Disruption of beech forests by human development and pathogens has had long lasting effects. Entire forests were clear cut in the Eastern United States in the 1800s. Even before the stands recovered, beech bark disease arrived in Maine (via Nova Scotia) in the early 1900s. Beech bark disease (BBD) is the result of a scale infection (Cryptococcus fagisuga Lind.) followed by a Neonectria fungal invasion (Figure 2). The scale insect was inadvertently brought to North America by importation of ornamental European beech. European beech are more tolerant to infection and show more resistance to the beech scale relative to their North American counterparts [24]. The infection rates of beech by this pair have proved to be devastating in the United States; an 80–90% rate of infection within a stand is not atypical. Today, in measured stands, scale-free trees (resistant to BBD) have been reported to range from only 1% to 3% [25,26] with estimates ranging from 80–95% for overall infestation (for all beech within the current North American range) with 50% dead or dying from BBD [27]. The scale is often hidden under the leaf litter and will begin its attack at the tree base. Unless the litter is removed and the bark at the tree base is carefully examined, a tree may only appear (temporarily) to be resistant to BBD.
Pioneering work in understanding the mechanism of this disease was carried out by Ehrlich. He was the first to effectively and systematically document BBD in the United States within approximately thirty years of its arrival [28]. Continued efforts by a number of scientists, including notably among others, Shigo, Houston, and Wainhouse, have increased our understanding of the pathology of this disease [19,24,25,29,30,31,32,33]. The fungus primarily associated with BBD was originally identified and named as Neonectria coccinea var. faginata [34], but this is now understood to be two species, namely Neonectria faginata and Neonectria coccinea. Neonectria faginata is considered a North American species and Neonectria coccinea is thought to be a European species, whereas Neonectria ditissima (previously known as Neonectria gallignea) is found on both continents [35] and is considered most responsible for BBD in Europe [27]. In North America, Neonectria ditissima and Neonectria faginata are considered the pathogens specifically responsible for BBD. Photographs provided in Figure 2 represent the phenotypic expression of this fungal genus, and are not intended to allow one to differentiate these species. Bionectria ochrolueca is another species that has been documented on severely infected and dying beech in both the United States and Canada [36]. Work by Jung has elucidated another potential pathogen, namely Phytophthora, that may also be contributing to BBD in Europe [37].
Current projections from the USDA Forest Service predict that BBD will spread throughout the entire beech range by 2025 [1,27]. Conservative models (without “stochastic jumps”) estimate that the front will advance approximately 15 km/year [38]. The spread of BBD is initiated by an “advancing front”; beech scale is carried by wind, animals or even by humans moving harvested or downed infested beech [20,27,39]. Stages relative to BBD infestation have been designated as pre-infestation, advancing front, killing front and aftermath [20,32].
Unfortunately for American beech, selection for resistance to scale infestation has only just begun as they have not co-evolved with the exotic and invasive European pathogens introduced to the continent in the late 1800s [24,40]. A 2015 study in Germany designed to examine the effect of forest stand management on scale infestation reported rates of infestation ranging from 16% to 35% [41], compared to much greater rates of infestation in American beech. European beech is more resistant than American beech to BBD based on overall infestation rates in each species [24].
In Europe, BBD is an endemic disease whereas in North America BBD displays characteristics of an epiphytotic disease, typical for epidemic as well as endemic development of the disease [20,42]. We cannot ignore the concept of the “climate envelope” and the fact that the geographic range of beech has been speculated to move north [43] in light of what can already be considered an epidemic. It had been assumed, prior to 2005, that the beech range would be able to migrate rapidly based on fossil pollen data, yet new evidence has required revisiting these older models. Based on genetic analyses utilizing 17 beech haplotypes, the spread of the tree is no longer estimated to track with the predicted warming. The rate is now less than the required minimum of 100 m/year [43]. Richard Pearson provides a commentary on McLachlan et al. and agrees that the molecular evidence helps to more fully examine the question of migration rates. He does question the climate envelope model itself and points to the need for more inclusive modeling that not only incorporates envelope models and species dispersal, but life-history dynamics, land use and human-induced climate change [44]. These data do not take into account that more than 90–95% (estimates will vary regionally but are very high regardless) of the trees are infected, dying or dead from BBD in the United States or the overall reduction in mast (viable seed) over time that results from BBD. Undoubtedly BBD, if it does not kill highly susceptible trees within a few growing seasons, it will hinder the growth of a wide range of susceptible trees, in both second growth and old growth forests [45], and also decrease the overall mast of a stand.
Morin and Liebhold also point out that beech bark disease is not working in isolation—additional pests, such as the wooly adelgid, are simultaneously attacking hemlocks in the same forests [46]. Impacts on not one, but two late-successional species, could have compounding effects on forest structure. An additional study by Krasny and DiGregorio looked at the concomitant impact of BBD and gypsy moths on beech as gap makers (defined as standing live trees with >50% branch loss, standing dead, and treefall). Over the six year study, the land gap area increased from approximately 20% to slightly over 30% [47].
Evans et al. have postulated comprehensive models to evaluate potential and, in some scenarios, substantial changes, that BBD may have on forest structure [48]. Incorporated into these hypothetical models are the assumptions of a disturbance threshold effect on BBD severity and a significant ecological role for beech root sprouting, both of which are predicted to contribute to markedly different outcomes for regional forest structure depending on current and historical forest composition.
4. The Impacts of Climate Change on American Beech
Geßler et al. have provided an excellent review specifically on projected impacts of climate change for European beech pointing out that by 2100, there is an expectation that global surface temperature will be elevated by 1.5–5.8 °C [49]. Correlates for American beech that can be drawn from this study include high susceptibility to the impacts of drought, as well as the opposite extreme, water-logging and flooding [49] that will undoubtedly impact both seedlings and adult beech. Availability of water has a tremendous impact on beech biomass relative to other suspected factors such as interspecies competition.
Droughts and elevated summer temperatures have already been attributed to global climate change. These circumstances are not unexpected conditions for North American beech stands. Systematic assessment of environmental factors that influence the overall health of European beech in southern Britain can provide a reasonable framework for the closely related American beech. One particular study used the extent of crown thinness, overall crown architecture and leaf color to determine the health of beech in different environments. A significant negative impact on crown chlorosis in response to drought and particularly high summer temperatures was documented [14].
As previously mentioned, in line with its large geographic distribution, American beech grows in a range of soil conditions. Due to the variable soil conditions in which American beech exists, the complexity of these effects insofar as they are related to changes in soil dynamics is likely to be even greater still. In recent years, there has been a concerted effort, especially within the realm of biogeochemistry, to understand the interaction between soil and atmospheric carbon dynamics with special attention given to the role of climate change in modulating these interactions [50,51,52]. Much of the research of this nature is of great relevance to the future of American beech stands. For instance, some experimental studies have suggested that decreased snow cover in the winter (a result of increased winter temperatures) that leads to soil freezing may significantly alter the leaching of inorganic nitrogen and phosphorus in northern hardwood forests [53,54,55,56], in which American beech is frequently a component. However, there exists no clear, specific consensus in the scientific literature as to how soil ecosystems will respond to global climate change or how those changes will translate into specific changes in forest communities, insofar as we are aware. Even general changes that might be expected do not have universal acceptance. For example, some researchers agree that an increase in average temperature ought to result in increased soil microbial respiration rates and, thus, an increase in the amount of carbon lost from soil ecosystems [57]. Yet, regional warming could increase evaporation rates, drying soil, with different impacts on microbial respiration rates. Also, much of the North American beech population is covered in snow pack at least during part of the year—so water in this case may not be a limiting factor for overall microbial activity. As Campbell et al. make evident in their extensive review of the literature concerning climate change and biogeochemical cycling in the northeastern forests of North America, the predictability of climate change impacts on soil dynamics is fraught with uncertainty, particularly with respect to the interactions between various effects of climate change (such as that between nitrogen deposition and increases in atmospheric CO2 concentrations), the extent of the CO2 fertilizer effect, and the potential feedback mechanisms at play in various climate change scenarios [57].
In addition to variability in soil conditions, American beech is also found at a wide range of elevations, specifically at higher elevations in the southeastern United States and lower elevations in northeastern North America [4]. This will be important to consider as the range of beech moves both in latitude (northward) and to higher elevations (assuming increased temperatures) as predicted with climate change. The beech range itself is expected to be displaced farther north in response to climate change over the next century; changes in the latitudinal range of beech may be a reasonable ecological response to attempt to effectively compensate for increasing temperatures along the Eastern United States but caveats to this theory will be discussed later (see section on Considerations for Beech Forest Management). Additionally, the United States Forest Service has modeled predictions in the change of suitable habitat for 134 tree species endemic to the United States. Under the assumptions of the General Circulation Model scenarios, the Forest Service predicts both a northern and westward expansion of the potential suitable habitat for American beech. Cumulatively, they predict that there will be an approximate increase in the area of land that is suitable for beech growth of 10.7 percent, assuming that global carbon emissions remain at their current level [58].
It should be noted that studies in European beech have shown that mature beech leaves increase their antioxidative properties (by increasing the expression of key protective enzymes such as superoxide dismutase and glutathione reductase) in response to increased temperatures. Although this adaption is not specific to beech, Polle and Morawe speculate that this might be an adaptive measure to protect beech in light of increasing temperatures and elevated ozone during the summer months [59]. Perhaps in light of climate change impacts, natural selection will favor beech with protective genomic adaptations. Although beech trees are considered to be relative newcomers on a geological time scale [2], on an evolutionary time scale since the Holocene migrations, genetic selection has occurred.
In 2010, Kramer et al. put forth a compelling argument for the need to consider “local differentiation” when developing species expansion models in response to climate change (in this case with evidence from European beech) [60]. Allozyme analysis, haplotype analyses and proteomic studies have shown genetic variation among several tested populations [61,62,63]. Our laboratory has found genetic heterogeneity both among and between large tree stands in the Green and Adirondack Mountains through microsatellite analyses (Cohojari, O., Malik, E. and Coe, N., manuscript in preparation). Our results show genetic variation in trees within Vermont as compared to New York, and both Vermont and New York populations are genetically distinct from published data for Michigan [64]. There is no reason to not consider regional adaptations to particular microenvironments and a variation in the responses of sub-populations of beech to expected changes in precipitation and elevated temperatures expected due to climate change.
5. Impacts of Climate Change on BBD-Infested American Beech: Lessons and Projections from Studies in American and European Beech
Drought conditions will decrease nitrogen soil uptake capacity of beech which will in turn reduce growth [49]. Interestingly, should bark nitrogen levels be lower in certain American beech populations, this may inadvertently protect them from host invasion in that the scale prefer old growth (to secondary growth) trees, presumably because of higher bark nitrogen levels. This protection will be only short term and relative to the natural range of nitrogen bark concentration in a given stand. A study looking specifically at the effect of bark nitrogen levels on infestation in both old-growth and second-growth forests showed a positive correlation between bark nitrogen concentration and infestation [65]. Lower bark nitrogen may benefit a stand overall, but will not prevent infestation. This phenomenon has also been noted in European beech [41]. Additionally, Wargo has documented an increase in both specific amino acids and the total amino acid concentration in bark of infested compared to non-infested trees [66]. It should be noted that Latty and colleagues showed that the increased nitrogen levels in bark in diseased trees is a cause, not the result, of infestation [65]. Importantly, regardless of total bark nitrogen content in an individual tree or forest type, overall tree health will be compromised as drought will reduce stomatal conductance and the impacts can be exacerbated if there is more understory competition.
Dukes et al. present a synthesis of the current literature regarding the interaction between “nuisance”’ species and changing climate dynamics, wherein BBD is used as one of six case studies [67]. In this portion of the review, the authors note that the scale insect associated with BBD is not able to withstand minimum winter daily temperatures below −34 °C and, as such, BBD has historically been absent from certain regions of northern Maine and Canada. However, they mention research conducted by Kasson and Livingston that demonstrates a northern movement of BBD that may be attributable to increased winter temperatures and, therefore, more suitable habitat for the scale insect [26]. Furthermore, Dukes et al. note the complexity of the issue by pointing out that the scale insects, as demonstrated by Grove and Houston [68] use insulated space beneath the snowpack at the base of the tree as refugia during the winter. Thus, there is also potential for increased scale mortality as warming winter temperatures decrease snowpack.
As the range of the American beech marches northward, there is no reason to not assume that the pathogen will accompany its host. The scale and fungus have robustly moved to new host beech and are expected to reach the entire range. This would mean that the pathogens were able to travel the entire latitudinal range of the United States in less than one hundred years. The Canadian Maritime provinces have already endured massive beech die off, estimating only 3.3% to be resistant to BBD there now [69]. Therefore moving north into farther regions of Canada would not be unprecedented and can be predicted with reasonable confidence as average minimum temperatures during winter are expected to rise. The likely pattern we will see may mimic what has already occurred although with some caveats and perhaps a bit of good news. Beech scale prefer larger beech trees. As mentioned previously, the spread of BBD is initiated by an “advancing front”. During this stage, beech trees with relatively larger dbhs may be preferentially attacked. This supposition is based on evidence that larger trees with greater concentrations of bark nitrogen and old growth forests are particularly hard hit initially [27,70,71]. As beech move north, it may take some time for mature and larger trees to become established and the scale infestation may lag behind significantly. Once the scale has entered a stand though, even the smallest of susceptible trees are not safe. It is not uncommon to see saplings in the Northeastern United States covered with scale. Beech trees are strikingly resilient though, and trees that have been removed from campsites due to obvious peripheral damage by BBD, have surprisingly appeared to maintain inner bark integrity. This may be the case for extended periods of time which can explain why a visibly infected beech can still have a reasonably full crown with little sign of chlorosis. However, highly infected trees are also more susceptible to beech snap and beech snag (loss of tree limbs) so the risk foresters managing heavily infested beech have to consider is well substantiated. As the scale is able to take hold in a region, this area becomes known as the “killing front” [27]. BBD resistance is at the level of the scale and potentially resistant trees are easier to identify at this stage. The “aftermath zone” is just as its name implies; this is the term referred to the forest once the scale and the fungus have ravaged through. The first report of a killing phase of beech in an aftermath forest was reported in 2012. A comprehensive study in Northern Maine provides direct evidence as to the potential impacts of climate change in the region. Over a four year period, beginning in 2000, a series of mild winters occurred, with temperatures remaining above −30 °F. This time period coincided with less than average autumn rainfall (2000–2003) and a severe drought (1999–2002). The result was a resurgence in the death of beech trees [26].
6. Impacts of Climate Change on BBD-Infested American Beech: Applied Examples from Other Pathogen-Host Models
Climate change will have an impact on susceptibility and spread of BBD as it will influence overall tree health. Although we do not have deterministic modeling for how climate change will impact different regions and forest types in the Eastern United States specifically, climate change will likely influence biotic and abiotic factors that will in turn influence BBD. The impacts to be considered are nutrient and water availability and temperature. Nutrient and water availability will likely favor and increase the reproductive capacity of the scale as average temperatures begin to rise and winters become milder, providing less robust seasonal checks on population growth and maintenance. Temperatures are expected to rise 1 °C in the next eighty years and strongly favor insect pest survival and reproduction, yet the impact of climate change on fungal populations is admittedly less clear [67]. Recent work has explored the expected impacts of climate change on the range of the hemlock wooly adelgid (Adelges tsugae) [71]. This pest has a similar lifecycle and attack mechanism as compared to the beech scale. Both organisms infect by way of a stylet insertion. The hemlock wooly adelgid attacks at the base of needles at the twig level, whereas the beech scale attacks the bark itself. Paradis et al. provide convincing arguments that as climate change promotes elevated winter temperatures, the wooly adelgid will be well suited to move farther north [71]. Although modeling data focused on the specific projected geographic expansion of beech scale is not available as we are aware to date, drawing correlates to expectations for another scale pathogen in our region seems reasonable.
Given the subsequent changes in environmental characteristics between varying elevations, and considering the potential differential effects of climate change on high and low elevation ecosystems, researchers should consider how changes in elevation may modulate the dispersal and impact of tree disease. For example, some research has suggested that increasing outbreaks of mountain pine beetle (Dendroctonus ponderosae) infestations in previously unaffected, high elevation, white bark pine (Pinus ablicaulis) stands are driven by a warming climate which expands the beetle’s habitat suitability to higher elevations [72], whereas other researchers have demonstrated that the susceptibility of sessile oak (Quercus petraea) to infection with powdery mildew may naturally increase with elevation [73]. Our laboratory has found similar results in American beech that align with the pathogen-host patterns found between powdery mildew and sessile oak wherein higher elevations do not appear to provide expected protection from persistent pests.
Mitton and Ferrenberg have published a 2012 article on the impact of climate change on the lifecycle of the mountain pine beetle. They have shown that flight stage starts earlier (and therefore lasts longer) and provided evidence for two generations per year for certain brood [74] versus the typically expected one generation per year. These findings have been met with resistance by Bentz and Powell that argue against the possibility of a bivoltine life cycle in mountain pine beetle based on the lack of thermal energy available and required for this phenomenon to occur in the Western United States [75]. Although specific changes in pathogen life cycle(s) will need to further examined, it is agreed that ranges are most definitely changing. Areas that had been inhospitable in prior decades (too cold) are now becoming more ideal.
Given the geographical extent of American beech, we expect the effects of global climate change on BBD progression to be diverse in some respects. The scale can overwinter quite efficiently in even bitterly cold conditions. Warmer winters may favor the survival and fecundity of both the scale and fungus. The scale will typically infect beech starting from the base (protected by leaf cover) but regardless of its first point of attack, it can cover the full perimeter of the trunk relatively quickly. A highly susceptible tree will go from little to no infection to dead from BBD in less than ten years. Regardless of the potential subtype or variation of regional Nectria fungi, the genus has proved to be quite deadly to beech. There is no doubt that climate change favors propagation of the pathogen duo responsible for BBD, with particular support for the scale, and although the beech range is expected to expand north, if temperatures continue to rise, this adaptive advantage to the pathogen is lost.
7. Considerations for Beech Forest Management
Multiple studies support the hypothesis that resistance to the scale is a genetically inheritable trait in both European beech [40] and American beech [63,76,77,78]. Field testing has shown that trees that remain symptom free in stands long affected by BBD are resistant to the beech scale insect [25,79,80]. Several research studies have looked for the underlying mechanisms of resistance to the scale insect and have indicated that structural differences, nutrient partitioning, or the overexpression of pathogen fighting enzymes may play a role. A review of the literature to systematically present multiple factors that may influence resistance is outlined in Table 1. Increasing our understanding of possible mechanisms of resistance to BBD and environmental factors that may influence susceptibility of American beech to BBD can inform proactive management strategies in light of climate change.
Additionally, Wargo has argued for the potential role of the concentration of phenolic compounds in inner versus outer bark of infected trees [71]. These types of studies can be confounded by the fact that it is not clear whether a chemical change confers (or does not confer) resistance to BBD or is actually a result of the infestation itself. Information from microarray and proteomic comparative data sets will ideally move efforts for screening for resistance and restoring beech forests in light of stressors of BBD compounded by climate change forward. Two groups have put forth data to evaluate differential mRNA [76] and protein profiles [62] of bark from trees that are more or less susceptible to beech bark disease but the pace of this work has been limited by the lack of genomic and cDNA sequences widely available for American beech to date.
Ideal management now would include the introduction of disease-resistant beech into more northern reaches of the current range to help expedite the migration and increase the likelihood that seed will be dispersed at a rate demanded by elevated temperature and drought conditions. As beech move north, it may take some time for mature and larger trees to become established and the scale infestation may lag behind significantly. Perhaps before this potential pathogen-host match up occurs, our ability to screen American beech trees for resistance, or purposefully plant resistant trees may supersede the disease progression and allow us to prevent and control further infestation.
In anticipation of this, Jennifer Koch and her colleagues at the USDA Forest Service Northern Research Station are currently developing an American beech breeding program focused on the propagation of BBD-resistant beech [83]. Our laboratory has worked with the USFS to use the technique first developed by Houston [79] and then expanded upon by Koch [84] to allow us to successfully screen beech for BBD-resistance at our own sites, as have additional researchers. Through this collaborative approach, we can work together to test, protect and propagate American beech truly resistant to BBD. These trees are currently being developed as potential regional stock for grafting and restoration efforts [78].
Nearly twenty years ago, Lindner put forward a forest gap model analysis to help evaluate the impacts of climate change on forest ecosystems [85]. He pointed out the inevitable interspecific competitions that will occur in light of limited resources. Interestingly, with beech, we have already seen the impacts of excessive beech sprouting (‘beech hell’) on sugar maple populations in the Northern United States [18,48]. Although American beech does not garner the interest of European beech for commercial harvesting, it is important that we learn from what has been done in Europe when considering management of American beech. Biomass production (wood density) has been shown to be influenced by temperature in European beech [86]. Concerning beech forests in the United States, managing for “climatically well-adapted forest composition” and “maximized tree species diversity” as initially described by Lindner certainly make the most sense still today [85]. Studies in European beech have shown a positive response of beech to thinning to reduce competition. It is speculated that thinning can counteract the impacts of increased temperature as beech are particular sensitive to water availability [87].
Based on an extensive comparative study of BBD in North America, Great Britain, France and Germany, “there is relatively little consistent or conclusive evidence to link environmental factors to population trends of C. fagi or to the cause of beech bark disease” [88]. A recent study in Germany focused on European beech management has shown no correlation between management practices and the infestation rate of the beech scale [41]. On the other hand, necrotic disease of beech was shown to increase dramatically (approximately 2% to nearly 18%) after less ideal forestry practices (e.g., cutting interventions mid-summer) were used [89]. Furthermore, research in 2006 found differential survival rates and health outcomes in high vigor American beech trees over a ten-year period dependent on whether the area was clear-cut or a buffer of surrounding trees was left post-harvest. In the study, high vigor American beech was more likely to experience decline or death if left standing amid a clear-cut site. The authors ultimately recommended that a buffer be left around beech trees when a forest is harvested for its timber [90]. Thus, both the season of harvest and the specific harvesting methods may affect the long-term health of beech stands and, as a result, their chances of survival. As we consider more careful monitoring and management of beech in North America, it will be ideal to not inadvertently increase the spread of BBD or adversely impact stands of American beech that may be otherwise healthy.
Hamelin of Vermont Fish and Wildlife has developed a management program to specifically enhance mast in BBD-infested forests in the Northeast [91]. Within five years of the discovery of BBD in Michigan, and with an expectation that it would kill at least half of the mature beech trees [92], McCullough and colleagues developed management strategies for beech forests at various stages (pre-infestation, advancing front, killing front and aftermath) [20]. Cale et al. warn that “conditions within the current aftermath zone are a harbinger of what most beech stands may look like in the near future” in anticipation of additional and substantial Neonectria invasions [93] and in fact, bark phosphorous levels may be more relevant [94]. This is a more recent and disturbing finding when considering overall beech health.
8. Conclusions
The environmental impact of drought and elevated temperatures in North America, coupled with the continuing progression of BBD, is poised to greatly diminish the integrity of beech-dominated forests. The impacts of climate change will not brighten the future for BBD-infected forests from the perspective of the American beech. The expected changes in temperature and precipitation favor the exotic, invasive pathogens, not the beech. As beech have been selected for BBD-resistance in the microevolution of the past century, climate change will yet again provide additional pressures and selection. The genetic heterogeneity of the current beech populations will most surely be tested, but with concrete restoration efforts currently underway, we may have an anthropocentric solution to a human induced problem.
Leak has looked at the impacts of BBD in the Bartlett Experimental Forest in New Hampshire for over fifty years. His findings support the positive impact of selective removal of infested beech on overall beech stand health. However, as we continue to manage our forests in light of the impacts of climate change, Leak himself notes that we should treat these results with caution. “It is quite likely that other regions have different levels of genetic resistance to the beech bark disease, and possibly different strains of scale and Nectria as well. Possibly, trees in the Bartlett stands that we thought to be resistant could be reinfected by new strains of the disease complex” [95].
Acknowledgments
We would like to thank Joshua Duncan for expertise provided to create the map (American beech’s geographic range).
Author Contributions
Both authors wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Coe, N. Life as Beech, Survival in the New England Forest. In Nature and Culture in the Northern Forest; University of Iowa Press: Iowa, IA, USA, 2010; pp. 49–58. [Google Scholar]
- Huntley, B.; Bartlein, P.J.; Prentice, I.C. Climatic control of the distribution and abundance of beech (Fagus L.) in Europe and North America. J. Biogeogr. 1989, 16, 551–560. [Google Scholar] [CrossRef]
- Lassoie, J.; Luzadis, V.; Grover, D. Forest Trees of the Northeast; Cornell Cooperative Extension: Ithaca, NY, USA, 1996. [Google Scholar]
- Tubbs, C.; Houston, D.R. Fagus grandifolia: American Beech. In Silvics of North America; United States Department of Agriculture: Washington, DC, USA, 1990; pp. 335–342. [Google Scholar]
- GECSC: Tree Species Distribution Maps for North America. Available online: https://esp.cr.usgs.gov/data/little (accessed on 1 May 2017).
- Williams-Linera, G.; Devall, M.S.; Alvarez-Aquino, C. A relict population of Fagus grandfolia var. mexicana at the Acatlan Volcano, Mexico: Structure, litterfall, phenology and dendroecology. J. Biogeogr. 2000, 27, 1297–1309. [Google Scholar]
- Busby, P.E.; Motzkin, G.; Hall, B. Distribution and dynamics of American beech in Coastal Southern New England. Northeast. Nat. 2009, 16, 159–176. [Google Scholar] [CrossRef]
- Parker, J. Cold resistance in woody plants. Bot. Rev. 1963, 29, 123–201. [Google Scholar] [CrossRef]
- Sakai, A.; Weiser, C.J. Freezing resistance of trees in North America with reference to tree regions. Ecology 1973, 54, 118–126. [Google Scholar] [CrossRef]
- Fang, J.; Lechowicz, M.J. Climatic limits for the present distribution of beech (Fagus L.) species in the world. J. Biogeogr. 2006, 33, 1804–1819. [Google Scholar] [CrossRef]
- Hershey, R.; Befort, W. Aerial Photo Guide to New England Forest Cover Types; USDA Forest Service: San Francisco, CA, USA, 1993.
- Burns, R. Silvicultural Systems for the Major Forest Types of the United States; United States Department of Agriculture: Washington, DC, USA, 1983.
- Outcalt, K. Magnolia grandiflora: Southern Magnolia. In Silvics of North America; United States Department of Agriculture: Washington, DC, USA, 1990. [Google Scholar]
- Ling, K.A.; Power, S.A.; Ashmore, M.R. A survey of the health of Fagus sylvatica in Southern Britain. J. Appl. Ecol. 2004, 30, 295–306. [Google Scholar] [CrossRef]
- Burns, R.; Honkala, B. Summary of Tree Characteristics. In Silvics of North America; United States Department of Agriculture: Washington, DC, USA, 1990. [Google Scholar]
- Held, M.E. Pattern of beech regeneration in the East-Central United States. Bull. Torrey Bot. Club 1983, 110, 55–62. [Google Scholar] [CrossRef]
- Jones, R.H.; Raynal, D.J. Root sprouting in American beech: Production, survival, and the effect of parent tree vigor. Can. J. For. Res. 1987, 17, 539–544. [Google Scholar] [CrossRef]
- Hane, E.N. Indirect effects of beech bark disease on sugar maple seedling survival. Can. J. For. Res. 2003, 33, 807–813. [Google Scholar] [CrossRef]
- Houston, D.R. Beech Bark Disease: 1934 to 2004: What’s New Since Ehrlich? In Proceedings of the Beech Bark Disease Symposium, New York, NY, USA, 16–18 June 2004. [Google Scholar]
- McCullough, D.G.; Heyd, R.L.; O’Brien, J.G. Biology and Management of Beech Bark Disease; Michigan State University: East Lansing, MI, USA, 2005. [Google Scholar]
- Bucher, E.H. The Causes of Extinction of the Passenger Pigeon. In Current Ornithology; Power, D.M., Ed.; Springer: New York, NY, USA, 1992; pp. 1–36. [Google Scholar]
- Forbush, E. Birds of Massachusetts and Other New England States; Massachusetts Department of Agriculture: Boston, MA, USA, 1927.
- Garneau, D.E.; Lawler, M.E.; Rumpf, A.S.; Weyburne, E.S.; Cuppernull, T.M.; Boe, A.G. Potential effects of beech bark disease on small mammals and invertebrates in Northeastern US forests. Northeast. Nat. 2012, 19, 391–410. [Google Scholar] [CrossRef]
- Houston, D.R. Major new tree disease epidemics: Beech bark disease. Annu. Rev. Phytopathol. 1994, 32, 75–87. [Google Scholar] [CrossRef]
- Houston, D.R. American Beech Resistance to Cryptococcus fagisuga; USDA Forest Service: Washington, DC, USA, 1983.
- Kasson, M.T.; Livingston, W.H. Relationships among beech bark disease, climate, radial growth response and mortality of American beech in northern Maine, USA. For. Pathol. 2012, 42, 199–212. [Google Scholar] [CrossRef]
- Houston, D.R.; O’Brien, J.T. Beech Bark Disease; U.S. Department of Agriculture Forest Service: Washington, DC, USA, 2003.
- Ehrlich, J. The beech bark disease. A Nectria disease of Fagus, following Cryptococcus fagi (BAER.). Can. J. For. Res. 1934, 10, 593–692. [Google Scholar] [CrossRef]
- Houston, D.R. Temporal and spatial shift within the Nectria pathogen complex associated with beech bark disease of Fagus grandifolia. Can. J. For. Res. 1993, 24, 960–968. [Google Scholar] [CrossRef]
- Morris, A.B.; Small, R.L.; Cruzan, M.B. Investing the relationship between Cryptococcus fagisuga and Fagus Grandifolia in Great Smoky Mountains National Park. Southeast. Nat. 2002, 1, 415–424. [Google Scholar] [CrossRef]
- Wainhouse, D. Dispersal of first instar larvae of the felted beech scale, Cryptococcus fagisuga. J. Appl. Ecol. 1980, 17, 523–532. [Google Scholar] [CrossRef]
- Shigo, A.L. The beech bark disease today in the Northeastern U.S. J. For. 1972, 70, 286–289. [Google Scholar]
- Shigo, A.L. Organism interactions in the beech bark disease. Phytopathology 1964, 54, 263–269. [Google Scholar]
- Lohman, M.L.; Watson, A.J. Identity and host relations of Nectria species associated with diseases of hardwoods in the eastern states. Lloydia 1943, 6, 77–108. [Google Scholar]
- Castlebury, L.A.; Rossman, A.Y.; Hyten, A.S. Phylogenetic relationships of Neonectria/Cylindrocarpon on Fagus in North America. Can. J. Bot. 2006, 84, 1417–1433. [Google Scholar] [CrossRef]
- Houston, D.R.; Mahoney, E.M. Beech bark disease: Association of Nectria ochroleuca in W.Va., Pa., and Ontario. Phytopathol. Abstr. 1987, 77, 1615. [Google Scholar]
- Jung, T. Beech decline in Central Europe driven by the interaction between Phytophthora infections and climatic extremes. For. Pathol. 2009, 39, 73–94. [Google Scholar] [CrossRef]
- Morin, R.S.; Liebhold, A.M.; Tobin, P.C.; Gottschalk, K.W.; Luzader, E. Spread of beech bark disease in the eastern United States and its relationship to regional forest composition. Can. J. For. Res. 2007, 37, 726–736. [Google Scholar] [CrossRef]
- Beech Bark Disease. Available online: http://forestinvasives.ca/Meet-the-Species/Pathogens/Beech-Bark-Disease#70229-impacts (accessed on 17 April 2017).
- Krabel, D.; Petercord, R. Genetic diversity and bark physiology of the European beech (Fagus sylvatica): A coevolutionary relationship with the beech scale (Cryptococcus fagisuga). Tree Physiol. 2000, 20, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Köhler, G.; Pašalić, E.; Weisser, W.W.; Gossner, M.M. Beech forest management does not affect the infestation rate of the beech scale Cryptococcus fagisuga across three regions in Germany. Agric. For. Entomol. 2015, 17, 197–204. [Google Scholar] [CrossRef]
- Houston, D.R. Beech bark disease: The result of an invasive causal complex. In Ecology of Invasive Species; Yale University: New Haven, CT, USA, 1999; pp. 16–17. [Google Scholar]
- McLachlan, J.S.; Clark, J.S.; Manos, P.S. Molecular indicators of tree migration capacity under rapid climate change. Ecology 2005, 86, 2088–2098. [Google Scholar] [CrossRef]
- Pearson, R.G. Climate change and the migration capacity of species. Trends Ecol. Evol. 2006, 21, 111–113. [Google Scholar] [CrossRef] [PubMed]
- Gavin, D.G.; Peart, D.R. Effects of beech bark disease on the growth of American Beech (Fagus grandifollia). Can. J. For. Res. 1993, 23, 1566–1575. [Google Scholar] [CrossRef]
- Morin, R.S.; Liebhold, A.M. Invasions by two non-native insects alter regional forest species composition and successional trajectories. For. Ecol. Manag. 2015, 341, 67–74. [Google Scholar] [CrossRef]
- Krasny, M.E.; DiGregorio, L.M. Gap dynamics in Allegheny northern hardwood forests in the presence of beech bark disease and gypsy moth disturbances. For. Ecol. Manag. 2001, 144, 265–274. [Google Scholar] [CrossRef]
- Evans, C.A.; Ayres, M.P.; Twery, M.J.; Houston, D.R. Using Models to Identify Forests at Risk of Major Structure and Compositional Change due to Beech Bark Disease. In Proceedings of the Beech Bark Disease Symposium, New York, NY, USA, 16–18 June 2004. [Google Scholar]
- Geßler, A.; Keitel, C.; Kreuzwieser, J.; Matyssek, R.; Seiler, W.; Rennenberg, H. Potential risks for European beech (Fagus sylvatica L.) in a changing climate. Trees 2007, 21, 1–11. [Google Scholar] [CrossRef]
- Schlesinger, W.H.; Andrews, J.A. Soil respiration and the global carbon cycle. Biogeochemistry 2000, 48, 7–20. [Google Scholar] [CrossRef]
- Kirschbaum, M.U.F. Will changes in soil organic carbon act as a positive or negative feedback on global warming? Biogeochemistry 2000, 48, 21–51. [Google Scholar] [CrossRef]
- Davidson, E.A.; Janssens, I.A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 2006, 440, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Fitzhugh, R.D.; Driscoll, C.T.; Groffman, P.M.; Tierney, G.L.; Fahey, T.J.; Hardy, J.P. Effects of soil freezing disturbance on soil solution nitrogen, phosphorus, and carbon chemistry in a northern hardwood ecosystem. Biogeochemistry 2001, 56, 215–238. [Google Scholar] [CrossRef]
- Groffman, P.; Hardy, J.P.; Fashu-Kanu, S.; Driscoll, C.T.; Cleavitt, N.L.; Fahey, T.J.; Fisk, M.C. Snow depth, soil freezing and nitrogen cycling in a Northern Hardwood forest landscape. Biogeochemistry 2011, 102, 223–238. [Google Scholar] [CrossRef]
- Groffman, P.M.; Driscoll, C.T.; Fahey, T.J.; Hardy, J.P.; Fitzhugh, R.D.; Tierney, G.L. Colder soils in a warmer world: A snow manipulation study in a Northern Hardwood forest ecosystem. Biogeochemistry 2001, 56, 135–150. [Google Scholar] [CrossRef]
- Tierney, G.L.; Fahey, T.J.; Groffman, P.M.; Hardy, J.P.; Fitzhugh, R.D.; Driscoll, C.T. Soil freezing alters fine root dynamics in a Northern Hardwood forest. Biogeochemistry 2001, 56, 175–190. [Google Scholar] [CrossRef]
- Rustad, L.E.; Boyer, E.W.; Christopher, S.F.; Driscoll, C.T.; Fernandez, I.J.; Groffman, P.M.; Houle, D.; Kiekbusch, J.; Magill, A.H.; Mitchell, M.J.; et al. Consequences of climate change for biogeochemical cycling in forests of northeastern North America. Can. J. For. Res. 2009, 39, 264–284. [Google Scholar]
- Prasad, A.M.; Iverson, L.R.; Matthews, S.; Peters, M. A Climate Change Atlas for 134 Forest Tree Species of the Eastern United States; Northern Research Station, USDA Forest Service: Delaware, OH, USA, 2007.
- Polle, A.; Morawe, B. Seasonal changes of the antioxidative systems in foliar buds and leaves of field-grown beech trees (Fagus sylvatica, L.) in a stressful climate. Bot. Acta 1995, 108, 314–320. [Google Scholar] [CrossRef]
- Kramer, K.; Degen, B.; Buschbom, J.; Hickler, T.; Thuiller, W.; Sykes, M.; de Winter, W. Modelling exploration of the future of European beech (Fagus sylvatica L.) under climate change—Range, abundance, genetic diversity and adaptive response. For. Ecol. Manag. 2010, 259, 2213–2222. [Google Scholar] [CrossRef]
- Houston, D.B.; Houston, D.R. Allozyme genetic diversity among Fagus grandifolia trees resistant or susceptible to beech bark disease in natural populations. Can. J. For. Res. 2000, 30, 778–789. [Google Scholar] [CrossRef]
- Mason, M.E.; Koch, J.L.K.; Loo, J. Comparisons of protein profiles of beech bark disease resistant and susceptible American Beech (Fagus grandifolia). Proteome Sci. 2013, 11, 2. [Google Scholar] [CrossRef] [PubMed]
- Koch, J.L.; Carey, D.W.; Mason, M.E.; Nelson, C.D. Assessment of beech scale resistance in full- and half-sibling American beech families. Can. J. For. Res. 2010, 40, 265–272. [Google Scholar] [CrossRef]
- Kubisiak, T.; Carey, D.W.; Koch, J.L. Charcterization of ten EST-based Polymorphic SSR Loci Isolated from American Beech, Fagus grandifolia Ehrh; USDA Forest Service, Southern Research Station, Southern Institute of Forest Genetics: Washington, DC, USA, 2010.
- Latty, E.F.; Canham, C.D.; Marks, P.L. Beech bark disease in northern hardwood forests: The importance of nitrogen dynamics and forest history for disease severity. Can. J. For. Res. 2003, 33, 257–268. [Google Scholar] [CrossRef]
- Wargo, P.M. Amino nitrogen and phenolic constituents of bark of American Beech, Fagus grandifolia, and infestation by beech scale, Cryptococcus fagisuga. For. Pathol. 1988, 18, 279–290. [Google Scholar] [CrossRef]
- Dukes, J.S.; Pontius, J.; Orwig, D.; Garnas, J.R.; Rodgers, V.; Brazee, N.; Cook, B.; Theoharides, K.; Stange, E.; Harrington, R.; et al. Response of insect pests, pathogens, and invasive plant species to climate change in the forests of northeastern North America: What can we predict? Can. J. For. Res. 2009, 39, 231–248. [Google Scholar] [CrossRef]
- Gove, J.H.; Houston, D.R. Monitoring the growth of American beech affected by beech bark disease in Maine using the Kalman filter. Environ. Ecol. Stat. 1996, 3, 167–187. [Google Scholar] [CrossRef]
- Taylor, A.R. Putting healthy beech on the map. Atl. For. Rev. 2013, 20, 38–41. [Google Scholar]
- Forrester, J.A.; Runkle, J.R. Mortality and Replacement patterns of an old-growth Acer fagus woods in the Holden Arboretum, Northeastern Ohio. Am. Midl. Nat. 2000, 144, 227–242. [Google Scholar] [CrossRef]
- Paradis, A.; Elkinton, J.; Hayhoe, K.; Buonaccorsi, J. Role of winter temperature and climate change on the survival and future range expansion of the hemlock woolly adelgid (Adelges tsugae) in eastern North America. Mitig. Adapt. Strateg. Glob. Chang. 2007, 13, 541–554. [Google Scholar] [CrossRef]
- Logan, J.A.; Macfarlane, W.W.; Willcox, L. Whitebark pine vulnerability to climate-driven mountain pine beetle disturbance in the Greater Yellowstone ecosystem. Ecol. Appl. 2010, 20, 895–902. [Google Scholar] [CrossRef] [PubMed]
- Dantec, C.F.; Ducasse, H.; Capdevielle, X.; Fabreguettes, O.; Delzon, S.; Deprez-Loustau, M.-L. Escape of spring frost and disease through phenological variations in oak populations along elevation gradients. J. Ecol. 2015, 103, 1044–1056. [Google Scholar] [CrossRef]
- Mitton, J.B.; Ferrenberg, S.M. Mountain pine beetle develops an unprecedented summer generation in response to climate warming. Am. Nat. 2012, 179, E163–E171. [Google Scholar] [CrossRef] [PubMed]
- Bentz, B.J.; Powell, J.A. Mountain pine beetle seasonal timing and constraints to bivoltinism. Am. Nat. 2014, 184, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Seitler, K.; Coe, N. cDNA primers for quantitative analysis of protective compounds implicated in beech bark disease resistance in American beech, Fagus grandifolia. Conserv. Genet. Resour. 2015, 7, 689–691. [Google Scholar] [CrossRef]
- Koch, J.L.; Carey, D.W. The Genetics of Resistance of American Beech to Beech Bark Disease: Knowledge through 2004. In Proceedings of the Beech Bark Disease Symposium, New York, NY, USA, 16–18 June 2004. [Google Scholar]
- Koch, J.L.; Mason, M.E.; Carey, D.W. Screening for Resistance to Beech Bark Disease: Improvements and Results from Seedlings and Grafted Filed Selections; General Technical PSW-GTR-240; U.S. Department of Agriculture: Washington, DC, USA, 2012.
- Houston, D.R. A Technique to Artificially Infest Beech Bark Disease with the Beech Scale, Cryptococcus fagisuga (Lindinger); USDA Forest Service Research Paper NE-507; USDA Forest Service: Washington, DC, USA, 1982.
- Londsdale, D. Wood and Bark Anatomy of Young Beech in Relation to Cryptococcus Attack. In Proceedings of the IUFRO Beech Bark Disease Working Party Conference, Hamden, CT, USA, 26 September–8 October 1982. [Google Scholar]
- Griffin, J.M.; Lovett, G.M.; Arthur, M.A.; Weathers, K.C. The distribution and severity of beech bark disease in the Catskill Mountains, N.Y. Can. J. For. Res. 2003, 33, 1754–1760. [Google Scholar] [CrossRef]
- Twery, M.J. Effects of Species Composition and Site Factors on the Severity of Beech Bark Disease in Western Massachusetts and the White Mountains of New Hampshire: A Preliminary Report. In Proceedings of the I.U.F.R.O. Beech Bark Disease Working Party Conference, Hamden, CT, USA, 26 September–8 October 1982. [Google Scholar]
- Development of an American Beech Breeding Program—Managing with Disturbance—Northern Research Station—USDA Forest Service. Available online: https://www.nrs.fs.fed.us/disturbance/invasive_species/beech_breeding (accessed on 17 April 2017).
- Koch, J.L.; Carey, D.W. A technique to screen American Beech for resistance to the beech scale insect (Cryptococcus fagisuga Lind.). J. Vis. Exp. 2014. [Google Scholar] [CrossRef] [PubMed]
- Lindner, M. Developing adaptive forest management strategies to cope with climate change. Tree Physiol. 2000, 20, 299–307. [Google Scholar] [CrossRef]
- Bouriaud, O.; Bréda, N.; Moguédec, G.L.; Nepveu, G. Modelling variability of wood density in beech as affected by ring age, radial growth and climate. Trees 2004, 18, 264–276. [Google Scholar] [CrossRef]
- Cescatti, A.; Piutti, E. Silvicultural alternatives, competition regime and sensitivity to climate in a European beech forest. For. Ecol. Manag. 1998, 102, 213–223. [Google Scholar] [CrossRef]
- Houston, D.R.; Parker, E.J.; Perrin, R.; Lang, K.J. Beech bark disease: A comparison of the disease in North America, Great Britain, France, and Germany. Eur. J. For. Pathol. 1979, 9, 199–211. [Google Scholar] [CrossRef]
- Cicák, A.; Mihál, L. T-disease—A little investigated phenomenon of the beech (Fagus sylvatica L.) necrotic damage. J. For. Sci. 2001, 47, 38–41. [Google Scholar]
- Farrar, A.; Ostrofsky, W.D. Dynamics of American Beech regeneration 10 years following harvesting in a beech bark disease—Affected stand in Maine. J. Appl. For. 2006, 23, 192–196. [Google Scholar]
- Hamelin, P.L. VT ANR Management Guidelines for Optimizing Mast Yields in Beech Mast Production Areas; Vermont Fish & Wildlife Department: Waterbury, VT, USA, 2011.
- Kearney, A.; McCullough, D.; Walters, M. Impact of beech bark disease on understory composition in Michigan. In Beech Bark Disease; Michigan State University: East Lansing, MI, USA, 2005; pp. 58–59. [Google Scholar]
- Cale, J.A.; Letkowski, S.K.; Teale, S.A.; Castello, J.D. Beech bark disease: An evaluation of the predisposition hypothesis in an aftermath forest. For. Pathol. 2012, 42, 52–56. [Google Scholar] [CrossRef]
- Cale, J.A.; Teale, S.A.; Johnston, M.T.; Boyer, G.L.; Perri, K.A.; Castello, J.D. New ecological and physiological dimensions of beech bark disease development in aftermath forests. For. Ecol. Manag. 2015, 336, 99–108. [Google Scholar] [CrossRef]
- Leak, W.B. Fifty-year impacts of the beech bark disease in the Bartlett experimental forest, New Hampshire. North. J. Appl. For. 2006, 23, 141–143. [Google Scholar]
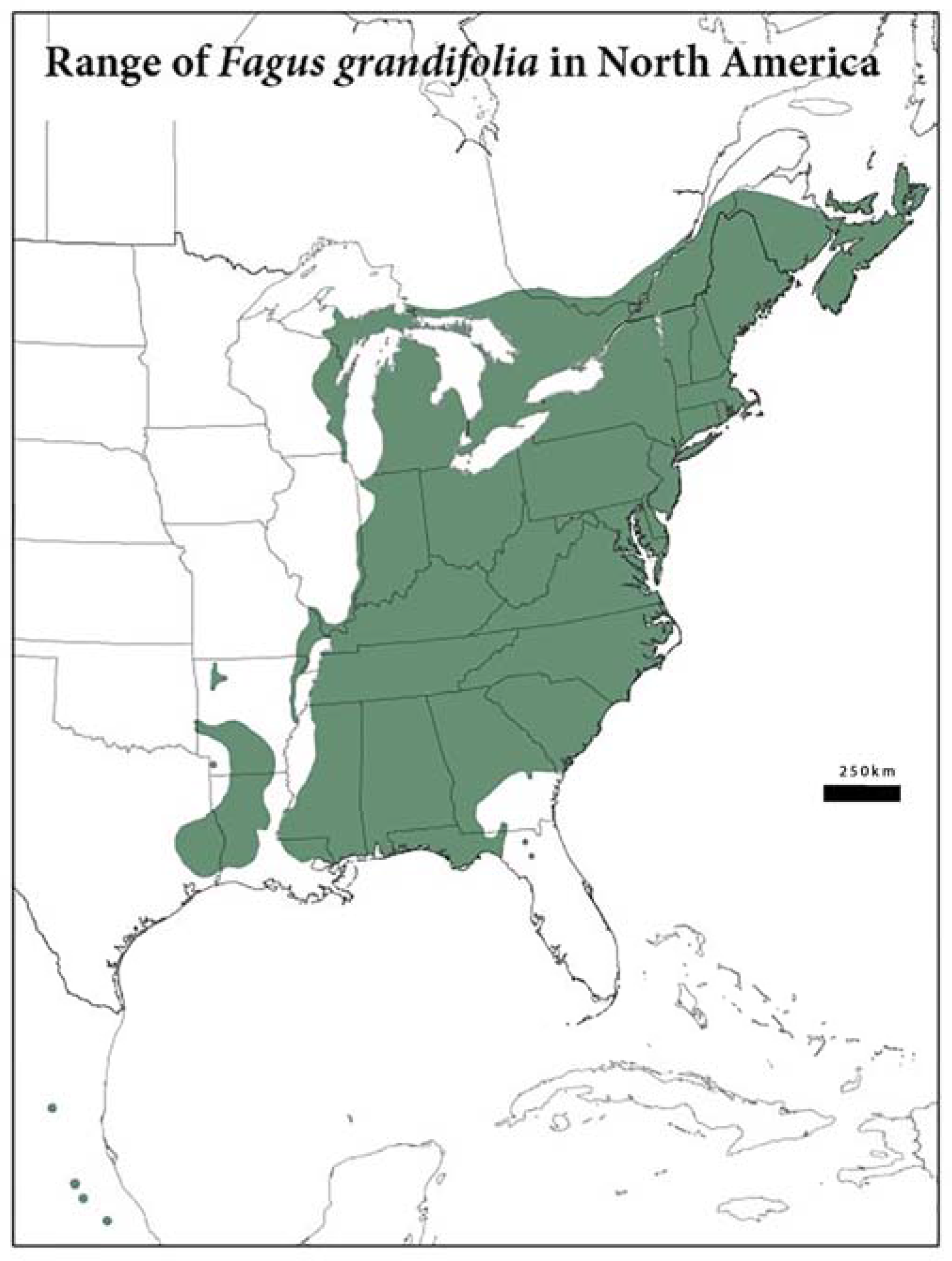
Figure 1.
Present geographic distribution of the American Beech. This map was created using ArcGIS and the documented range of American Beech is depicted in green. The data were downloaded from the United States Geological Survey’s Geosciences and Environmental Changed center website [5].
Figure 1.
Present geographic distribution of the American Beech. This map was created using ArcGIS and the documented range of American Beech is depicted in green. The data were downloaded from the United States Geological Survey’s Geosciences and Environmental Changed center website [5].

Figure 2.
Visible signs of pathogens responsible for Beech Bark Disease. (A) Intense patch of wax created by overwintering Beech scale (Cryptococcus fagisuga Lind.); (B) Fruiting bodies of the Neonectria fungal infection (typically N. ditissima and/or N. faginata); (C) Typical “whitewashing” phenotype of scale infested Beesh; (D) Neonectria fungal infection. Multiple raised circular red welts are commonly seen. Photos by Natalie Coe.
Figure 2.
Visible signs of pathogens responsible for Beech Bark Disease. (A) Intense patch of wax created by overwintering Beech scale (Cryptococcus fagisuga Lind.); (B) Fruiting bodies of the Neonectria fungal infection (typically N. ditissima and/or N. faginata); (C) Typical “whitewashing” phenotype of scale infested Beesh; (D) Neonectria fungal infection. Multiple raised circular red welts are commonly seen. Photos by Natalie Coe.


{kind=link}
{kind=link}
Table 1.
An overview of the possible mechanisms of resistance and environment factors that may influence susceptibility of American beech to Beech Bark Disease (BBD) during the “killing front”.
Table 1.
An overview of the possible mechanisms of resistance and environment factors that may influence susceptibility of American beech to Beech Bark Disease (BBD) during the “killing front”.
| Possible Mechanisms of Resistance | Environmental Factors |
|---|---|
| Structural (endogenous to host) Stone cell layers of cortex may provide structural barrier(s) to scale stylet [80] | Structural (exogenous to host) Forest type and forest dynamics [81] Forest structure [38] Species composition and site factors [82] |
| Nutritional access (pathogen perspective) Nutrition partitioning allows tree to discourage infestation [40] Higher bark nitrogen may provide more desirable habitat for the beech scale [65,66] | Nutritional availability (host perspective) Nutrients and water [26] Drought conditions [14] |
| Differential gene expression Differential gene expression in disease resistant versus BBD-susceptible beech trees [62,76] | Location attributes Stand-level [28] |
In considering the influence of climate change in synergy with disease pathology, it is necessary to consider all the environmental factors that may be influenced or linked to climate change within the context of both physical parameters (slope, aspect, etc.) and genetic conditions that may predispose trees to susceptibility or resistance.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Stephanson, C.A.; Ribarik Coe, N. Impacts of Beech Bark Disease and Climate Change on American Beech. Forests 2017, 8, 155. https://doi.org/10.3390/f8050155
AMA Style
Stephanson CA, Ribarik Coe N. Impacts of Beech Bark Disease and Climate Change on American Beech. Forests. 2017; 8(5):155. https://doi.org/10.3390/f8050155
Chicago/Turabian StyleStephanson, Christopher Alexander, and Natalie Ribarik Coe. 2017. "Impacts of Beech Bark Disease and Climate Change on American Beech" Forests 8, no. 5: 155. https://doi.org/10.3390/f8050155
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.